Intensification of Organophosphorus Hydrolase Synthesis by Using Substances with Gas-Transport Function

Abstract
:1. Introduction
2. Materials and Methods
2.1. Biological and Chemical Materials
2.2. Synthesis of Perfluoroethers
2.3. General Experimental Procedures
2.4. Analysis Methods
3. Results
3.1. Cultivation of Recombinant Protein-Producing Cells in Media Containing PFCs
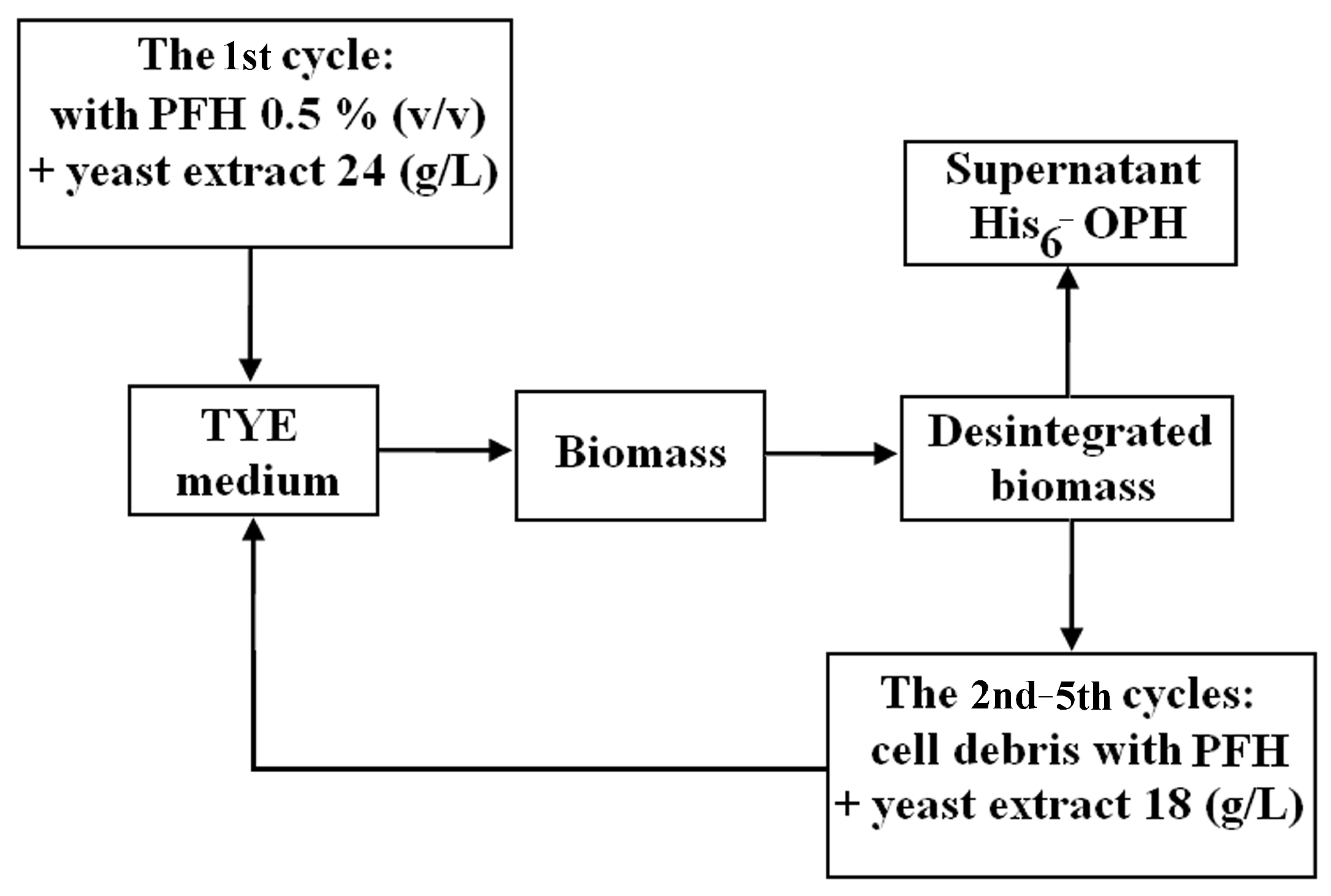
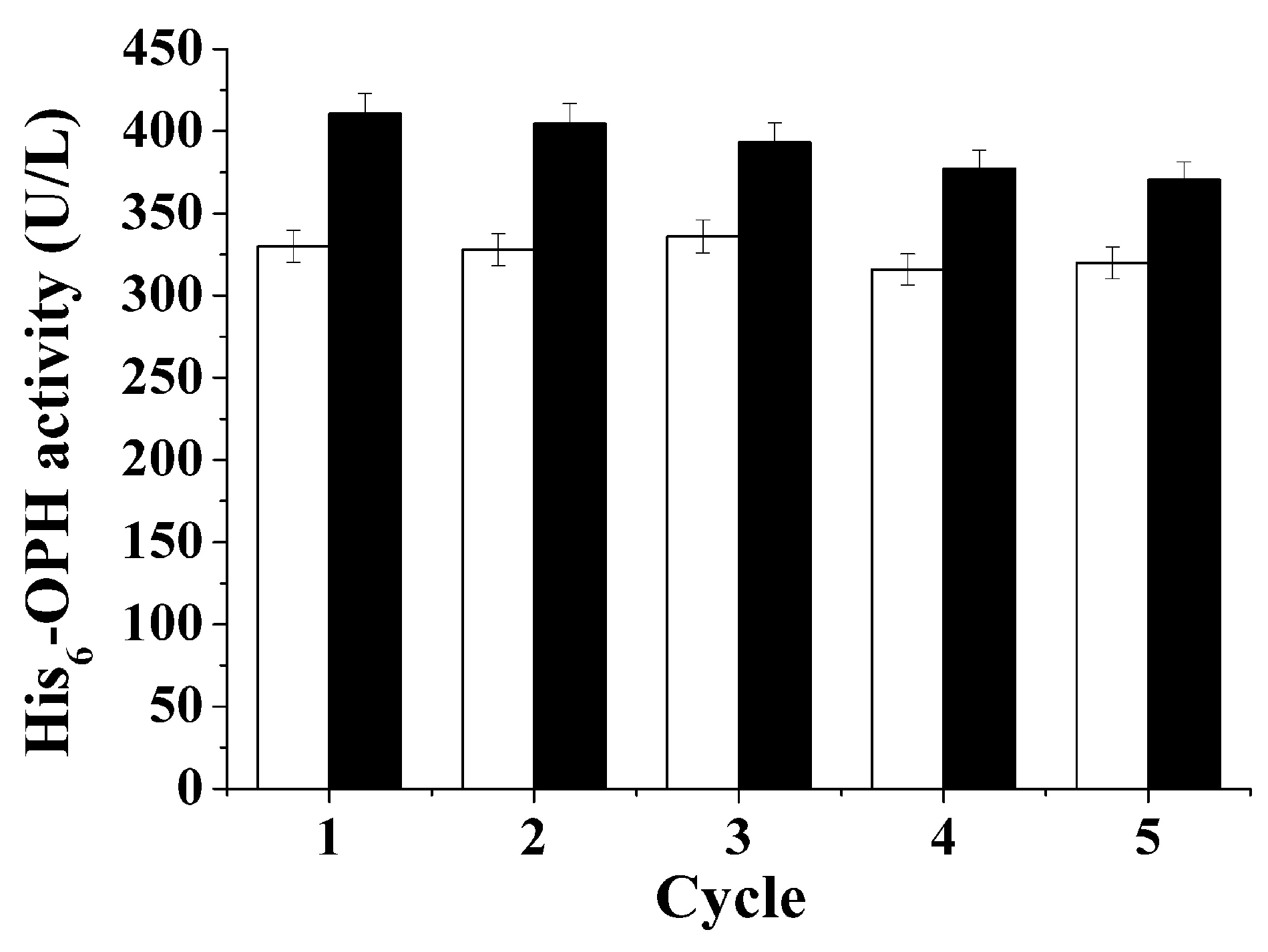
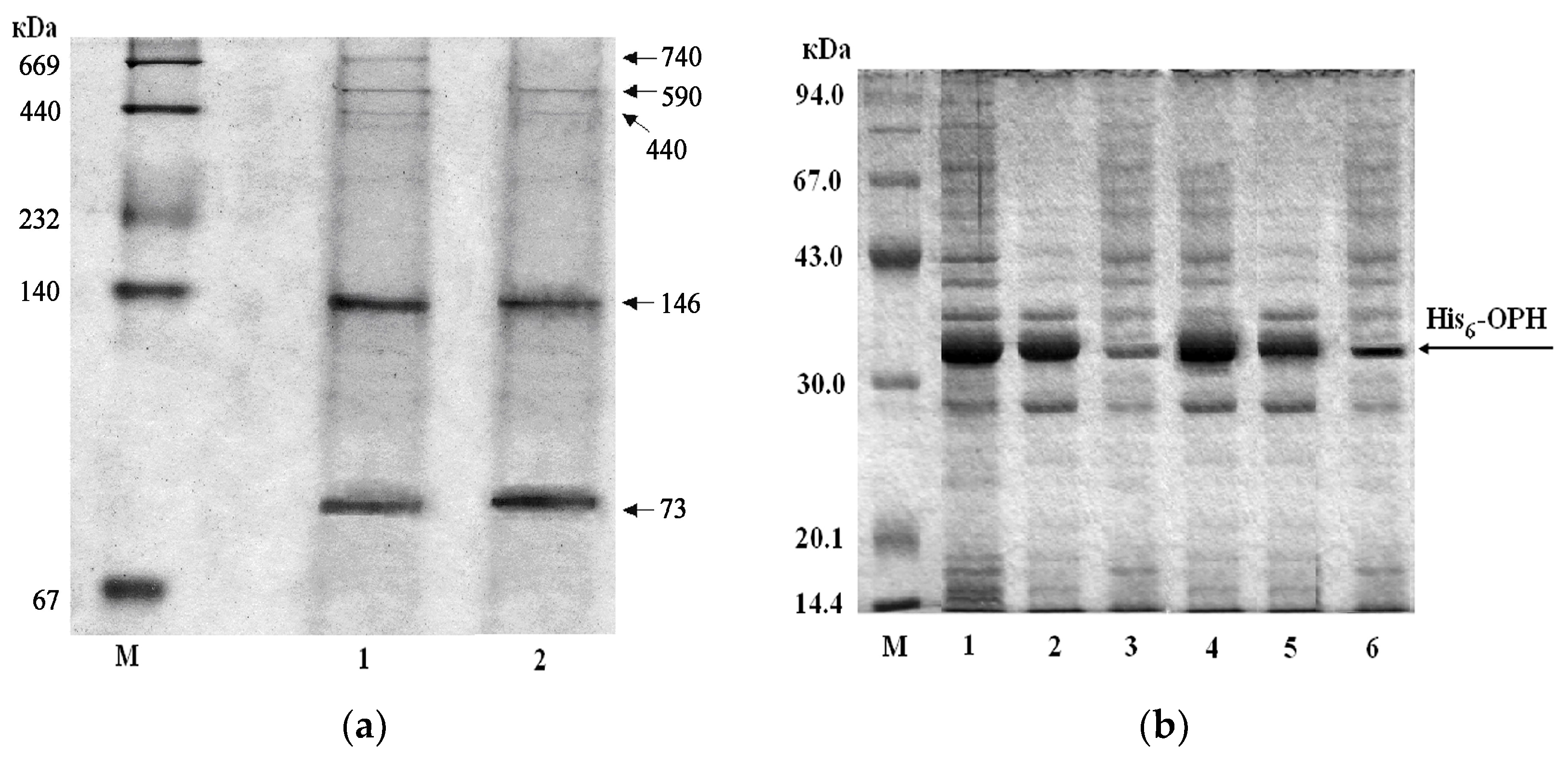
3.2. Studying the Possibility of muLtiple re-Use of Gas Transport-Capable Substances in Biosynthesis of Intracellular Proteins
4. Discussion
5. Conclusions
6. Patents
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Rosano, G.L.; Ceccarelli, E.A. Recombinant protein expression in Escherichia coli: Advances and challenges. Front. Microbiol. 2014, 5, 172. [Google Scholar] [CrossRef] [PubMed]
- Jia, B.; Jeon, C.O. High-throughput recombinant protein expression in Escherichia coli: Current status and future perspectives. Open Biol. 2016, 6, 160196. [Google Scholar] [CrossRef] [PubMed]
- Fakruddin, M.; Mohammad Mazumdar, R.; Bin Mannan, K.S.; Chowdhury, A.; Hossain, M.N. Critical factors affecting the success of cloning, expression, and mass production of enzymes by recombinant E. coli. ISRN Biotechnol. 2013, 2013, 590587. [Google Scholar] [CrossRef] [PubMed]
- Spadiut, O.; Capone, S.; Krainer, F.; Glieder, A.; Herwig, C. Microbials for the production of monoclonal antibodies and antibody fragments. Trends Biotechnol. 2014, 32, 54–60. [Google Scholar] [CrossRef] [PubMed]
- Tripathi, N.K.; Shrivastva, A.; Biswal, K.C.; Lakshmana Rao, P.V. Optimization of culture medium for production of recombinant dengue protein in Escherichia coli. Ind. Biotechnol. 2009, 5, 179–183. [Google Scholar] [CrossRef]
- Kamionka, M. Engineering of therapeutic proteins production in Escherichia coli. Curr. Pharm. Biotechnol. 2011, 12, 268–274. [Google Scholar] [CrossRef] [PubMed]
- Baeshen, M.N.; Al-Hejin, A.M.; Bora, R.S.; Ahmed, M.M.; Ramadan, H.A.; Saini, K.S.; Baeshen, N.A.; Redwan, E.M. Production of biopharmaceuticals in E. coli: Current scenario and future perspectives. J. Microbiol. Biotechnol. 2015, 25, 953–962. [Google Scholar] [CrossRef] [PubMed]
- Garcia-Ochoa, F.; Gomez, E. Bioreactor scale-up and oxygen transfer rate in microbial processes: An overview. Biotechnol. Adv. 2009, 27, 153–176. [Google Scholar] [CrossRef] [PubMed]
- Ibrahim, D.; Weloosamy, H.; Lim, S.-H. Effect of agitation speed on the morphology of Aspergillus niger HFD5A-1 hyphae and its pectinase production in submerged fermentation. World J. Biol. Chem. 2015, 6, 265–271. [Google Scholar] [CrossRef] [PubMed]
- Lowe, K.C. Perfluorochemical respiratory gas carriers: Benefits to cell culture systems. J. Fluorine Chem. 2002, 118, 19–26. [Google Scholar] [CrossRef]
- Pilarek, M.; Szewczyk, K.W. Effects of perfluorinated oxygen carrier application in yeast, fungi and plant cell suspension cultures. Biochem. Eng. J. 2008, 41, 38–42. [Google Scholar] [CrossRef]
- Vieira, E.S.; de Oliveira Fontes, K.T.; Pereira, M.M.; Alexandre, H.V.; da Silva, D.P.; Soares, C.M.F.; Lima, A.S. New strategy to apply perfluorodecalin as an oxygen carrier in lipase production: Minimisation and reuse. Bioprocess. Biosyst. Eng. 2015, 38, 721–728. [Google Scholar] [CrossRef] [PubMed]
- Martinson, E.A.; Birjukov, V.V.; Bakulin, V.M.; Kruchkov, A.V.; Sincov, K.N. Influence of perfluorodecalin on growth of Escherichia coli of М-17 at deep cultivation. Fundam. Res. 2014, 5, 71–74. [Google Scholar]
- Pilarek, M. Liquid perfluorochemicals as flexible and efficient gas carrier applied in bioprocess engineering an updated overview and future prospect. Chem. Process. Eng. 2014, 35, 463–487. [Google Scholar] [CrossRef]
- Pilarek, M.; Brand, E.; Hillig, F.; Krause, M.; Neubauer, P. Enhanced plasmid production in miniaturized high-cell-density cultures of Escherichia coli supported with perfluorinated oxygen carrier. Bioprocess. Biosyst. Eng. 2013, 36, 1079–1086. [Google Scholar] [CrossRef] [PubMed]
- Lowe, K.C.; Davey, M.R.; Power, J.B. Perfluorochemicals: Their applications and benefits to cell culture. Trends Biotechnol. 1998, 16, 272–277. [Google Scholar] [CrossRef]
- Pilarek, M.; Glazyrina, J.; Neubauer, P. Enhanced growth and recombinant protein production of Escherichia coli by a perfluorinated oxygen carrier in miniaturized fed-batch cultures. Microb. Cell Fact. 2011, 10, 50. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Grunzel, P.; Pilarek, M.; Steinbrück, D.; Neubauer, A.; Brand, E.; Kumke, M.U.; Neubauer, P.; Krause, M. Mini-scale cultivation method enables expeditious plasmid production in Escherichia coli. Biotechnol. J. 2014, 9, 128–136. [Google Scholar] [CrossRef] [PubMed]
- Bakulin, M.K.; Grudtsyna, A.S.; Pletneva, A.Y.; Kucherenko, A.S.; Lyapustin, A.V.; Malakhov, I.G. Effect of perfluorodecalin, carbogal, and perfluoromethyldecalin on growth and ice-forming activity of bacteria. Microbiology 2006, 75, 312–316. [Google Scholar] [CrossRef]
- Su, L.; Shen, Y.; Gao, T.; Luo, J.; Wang, M. Improvement of AD biosynthesis response to enhanced oxygen transfer by oxygen vectors in Mycobacterium neoaurum TCCC 11979. Appl. Biochem. Biotechnol. 2017, 182, 1564–1574. [Google Scholar] [CrossRef] [PubMed]
- Lyagin, I.V.; Andrianova, M.S.; Efremenko, E.N. Extensive hydrolysis of phosphonates as unexpected behaviour of the known His6-organophosphorus hydrolase. Appl. Microbiol. Biotechnol. 2016, 100, 5829–5838. [Google Scholar] [CrossRef] [PubMed]
- Efremenko, E.N.; Lyagin, I.V.; Klyachko, N.L.; Bronich, T.; Zavyalova, N.V.; Jiang, Y.; Kabanov, A.V. A simple and highly effective catalytic nanozyme scavenger for organophosphorus neurotoxins. J. Control Release 2017, 247, 175–181. [Google Scholar] [CrossRef] [PubMed]
- Sirotkina, M.; Efremenko, E.N. Rhodococcus lactonase with organophosphate hydrolase (oph) activity and his-tagged oph with lactonase activity: Evolutionary proximity of the enzymes and new possibilities in their application. Appl. Microbiol. Biotechnol. 2014, 98, 2647–2656. [Google Scholar] [CrossRef] [PubMed]
- Maslova, O.V.; Senko, O.V.; Stepanov, N.A.; Aslanli, A.G.; Efremenko, E.N. His6-OPH and its stabilized forms combating quorum sensing molecules of gram-negative bacteria in combination with antibiotics. Jundishapur J. Nat. Pharm. Prod. 2017, in press. [Google Scholar] [CrossRef]
- Efremenko, E.N.; Votchitseva, Y.A.; Aliev, T.K.; Varfolomeev, S.D. Recombinant Plasmid DNA pTES-HIS-OPH and Producer of Oligohistidine-Containing Organophosphate Hydrolase. RU Patent 2255975 C 1, 10 July 2005. [Google Scholar]
- Lyagin, I.V.; Efremenko, E.N. Biomolecular engineering of biocatalysts hydrolyzing neurotoxic organophosphates. Biochimie 2018, 144, 115–121. [Google Scholar] [CrossRef] [PubMed]
- Freire, M.G.; Dias, A.M.A.; Coelho, M.A.Z.; Coutinho, J.A.P.; Marrucho, I.M. Aging mechanisms of perfluorocarbon emulsions using image analysis. J. Colloid Interface Sci. 2005, 286, 224–232. [Google Scholar] [CrossRef] [PubMed]
- Efremenko, E.; Votchitseva, Y.; Plieva, F.; Galaev, I.; Mattiasson, B. Purification of His6-organophosphate hydrolase using monolithic supermacroporous polyacrylamide cryogels developed for immobilized metal affinity chromatography. Appl. Microbiol. Biotechnol. 2006, 70, 558–563. [Google Scholar] [CrossRef] [PubMed]
- Berney, M.; Weilenmann, H.-U.; Julian Ihssen, J.; Bassin, C.; Egli, T. Specific growth rate determines the sensitivity of Escherichia coli to thermal, UVA, and solar disinfection. Appl. Environ. Microbiol. 2006, 72, 2586–2593. [Google Scholar] [CrossRef] [PubMed]
- Ntwampe, S.K.O.; Williams, C.C.; Sheldon, M.S. Influence of perfluorocarbons on Phanerochaete. chrysosporium biomass development, substrate consumption and enzyme production. Chem. Biochem. Eng. Q. 2010, 24, 187–194. [Google Scholar]
- Elibola, M.; Mavitunab, F. A remedy to oxygen limitation problem in antibiotic production: Addition of perfluorocarbon. Biochem. Eng. J. 1999, 3, 1–7. [Google Scholar] [CrossRef]
- Efremenko, E.; Lyagin, I.; Votchitseva, Y.; Sirotkina, M.; Varfolomeyev, S. Polyhistidine-containing organophosphorus hydrolase with outstanding properties. Biocatal. Biotransform. 2007, 25, 103–108. [Google Scholar] [CrossRef]
- Efremenko, E.N.; Votchitseva, Y.A.; Aliev, T.K.; Varfolomeyev, S.D. Expression of recombinant organophosphorus hydrolase in active form. In Biocatalytic Technology and Nanotechnology; Zaikov, G.E., Ed.; Nova Science Publishers, Inc.: New York, NY, USA, 2004; pp. 65–71. ISBN 1-59454-117-5. [Google Scholar]
- Efremenko, E.N.; Lyagin, I.V.; Votchitseva, Y.V.; Gudkov, D.A.; Peregudov, A.A.; Aliev, T.K.; Varfolomeev, S.D. The influence of length and localization of polyhistidine tag in the molecule of organophosphorus hydrolase on the biosynthesis and behavior of fusion protein. In Biotechnology: State of the Art and Prospects for Development; Zaikov, G.E., Ed.; Nova Science Publishers, Inc.: New York, NY, USA, 2008; pp. 87–101. ISBN 978-1-60456-015-2. [Google Scholar]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| PFCs | Molecular Mass (g/mol) | О2 Solubility at 298 K (% v/v) | Density (kg/m3) |
|---|---|---|---|
| 1,1,1,2,2,3,3,4,4,5,5,6,6,6-tetradecafluorohexane Perfluorohexane CF3(CF2)4CF3 | 337.90 * | 55.5 | 1.678 * |
| 3,6-dioxaperfluoro-5-methylnonane (Polyether I) C3F7OCF(CF3)CF2OC2F5 | 470.05 | 52.0 | 1.790 |
| 1,1,2,2,3,3,4,4a,5,5,6,6,7,7,8,8,8a-octadecafluorodecalinPerfluorodecalin C10F18 | 461.98 * | 45.0 | 1.930 * |
| 4,7,10,13,16,19,22,25-octaoxaperfluoro- 5,8,11,14,17,18,21,24-octamethyloctacosane (Polyether II) [С3F3O(C2F4CF2O)4C2F4]2 | 1746.26 | 53.0 | 1.804 |
| PFC (% v/v) | 0.3 | 0.5 | 1 | 3 | 5 |
|---|---|---|---|---|---|
| PFH | 0.363 ± 0.015 | 0.368 ± 0.017 | 0.374 ± 0.017 | 0.351 ± 0.014 | 0.349 ± 0.013 |
| PFD | 0.345 ± 0.012 | 0.345 ± 0.012 | 0.345 ± 0.012 | 0.346 ± 0.012 | 0.345 ± 0.012 |
| Polyether I | 0.352 ± 0.014 | 0.357 ± 0.015 | 0.358 ± 0.014 | 0.353 ± 0.014 | 0.347 ± 0.012 |
| Polyether II | 0.367 ± 0.016 | 0.369 ± 0.016 | 0.376 ± 0.017 | 0.352 ± 0.014 | 0.349 ± 0.013 |
| PFCs | Concentration (% v/v) | |||||
|---|---|---|---|---|---|---|
| 0.3 | 0.5 | 1 | 2 | 3 | 5 | |
| Perfluorohexane | 389.4 ± 19.2 | 410.7 ± 19.8 | 384.8 ± 18.9 | 325.6 ± 18.0 | 335.0 ± 16.2 | 271.9 ± 13.2 |
| Perfluorodecalin | 334.4 ± 16.1 | 339.9 ± 16.8 | 341.7 ± 16.8 | 360.1 ± 16.9 | 337.0 ± 16.5 | 325.8 ± 15.3 |
| Polyether I | 373.9 ± 18.3 | 384.2 ± 18.7 | 396.7 ± 19.6 | 337.7 ± 19.2 | 371.6 ± 18.2 | 350.3 ± 16.5 |
| Polyether II | 361.7 ± 18.0 | 441.1 ± 21.2 | 463.5 ± 22.3 | 383.5 ± 20.7 | 380.0 ± 18.6 | 308.2 ± 14.9 |
| E. coli Strain | Yield of Biomass (CDW/L) | Amax * in the Cells (U/g) | Process Productivity (U/L/h) | Total His6-OPH Activity (U/L) |
|---|---|---|---|---|
| DH5a [33] | 2.8 | 130 | 17.3 | 364.0 |
| 5.8 | 44 | 12.2 | 255.2 | |
| W3110 [34] | 2.7 | 38 | 5.1 | 102.6 |
| SG13009[pREP4] [34] | 4.3 | 34 | 6.1 | 146.2 |
| SG13009[pREP4] (this work) | a 3.7 | 88 | 13.6 | 325.6 |
| b 5.1 | 91.5 | 19.3 | 463.5 | |
| c 4.4 | 93.3 | 17.1 | 410.7 |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Senko, O.; Stepanov, N.; Tyutyunov, A.; Sterlin, S.; Grinberg, V.; Makhlis, T.; Efremenko, E. Intensification of Organophosphorus Hydrolase Synthesis by Using Substances with Gas-Transport Function. Appl. Sci. 2017, 7, 1305. https://doi.org/10.3390/app7121305
Senko O, Stepanov N, Tyutyunov A, Sterlin S, Grinberg V, Makhlis T, Efremenko E. Intensification of Organophosphorus Hydrolase Synthesis by Using Substances with Gas-Transport Function. Applied Sciences. 2017; 7(12):1305. https://doi.org/10.3390/app7121305
Chicago/Turabian StyleSenko, Olga, Nikolay Stepanov, Andrey Tyutyunov, Sergey Sterlin, Vitaly Grinberg, Tatiana Makhlis, and Elena Efremenko. 2017. "Intensification of Organophosphorus Hydrolase Synthesis by Using Substances with Gas-Transport Function" Applied Sciences 7, no. 12: 1305. https://doi.org/10.3390/app7121305
APA StyleSenko, O., Stepanov, N., Tyutyunov, A., Sterlin, S., Grinberg, V., Makhlis, T., & Efremenko, E. (2017). Intensification of Organophosphorus Hydrolase Synthesis by Using Substances with Gas-Transport Function. Applied Sciences, 7(12), 1305. https://doi.org/10.3390/app7121305