Valorization of Waste Glycerol to Dihydroxyacetone with Biocatalysts Obtained from Gluconobacter oxydans



Abstract
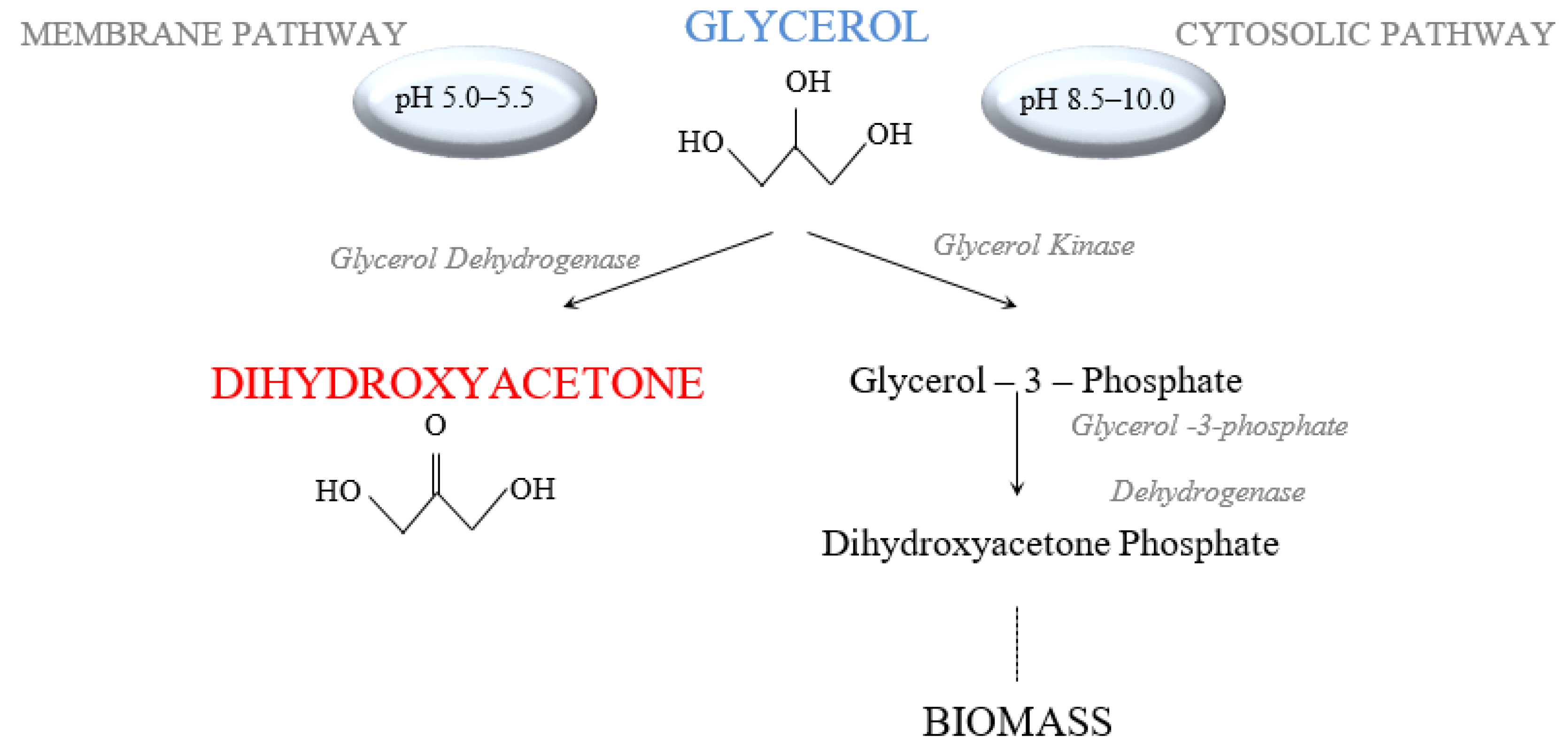
:1. Introduction
2. Materials and Methods
2.1. Biological Material
2.2. Media Composition and Culture Conditions
2.3. Preparation of Biocatalysts
2.4. Utilization of a Waste G·Lycerol
2.5. Determination of the Concentration of Glycerol and Dihydroxyacetone by Gas Chromatography
2.6. Analysis
3. Results
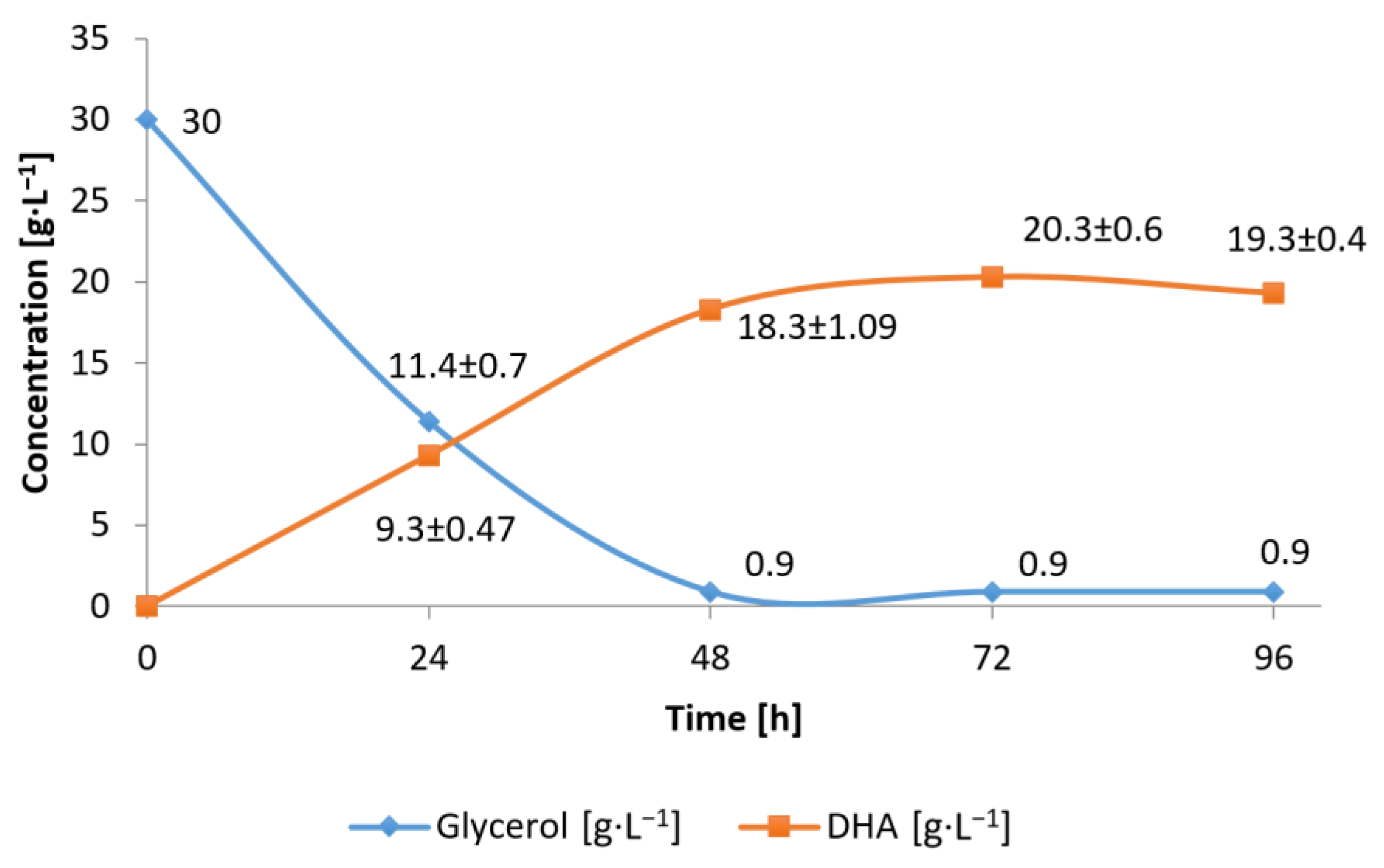
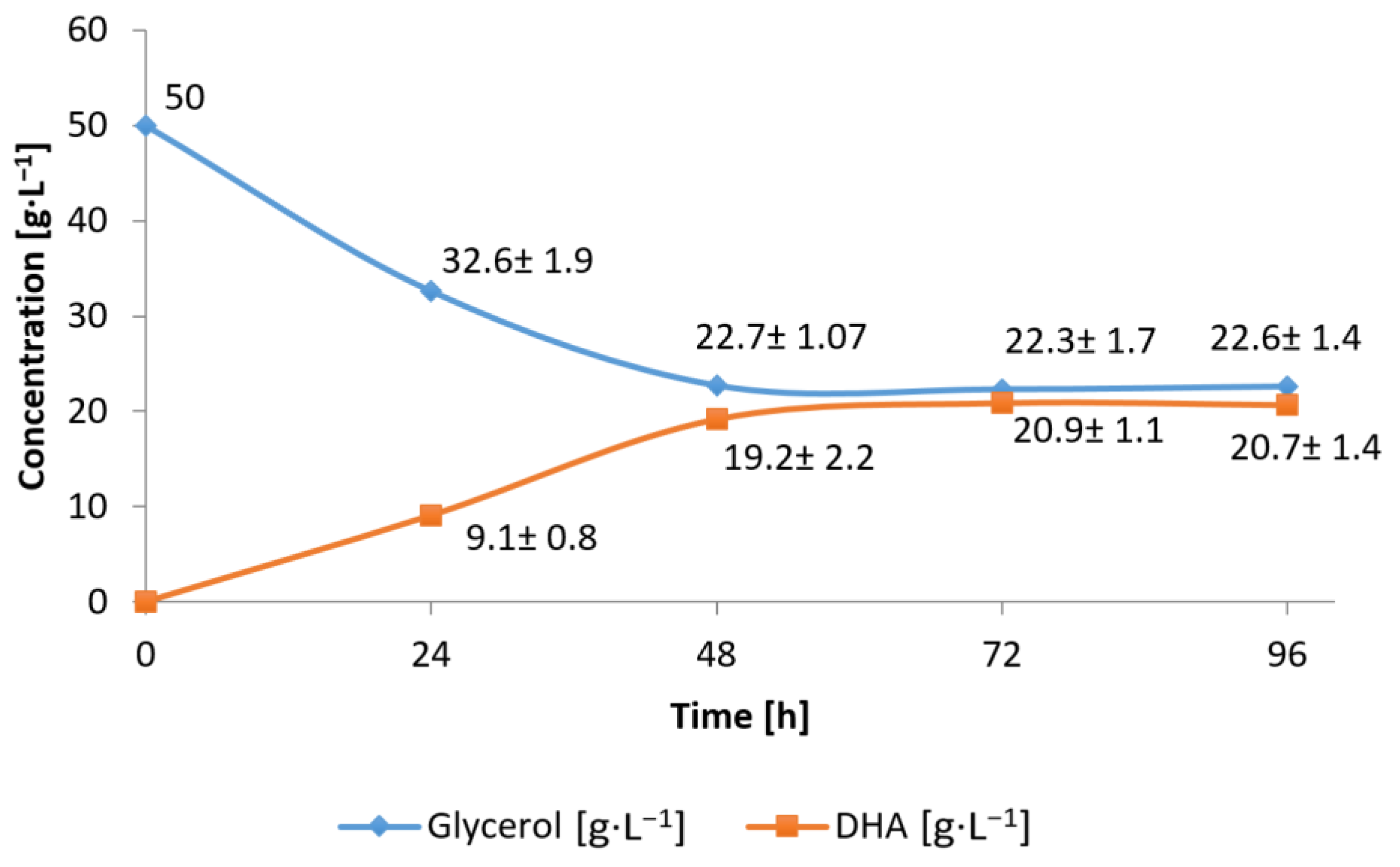
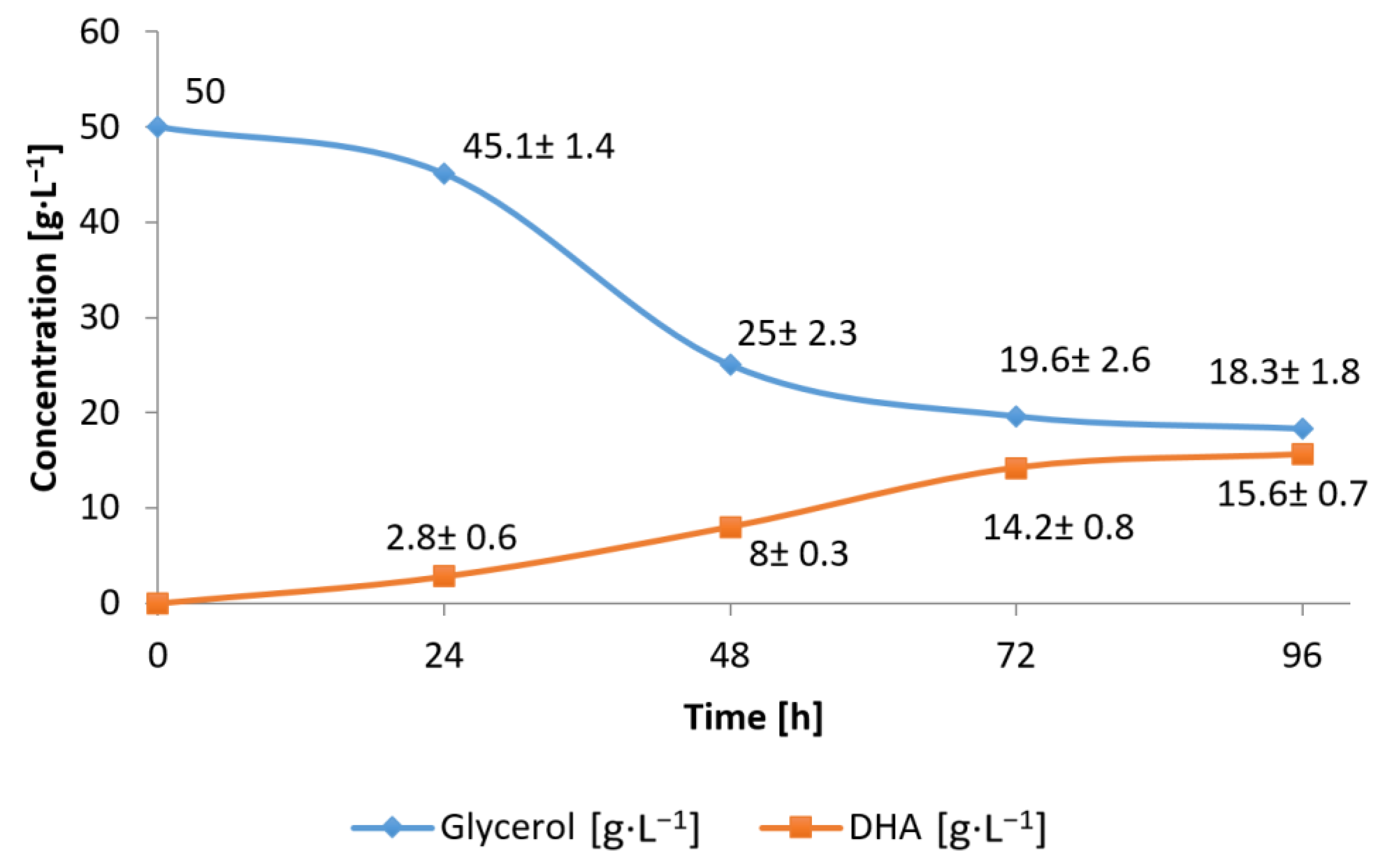
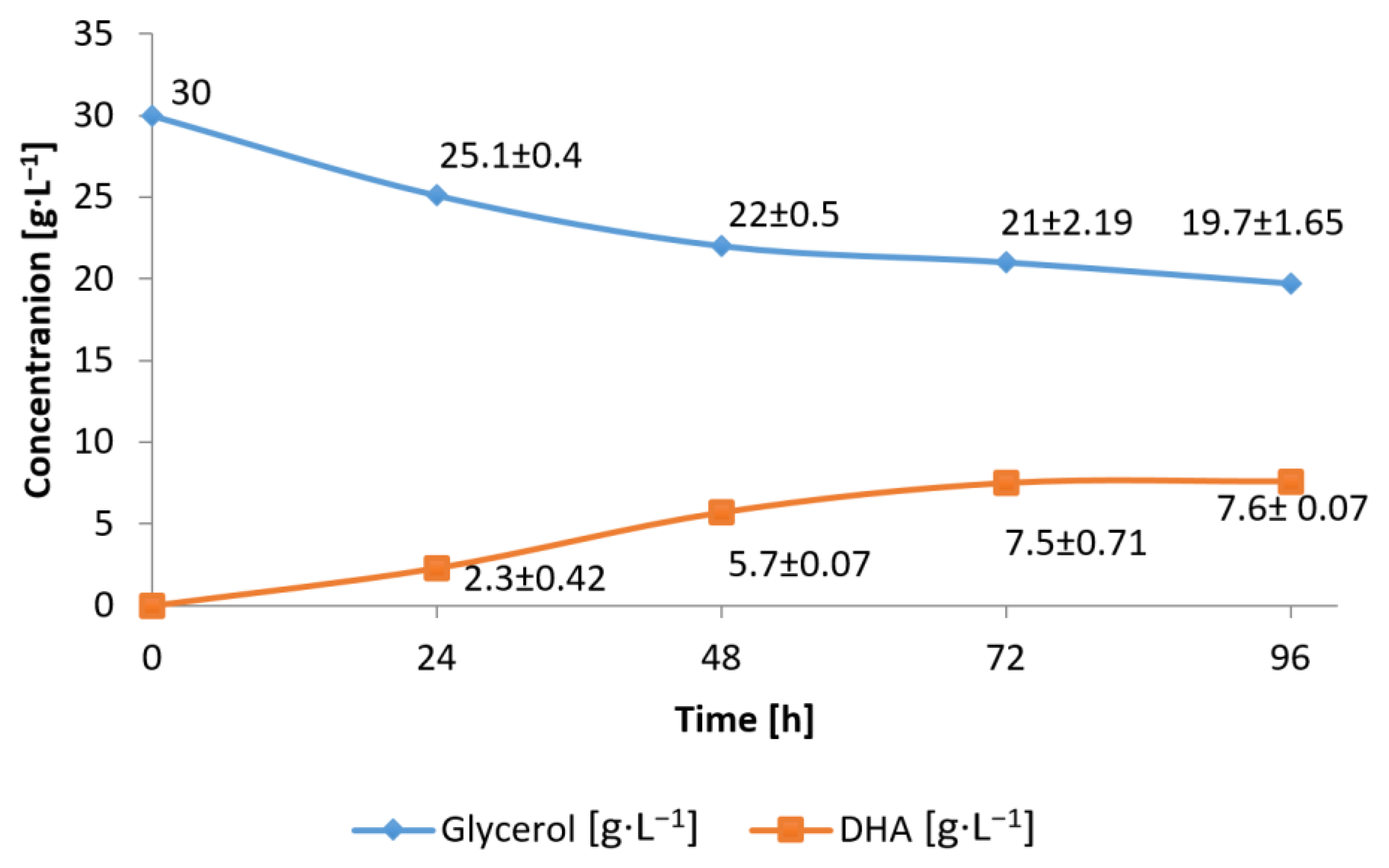
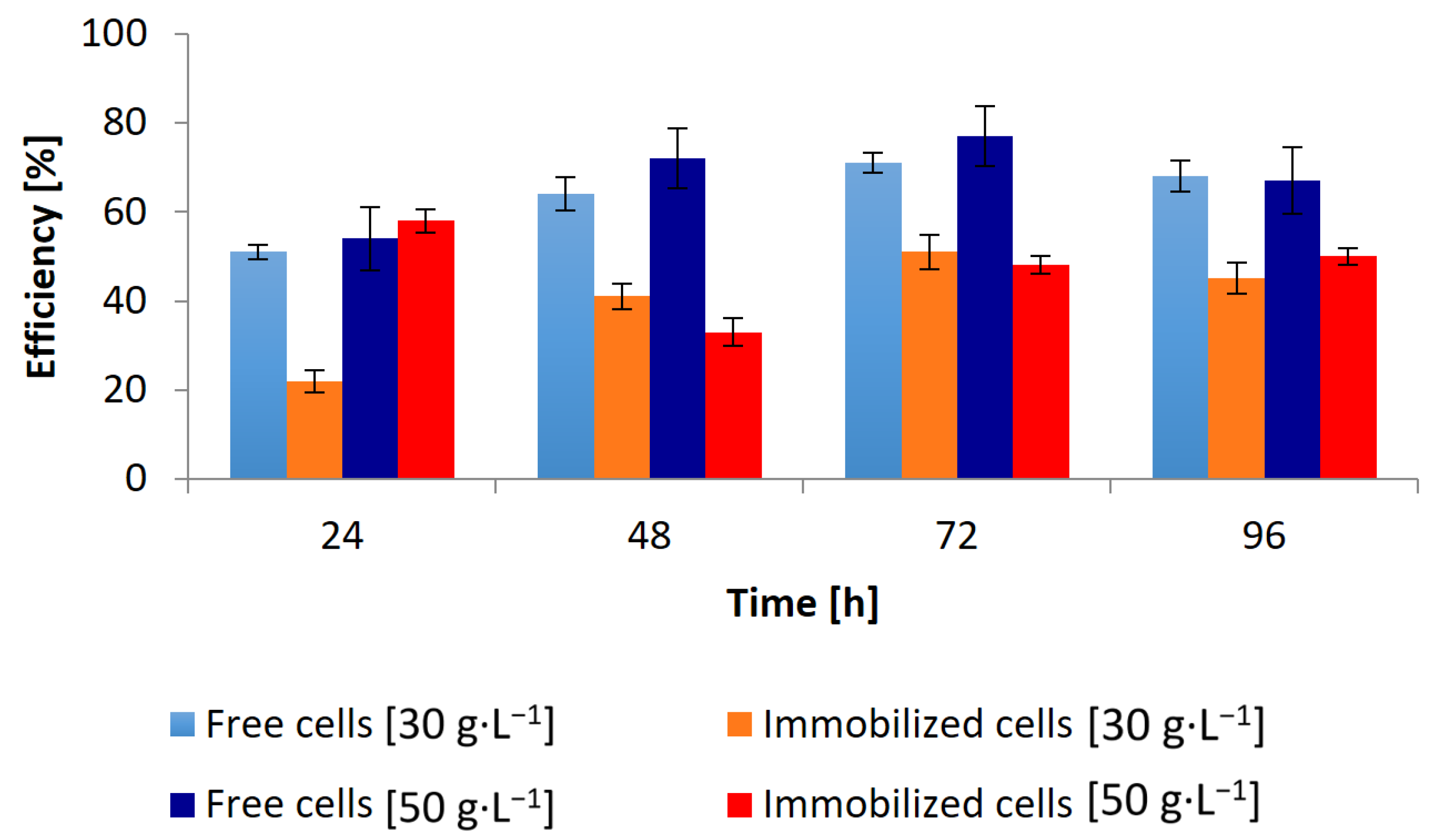
3.1. Conversion of Waste Glycerol with Free and Immobilized Cells of G. oxydans ATCC 621
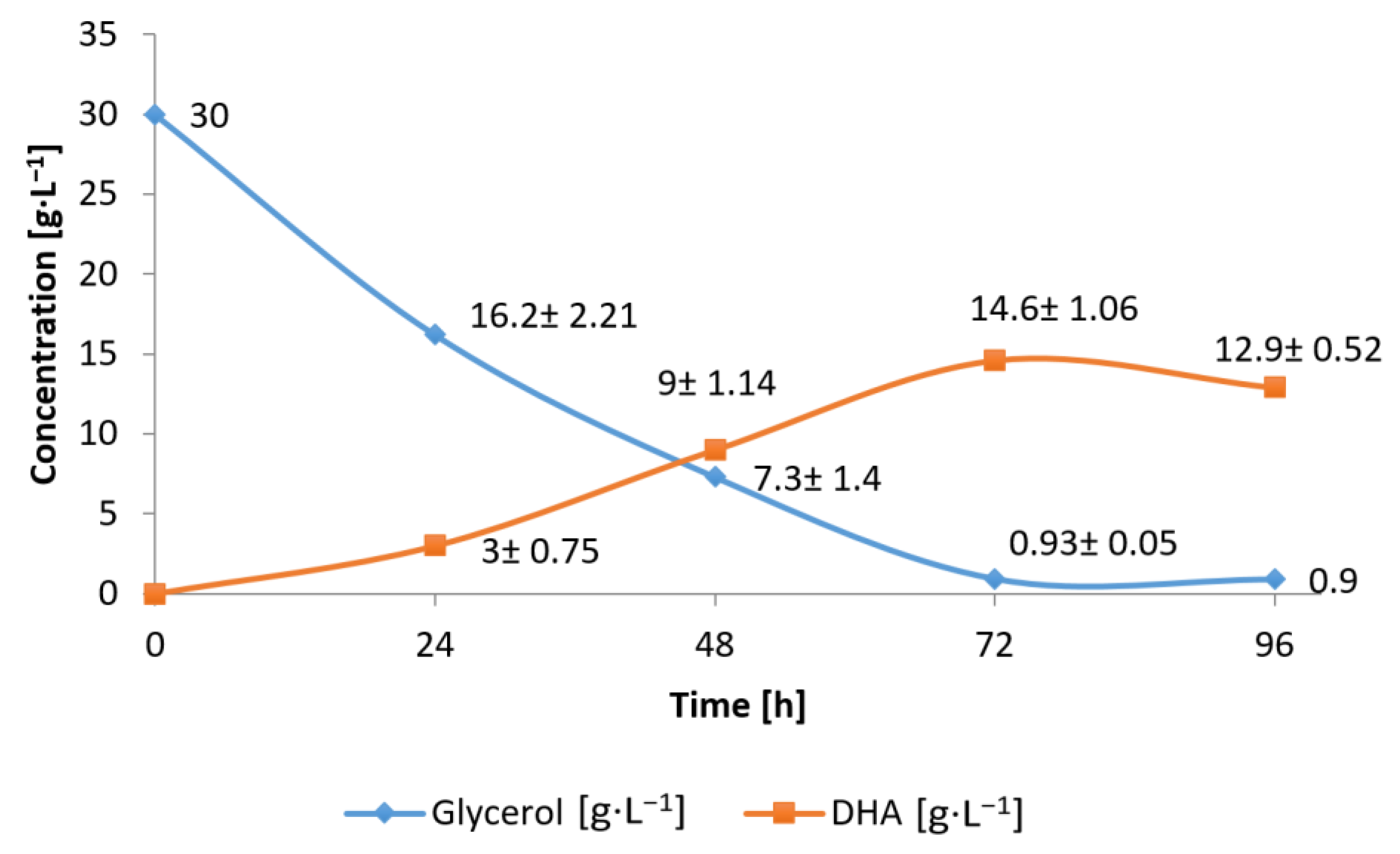
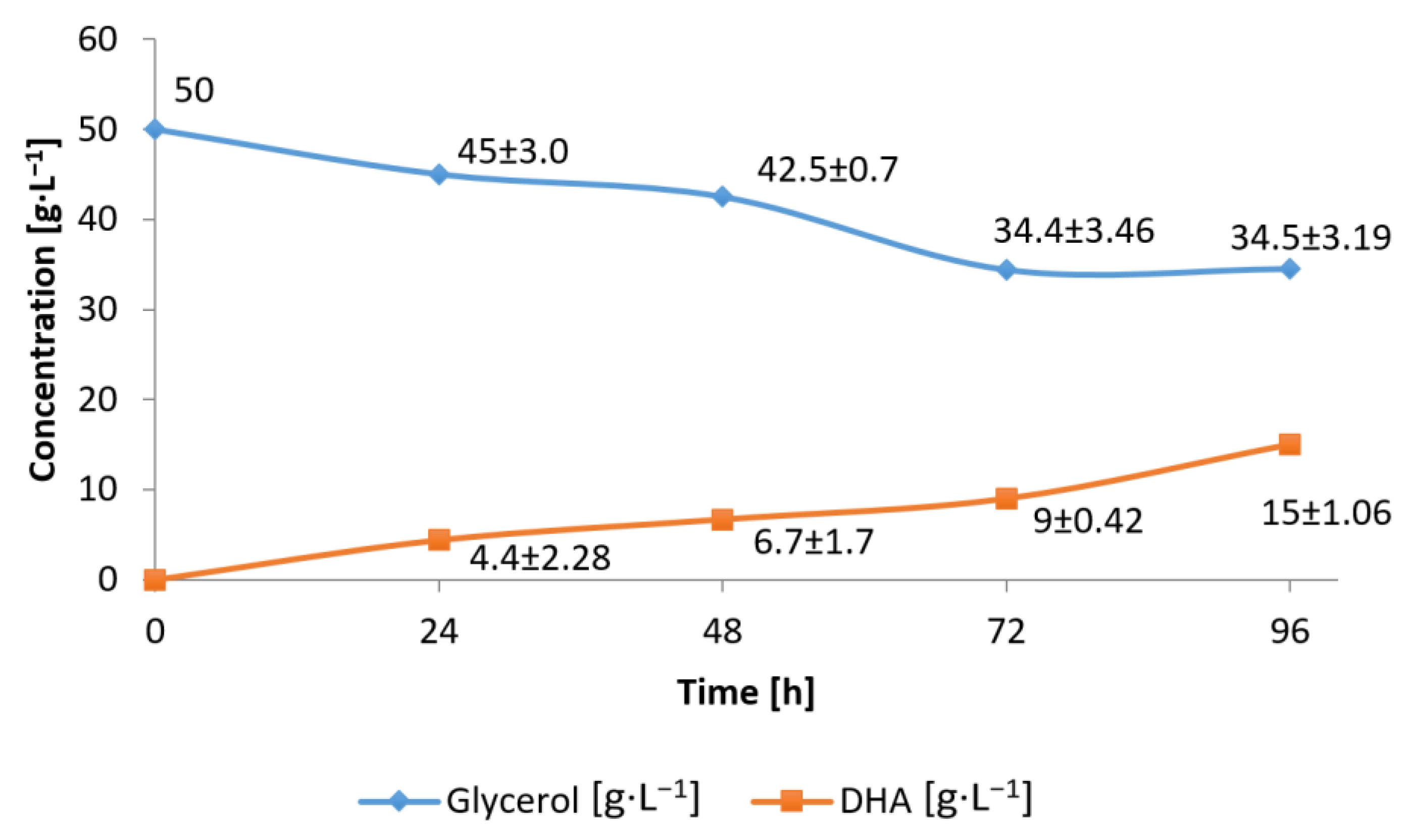
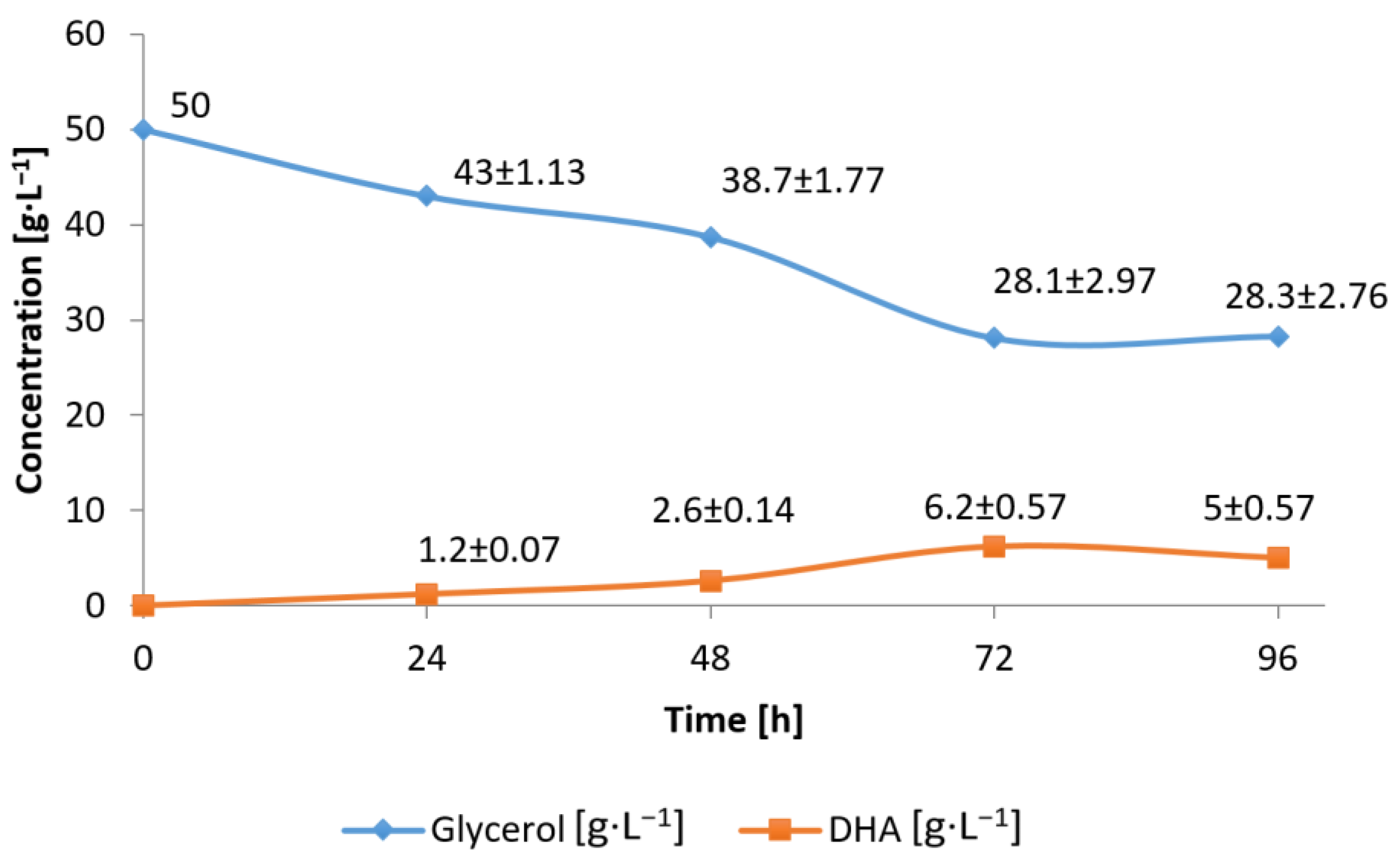
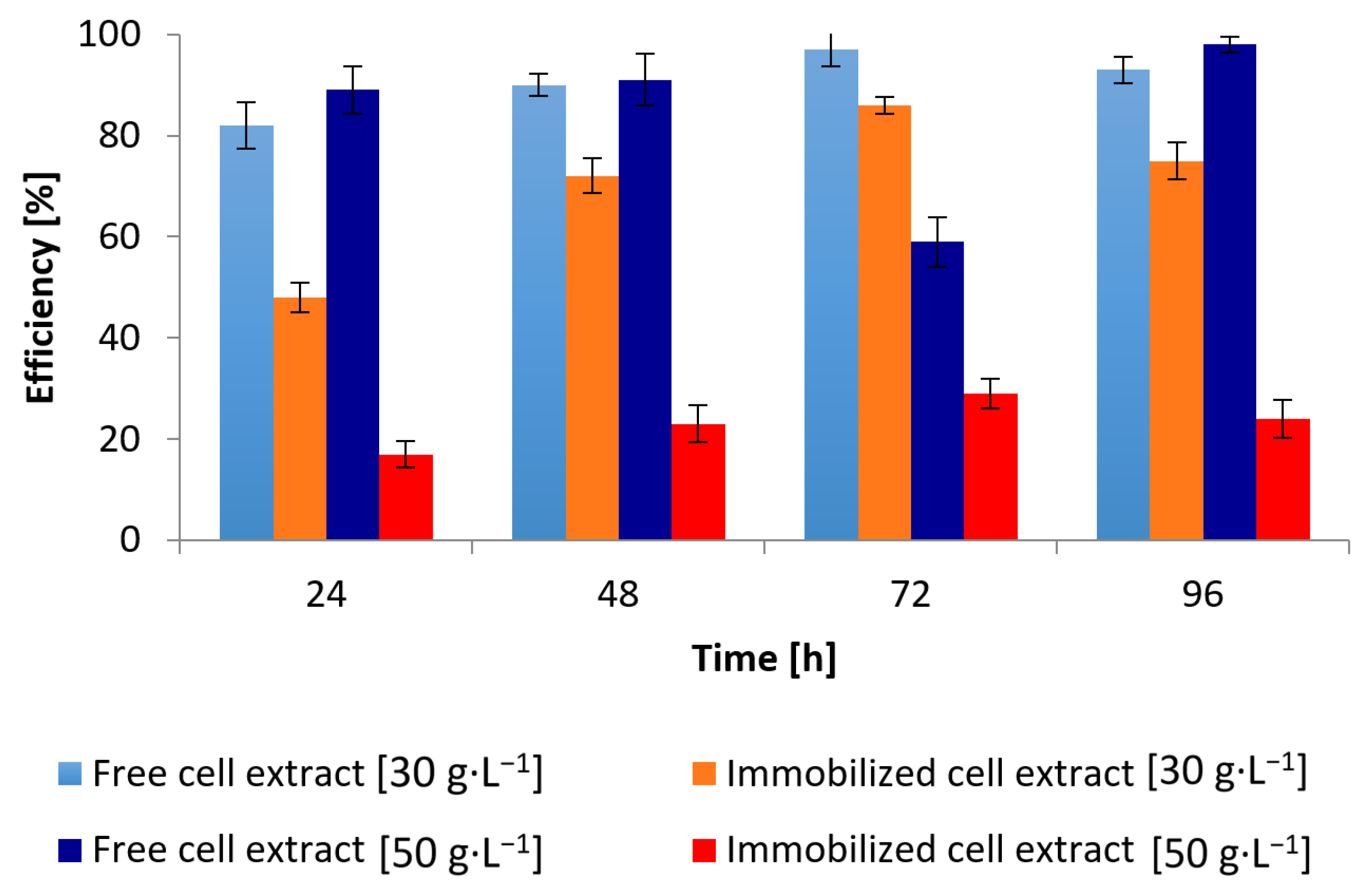
3.2. Conversion of Waste Glycerol with Free and Immobilized Cell Extract Obtained from G. oxydans ATCC 621
3.3. Substrate Consumption Rate and DHA Production Yield during Bioconversion of Waste Glycerol by G. oxydans ATCC 621 Cells and Their Extract
3.4. Efficiency of Waste Glycerol Bioconversion by G. oxydans ATCC 621 Cells and Their Extracts
4. Discussion
Author Contributions
Funding
Conflicts of Interest
References
- Binhayeeding, N.; Klomklao, S.; Sangkharak, K. Utilization of Waste G·Lycerol from Biodiesel Process as a Substrate for Mono-, Di-, and Triacylg·Lycerol Production. In Proceedings of the International Conference on Alternative Energy in Developing Countries and Emerging Economies, Bangkok, Thailand, 25–26 May 2017; Volume 138, pp. 895–900. [Google Scholar] [CrossRef]
- Manara, P.; Zabaniotou, A. Co-valorization of crude g·Lycerol waste streams with conventional and/or renewable fuels for power generation and industrial symbiosis perspectives. Waste Biomass Valoriz. 2016, 7, 135–150. [Google Scholar] [CrossRef]
- Vasudevan, P.T.; Fu, B. Environmentally sustainable biofuels: Advances in biodiesel research. Waste Biomass Valoriz. 2010, 1, 47–63. [Google Scholar] [CrossRef]
- Stasiak-Różańska, L.; Błażejak, S.; Gientka, I.; Bzducha-Wróbel, A.; Lipińska, E. Utilization of a waste g·Lycerol fraction using and reusing immobilized G·Luconobacter oxydans ATCC 621 cell extract. Electron. J. Biotechnol. 2017, 27, 44–48. [Google Scholar] [CrossRef]
- Mostafa, N.A.; Maher, A.; Abdelmoez, W. Production of mono-, di-, and trig·Lycerides from waste fatty acids through esterification with g·Lycerol. Adv. Biosci. Biotechnol. 2013, 4, 900–907. [Google Scholar] [CrossRef]
- Quispe, C.A.G.; Coronado, C.J.R.; Carvalho, J.A., Jr. G·Lycerol: production, consumption, prices, characterization and new trends in combustion. Renew. Sustain. Energy Rev. 2013, 27, 475–493. [Google Scholar] [CrossRef]
- Nitayavardhana, S.; Khanal, S.K. Biodiesel-derived crude g·Lycerol bioconversion to animal feed: A sustainable option for a biodiesel refinery. Bioresour. Technol. 2011, 102, 5808–5814. [Google Scholar] [CrossRef]
- Bala-Litwiniak, A.; Radomiak, H. Pissibility of the utilization of waste g·Lycerol as an addition to wood pellets. Waste Biomass Valoriz. 2018, 9, 1–7. [Google Scholar] [CrossRef]
- Leoneti, A.B.; Aragão-Leoneti, V.; de Oliveira, S.V.W.B. G·Lycerol as a by-product of biodiesel production in Brazil: Alternatives for the use of unrefined g·Lycerol. Renew. Energ. 2012, 45, 138–145. [Google Scholar] [CrossRef]
- Poljungreed, I.; Boonyarattanakalin, S. Low-cost biotransformation of g·Lycerol to 1,3-dihydroxyacetone through G·Luconobacter frateurii in medium with inorganic salts only. Lett. Appl. Microbiol. 2018, 67, 39–46. [Google Scholar] [CrossRef] [PubMed]
- Hekmat, D.; Bauer, R.; Neff, V. Optimization of the microbial synthesis of dihydroxyacetone in a semi–continuous repeated–fed–batch process by in situ immobilization of G·Luconobacter oxydans. Process Biochem. 2007, 42, 71–76. [Google Scholar] [CrossRef]
- Dikshit, P.K.; Moholkar, V.S. Optimization of 1,3-dihydroxyacetone production from crude g·Lycerol by immobilized G·Luconobacter oxydans MTCC 904. Bioresour. Technol. 2016, 216, 1058–1065. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Z.; Li, G.; Li, Y. Immobilization of Saccharomyces cerevisiae alcohol dehydrogenase on hybrid alginate-chitosan beads. Int. J. Biol. Macromol. 2010, 3, 21–26. [Google Scholar] [CrossRef] [PubMed]
- Mamlouk, D.; Gullo, M. Acetic Acid Bacteria: Physiology and Carbon Sources Oxidation. Indian J Microbiol. 2013, 53, 377–384. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stasiak-Różańska, L.; Błażejak, S. Dihydroxyacetone—Characterictic, application and recive. Acta Sci. Pol. Biotechnol. 2011, 11, 17–28. [Google Scholar]
- Ahmadi, J.; Joukar, S.; Anani, H.; Karami-Mohajeri, S. Dihydroxyacetone as a definitive treatment for aluminium phosphide poisoning in rats. Arch. Ind. Hyg. Toxicol. 2018, 69, 169–177. [Google Scholar] [CrossRef]
- Braunberger, T.L.; Nahhas, A.F.; Katz, L.M.; Sadrieh, N.; Lim, H.W. Dihydroxyacetone: A review. J. Drugs Dermatol. 2018, 17, 387–391. Available online: https://www.ncbi.nlm.nih.gov/pubmed/29601614 (accessed on 8 August 2018).
- Ciriminna, R.; Fidalgo, A.; Ilharco, L.M.; Pag·Liaro, M. Dihydroxyacetone: An Updated Insight into an Important Bioproduct. Chem. Open 2018, 7, 233–236. [Google Scholar] [CrossRef] [Green Version]
- Stasiak-Różańska, L.; Płoska, J. Study on the use of microbial cellulose as a biocarrier for 1,3-dihydroxy-2-propanone and its potential application in industry. Polymers 2018, 10, 438. [Google Scholar] [CrossRef]
- Peters, B.; Mientus, M.; Kostner, D.; Junker, A.; Liebl, W.; Ehrenreich, A. Characterization of membrane-bound dehydrogenases from G·Luconobacter oxydans 621H via whole-cell activity assays using multideletion strains. Appl. Microbiol. Biotechnol. 2013, 97, 6397–6412. [Google Scholar] [CrossRef]
- Habe, H.; Fukuoka, T.; Morita, T.; Kitamoto, D.; Yakushi, T.; Matsushita, K.; Sakaki, K. Disruption of the membrane-bound alcohol dehydrogenase-encoding gene improved g·Lycerol use and dihydroxyacetone productivity in G·Luconobacter oxydans. Biosci. Biotechnol. Biochem. 2010, 74, 1391–1395. [Google Scholar] [CrossRef]
- De Muynck, C.; Pereira, C.S.S.; Naessens, M.; Parmentier, S.; Soetaert, W.; Vandamme, E.J. The Genus G·Luconobacter oxydans: Comprehensive Overview of Biochemistry and Biotechnological Applications. Crit. Rev. Biotechnol. 2007, 27, 147–171. [Google Scholar] [CrossRef]
- Ameyama, M.; Shinagawa, E.; Matsushita, K.; Adachi, O. Solubilization, Purification and Properties of Membrane-bound G·Lycerol Dehydrogenase from G·Luconobacter industrius. Agric. Biol. Chem. 1985, 49, 1001–1010. [Google Scholar] [CrossRef]
- Lapenaite, I.; Kurtinaitiene, B.; Razumiene, J.; Laurinavicius, V.; Marcinkeviciene, L.; Bachmatova, I.; Meskys, R.; Ramanavicius, A. Properties and analytical application of PQQ-dependent g·Lycerol dehydrogenase from G·Luconobacter sp. 33. Anal. Chim. Acta 2005, 549, 140–150. [Google Scholar] [CrossRef]
- Anitha, M.; Kamarudinab, S.K.; Koflia, N.T. The potential of g·Lycerol as a value-added commodity. Chem. Eng. J. 2016, 295, 119–130. [Google Scholar] [CrossRef]
- Stasiak-Różańska, L.; Błażejak, S. Production of dihydroxyacetone from an aqueous solution of g·Lycerol in the reaction catalyzed by an immobilized cell preparation of acetic acid bacteria G·Luconobacter oxydans ATCC 621. Eur. Food Res. Technol. 2012, 235, 1125–1132. [Google Scholar] [CrossRef]
- Ye, Z.; Song, J.; Zhu, E.; Song, X.; Chen, X.; Xiaoting, H. Alginate Adsorbent Immobilization Technique Promotes Biobutanol Production by Clostridium acetobutylicum Under Extreme Condition of High Concentration of Organic Solvent. Front. Microbiol. 2018, 9, 1071. [Google Scholar] [CrossRef] [PubMed]
- Orrego, D.; Zapata-Zapata, A.D.; Kim, D. Ethanol production from coffee mucilage fermentation by S. cerevisiae immobilized in calcium-alginate beads. Bioresour. Technol. Rep. 2018, 3, 200–204. [Google Scholar] [CrossRef]
- Gutenwik, J.; Nilsson, B.; Axelsson, A. Mass transfer effects on the reaction rate for heterogeneously distributed immobilized yeast cells. Biotechnol. Bioeng. 2002, 79, 664–673. [Google Scholar] [CrossRef] [PubMed]
- Chatzifragkou, A.; Papanikolaou, S. Effect of impurities in biodiesel-derived waste g·Lycerol on the performance and feasibility of biotechnological processes. Appl. Microbiol. Biotechnol. 2012, 95, 13–27. [Google Scholar] [CrossRef]
- Prust, C.; Hoffmeister, M.; Liesegang, H.; Wiezer, A.; Fricke, W.F.; Ehrenreich, A.; Gottschalk, G.; Deppenmeier, U. Complete genome sequence of the acetic acid bacterium G·Luconobacter oxydans. Nat. Biotechnol. 2005, 23, 195–200. [Google Scholar] [CrossRef] [PubMed]
- VanLare, I.J.; Claus, G.W. Purification and properties of NAD(P)-independent polyol dehydrogenase complex from the plasma membrane of G·Luconobacter oxydans. Can. J. Microbiol. 2007, 53, 504–508. [Google Scholar] [CrossRef] [PubMed]
- Matsushita, K.; Fujii, Y.; Ano, Y.; Toyama, H.; Shinjoh, M.; Tomiyama, N.; Miyazaki, T.; Sugisawa, T.; Hoshino, T.; Adachi, O. 5-Keto-D-g·Luconate production is catalyzed by a quinoprotein g·Lycerol dehydrogenase, major polyol dehydrogenase, in G·Luconobacter species. Appl. Environ. Microbiol. 2003, 69, 1959–1966. [Google Scholar] [CrossRef] [PubMed]
- Mientus, M.; Kostner, D.; Peters, B.; Liebl, W.; Ehrenreich, A. Characterization of membrane-bound dehydrogenases of G·Luconobacter oxydans 621H using a new system for their functional expression. Appl. Microbiol. Biotechnol. 2017, 101, 3189–3200. [Google Scholar] [CrossRef]
- Ikemoto, K.; Mori, S.; Mukai, K. Synthesis and crystal structure of pyrroloquinoline quinol (PQQH2) and pyrroloquinoline quinone (PQQ). Acta Crystallogr. B Struct. Sci. Cryst. Eng. Mater. 2017, 73, 489–497. [Google Scholar] [CrossRef] [PubMed]
- Gupta, A.; Singh, V.K.; Qazi, G.N.; Kumar, A. G·Luconobacter oxydans: Its biotechnological applications. J. Mol. Microbiol. Biotechnol. 2001, 3, 445–456. Available online: https://www.ncbi.nlm.nih.gov/pubmed/11361077 (accessed on 10 August 2018). [PubMed]
- La China, S.; Zanichelli, G.; De Vero, L.; Gullo, M. Oxidative fermentations and exopolysaccharides production by acetic acid bacteria: A mini review. Biotechnol. Lett. 2018, 40, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Erni, B.; Siebold, C.; Christen, S.; Srinivas, A.; Oberholzer, A.; Baumann, U. Small substrate, big surprise: Fold, function and phylogeny of dihydroxyacetone kinases. Cell. Mol. Life Sci. 2006, 63, 890–900. [Google Scholar] [CrossRef]
- Dikshit, P.K.; Kharmawlong, G.J.; Moholkar, V.S. Investigations in sonication-induced intensification of crude g·Lycerol fermentation to dihydroxyacetone by free and immobilized G·Luconobacter oxydans. Bioresour. Technol. 2018, 256, 302–311. [Google Scholar] [CrossRef]












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Initial Glycerol (g·L−1) | Type of Biocatalysts | Time | ||||
|---|---|---|---|---|---|---|
| 24 h | 48 h | 72 h | 96 h | |||
| 30 | Free cells | Glycerol consumption (g·L−1 h−1) | 0.78 | 0.61 | 0.40 | 0.30 |
| DHA production (g·L−1 h−1) | 0.39 | 0.38 | 0.28 | 0.20 | ||
| DHA yield (gg−1) | 0.50 | 0,63 | 0,70 | 0,66 | ||
| Immobilized cells | Glycerol consumption (g·L−1 h−1) | 0.58 | 0.47 | 0.40 | 0.30 | |
| DHA production (g·L−1 h−1) | 0.12 | 0.19 | 0.20 | 0.13 | ||
| DHA yield (gg−1) | 0.22 | 0.40 | 0.50 | 0.44 | ||
| 50 | Free cells | Glycerol consumption (g·L−1 h−1) | 0.73 | 0.57 | 0.11 | 0.08 |
| DHA production (g·L−1 h−1) | 0.38 | 0.40 | 0.29 | 0.22 | ||
| DHA yield (gg−1) | 0.52 | 0.70 | 0.75 | 0.76 | ||
| Immobilized cells | Glycerol consumption (g·L−1 h−1) | 0.21 | 0.52 | 0.42 | 0.33 | |
| DHA production (g·L−1 h−1) | 0.12 | 0.17 | 0.20 | 0.16 | ||
| DHA yield (gg−1) | 0.56 | 0.32 | 0.47 | 0.49 | ||
| Initial Glycerol (g·L−1) | Type of Biocatalysts | Time | ||||
|---|---|---|---|---|---|---|
| 24 h | 48 h | 72 h | 96 h | |||
| 30 | Free cell extract | Glycerol consumption (g·L−1 h−1) | 0.21 | 0.30 | 0.24 | 0.18 |
| DHA production (g·L−1 h−1) | 0.18 | 0.27 | 0.22 | 0.16 | ||
| DHA yield (gg−1) | 0.81 | 0.88 | 0.94 | 0.91 | ||
| Immobilized cell extract | Glycerol consumption (g·L−1 h−1) | 0.21 | 0.17 | 0.12 | 0.11 | |
| DHA production (g·L−1 h−1) | 0.10 | 0.12 | 0.10 | 0.08 | ||
| DHA yield (gg−1) | 0.47 | 0.71 | 0.83 | 0.74 | ||
| 50 | Free cell extract | Glycerol consumption (g·L−1 h−1) | 0.21 | 0.16 | 0.21 | 0.16 |
| DHA production (g·L−1 h−1) | 0.18 | 0.14 | 0.12 | 0.16 | ||
| DHA yield (gg−1) | 0.88 | 0.89 | 0.58 | 0.97 | ||
| Immobilized cell extract | Glycerol consumption (g·L−1 h−1) | 0.29 | 0.23 | 0.31 | 0.23 | |
| DHA production (g·L−1 h−1) | 0.05 | 0.05 | 0.09 | 0.05 | ||
| DHA yield (gg−1) | 0.17 | 0.23 | 0.28 | 0.23 | ||
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Stasiak-Różańska, L.; Berthold-Pluta, A.; Dikshit, P.K. Valorization of Waste Glycerol to Dihydroxyacetone with Biocatalysts Obtained from Gluconobacter oxydans. Appl. Sci. 2018, 8, 2517. https://doi.org/10.3390/app8122517
Stasiak-Różańska L, Berthold-Pluta A, Dikshit PK. Valorization of Waste Glycerol to Dihydroxyacetone with Biocatalysts Obtained from Gluconobacter oxydans. Applied Sciences. 2018; 8(12):2517. https://doi.org/10.3390/app8122517
Chicago/Turabian StyleStasiak-Różańska, Lidia, Anna Berthold-Pluta, and Pritam Kumar Dikshit. 2018. "Valorization of Waste Glycerol to Dihydroxyacetone with Biocatalysts Obtained from Gluconobacter oxydans" Applied Sciences 8, no. 12: 2517. https://doi.org/10.3390/app8122517
APA StyleStasiak-Różańska, L., Berthold-Pluta, A., & Dikshit, P. K. (2018). Valorization of Waste Glycerol to Dihydroxyacetone with Biocatalysts Obtained from Gluconobacter oxydans. Applied Sciences, 8(12), 2517. https://doi.org/10.3390/app8122517