Impact of Copper Oxide Nanoparticles on Enhancement of Bioactive Compounds Using Cell Suspension Cultures of Gymnema sylvestre (Retz.) R. Br





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. CSC
2.2. Elicitation of CuO NPs in CSC
2.3. Extraction and Estimation of Gymnemic Acid II (GA II) in CSC
2.4. Total Phenolic Content (TPC) and Total Flavonoid Content (TFC) in CSC
2.5. Biological Activities in CSC
2.5.1. Extract Preparation
2.5.2. Antioxidant Activity
Radical Scavenging Activity
Reductive Potential
Phosphomolybdenum Method
2.5.3. Antidiabetic Activity
2.5.4. Anti-Inflammatory Activity
2.5.5. Antibacterial and Antifungal Activities
2.5.6. Anticancer Activity
2.6. Statistical Analyses
3. Results and Discussion
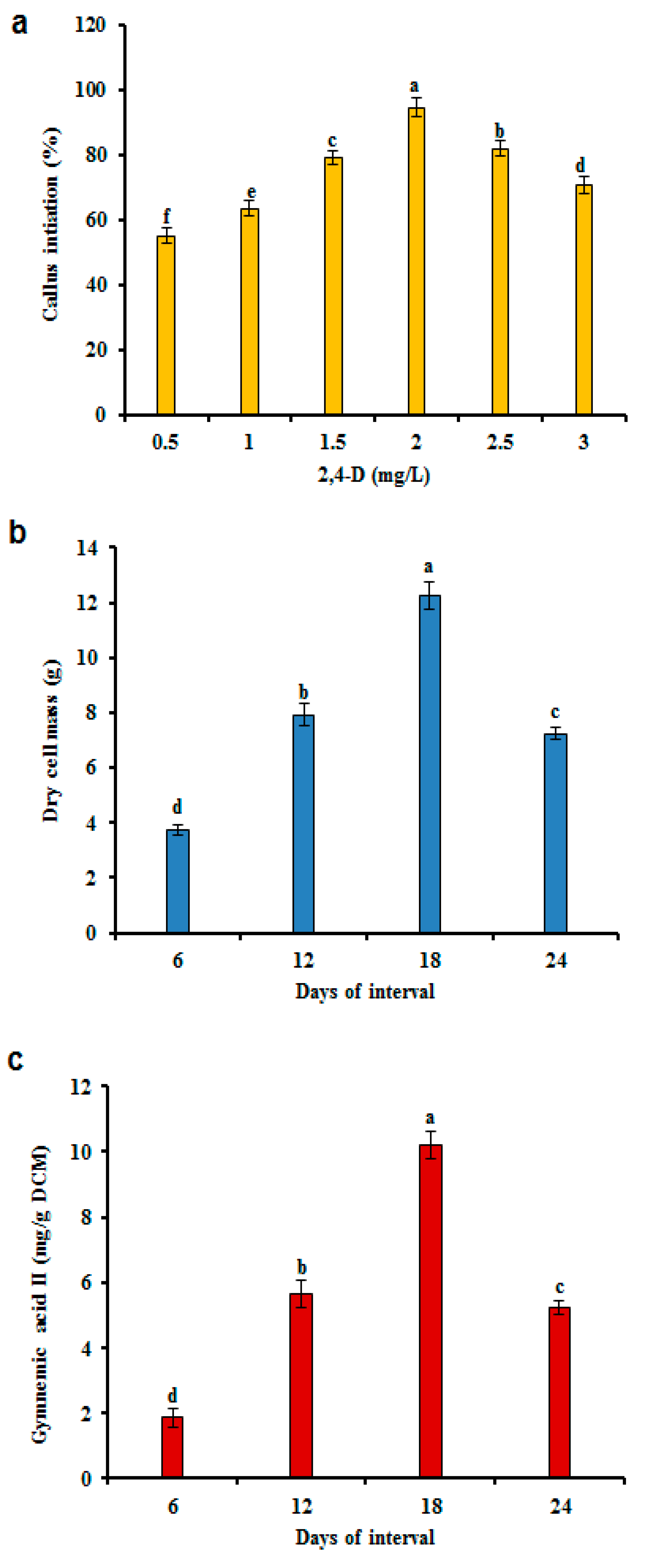
3.1. Gymnemic Acid II (GA II) Content in CSC
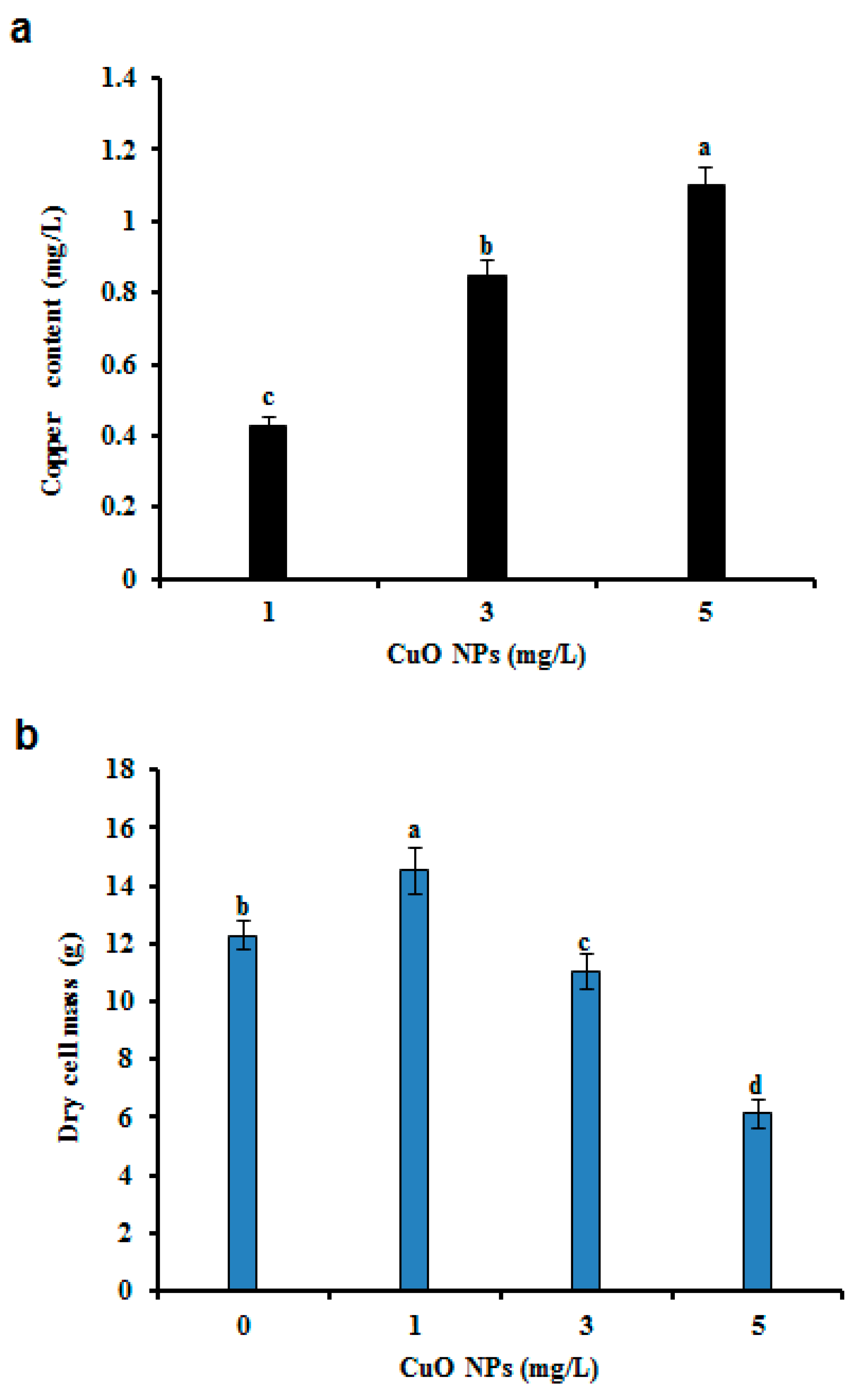
3.2. Biomass, Cu, and GA II Content in CuO NP Treated and Non-Treated CSC
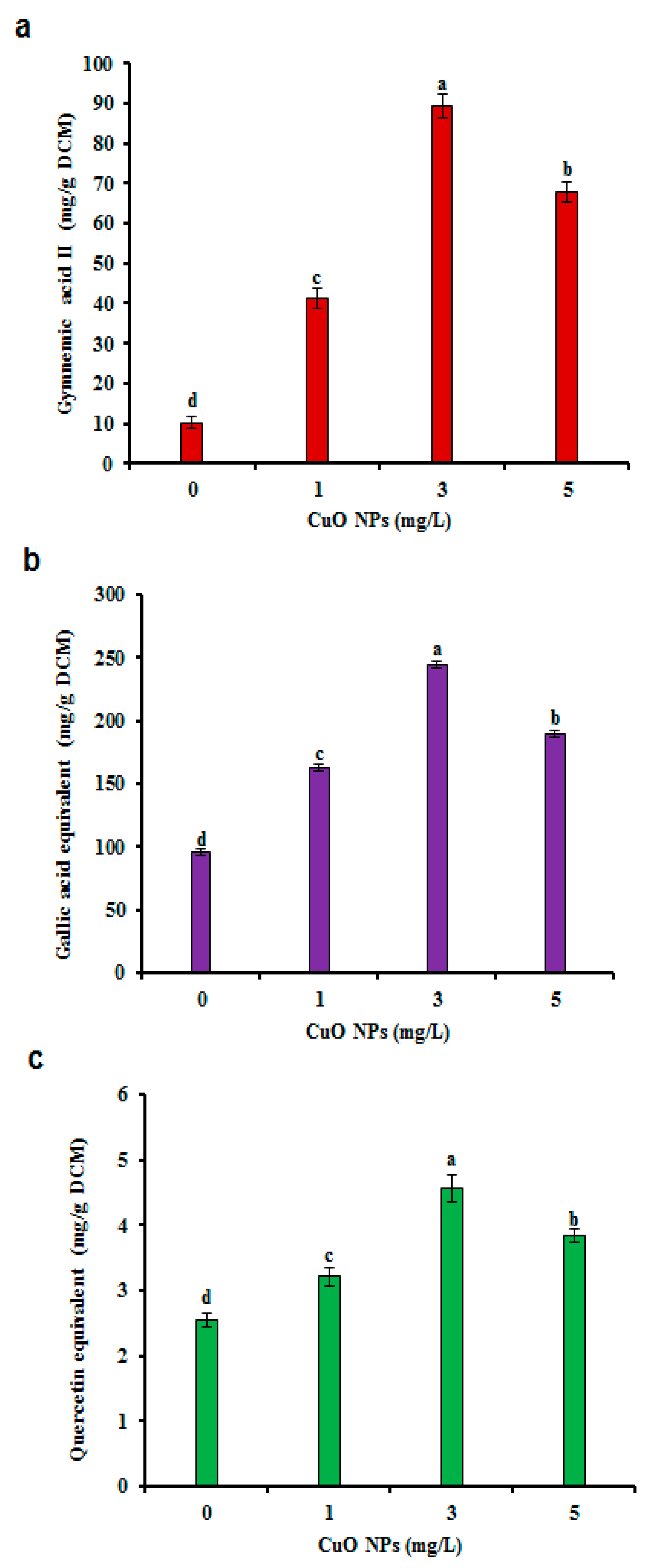
3.3. TPC and TFC in CSC
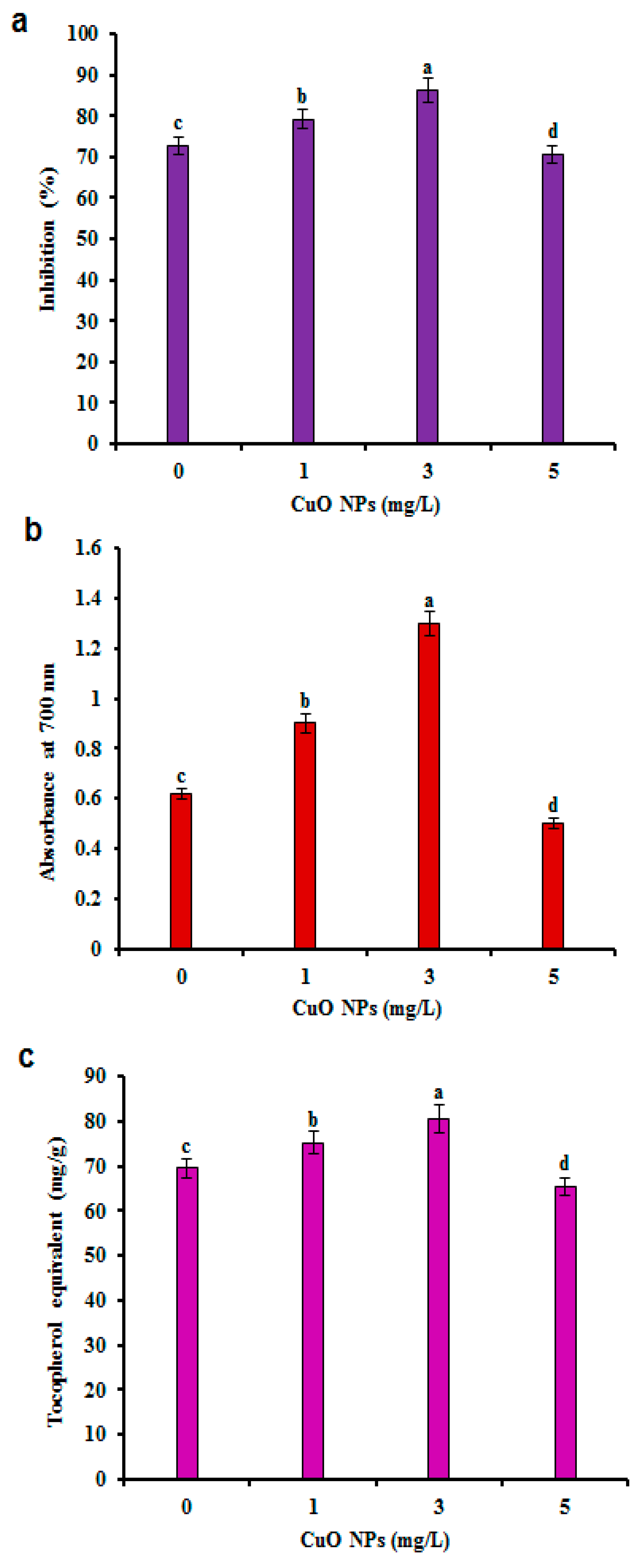
3.4. Antioxidant Activity
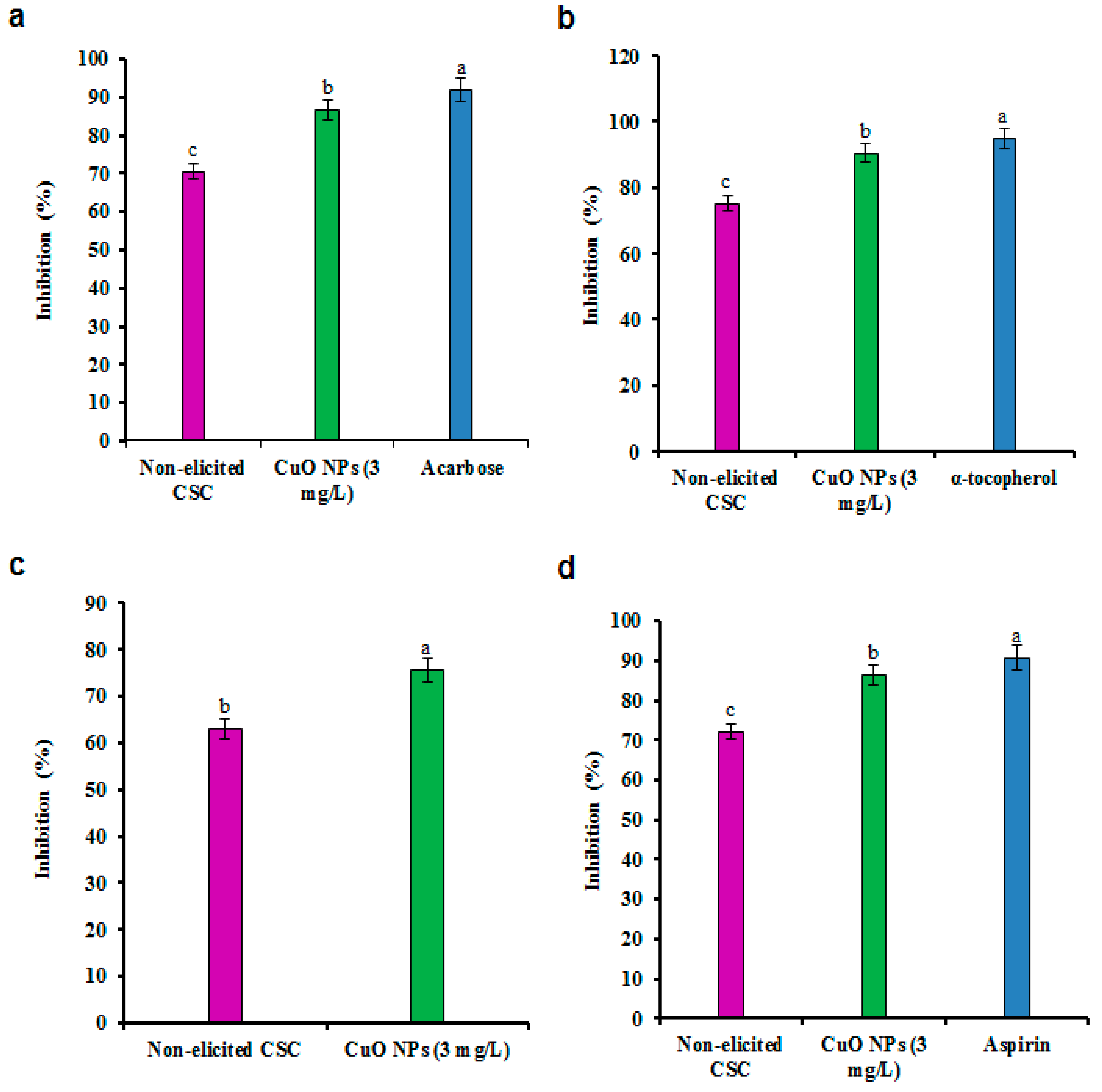
3.5. Antidiabetic Activity
3.6. Anti-Inflammatory Activity
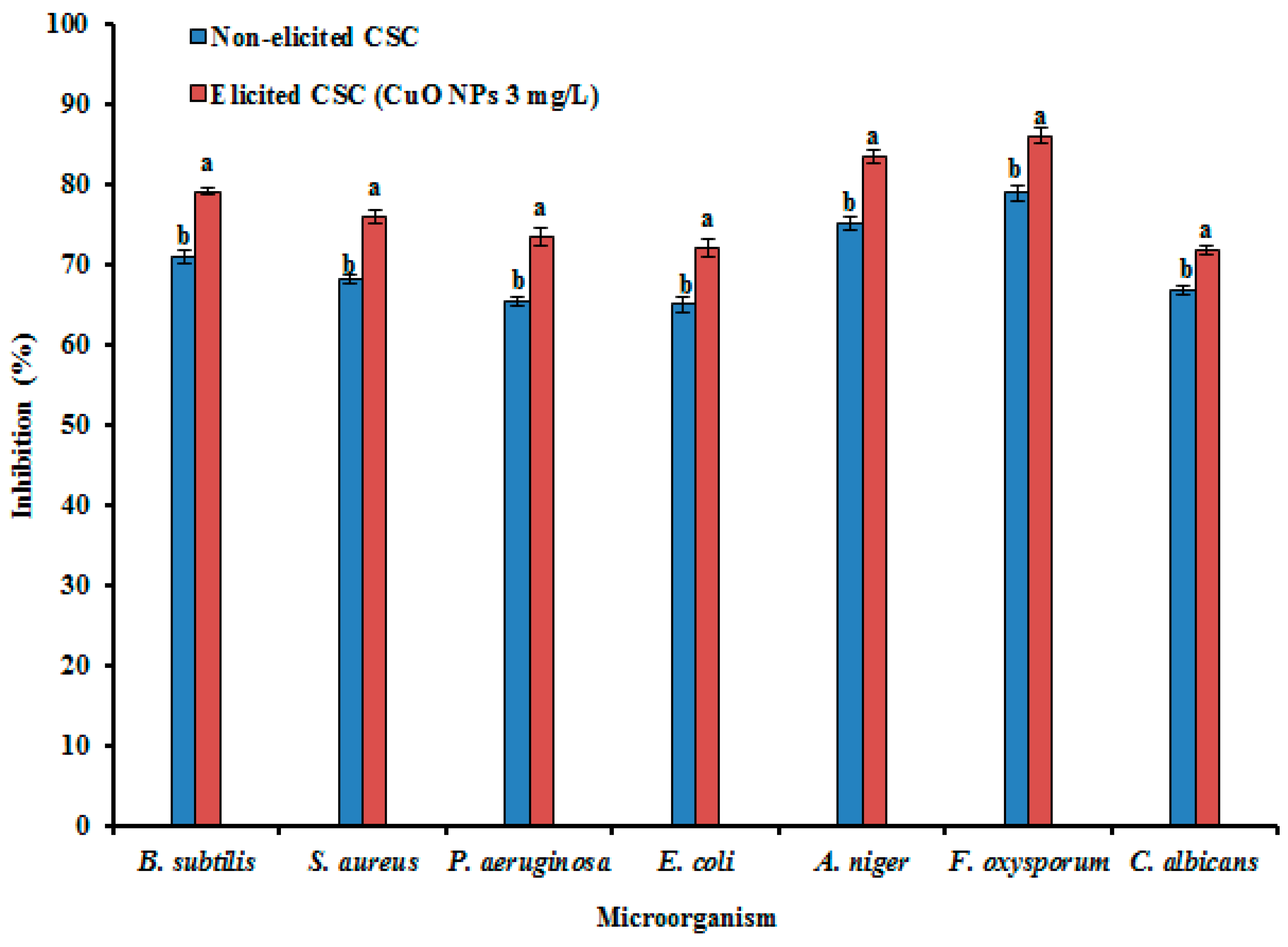
3.7. Antibacterial and Antifungal Activities
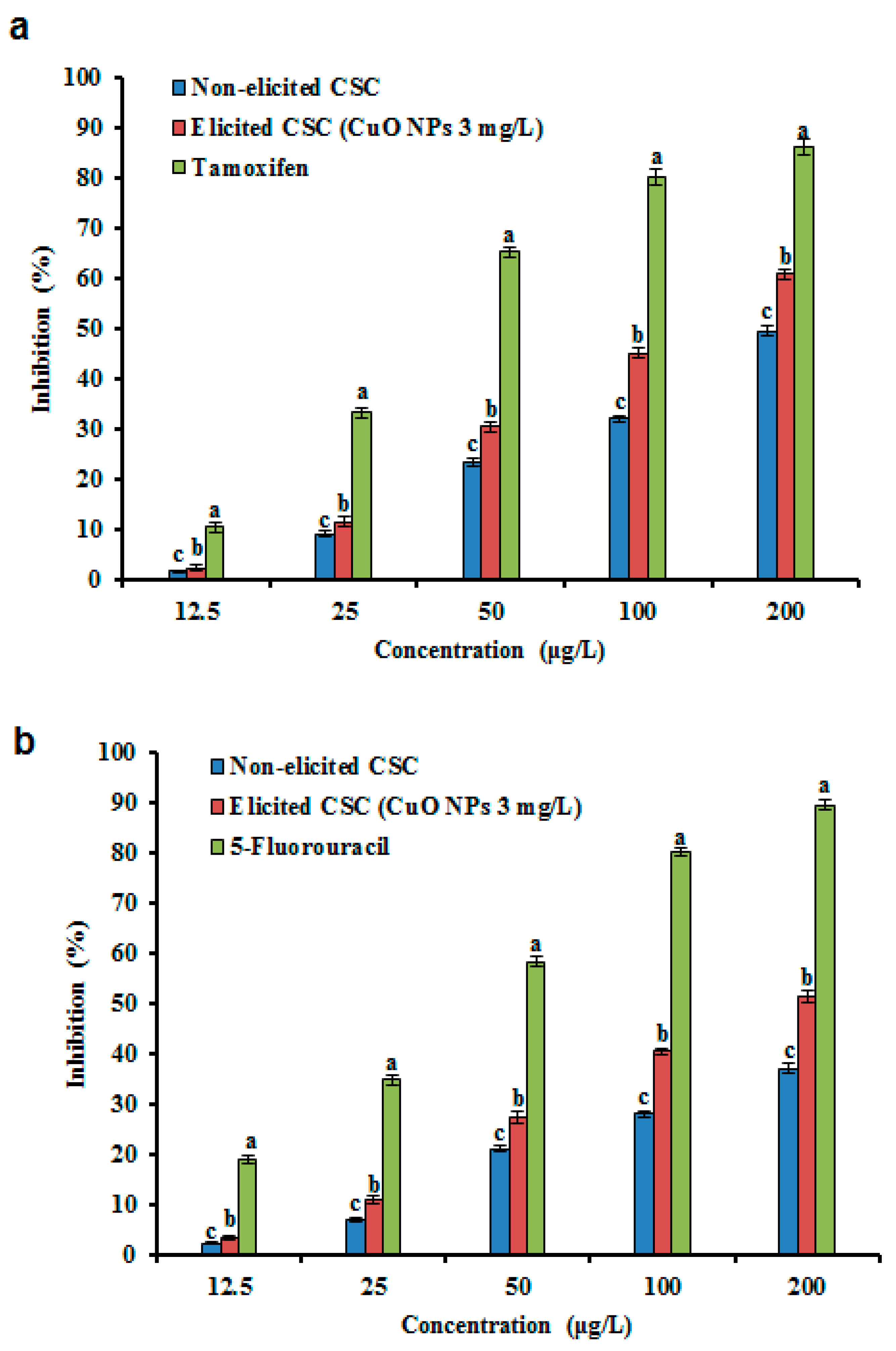
3.8. Anticancer Activity
4. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Pothuraju, R.; Sharma, R.K.; Chagalamarri, J.; Jangra, S.; Kumar Kavadi, P. A systematic review of Gymnema sylvestre in obesity and diabetes management. J. Sci. Food Agric. 2014, 94, 834–840. [Google Scholar] [CrossRef] [PubMed]
- Tiwari, P.; Mishra, B.N.; Sangwan, N.S. Phytochemical and pharmacological properties of Gymnema sylvestre: An important medicinal plant. BioMed Res. Int. 2014, 18, 830285. [Google Scholar]
- Nagella, P.; Thiruvengadam, M.; Jung, S.J.; Murthy, H.N.; Chung, I.M. Establishment of Gymnema sylvestre hairy root cultures for the production of gymnemic acid. Acta Physiol. Plant 2013, 35, 3067. [Google Scholar] [CrossRef]
- Praveen, N.; Thiruvengadam, M.; Yang, Y.S.; Kim, S.H.; Murthy, H.N.; Chung, I.M. Production of gymnemic acid from hairy root cultures of Gymnema sylvestre R. Br. as influenced by polyunsaturated fatty acids (PUFAs) and their antioxidant activity. Ind. Crop. Prod. 2014, 54, 54–61. [Google Scholar] [CrossRef]
- Georgiev, M.I.; Weber, J.; Maciuk, A. Bioprocessing of plant cell cultures for mass production of targeted compounds. Appl. Microbiol. Biotechnol. 2009, 83, 809–823. [Google Scholar] [CrossRef] [PubMed]
- Narayani, M.; Srivastava, S. Elicitation: A stimulation of stress in in vitro plant cell/tissue cultures for enhancement of secondary metabolite production. Phytochem. Rev. 2017, 16, 1227–1252. [Google Scholar] [CrossRef]
- Zhao, J.; Davis, L.C.; Verpoorte, R. Elicitor signal transduction leading to production of plant secondary metabolites. Biotechnol. Adv. 2005, 23, 283–333. [Google Scholar] [CrossRef]
- Tahsili, J.; Sharifi, M.; Safaie, N.; Esmaeilzadeh-Bahabadi, S.; Behmanesh, M. Induction of lignans and phenolic compounds in cell culture of Linum album by culture filtrate of Fusarium graminearum. J. Plant Interact. 2014, 9, 412–417. [Google Scholar] [CrossRef]
- Taghizadeh, M.; Nasibi, F.; Kalantari, K.M.; Ghanati, F. Evaluation of secondary metabolites and antioxidant activity in Dracocephalum polychaetum Bornm. cell suspension culture under magnetite nanoparticles and static magnetic field elicitation. Plant Cell Tiss. Org. Cult. 2019, 136, 489–498. [Google Scholar] [CrossRef]
- Ahmed, A.B.A.; Rao, A.S.; Rao, M.V. In vitro callus and in vivo leaf extract of Gymnema sylvestre stimulate β-cells regeneration and anti-diabetic activity in Wistar rats. Phytomedicine 2010, 17, 1033–1039. [Google Scholar] [CrossRef]
- Veerashree, V.; Anuradha, C.M.; Kumar, V. Elicitor-enhanced production of gymnemic acid in cell suspension cultures of Gymnema sylvestre R. Br. Plant Cell Tiss. Org. Cult. 2012, 108, 27–35. [Google Scholar] [CrossRef]
- Bhuvaneswari, C.; Rao, K.; Gandi, S.; Giri, A. Abiotic elicitation of gymnemic acid in the suspension cultures of Gymnema sylvestre. World J. Microbiol. Biotechnol. 2012, 28, 741–747. [Google Scholar]
- Yruela, I. Copper in plants. Braz. J. Plant Physiol. 2005, 17, 145–156. [Google Scholar] [CrossRef] [Green Version]
- Trettel, J.R.; Gazim, Z.C.; Gonçalves, J.E.; Stracieri, J.; Magalhães, H.M. Effects of copper sulphate (CuSO4) elicitation on the chemical constitution of volatile compounds and the in vitro development of Basil. Sci. Horticul. 2018, 234, 19–26. [Google Scholar] [CrossRef]
- Sharma, M.; Ahuja, A.; Gupta, R.; Mallubhotla, S. Enhanced bacoside production in shoot cultures of Bacopa monnieri under the influence of abiotic elicitors. Nat. Prod. Res. 2015, 29, 745–749. [Google Scholar] [CrossRef] [PubMed]
- Perotti, J.C.; Rodrigues, I.C.; Kleinowski, A.M.; Ribeiro, M.V.; Einhardt, A.M.; Peters, J.A.; Bacarin, M.A.; Braga, E.J.B. Betacyanin production in alligator weed, grown in vitro, with different concentrations of copper sulfate. Cienc. Rural. 2010, 40, 1874–1880. [Google Scholar] [CrossRef]
- Ahmed, B.; Khan, M.S.; Musarrat, J. Toxicity assessment of metal oxide nano-pollutants on tomato (Solanum lycopersicon): A study on growth dynamics and plant cell death. Environ. Pollut. 2018, 240, 802–816. [Google Scholar] [CrossRef] [PubMed]
- De la Rosa, G.; García-Castañeda, C.; Vázquez-Núñez, E.; Alonso-Castro, Á.J.; Basurto-Islas, G.; Mendoza, Á.; Cruz-Jiménez, G.; Molina, C. Physiological and biochemical response of plants to engineered NMs: Implications on future design. Plant Physiol. Biochem. 2017, 110, 226–235. [Google Scholar] [CrossRef] [PubMed]
- Fakruddin, M.D.; Hossain, Z.; Afroz, H. Prospects and applications of nanobiotechnology: A medical perspective. J. Nanobiotechnol. 2012, 10, 1–8. [Google Scholar] [CrossRef]
- Sharma, G.; Kumar, A.; Sharma, S.; Naushad, M.; Prakash Dwivedi, R.; ALOthman, Z.A.; Mola, G.T. Novel development of nanoparticles to bimetallic nanoparticles and their composites: A Review. J. King Saud Univ. Sci. 2019, 31, 257–269. [Google Scholar] [CrossRef]
- Huang, Y.; Li, W.; Minakova, A.S.; Anumol, T.; Keller, A.A. Quantitative analysis of changes in amino acids levels for cucumber (Cucumis sativus) exposed to nano copper. Nano Impact. 2018, 12, 9–17. [Google Scholar] [CrossRef]
- Juarez-Maldonado, A.; Ortega-Ortíz, H.; Pérez-Labrada, F.; Cadenas-Pliego, G.; Benavides-Mendoza, A. Cu nanoparticles absorbed on chitosan hydrogels positively alter morphological, production, and quality characteristics of tomato. J. Appl. Bot. Food Qual. 2016, 89, 183–189. [Google Scholar]
- Pinedo-Guerrero, Z.H.; Hernández-Fuentes, A.D.; Ortega-Ortiz, H.; Benavides-Mendoza, A.; Cadenas-Pliego, G.; Juárez-Maldonado, A. Cu nanoparticles in hydrogels of chitosan-PVA affects the characteristics of post-harvest and bioactive compounds of jalapeño pepper. Molecules 2017, 22, 926. [Google Scholar] [CrossRef] [PubMed]
- Fazal, H.; Abbasi, B.H.; Ahmad, N.; Ali, M. Elicitation of medicinally important antioxidant secondary metabolites with silver and gold nanoparticles in callus cultures of Prunella vulgaris L. Appl. Biochem. Biotechnol. 2016, 180, 1076–1092. [Google Scholar] [CrossRef] [PubMed]
- Chung, I.M.; Rekha, K.; Rajakumar, G.; Thiruvengadam, M. Elicitation of silver nanoparticles enhanced the secondary metabolites and pharmacological activities in cell suspension cultures of bitter gourd. 3 Biotech 2018, 10, 412. [Google Scholar] [CrossRef] [PubMed]
- Talankova-Sereda, T.E.; Liapina, K.V.; Shkopinskij, E.A.; Ustinov, A.I.; Kovalyova, A.V.; Dulnev, P.G.; Kucenko, N.I. The influence of Cu and Co nanoparticles on growth characteristics and biochemical structure of Mentha longifolia in vitro. Nanosci. Nanoeng. 2016, 4, 31–39. [Google Scholar]
- Genady, E.A.; Qaid, E.A.; Fahmy, A.H. Copper sulfate nanoparticales in vitro applications on Verbena bipinnatifida Nutt. Stimulating growth and total phenolic content increasments. Int. J. Pharm. Res. Allied Sci. 2016, 5, 196–202. [Google Scholar]
- Genady, E.A.; Ahmed, S.S.; Fahmy, A.H.; Ashour, R.M. Copper sulfate nanoparticles enhance growth parameters, flavonoid content and antimicrobial activity of Ocimum basilicum Linnaeus. J. Am. Sci. 2017, 13, 108–114. [Google Scholar]
- Chung, I.M.; Rekha, K.; Rajakumar, G.; Thiruvengadam, M. Production of bioactive compounds and gene expression alterations in hairy root cultures of Chinese cabbage elicited by copper oxide nanoparticles. Plant Cell Tiss. Org. Cult. 2018, 134, 95–106. [Google Scholar] [CrossRef]
- Murashige, T.; Skoog, F. A revised medium for rapid growth and bioassays with tobacco tissue cultures. Physiol. Plant. 1962, 15, 473–497. [Google Scholar] [CrossRef]
- Bao, J.S.; Cai, Y.; Sun, M.; Wang, G.; Corke, H. Anthocyanins, flavonols, and free radical scavenging activity of Chinese bayberry (Myrica rubra) extracts and their color properties and stability. J. Agric. Food Chem. 2005, 53, 2327–2332. [Google Scholar] [CrossRef] [PubMed]
- Brand-Williams, W.; Cuvelier, M.E.; Berset, C. Use of a free radical method to evaluate antioxidant activity. Lwt Food Sci. Technol. 1995, 28, 25–30. [Google Scholar] [CrossRef]
- Oyaizu, M. Studies on products of browning reactions: Antioxidative activities of browning reaction prepared from glucosamine. Jpn. J. Nutr. 1986, 44, 307–315. [Google Scholar] [CrossRef]
- Saleem, B.; Islam, M.; Saeed, H.; Imtiaz, F.; Asghar, M.; Saleem, Z.; Mehmood, A.; Naheed, S. Investigations of Acacia modesta Wall. leaves for in vitro anti-diabetic, proliferative and cytotoxic effects. Braz. J. Pharm. Sci. 2018, 54, e17467. [Google Scholar] [CrossRef]
- Shah, S.M.; Ashraf, M.; Ahmad, I.; Arshad, S.; Yar, M.; Latif, A. Anti-lipoxygenase activity of some indigenous medicinal plants. J. Med. Plants Res. 2013, 7, 219–222. [Google Scholar]
- Govindappa, M.; Hemashekhar, B.; Arthikala, M.K.; Ravishankar Rai, V.; Ramachandra, Y.L. Characterization, antibacterial, antioxidant, antidiabetic, anti-inflammatory and antityrosinase activity of green synthesized silver nanoparticles using Calophyllum tomentosum leaves extract. Results Phys. 2018, 9, 400. [Google Scholar] [CrossRef]
- Nagella, P.; Chung, I.M.; Murthy, H.N. In vitro production of gymnemic acid from cell suspension cultures of Gymnema Sylvestre R. Br. Eng. Life Sci. 2011, 11, 537–540. [Google Scholar] [CrossRef]
- Thiruvengadam, M.; Rekha, K.; Rajakumar, G.; Lee, T.J.; Kim, S.H.; Chung, I.M. Enhanced production of anthraquinones and phenolic compounds and biological activities in the cell suspension cultures of Polygonum multiflorum. Int. J. Mol. Sci. 2016, 17, 1912. [Google Scholar] [CrossRef]
- Van Assche, F.; Clijsters, H. Effects of metals on enzyme activity in plants. Plant Cell Environ. 1990, 13, 195–206. [Google Scholar] [CrossRef]
- Halliwell, B.; Gutteridge, J.M.C. Oxygen toxicity, oxygen radicals, transition metals and disease. Biochem. J. 1984, 219, 1–14. [Google Scholar] [CrossRef]
- Tahiliani, S.; Kothari, S.L. Increased copper content of the medium improves plant regeneration from immature embryos derived callus of wheat (Triticum aestivum). J. Plant Biochem. Biotechnol. 2004, 13, 85–88. [Google Scholar] [CrossRef]
- Javed, R.; Mohamed, A.; Yücesan, B.; Gürel, E.; Kausar, R.; Zia, M. CuO nanoparticles significantly influence in vitro culture, steviol glycosides, and antioxidant activities of Stevia rebaudiana. Bertoni. Plant Cell Tiss. Org. Cult. 2017, 131, 611–620. [Google Scholar] [CrossRef]
- Bhuvaneswari, C.; Rao, K.; Gandi, S.; Giri, A. Gymnemic acid enhancement in the suspension cultures of Gymnema sylvestre by using the signaling molecules-methyl jasmonate and salicylic acid. In Vitro Cell. Dev. Biol. Plant 2015, 51, 88–92. [Google Scholar]
- Zafar, H.; Ali, A.; Zia, M. CuO nanoparticles inhibited root growth from Brassica nigra seedlings but induced root from stem and leaf explants. Appl. Biochem. Biotechnol. 2017, 181, 365–378. [Google Scholar] [CrossRef]
- Nourozia, E.; Hosseini, B.; Maleki, R.; Mandoulakani, B.A. Pharmaceutical important phenolic compounds overproduction and gene expression analysis in Dracocephalum kotschyi hairy roots elicited by SiO2 nanoparticles. Ind. Crop. Prod. 2019, 133, 435–446. [Google Scholar] [CrossRef]
- Wang, J.W.; Wu, J.Y. Tanshinone biosynthesis in Salvia miltiorrhiza and production in plant tissue cultures. Appl. Microbiol. Biotechnol. 2010, 88, 437–449. [Google Scholar] [CrossRef]
- Večeřová, K.; Večeřa, Z.; Dočekal, B.; Oravec, M.; Pompeiano, A.; Tříska, J.; Urban, O. Changes of primary and secondary metabolites in barley plants exposed to CdO nanoparticles. Environ. Pollut. 2016, 218, 207–218. [Google Scholar] [CrossRef]
- Chung, I.M.; Rekha, K.; Venkidasamy, B.; Thiruvengadam, M. Effect of copper oxide nanoparticles on the physiology, bioactive molecules, and transcriptional changes in Brassica rapa ssp. rapa seedlings. Water Air Soil Pollut. 2019, 230, 48. [Google Scholar] [CrossRef]
- Janas, K.M.; Amarowicz, R.; Zielińska-Tomaszewska, J.; Kosińska, A.; Posmyk, M.M. Induction of phenolic compounds in two darkgrown lentil cultivars with different tolerance to copper ions. Acta Physiol. Plant. 2009, 31, 587–595. [Google Scholar] [CrossRef]
- Rachh, P.R.; Patel, S.R.; Hirpara, H.V.; Rupareliya, M.T.; Rachh, M.R.; Bhargava, A.S.; Patel, N.M.; Modi, D.C. In vitro evaluation of antioxidant activity of Gymnema sylvestre R. br. leaf extract. Rom. J. Biol. Plant Biol. 2009, 54, 141–148. [Google Scholar]
- Ruiz-Ruiz, J.C.; Matus-Basto, A.J.; Acereto-Escoffié, P.; Segura-Campos, M.R. Antioxidant and anti-inflammatory activities of phenolic compounds isolated from Melipona beecheii honey. Food Agric. Immunol. 2018, 28, 61424–61437. [Google Scholar] [CrossRef]
- Asmat, U.; Abad, K.; Ismail, K. Diabetes mellitus and oxidative stress - A concise review. Saudi Pharm. J. 2016, 24, 547–553. [Google Scholar] [CrossRef]
- Tripathi, Y.B.; Singh, V.P. Role of Tamra bhasma, an Ayurvedic preparation, in the management of lipid peroxidation in liver of albino rats. Indian J. Exp. Biol. 1996, 34, 66–70. [Google Scholar] [PubMed]
- Nirmala, S.; Ravichandiran, V.; Vijayalakshmi, A. In vitro assay of alpha amylase inhibitory activity of gymnemic acid isolated from Gymnema Sylvestre leaves. Der Pharm. Lett. 2016, 8, 29–32. [Google Scholar]
- Shenoy, R.S.; Prashanth, K.V.H.; Manonmani, H.K. In vitro antidiabetic effects of isolated triterpene glycoside fraction from Gymnema sylvestre. Evid.-Based Complement. Alternat. Med. 2018, 7154702. [Google Scholar] [CrossRef] [PubMed]
- Kalita, D.; Holm, D.G.; LaBarbera, D.V.; Petrash, J.M.; Jayanty, S.S. Inhibition of α-glucosidase, α-amylase, and aldose reductase by potato polyphenolic compounds. PLoS ONE 2018, 13, e0191025. [Google Scholar] [CrossRef] [PubMed]
- Ratnasari, N.; Walters, M.; Tsopmo, A. Antioxidant and lipoxygenase activities of polyphenol extracts from oat brans treated with polysaccharide degrading enzymes. Heliyon 2017, 3, e00351. [Google Scholar] [CrossRef] [Green Version]
- Umapathy, E.; Ndebia, E.J.; Meeme, A.; Adam, B.; Menziwa, P.; Nkeh-Chungag, B.N.; Iputo, J.E. An experimental evaluation of Albuca setosa aqueous extract on membrane stabilization, protein denaturation and white blood cell migration during acute inflammation. J. Med. Plants Res. 2010, 4, 789–795. [Google Scholar]
- David, B.C.; Sudarsanam, G. Antimicrobial activity of Gymnema sylvestre (Asclepiadaceae). J. Acute Dis. 2013, 222–225. [Google Scholar] [CrossRef]
- Tusevski, B.; Vinterhalter, D.; Krstic Milosevic, M.; Sokovic, A.; Ciric, D.; Vinterhalter, S.; Zdravkovic Korac, J.; Petreska Stanoeva, M.; Stefova, S.; Simic, G. Production of phenolic compounds, antioxidant and antimicrobial activities in hairy root and shoot cultures of Hypericum perforatum L. Plant Cell Tiss. Org. Cult. 2017, 128, 589–605. [Google Scholar] [CrossRef]
- Saumendu, D.R.; Sarkar, K.; Dipankar, S.; Singh, T.; Prabha, B. In vitro antibiotic activity of various extracts of Gymnema sylvestre. Int. J. Pharm. Res. Dev. 2010, 2, 1–3. [Google Scholar]
- Ghanati, F.; Bakhtiarian, S.; Parast, B.M.; Behrooz, M.K. Production of new active phytocompounds by Achillea millefolium L. after elicitation with silver nanoparticles and methyl jasmonate. Biosci. Biotechnol. Res. Asia 2014, 11, 391–399. [Google Scholar] [CrossRef]
- Khanna, V.G.; Kannabiran, K. Anticancer-cytotoxic activity of saponins isolated from the leaves of Gymnema sylvestre and Eclipta prostrata on HeLa cells. Int. J. Green Pharm. 2009, 3, 227–229. [Google Scholar]
- Srikanth, A.V.; Maricar, S.; Lakshmi, M.N.; Ravi Kumar, P.; Madhava Reddy, B. Anticancer activity of Gymnema Sylv. R. Br. Int. J. Pharm. Sci. Nanotechnol. 2010, 3, 2–4. [Google Scholar]







© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chung, I.-M.; Rajakumar, G.; Subramanian, U.; Venkidasamy, B.; Thiruvengadam, M. Impact of Copper Oxide Nanoparticles on Enhancement of Bioactive Compounds Using Cell Suspension Cultures of Gymnema sylvestre (Retz.) R. Br. Appl. Sci. 2019, 9, 2165. https://doi.org/10.3390/app9102165
Chung I-M, Rajakumar G, Subramanian U, Venkidasamy B, Thiruvengadam M. Impact of Copper Oxide Nanoparticles on Enhancement of Bioactive Compounds Using Cell Suspension Cultures of Gymnema sylvestre (Retz.) R. Br. Applied Sciences. 2019; 9(10):2165. https://doi.org/10.3390/app9102165
Chicago/Turabian StyleChung, Ill-Min, Govindasamy Rajakumar, Umadevi Subramanian, Baskar Venkidasamy, and Muthu Thiruvengadam. 2019. "Impact of Copper Oxide Nanoparticles on Enhancement of Bioactive Compounds Using Cell Suspension Cultures of Gymnema sylvestre (Retz.) R. Br" Applied Sciences 9, no. 10: 2165. https://doi.org/10.3390/app9102165
APA StyleChung, I. -M., Rajakumar, G., Subramanian, U., Venkidasamy, B., & Thiruvengadam, M. (2019). Impact of Copper Oxide Nanoparticles on Enhancement of Bioactive Compounds Using Cell Suspension Cultures of Gymnema sylvestre (Retz.) R. Br. Applied Sciences, 9(10), 2165. https://doi.org/10.3390/app9102165