Use of Anionic Polysaccharides in the Development of 3D Bioprinting Technology
 , , and
, , and 
Abstract
:1. Introduction
2. Overview of Natural Anionic Polysaccharides from Plant, Bacteria and Algae
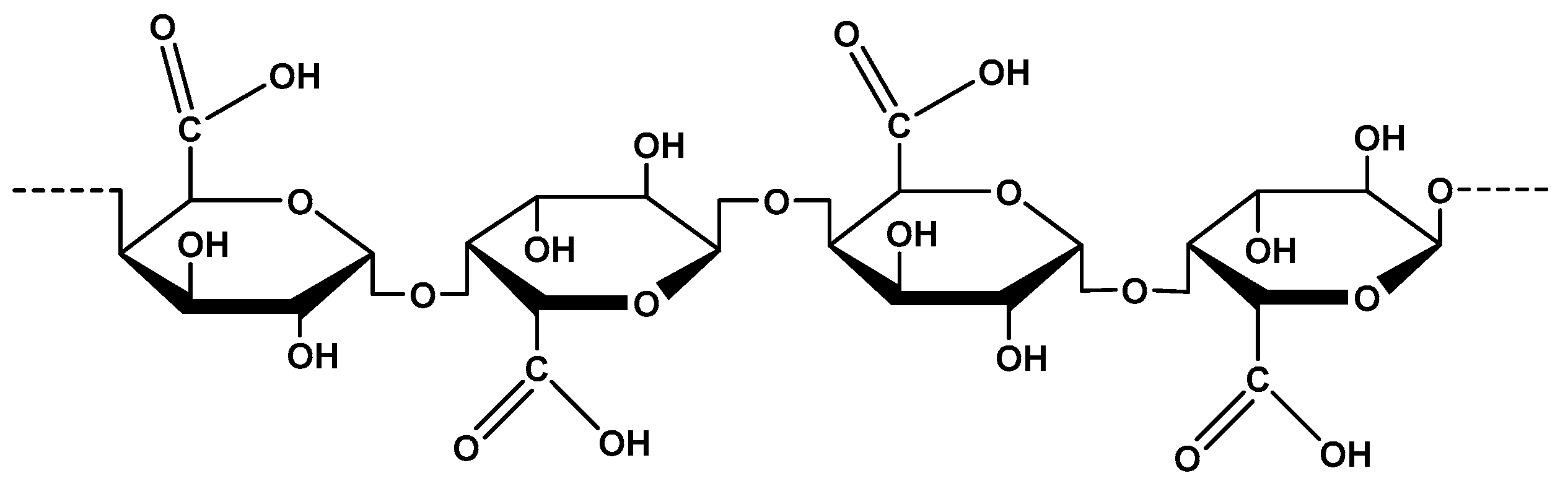
2.1. Polygalacturonic Acid from Pectin
2.2. Polyglucuronic Acid from Bacteria, Fungi and Green Algae
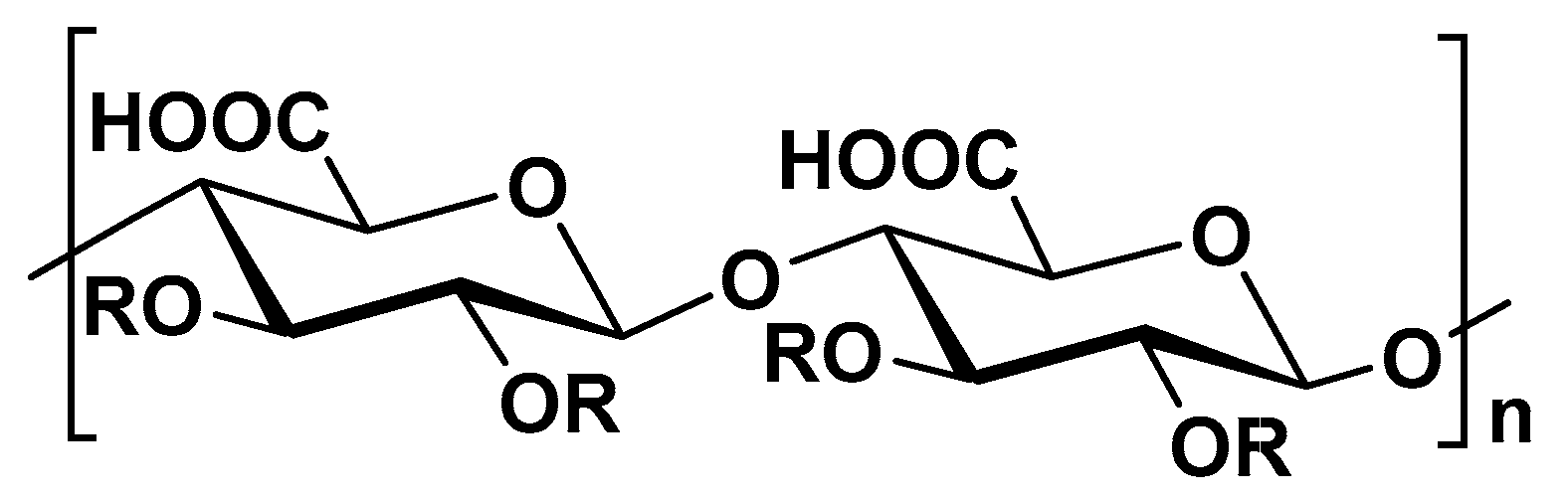
2.3. Alginate from Brown Algae and Bacteria
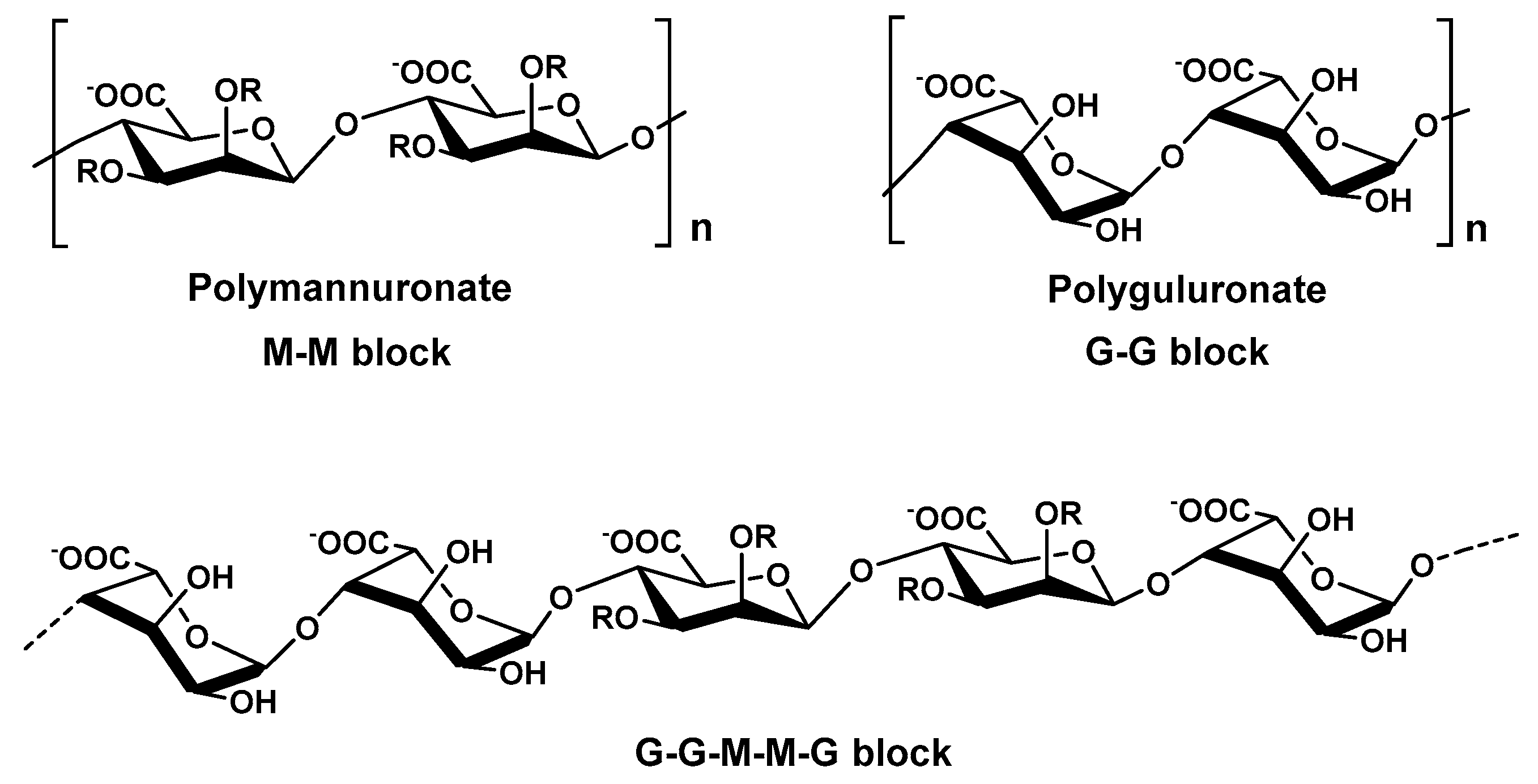
2.3.1. Alginate from Brown Algae
2.3.2. Alginate from Azotobacter Vinelandii and Pseudomonas spp.
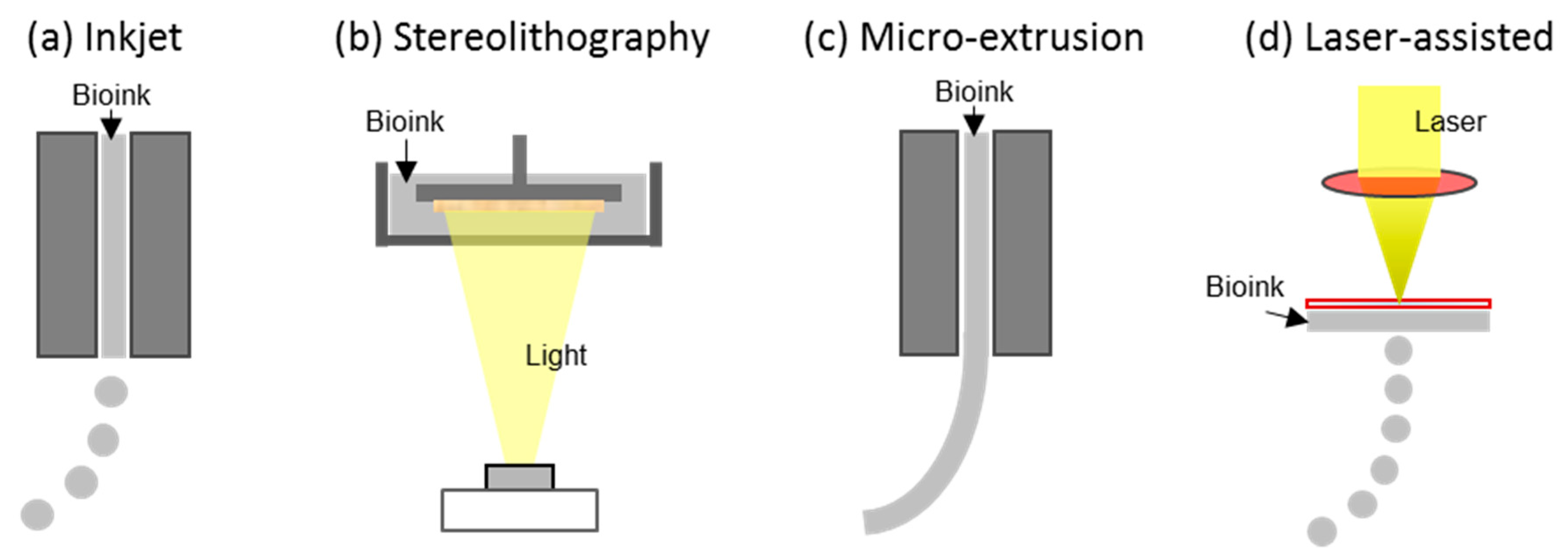
3. 3D Bioprinting Technology Concept
4. Biomaterial Application of Alginate in 3D Bioprinting and Perspectives Using Other Polyuronic Acid Polysaccharides
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Kodama, H. Automatic method for fabricating a three-dimensional plastic model with photo-hardening polymer. Rev. Sci. Instrum. 1981, 52, 1770–1773. [Google Scholar] [CrossRef]
- Crump, S.S. Apparatus and Method for Creating Three-Dimensional Objects. U.S. Patent No. 5,121,329, 9 June 1992. [Google Scholar]
- Liu, J.; Sun, L.; Xu, W.; Wang, Q.; Yu, S.; Sun, J. Current advances and future perspectives of 3D printing natural-derived biopolymers. Carbohyd. Polym. 2019, 207, 297–316. [Google Scholar] [CrossRef] [PubMed]
- Yanagawa, F.; Sugiura, S.; Kanamori, T. Hydrogel microfabrication technology toward three dimensional tissue engineering. Regen. Ther. 2016, 3, 45–57. [Google Scholar] [CrossRef] [Green Version]
- Hong, N.; Yang, G.H.; Lee, J.; Kim, G. 3D bioprinting and its in vivo applications. J. Biomed. Mater. Res. B Appl. Biomater. 2018, 106, 444–459. [Google Scholar] [CrossRef]
- Sutherland, I.W. Bacterial exopolysaccharides. In Comprehensive Glycoscience: From Chemistry to System Biology; Boons, G.-J., Kamerling, J.P., Lee, Y.C., Suzuki, A., Taniguchi, N., Voragen, A.G.J., Eds.; Elsevier B.V.: Oxford, UK, 2007; Volume 2, pp. 521–557. [Google Scholar]
- Laroche, C.; Michaud, P. New developments and prospective applications for (1,3) glucans. Rec. Pat. Biotechnol. 2007, 1, 59–73. [Google Scholar] [CrossRef]
- Delattre, C.; Pierre, G.; Laroche, C.; Michaud, P. Production, extraction and characterization of microalgal and cyanobacterial exopolysaccharides. Biotechnol. Adv. 2016, 34, 1159–1179. [Google Scholar] [CrossRef] [PubMed]
- Pierre, G.; Delattre, C.; Laroche, C.; Michaud, P. Galactans and its applications. In Polysaccharides; Springer International Publishing: Cham, Switzerland, 2014. [Google Scholar] [CrossRef]
- Giustina, G.D.; Gandin, A.; Brigo, L.; Panciera, T.; Giulitti, S.; Sgarbossa, P.; D’Alessandro, D.; Trombi, L.; Danti, S.; Brusatin, G. Polysaccharide hydrogels for multiscale 3D printing of pullulan scaffolds. Mater. Des. 2019, 165, 107566. [Google Scholar] [CrossRef]
- Mohan, T.; Maver, T.; Štiglic, A.D.; Stana-Kleinschek, K.; Kargl, R. 3D bioprinting of polysaccharides and their derivatives: From characterization to application. In Fundamental Biomaterials: Polymers; Elsevier Ltd.: Amsterdam, The Netherlands, 2018. [Google Scholar] [CrossRef]
- Murphy, S.V.; Atala, A. 3D bioprinting of tissues and organs. Nat. Biotechnol. 2014, 32, 773–785. [Google Scholar] [CrossRef]
- Ab-Rahim, S.; Selvaratnam, L.; Raghavendran, H.R.B.; Kamarul, T. Chondrocyte-alginate constructs with or without TGF-1 produces superior extracellular matrix expression than monolayer cultures. Mol. Cell. Biochem. 2013, 376, 11–20. [Google Scholar] [CrossRef]
- Elboutachfaiti, R.; Delattre, C.; Petit, E.; Michaud, P. Polyglucuronics acids: Structures, functions and degrading enzymes. Carbohydr. Polym. 2011, 84, 1–13. [Google Scholar] [CrossRef]
- Purcell, E.K.; Singh, A.; Kipke, D.R. Alginate composition effects on a neural stem cell-seeded scaffold. Tissue Eng. Part C Methods 2009, 15, 541–550. [Google Scholar] [CrossRef]
- Luo, S.J.; Chen, R.Y.; Huang, L.; Liang, R.H.; Liu, C.-M.; Chen, J. Investigation on the influence of pectin structures on the pasting properties of rice starch by multiple regression. Food Hydrocoll. 2017, 63, 580–584. [Google Scholar] [CrossRef]
- Pérez, S.; Rodríguez-Carvajal, M.A.; Doco, T. A complex plant cell wall polysaccharide: Rhamnogalacturonan II.A structure in quest of a function. Biochimie 2003, 85, 109–121. [Google Scholar] [CrossRef]
- Taylor, K.A.; Buchanan-Smith, J.G. A colorimetric method for the quantitation of uronic acids and a specific assay for galacturonic acid. Anal. Biochem. 1992, 201, 190–196. [Google Scholar] [CrossRef]
- Aspinall, G.O. Chemistry of cell wall polysaccharides. In The Biochemistry of Plants; Preiss, J., Ed.; Academic Press: New York, NY, USA, 1980; pp. 473–500. [Google Scholar]
- Voragen, A.G.J.; Pilnik, W.; Thibault, J.F.; Axelos, M.A.V.; Renard, C.M.G.C. Pectins. In Food Polysaccharides and Their Applications; Stephen, A.M., Ed.; Marcel Dekker: New York, NY, USA, 1995; pp. 287–339. [Google Scholar]
- Axelos, M.A.V.; Thibault, J.F. Influence of the substituents of the carboxyl groups and the rhamnose content on the solution properties and flexibility of pectins. Int. J. Biol. Macromol. 1991, 13, 77–82. [Google Scholar] [CrossRef]
- Renard, C.M.G.C.; Jarvis, M.C. Acetylation and methylation of homogalacturonans. Part II. Effect on ion-binding properties and conformations. Carbohydr. Polym. 1999, 39, 209–216. [Google Scholar] [CrossRef]
- Massiot, P.; Rouau, X.; Thibault, J.F. characterisation of the extractable pectins and hemicelluloses of the cell wall of carrot. Carbohydr. Res. 1988, 172, 229–242. [Google Scholar] [CrossRef]
- Guillon, F.; Thibault, J.F. Further characterization of acid- and alkali-soluble pectins from sugar beet pulp. LMT-Food Sci. Technol. 1988, 21, 198–205. [Google Scholar]
- Saulnier, L.; Thibault, J.F. Extraction and characterization of pectic substances from pulp of grape berries. Carbohydr. Polym. 1987, 7, 329–343. [Google Scholar] [CrossRef] [Green Version]
- Renard, C.M.G.C.; Weightman, R.M.; Thibault, J.F. Structure of the xylose-rich pectins from pea hulls. Int. J. Biol. Macromol. 1997, 21, 155–162. [Google Scholar] [CrossRef]
- Renard, C.M.G.C.; Champenois, Y.; Thibault, J.F. Characterization of the extractable pectins and hemicelluloses of the cell wall of glasswort, Salicornia ramosissima. Carbohydr. Polym. 1993, 22, 239–245. [Google Scholar] [CrossRef]
- Blumenkrantz, N.; Asboe-Hansen, G. New method for quantitative determination of uronic acids. Anal. Biochem. 1973, 54, 484–489. [Google Scholar] [CrossRef]
- Scott, R.W. Colorimetric determination of hexuronic acids in plant materials. Anal. Chem. 1979, 51, 936–941. [Google Scholar] [CrossRef]
- Thibault, J.F. Automatisation du dosage des substances pectiques par la méthode au métahydroxydiphényle. LMT-Food Sci. Technol. 1979, 12, 247–251. [Google Scholar]
- Heyraud, A.; Courtois, J.; Dantas, L.; Colin-Morel, P.; Courtois, B. Structural characterization and rheological properties of an extracellular glucuronan produced by a Rhizobium meliloti M5N1 mutant strain. Carbohydr. Res. 1993, 240, 71–78. [Google Scholar] [CrossRef]
- Dantas, L.; Courtois, J.; Courtois, B.; Séguin, J.P.; Gey, C.; Heyraud, A. NMR spectroscopic investigation of oligoglucuronates prepared by enzymic hydrolysis of a (1,4) beta-d-glucuronan. Carbohydr. Res. 1994, 265, 303–310. [Google Scholar] [CrossRef]
- Courtois-Sambourg, J.; Courtois, B.; Heyraud, A.; Colin-Morel, P.; Rinaudo-Duhem, M. Polymer Compounds of the Glycuronic Acid, Method of Preparation and Utilization Particularly as Gelifying, Thickening, Hydrating, Stabilizing, Chelating or Floculating Means. Patent WO1,993,018,174, 16 September 1993. [Google Scholar]
- Tavernier, M.L.; Petit, E.; Delattre, C.; Courtois, B.; Courtois, J.; Strancar, A. Production of oligoglucuronans using a monolithic enzymatic microreactor. Carbohydr. Res. 2008, 343, 2687–2691. [Google Scholar] [CrossRef] [PubMed]
- Park, J.K.; Park, Y.H.; Jung, J.Y. Production of bacterial cellulose by Gluconacetobacter hansenii PJK isolated from rotten apple. Biotechnol. Biop. Eng. 2003, 8, 83–88. [Google Scholar] [CrossRef]
- Jung, J.Y.; Park, J.K.; Chang, H.N. Bacterial cellulose production by Gluconacetobacter hansenii in an agitated culture without living non cellulose producing cells. Enzym. Microb. Technol. 2005, 37, 347–354. [Google Scholar] [CrossRef]
- Park, J.K.; Hyun, S.H.; Jung, J.Y. Conversion of G. hansenii PJK into noncellulose-producing mutants. Biotechnol. Bioprocess Eng. 2004, 9, 383–388. [Google Scholar] [CrossRef]
- Shah, N.; Ha, J.H.; Park, J.K. Effect of reactor surface on production of bacterial cellulose and water-soluble oligosaccharides by Gluconacetobacter hansenii PJK. Biotechnol. Bioprocess Eng. 2010, 15, 110–118. [Google Scholar] [CrossRef]
- Khan, T.; Hyun, S.H.; Park, J.K. Physical properties of a single sugar-linked glucuronic acid-based oligosaccharides produced by a Gluconacetobacter hansenii strain. Process Biochem. 2007, 42, 252–257. [Google Scholar] [CrossRef]
- Park, J.K.; Khan, T.; Jung, J.Y. Structural studies of the glucuronic acid oligomers produced by Gluconacetobacter hansenii strain. Carbohydr. Polym. 2006, 63, 482–486. [Google Scholar] [CrossRef]
- Khan, T.; Park, J.K. The structure and physical properties of glucuronic acid oligomers produced by a Gluconacetobacter hansenii strain using the waste from beer fermentation broth. Carbohydr. Polym. 2008, 73, 438–445. [Google Scholar] [CrossRef]
- Bartnicki-Garcia, S.; Nickerson, W.J. Isolation, composition, and structure of cell walls of filaments and yeast-like forms of Mucor rouxii. Biochim. Biophys. Acta 1962, 58, 102–119. [Google Scholar] [CrossRef]
- Bartnicki-Garcia, S.; Reyes, E. Polyuronides in the cell wall of Mucor rouxii. Biochim. Biophys. Acta 1968, 170, 54–62. [Google Scholar] [CrossRef]
- Dow, J.M.; Darnall, D.W.; Villa, V.D. Two distinct classes of polyuronide from the cell walls of a dimorphic fungus, Mucor rouxii. J. Bacteriol. 1983, 155, 1088–1093. [Google Scholar]
- Ray, B. Polysaccharides from Enteromorpha compressa: Isolation, purification and structural features. Carbohydr. Polym. 2006, 66, 408–416. [Google Scholar] [CrossRef]
- Ray, B.; Lahaye, M. Cell-wall polysaccharides from the marine green alga Ulva “rigida” (Ulvales, Chlorophyta). Chemical structure of ulvan. Carbohydr Res. 1995, 274, 313–318. [Google Scholar] [CrossRef]
- Hentati, F.; Delattre, C.; Ursu, A.V.; Desbrières, J.; Le Cerf, D.; Gardarin, C.; Abdelkafi, S.; Michaudn, P.; Pierre, G. Structural characterization and antioxidant activity of water-soluble polysaccharides from the Tunisian brown seaweed Cystoseira compressa. Carbohydr. Polym. 2018, 198, 589–600. [Google Scholar] [CrossRef]
- Gacesa, P. Alginates. Carbohydr. Polym. 1988, 8, 161–182. [Google Scholar] [CrossRef]
- Draget, K.I.; Smidsrød, O.; Skjak-Braek, G. Alginates from Algae. In Biopolymers: Polysaccharides from Eukaryotes; De Baets, S., Vandamme, E.J., Steinbüchel, A., Eds.; Wileg-VCH: Weinheim, Germany, 2002; pp. 215–244. [Google Scholar]
- Smidsrod, O.; Draget, K.I. Alginates: Chemistry and physical properties. Carbohydr. Eur. 1996, 14, 6–13. [Google Scholar]
- Rinaudo, M. Seaweed polysaccharides. In Comprehensive Glycoscience: From Chemistry to Systems Biology; Kamerling, J.P., Ed.; Elsevier Science: Amsterdam, The Netherland, 2007; Volume 2, pp. 691–735. [Google Scholar]
- Cosenza, V.A.; Navarro, D.A.; Ponce, N.M.A. Seaweed Polysaccharides: Structure and Applications. In Industrial Applications of Renewable Biomass Products; Goyanes, S., D’Accorso, N., Eds.; Springer: Cham, Switzerland, 2017. [Google Scholar]
- Laurienzo, P. Marine polysaccharides in pharmaceutical applications: An overview. Mar. Drugs 2010, 8, 2435–2465. [Google Scholar] [CrossRef] [PubMed]
- Ertesvåg, H.; Valla, S. Biosynthesis and applications of alginates. Polym. Degr. Stab. 1998, 59, 85–91. [Google Scholar] [CrossRef]
- Lim, S.J.; Aida, W.M.W. Chapter 3—Extraction of Sulfated Polysaccharides (Fucoidan) from Brown Seaweed. In Seaweed Polysaccharides: Isolation, Biological and Biomedical Applications; Elsevier: Cham, Switzerland, 2017; pp. 27–46. [Google Scholar]
- Galindo, E.; Peña, C.; Núñez, C.; Segura, D.; Espin, G. Molecular and bioengineering strategies to improve alginate and polyhydroxyalkanoate production by Azotobacter vinelandii. Microb. Cell Fact. 2007, 6, 1–16. [Google Scholar] [CrossRef]
- Rehm, B.H. Bacterial polymers: Biosynthesis, modifications and applications. Nat. Rev. Microbiol. 2010, 8, 578–592. [Google Scholar] [CrossRef] [PubMed]
- Gacesa, P. Bacterial alginate biosynthesis-recent progress and future prospects. J. Microbiol. 1998, 144, 1133–1143. [Google Scholar] [CrossRef]
- Hay, I.D.; Wang, Y.; Moradali, M.F.; Rehman, Z.U.; Rehm, B.H. Genetics and regulation of bacterial alginate production. Appl. Environ. Microbiol. 2014, 16, 2997–3011. [Google Scholar] [CrossRef]
- Remminghorst, U.; Rehm, B.H.A. In vitro alginate polymerization and the functional role of Alg8 in alginate production by Pseudomonas aeruginosa. Appl. Environ. Mirobiol. 2006, 72, 298–305. [Google Scholar] [CrossRef]
- Sabra, W.; Zeng, A.P.; Lünsdorf, H.; Deckwer, W.D. Effect of oxygen on formation and structure of Azotobacter vinelandii and its role in protecting nitrogenase. Appl. Environ. Microbiol. 2000, 66, 4037–4044. [Google Scholar] [CrossRef]
- Oelze, J. Respiratory protection of nitrogenase in Azotobacter species: Is a widely held hypothesis unequivocally supported by experimental evidence. FEMS Microbiol. Rev. 2000, 24, 321–333. [Google Scholar] [CrossRef] [PubMed]
- Ramphal, R.; Pier, G.B. Role of Pseudomonas aeruginosa mucoid exopolysaccharide in adherence to tracheal cell. Infect. Inmmun. 1985, 47, 1–4. [Google Scholar]
- Franklin, M.J.; Nivens, D.E.; Weadge, J.T.; Lynne Howell, P. Biosynthesis of the Pseudomonas aeruginosa extracellular polysaccharides, alginate, Pel and Psl. Front. Microbiol. 2011, 2, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Skajak-Braek, G.; Larsen, B.; Grasdalen, H. Monomer sequence and acetylation pattern in some bacterial alginates. Carbohydr. Res. 1986, 154, 239–250. [Google Scholar] [CrossRef]
- Holzl, K.; Lin, S.M.; Tytgat, L.; Van Vlierberghe, S.; Gu, L.X.; Ovsianikov, A. Bioink properties before, during and after 3D bioprinting. Biofabrication 2016, 8, 032002. [Google Scholar] [CrossRef] [PubMed]
- Gudapati, H.; Dey, M.; Ozbolat, I. A comprehensive review on droplet-based bioprinting: Past, present and future. Biomaterials 2016, 102, 20–42. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pedde, R.D.; Mirani, B.; Navaei, A.; Styan, T.; Wong, S.; Mehrali, M.; Thakur, A.; Mohtaram, N.K.; Bayati, A.; Dolatshahi-Pirouz, A.; et al. Emerging biofabrication strategies for engineering complex tissue constructs. Adv. Mater. 2017, 29, 1606061. [Google Scholar] [CrossRef] [PubMed]
- Hopp, B.; Smausz, T.; Szabo, G.; Kolozsvari, L.; Kafetzopoulos, D.; Fotakis, C.; Ngrdi, A. Femtosecond laser printing of living cells using absorbing film-assisted laser-induced forward transfer. Opt. Eng. 2012, 51, 014302. [Google Scholar] [CrossRef]
- Lee, K.Y.; Mooney, D.J. Alginate: Properties and biomedical applications. Prog. Polym. Sci. 2012, 37, 106–126. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nakamura, M.; Kobayashi, A.; Takagi, F.; Watanabe, A.; Hiruma, Y.; Ohuchi, K.; Iwasaki, Y.; Horie, M.; Morita, I.; Takatani, S. Biocompatible inkjet printing technique for designed seeding of individual living cells. Tissue Eng. 2005, 11, 1658–1666. [Google Scholar] [CrossRef]
- Nakamura, M.; Nishiyama, Y.; Henmi, C.; Iwanaga, S.; Nakagawa, H.; Yamaguchi, K.; Akita, K.; Mochizuki, S.; Takiura, K. Ink jet three-dimensional digital fabrication for biological tissue manufacturing: Analysis of alginate microgel beads produced by ink jet droplets for three dimensional tissue fabrication. J. Imaging Sci. Technol. 2008, 52, 060201–060206. [Google Scholar] [CrossRef]
- Nishiyama, Y.; Nakamura, M.; Henmi, C.; Yamaguchi, K.; Mochizuki, S.; Nakagawa, H.; Takiura, K. Development of a three-dimensional bioprinter: Construction of cell supporting structures using hydrogel and state-of-the-art inkjet technology. J. Biomech. Eng. 2009, 131, 035001. [Google Scholar] [CrossRef] [PubMed]
- Jia, J.; Richards, D.J.; Pollard, S.; Tan, Y.; Rodriguez, J.; Visconti, R.P.; Trusk, T.C.; Yost, M.J.; Yao, H.; Markwald, R.R.; et al. Engineering alginate as bioink for bioprinting. Acta Biomater. 2014, 10, 4323–4331. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Q.; Xia, Q.; Wu, Y.; Zhang, X.; Wen, F.; Chen, X.; Zhang, S.; Heng, B.C.; He, Y.; Ouyang, H.W. 3D-printed Atsttrin-incorporated alginate/hydroxyapatite scaffold promotes bone defect regeneration with TNF/TNFR signaling involvement. Adv. Healthc. Mater. 2015, 4, 1701–1708. [Google Scholar] [CrossRef] [PubMed]
- Wust, S.; Godla, M.E.; Muller, R.; Hofmann, S. Tunable hydrogel composite with two-step processing in combination with innovative hardware upgrade for cell-based three-dimensional bioprinting. Acta Biomater. 2014, 10, 630–640. [Google Scholar] [CrossRef] [PubMed]
- Sun, J.; Wei, D.; Yang, K.; Yang, Y.; Liu, X.L.; Fan, H.S.; Zhang, X.D. The development of cell-initiated degradable hydrogel based on methacrylated alginate applicable to multiple microfabrication technologies. J. Mater. Chem. B. 2017, 5, 8060–8069. [Google Scholar] [CrossRef]
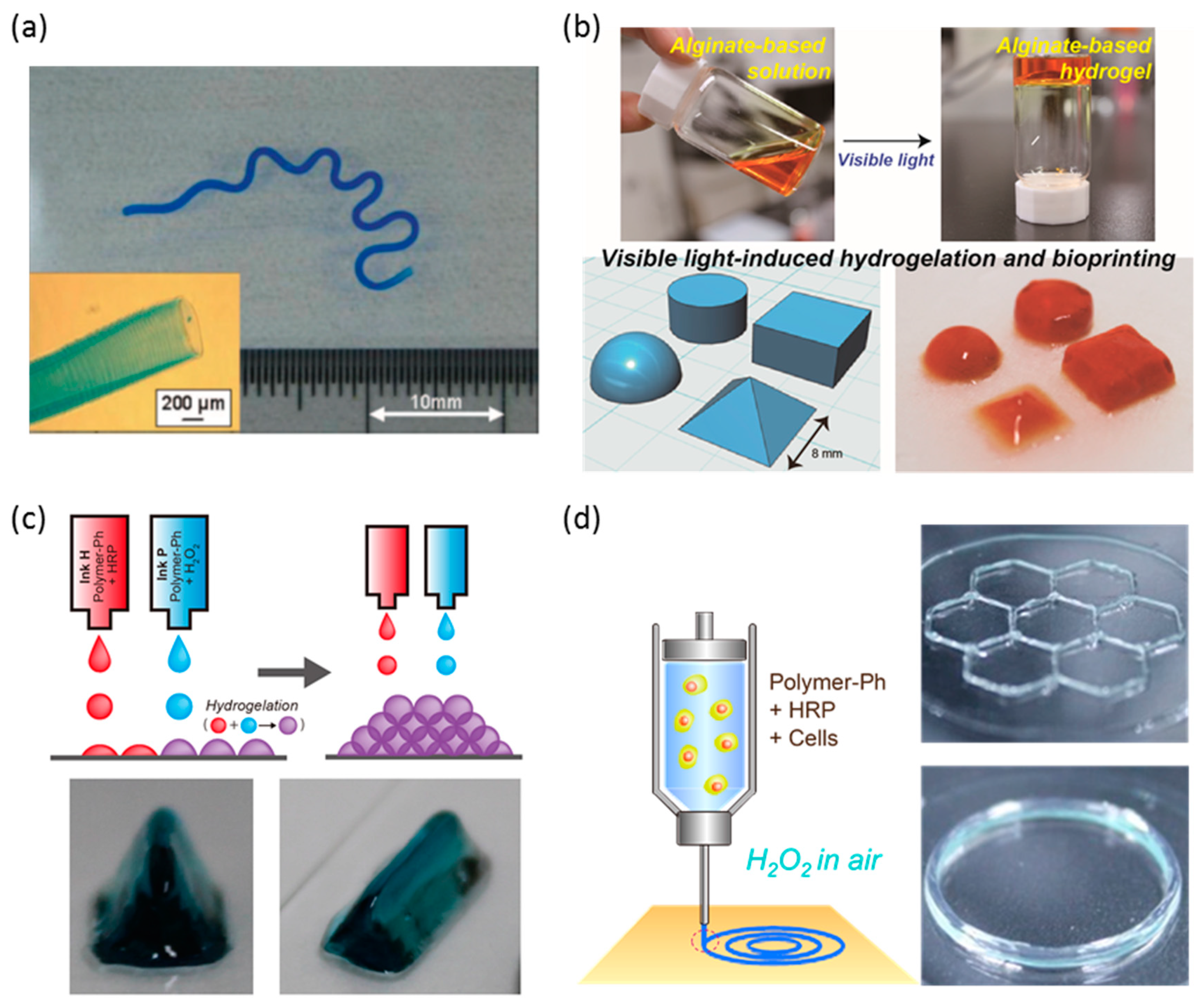
- Sakai, S.; Kamei, H.; Mori, T.; Hotta, T.; Ohi, H.; Nakahata, M.; Taya, M. Visible light-induced hydrogelation of an alginate derivative and application to stereolithographic bioprinting using a visible light projector and Acid Red. Biomacromolecules 2018, 19, 672–679. [Google Scholar] [CrossRef]
- Sakai, S.; Ueda, K.; Gantumur, E.; Taya, M.; Nakamura, M. Drop-On-Drop multimaterial 3D bioprinting realized by peroxidase-mediated cross-linking. Macromol. Rapid Commun. 2018, 39, 1700534. [Google Scholar] [CrossRef]
- Sakai, S.; Mochizuki, K.; Qu, Y.; Mail, M.; Nakahata, M.; Taya, M. Peroxidase-catalyzed microextrusion bioprinting of cell-laden hydrogel constructs in vaporized ppm-level hydrogen peroxide. Biofabrication 2018, 10, 045007. [Google Scholar] [CrossRef]
- Casale, M.; Moffa, A.; Vella, P.; Sabatino, L.; Capuano, F.; Salvinelli, B.; Lopez, M.A.; Carinci, F.; Salvinelli, F. Hyaluronic acid: Perspectives in dentistry. A systematic review. Int. J. Immunopathol. Pharmacol. 2016, 29, 572–582. [Google Scholar] [CrossRef]
- Pre, E.D.; Conti, G.; Sbarbati, A. Hyaluronic acid (HA) scaffolds and multipotent stromal cells (MSCs) in regenerative medicine. Stem Cell Rev. Rep. 2016, 12, 664–681. [Google Scholar] [CrossRef] [PubMed]
- Poldervaart, M.T.; Goversen, B.; de Ruijter, M.; Abbadessa, A.; Melchels, F.P.W.; Oner, F.C.; Dhert, W.J.A.; Vermonden, T.; Alblas, J. 3D bioprinting of methacrylated hyaluronic acid (MeHA) hydrogel with intrinsic osteogenicity. PLoS ONE 2017, 12, e0177628. [Google Scholar] [CrossRef] [PubMed]
- Sakai, S.; Ohi, H.; Hotta, T.; Kamei, H.; Taya, M. Differentiation potential of human adipose stem cells bioprinted with hyaluronic acid/gelatin-based bioink through microextrusion and visible light-initiated crosslinking. Biopolymers 2018, 109, e23080. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Origin | GalA | Rha | Ara | Xyl | Gal | FerA | Reference |
|---|---|---|---|---|---|---|---|
| Citrus | 792 | 14 | 11 | 2 | 24 | ND* | [21] |
| Apple | 731 | 23 | 44 | 17 | 42 | ND | [21] |
| Quinoa | 625 | 38 | 33 | 18 | 51 | 3 | [22] |
| Carrot | 613 | 43 | 35 | 3 | 79 | ND | [23] |
| Beet | 558 | 28 | 124 | 3 | 47 | 7 | [24] |
| Grape | 533 | 58 | 77 | 6 | 44 | ND | [25] |
| Pea | 449 | 102 | 144 | 155 | 75 | ND | [26] |
| Pear | 380 | ND | ND | ND | ND | ND | [20] |
| Potato | 350 | ND | ND | ND | ND | ND | [20] |
| Glasswort | 289 | 22 | 462 | 3 | 43 | 17 | [27] |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tai, C.; Bouissil, S.; Gantumur, E.; Carranza, M.S.; Yoshii, A.; Sakai, S.; Pierre, G.; Michaud, P.; Delattre, C. Use of Anionic Polysaccharides in the Development of 3D Bioprinting Technology. Appl. Sci. 2019, 9, 2596. https://doi.org/10.3390/app9132596
Tai C, Bouissil S, Gantumur E, Carranza MS, Yoshii A, Sakai S, Pierre G, Michaud P, Delattre C. Use of Anionic Polysaccharides in the Development of 3D Bioprinting Technology. Applied Sciences. 2019; 9(13):2596. https://doi.org/10.3390/app9132596
Chicago/Turabian StyleTai, Chia, Soukaina Bouissil, Enkhtuul Gantumur, Mary Stephanie Carranza, Ayano Yoshii, Shinji Sakai, Guillaume Pierre, Philippe Michaud, and Cédric Delattre. 2019. "Use of Anionic Polysaccharides in the Development of 3D Bioprinting Technology" Applied Sciences 9, no. 13: 2596. https://doi.org/10.3390/app9132596
APA StyleTai, C., Bouissil, S., Gantumur, E., Carranza, M. S., Yoshii, A., Sakai, S., Pierre, G., Michaud, P., & Delattre, C. (2019). Use of Anionic Polysaccharides in the Development of 3D Bioprinting Technology. Applied Sciences, 9(13), 2596. https://doi.org/10.3390/app9132596