The Role of Movement Representation Techniques in the Motor Learning Process: A Neurophysiological Hypothesis and a Narrative Review

 ,
,  ,
,  and
and 
Abstract
:1. Introduction
2. Effectiveness of AO and MI in the Motor Learning Process: A Minireview
3. Hypothesis
3.1. Shared Neurophysiological Activity
3.2. Magnitude of Brain Activity
3.3. Influence of Variables Related to Movement Representation
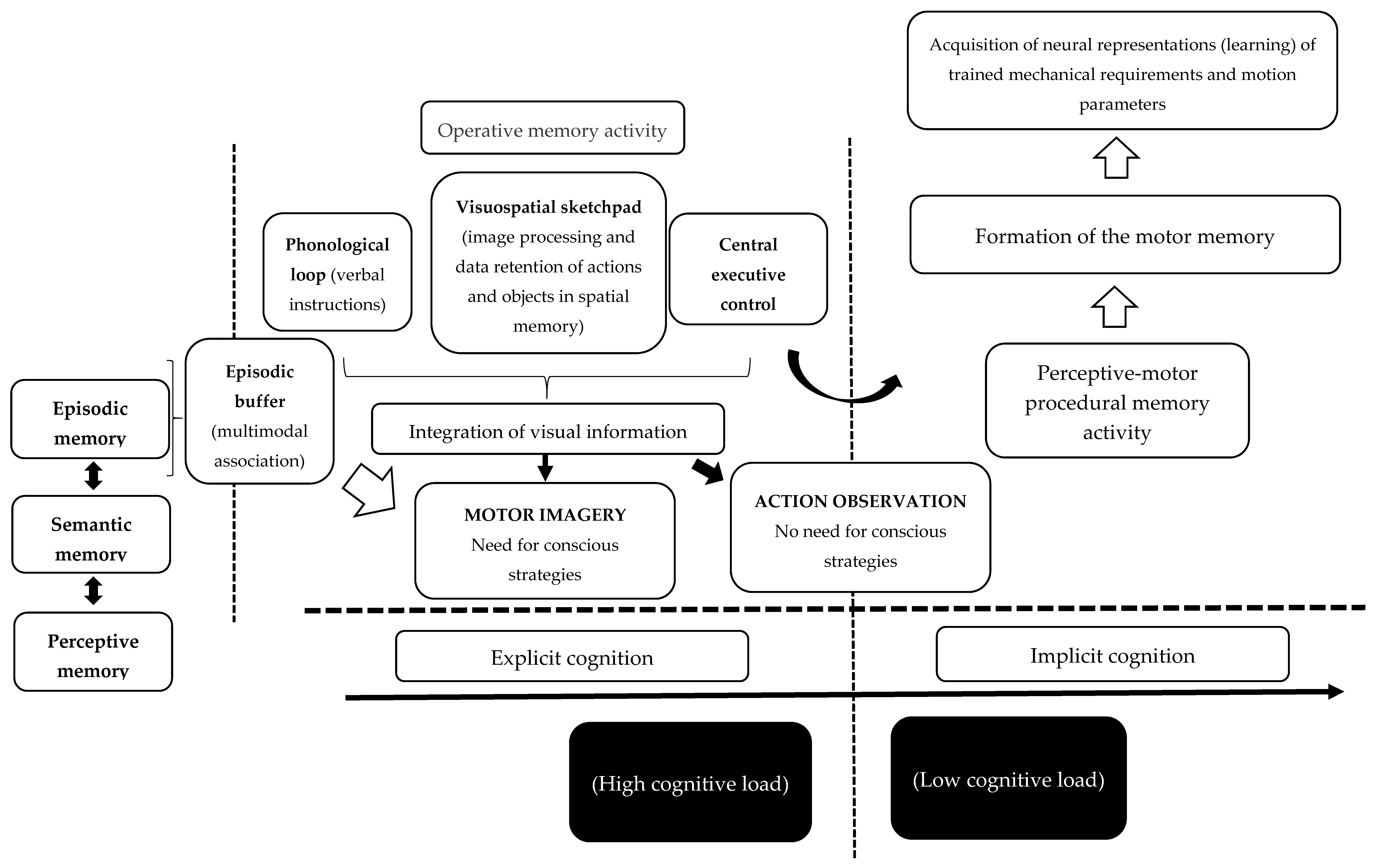
3.4. Differences in the Process of Creating Mnemonic Representations: Integration of Visual Information and Formation of Motor Memory
3.5. Observing and Imagining: Different Cognitive Demands
4. Theoretical Framework
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Isaac, A.R. Mental Practice—Does It Work in the Field? Sport Psychol. 1992, 6, 192–198. [Google Scholar] [CrossRef]
- Guillot, A.; Collet, C. Construction of the Motor Imagery Integrative Model in Sport: A review and theoretical investigation of motor imagery use. Int. Rev. Sport Exerc. Psychol. 2008, 1, 31–44. [Google Scholar] [CrossRef]
- Decety, J. The neurophysiological basis of motor imagery. Behav. Brain Res. 1996, 77, 45–52. [Google Scholar] [CrossRef]
- Buccino, G. Action observation treatment: A novel tool in neurorehabilitation. Philos. Trans. R. Soc. B Biol. Sci. 2014, 369, 20130185. [Google Scholar] [CrossRef]
- Dickstein, R.; Deutsch, J.E. Motor Imagery in Physical Therapist Practice. Phys. Ther. 2007, 87, 942–953. [Google Scholar] [CrossRef] [Green Version]
- Stefan, K.; Cohen, L.G.; Duque, J.; Mazzocchio, R.; Celnik, P.; Sawaki, L.; Ungerleider, L.; Classen, J. Formation of a Motor Memory by Action Observation. J. Neurosci. 2005, 25, 9339–9346. [Google Scholar] [CrossRef]
- Mouthon, A.; Ruffieux, J.; Waelchli, M.; Keller, M.; Taube, W. Task-Dependent Changes of Corticospinal Excitability During Observation and Motor Imagery of Balance Tasks. Neuroscience 2015, 303, 535–543. [Google Scholar] [CrossRef] [Green Version]
- Robin, N.; Dominique, L.; Toussaint, L.; Blandin, Y.; Guillot, A.; Her, M. The Effects of motor imagery training on service return accuracy in tennis: The role of imagery ability. Int. J. Sport Exerc. Psychol. 2007, 5, 175–186. [Google Scholar] [CrossRef]
- La Touche, R.; Sánchez-Vázquez, M.; Cuenca-Martínez, F.; Prieto-Aldana, M.; Paris-Alemany, A.; Navarro-Fernández, G. Instruction Modes for Motor Control Skills Acquisition: A Randomized Controlled Trial. J. Mot. Behav. 2019, 1–12. [Google Scholar] [CrossRef]
- Grush, R. The emulation theory of representation: Motor control, imagery, and perception. Behav. Brain Sci. 2004, 27, 377–396. [Google Scholar] [CrossRef] [Green Version]
- Glover, S.; Baran, M. The motor-cognitive model of motor imagery: Evidence from timing errors in simulated reaching and grasping. J. Exp. Psychol. Hum. Percept. Perform. 2017, 43, 1359–1375. [Google Scholar] [CrossRef] [PubMed]
- Fitts, P.M.; Posner, M.I. Human Performance; Brooks/Cole: Oxford, UK, 1967. [Google Scholar]
- Tinazzi, M.; Zanette, G. Modulation of ipsilateral motor cortex in man during unimanual finger movements of different complexities. Neurosci. Lett. 1998, 244, 121–124. [Google Scholar] [CrossRef]
- Cano-de-la-Cuerda, R.; Molero-Sánchez, A.; Carratalá-Tejada, M.; Alguacil-Diego, I.M.; Molina-Rueda, F.; Miangolarra-Page, J.C.; Torricelli, D. Teorías y modelos de control y aprendizaje motor. Aplicaciones clínicas en neurorrehabilitación. Neurologia 2015, 30, 32–41. [Google Scholar] [CrossRef] [PubMed]
- Bilodeau, E.A.; Bilodeau, I.M.; Schumsky, D.A. Some effects of introducing and withdrawing knowledge of results early and late in practice. J. Exp. Psychol. 1959, 58, 142–144. [Google Scholar] [CrossRef] [PubMed]
- Dahm, S.F.; Rieger, M. Is imagery better than reality? Performance in imagined dart throwing. Hum. Mov. Sci. 2019, 66, 38–52. [Google Scholar] [CrossRef]
- Stone, P.W. Popping the (PICO) question in research and evidence-based practice. Appl. Nurs. Res. 2002, 15, 197–198. [Google Scholar] [CrossRef]
- Maher, C.G.; Sherrington, C.; Herbert, R.D.; Moseley, A.M.; Elkins, M. Reliability of the PEDro Scale for Rating Quality of Randomized Controlled Trials. Phys. Ther. 2003, 83, 713–721. [Google Scholar] [CrossRef] [Green Version]
- Cohen, J. A Coefficient of Agreement for Nominal Scales. Educ. Psychol. Meas. 1960, 20, 37–46. [Google Scholar] [CrossRef]
- Cabral-Sequeira, A.S.; Coelho, D.B.; Teixeira, L.A. Motor imagery training promotes motor learning in adolescents with cerebral palsy: Comparison between left and right hemiparesis. Exp. Brain Res. 2016, 234, 1515–1524. [Google Scholar] [CrossRef]
- Kumar, V.K. Motor Imagery Training on Muscle Strength and Gait Performance in Ambulant Stroke Subjects-A Randomized Clinical Trial. J. Clin. Diagn. Res. 2016, 10, YC01–YC04. [Google Scholar]
- Kleynen, M.; Jie, L.J.; Theunissen, K.; Rasquin, S.M.C.; Masters, R.S.W.; Meijer, K.; Beurskens, A.J.; Braun, S.M. The immediate influence of implicit motor learning strategies on spatiotemporal gait parameters in stroke patients: A randomized within-subjects design. Clin. Rehabil. 2019, 33, 619–630. [Google Scholar] [CrossRef] [PubMed]
- Moukarzel, M.; Di Rienzo, F.; Lahoud, J.C.; Hoyek, F.; Collet, C.; Guillot, A.; Hoyek, N. The therapeutic role of motor imagery during the acute phase after total knee arthroplasty: A pilot study. Disabil. Rehabil. 2019, 41, 926–933. [Google Scholar] [CrossRef] [PubMed]
- Cuenca-Martínez, F.; Suso-Martí, L.; Sánchez-Martín, D.; Soria-Soria, C.; Serrano-Santos, J.; Paris-Alemany, A.; La Touche, R.; León-Hernández, J.V. Effects of Motor Imagery and Action Observation on Lumbo-pelvic Motor Control, Trunk Muscles Strength and Level of Perceived Fatigue: A Randomized Controlled Trial. Res. Q. Exerc. Sport 2019, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Bek, J.; Poliakoff, E.; Marshall, H.; Trueman, S.; Gowen, E. Enhancing voluntary imitation through attention and motor imagery. Exp. Brain Res. 2016, 234, 1819–1828. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sheahan, H.R.; Ingram, J.N.; Žalalytė, G.M.; Wolpert, D.M. Imagery of movements immediately following performance allows learning of motor skills that interfere. Sci. Rep. 2018, 8, 14330. [Google Scholar] [CrossRef] [Green Version]
- Dana, A.; Gozalzadeh, E. Internal and External Imagery Effects on Tennis Skills Among Novices. Percept. Mot. Skills 2017, 124, 1022–1043. [Google Scholar] [CrossRef]
- Kim, T.; Frank, C.; Schack, T. A systematic investigation of the effect of action observation training and motor imagery training on the development of mental representation structure and skill performance. Front. Hum. Neurosci. 2017, 11, 499. [Google Scholar] [CrossRef]
- Gonzalez-Rosa, J.J.; Natali, F.; Tettamanti, A.; Cursi, M.; Velikova, S.; Comi, G.; Gatti, R.; Leocani, L. Action observation and motor imagery in performance of complex movements: Evidence from EEG and kinematics analysis. Behav. Brain Res. 2015, 281, 290–300. [Google Scholar] [CrossRef] [Green Version]
- Hidalgo-Peréz, A.; Fernández-García, Á.; López-de-Uralde-Villanueva, I.; Gil-Martínez, A.; Paris-Alemany, A.; Fernández-Carnero, J.; La Touche, R. Effectiveness of a motor control therapeutic exercise program combined with motor imagery on the sensorimotor function of the cervical spine: A randomized controlled trial. Int. J. Sports Phys. Ther. 2015, 10, 877–892. [Google Scholar]
- Ingram, T.G.J.; Kraeutner, S.N.; Solomon, J.P.; Westwood, D.A.; Boe, S.G. Skill acquisition via motor imagery relies on both motor and perceptual learning. Behav. Neurosci. 2016, 130, 252–260. [Google Scholar] [CrossRef]
- Nishizawa, H.; Kimura, T. Enhancement of motor skill learning by a combination of ideal model-observation and self-observation. J. Phys. Ther. Sci. 2017, 29, 1555–1560. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kawasaki, T.; Tozawa, R.; Aramaki, H. Effectiveness of using an unskilled model in action observation combined with motor imagery training for early motor learning in elderly people: A preliminary study. Somatosens. Mot. Res. 2018, 35, 204–211. [Google Scholar] [CrossRef] [PubMed]
- Kraeutner, S.N.; MacKenzie, L.A.; Westwood, D.A.; Boe, S.G. Characterizing skill acquisition through motor imagery with no prior physical practice. J. Exp. Psychol. Hum. Percept. Perform. 2016, 42, 257–265. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kraeutner, S.N.; Gaughan, T.C.; Eppler, S.N.; Boe, S.G. Motor imagery-based implicit sequence learning depends on the formation of stimulus-response associations. Acta Psychol. 2017, 178, 48–55. [Google Scholar] [CrossRef]
- Lagravinese, G.; Bisio, A.; Ruggeri, P.; Bove, M.; Avanzino, L. Learning by observing: The effect of multiple sessions of action-observation training on the spontaneous movement tempo and motor resonance. Neuropsychologia 2017, 96, 89–95. [Google Scholar] [CrossRef]
- Lei, Y.; Bao, S.; Wang, J. The combined effects of action observation and passive proprioceptive training on adaptive motor learning. Neuroscience 2016, 331, 91–98. [Google Scholar] [CrossRef]
- Salfi, F.; Tempesta, D.; De Gennaro, L.; Ferrara, M. Cued Memory Reactivation during Motor Imagery Practice Influences Early Improvement of Procedural Skill Learning. Neuroscience 2019, 418, 244–253. [Google Scholar] [CrossRef]
- Sobierajewicz, J.; Przekoracka-Krawczyk, A.; Jaśkowski, W.; Verwey, W.B.; van der Lubbe, R. The influence of motor imagery on the learning of a fine hand motor skill. Exp. Brain Res. 2017, 235, 305–320. [Google Scholar] [CrossRef]
- Marusic, U.; Grosprêtre, S.; Paravlic, A.; Kovač, S.; Pišot, R.; Taube, W. Motor Imagery during Action Observation of Locomotor Tasks Improves Rehabilitation Outcome in Older Adults after Total Hip Arthroplasty. Neural Plast. 2018, 2018, 1–9. [Google Scholar] [CrossRef]
- Bassolino, M.; Campanella, M.; Bove, M.; Pozzo, T.; Fadiga, L. Training the Motor Cortex by Observing the Actions of Others During Immobilization. Cereb. Cortex 2014, 24, 3268–3276. [Google Scholar] [CrossRef]
- Sakamoto, M.; Muraoka, T.; Mizuguchi, N.; Kanosue, K. Combining observation and imagery of an action enhances human corticospinal excitability. Neurosci. Res. 2009, 65, 23–27. [Google Scholar] [CrossRef] [PubMed]
- Vogt, S.; Rienzo, F.D.; Collet, C.; Collins, A.; Guillot, A. Multiple roles of motor imagery during action observation. Front. Hum. Neurosci. 2013, 7, 807. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Collet, C.; Di Rienzo, F.; El Hoyek, N.; Guillot, A. Autonomic nervous system correlates in movement observation and motor imagery. Front. Hum. Neurosci. 2013, 7, 415. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wriessnegger, S.C.; Kirchmeyr, D.; Bauernfeind, G.; Müller-Putz, G.R. Force related hemodynamic responses during execution and imagery of a hand grip task: A functional near infrared spectroscopy study. Brain Cogn. 2017, 117, 108–116. [Google Scholar] [CrossRef]
- Williams, S.E.; Guillot, A.; Di Rienzo, F.; Cumming, J. Comparing self-report and mental chronometry measures of motor imagery ability. Eur. J. Sport Sci. 2015, 15, 703–711. [Google Scholar] [CrossRef]
- Di Corrado, D.; Guarnera, M.; Quartiroli, A. Vividness and Transformation of Mental Images in Karate and Ballet. Percept. Mot. Skills 2014, 119, 764–773. [Google Scholar] [CrossRef] [Green Version]
- Paris-Alemany, A.; La Touche, R.; Agudo-Carmona, D.; Fernández-Carnero, J.; Gadea-Mateos, L.; Suso-Martí, L.; Cuenca-Martínez, F. Visual motor imagery predominance in professional Spanish dancers. Somatosens. Mot. Res. 2019, 36, 179–188. [Google Scholar]
- La Touche, R.; Grande-Alonso, M.; Cuenca-Martínez, F.; Gónzález-Ferrero, L.; Suso-Martí, L.; Paris-Alemany, A. Diminished Kinesthetic and Visual Motor Imagery Ability in Adults with Chronic Low Back Pain. PMR 2018, 11, 227–235. [Google Scholar] [CrossRef]
- Pijnenburg, M.; Brumagne, S.; Caeyenberghs, K.; Janssens, L.; Goossens, N.; Marinazzo, D.; Swinnen, S.P.; Claeys, K.; Siugzdaite, R. Resting-State Functional Connectivity of the Sensorimotor and the Association with the Sit-to-Stand-to-Sit Task. Brain Connect. 2015, 5, 303–311. [Google Scholar] [CrossRef]
- Taube, W.; Mouthon, M.; Leukel, C.; Hoogewoud, H.-M.; Annoni, J.-M.; Keller, M. Brain activity during observation and motor imagery of different balance tasks: An fMRI study. Cortex 2015, 64, 102–114. [Google Scholar] [CrossRef] [Green Version]
- Isaac, A.R.; Marks, D.F. Individual differences in mental imagery experience: Developmental changes and specialization. Br. J. Psychol. 1994, 85 Pt 4, 479–500. [Google Scholar] [CrossRef]
- Cuenca-Martínez, F.; Suso-Martí, L.; Grande-Alonso, M.; Paris-Alemany, A.; La Touche, R. Combining motor imagery with action observation training does not lead to a greater autonomic nervous system response than motor imagery alone during simple and functional movements: A randomized controlled trial. PeerJ 2018, 6, e5142. [Google Scholar] [CrossRef] [PubMed]
- Parsons, L.M. Temporal and kinematic properties of motor behavior reflected in mentally simulated action. J. Exp. Psychol. Hum. Percept. Perform. 1994, 20, 709–730. [Google Scholar] [CrossRef] [PubMed]
- Rieger, M. Motor imagery in typing: Effects of typing style and action familiarity. Psychon. Bull. Rev. 2012, 19, 101–107. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gatti, R.; Tettamanti, A.; Gough, P.M.; Riboldi, E.; Marinoni, L.; Buccino, G. Action observation versus motor imagery in learning a complex motor task: A short review of literature and a kinematics study. Neurosci. Lett. 2013, 540, 37–42. [Google Scholar] [CrossRef] [Green Version]
- Finke, R.A. The functional equivalence of mental images and errors of movement. Cogn. Psychol. 1979, 11, 235–264. [Google Scholar] [CrossRef]
- Lotze, M.; Montoya, P.; Erb, M.; Hülsmann, E.; Flor, H.; Klose, U.; Birbaumer, N.; Grodd, W. Activation of Cortical and Cerebellar Motor Areas during Executed and Imagined Hand Movements: An fMRI Study. J. Cogn. Neurosci. 1999, 11, 491–501. [Google Scholar] [CrossRef] [Green Version]
- Kim, S.-G.; Jennings, J.E.; Strupp, J.P.; Andersen, P.; Ugurbil, K. Functional MRI of human motor cortices during overt and imagined finger movements. Int. J. Imaging Syst. Technol. 1995, 6, 271–279. [Google Scholar] [CrossRef]
- Lacourse, M.G.; Orr, E.L.R.; Cramer, S.C.; Cohen, M.J. Brain activation during execution and motor imagery of novel and skilled sequential hand movements. Neuroimage 2005, 27, 505–519. [Google Scholar] [CrossRef] [Green Version]
- Jeannerod, M. The representing brain: Neural correlates of motor intention and imagery. Behav. Brain Sci. 1994, 17, 187–202. [Google Scholar] [CrossRef] [Green Version]
- Lindsay, P.H.; Norman, D.A. Human Information Processing: An Introduction to Psychology; Academic Press: Cambridge, MA, USA, 1972; ISBN 9780124509504. [Google Scholar]
- Guillot, A.; Collet, C. Duration of Mentally Simulated Movement: A Review. J. Mot. Behav. 2005, 37, 10–20. [Google Scholar] [CrossRef] [PubMed]
- Papaxanthis, C.; Schieppati, M.; Gentili, R.; Pozzo, T. Imagined and actual arm movements have similar durations when performed under different conditions of direction and mass. Exp. Brain Res. 2002, 143, 447–452. [Google Scholar] [CrossRef] [PubMed]
- Vissing, S.F.; Scherrer, U.; Victor, R.G. Stimulation of skin sympathetic nerve discharge by central command. Differential control of sympathetic outflow to skin and skeletal muscle during static exercise. Circ. Res. 1991, 69, 228–238. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Decety, J.; Jeannerod, M.; Durozard, D.; Baverel, G. Central activation of autonomic effectors during mental simulation of motor actions in man. J. Physiol. 1993, 461, 549–563. [Google Scholar] [CrossRef] [PubMed]
- Decety, J.; Jeannerod, M.; Germain, M.; Pastene, J. Vegetative response during imagined movement is proportional to mental effort. Behav. Brain Res. 1991, 42, 1–5. [Google Scholar] [CrossRef]
- Williams, J.; Pearce, A.J.; Loporto, M.; Morris, T.; Holmes, P.S. The relationship between corticospinal excitability during motor imagery and motor imagery ability. Behav. Brain Res. 2012, 226, 369–375. [Google Scholar] [CrossRef]
- Martin, K.A.; Moritz, S.E.; Hall, C.R. Imagery Use in Sport: A Literature Review and Applied Model. Sport Psychol. 1999, 13, 245–268. [Google Scholar] [CrossRef]
- Mattar, A.A.G.; Gribble, P.L. Motor Learning by Observing. Neuron 2005, 46, 153–160. [Google Scholar] [CrossRef] [Green Version]
- Destrebecqz, A.; Cleeremans, A. Can sequence learning be implicit? New evidence with the process dissociation procedure. Psychon. Bull. Rev. 2001, 8, 343–350. [Google Scholar] [CrossRef] [Green Version]
- Jongbloed-Pereboom, M.; Nijhuis-van der Sanden, M.W.G.; Steenbergen, B. Explicit and implicit motor sequence learning in children and adults; the role of age and visual working memory. Hum. Mov. Sci. 2019, 64, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Meissner, S.N.; Keitel, A.; Südmeyer, M.; Pollok, B. Implicit Motor Sequence Learning and Working Memory Performance Changes Across the Adult Life Span. Front. Aging Neurosci. 2016, 8, 89. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Steenbergen, B.; van der Kamp, J.; Verneau, M.; Jongbloed-Pereboom, M.; Masters, R.S.W. Implicit and explicit learning: Applications from basic research to sports for individuals with impaired movement dynamics. Disabil. Rehabil. 2010, 32, 1509–1516. [Google Scholar] [CrossRef] [PubMed]
- Postle, B.R. Working memory as an emergent property of the mind and brain. Neuroscience 2006, 139, 23–38. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bo, J.; Jennett, S.; Seidler, R.D. Working memory capacity correlates with implicit serial reaction time task performance. Exp. Brain Res. 2011, 214, 73–81. [Google Scholar] [CrossRef] [PubMed]
- Pascual-Leone, A.; Wassermann, E.; Grafman, J.; Hallett, M. The role of the dorsolateral prefrontal cortex in implicit procedural learning. Exp. Brain Res. 1996, 107, 479–485. [Google Scholar] [CrossRef]
- Baddeley, A. The episodic buffer: A new component of working memory? Trends Cogn. Sci. 2000, 4, 417–423. [Google Scholar] [CrossRef]
- Salmon, E.; Van der Linden, M.; Collette, F.; Delfiore, G.; Maquet, P.; Degueldre, C.; Luxen, A.; Franck, G. Regional brain activity during working memory tasks. Brain 1996, 119, 1617–1625. [Google Scholar] [CrossRef]
- De Zeeuw, C.I.; Ten Brinke, M.M. Motor Learning and the Cerebellum. Cold Spring Harb. Perspect. Biol. 2015, 7, a021683. [Google Scholar] [CrossRef] [Green Version]
- Thach, W.T. What is the role of the cerebellum in motor learning and cognition? Trends Cogn. Sci. 1998, 2, 331–337. [Google Scholar] [CrossRef]
- VanMeter, J.W.; Maisog, J.M.; Zeffiro, T.A.; Hallett, M.; Herscovitch, P.; Rapoport, S.I. Parametric Analysis of Functional Neuroimages: Application to a Variable-Rate Motor Task. Neuroimage 1995, 2, 273–283. [Google Scholar] [CrossRef]
- Lacourse, M.G.; Turner, J.A.; Randolph-Orr, E.; Schandler, S.L.; Cohen, M.J. Cerebral and cerebellar sensorimotor plasticity following motor imagery-based mental practice of a sequential movement. J. Rehabil. Res. Dev. 2004, 41, 505–524. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vogt, S.; Buccino, G.; Wohlschläger, A.M.; Canessa, N.; Shah, N.J.; Zilles, K.; Eickhoff, S.B.; Freund, H.-J.; Rizzolatti, G.; Fink, G.R. Prefrontal involvement in imitation learning of hand actions: Effects of practice and expertise. Neuroimage 2007, 37, 1371–1383. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Loporto, M.; McAllister, C.; Williams, J.; Hardwick, R.; Holmes, P. Investigating Central Mechanisms Underlying the Effects of Action Observation and Imagery Through Transcranial Magnetic Stimulation. J. Mot. Behav. 2011, 43, 361–373. [Google Scholar] [CrossRef] [PubMed]
- Roure, R.; Collet, C.; Deschaumes-Molinaro, C.; Delhomme, G.; Dittmar, A.; Vernet-Maury, E. Imagery quality estimated by autonomic response is correlated to sporting performance enhancement. Physiol. Behav. 1999, 66, 63–72. [Google Scholar] [CrossRef]
- Guillot, A.; Collet, C.; Dittmar, A. Relationship Between Visual and Kinesthetic Imagery, Field Dependence-Independence, and Complex Motor Skills. J. Psychophysiol. 2004, 18, 190–198. [Google Scholar] [CrossRef]
- Rizzolatti, G.; Fadiga, L.; Gallese, V.; Fogassi, L. Premotor cortex and the recognition of motor actions. Brain Res. Cogn. Brain Res. 1996, 3, 131–141. [Google Scholar] [CrossRef]
- Rizzolatti, G.; Craighero, L. The mirror-neuron system. Annu. Rev. Neurosci. 2004, 27, 169–192. [Google Scholar] [CrossRef] [Green Version]



{kind=link}
{kind=link}
| Trial | Population (Patients) | Intervention Data and Target | Results |
|---|---|---|---|
| Cabral-Sequeira et al. 2016 [20] | Adolescents with cerebral palsy: 11- to 16-year-old participants (mean = 13.58 years), who suffered left (n = 16) or right (n = 15) mild hemiparesis. | EG: Day 1: MI in isolation Day 2: MI plus physical training on motor learning an aiming task CG: Day 1: recreational activities Day 2: physical training on motor learning an aiming task. | MI increased motor learning as a function of side hemiparesis in comparison with a no MI intervention. |
| Kumar et al. 2016 [21] | Ambulant stroke subjects: 40 hemi paretic subjects (>3 months post-stroke) who were ambulant with good imagery ability. | EG (n = 20): task-oriented training group plus MI CG (n = 20): task-oriented training group in paretic lower extremity muscles strength and gait performance. | Additional task specific MI training improves paretic muscle strength and gait performance in ambulant stroke patients. |
| Keynen et al. 2018 [22] | Stroke patients: 56 patients with a stroke (>3 months ago), capacity to walk independently with or without a walking aid over 10 m (with a self-selected gait speed <1.2 m/s), and presence of hemiparesis (indicated by a score of <100 on the lower extremity part of the Motricity Index and a score <34 on the lower extremity part of the Brunnstrom Fugl–Meyer assessment). | AO group (n = 20) Analogy instruction (n = 19) Environmental group (n = 17) To explore immediate changes in walking performance when using the three implicit learning. | Analogy instructions and environmental constraints can lead to specific, immediate changes in the walking performance and were in general experienced as feasible by the participants. |
| La Touche et al. 2019 [9] | Patients with chronic non-specific low back pain: (low back pain for at least the prior three months; low back pain of nonspecific nature). | MI plus physical training (n = 16) Tactile feedback plus physical training (n = 16) CG: physical training in isolation (n = 16). Motor control gestures acquisition. | The MI strategy was the most effective mode for developing the motor control task in an accurate and controlled manner, obtaining better outcomes than tactile feedback or verbal instruction. |
| Moukarzel et al. 2019 [23] | Patients with total knee arthroplasty (n = 24). Four men and 20 women aged from 65 to 75 years (70 ± 2.89). | EG: MI plus physical therapy program (progressive lower-extremity strengthening exercises combined with electrical stimulation for quadriceps muscle, manual therapy, knee proprioceptive exercises, gait training, and functional exercises on stairs (n = 12). CG: physical therapy program in isolation (n = 12). Quadriceps strength, peak knee flexion during the swing phase, performance at the timed up and go test, stair climbing test, six-minute walk test, and Oxford knee score. | MI showed effectiveness in gait performance and functional recovery in a small sample of patients with total knee arthroplasty. |
| Trial | Population (Healthy Subjects) | Intervention Data and Target | Results |
| Cuenca-Martínez et al. 2019 [24] | HS (n = 45). Fourteen men and 31 women aged from 18 to 65 years. | MI plus physical training program for the lumbo-pelvic region (n = 15) AO plus program for the lumbo-pelvic (n = 15) CG: physical training in isolation (n = 15). Lumbo-pelvic motor control gestures acquisition. | AO training caused faster changes in lumbo-pelvic motor control compared with the CG group. All groups showed within-group significant differences between pre- and post-intervention. |
| Bek et al. 2016 [25] | HS (n = 50). The imagery group (n = 18, 5 males) had a mean age of 19.4 ± .98 years, the attention group (n = 15, 2 males) had a mean age of 19.9 ± 1.4 years, and the control group (n = 17, 1 male) had a mean age of 19.8 ± 1.7 years. | Two blocks of trials were completed, and after the first block, participants were instructed to imagine performing the observed movement (imagery group, n = 18) or attend closely to the characteristics of the movement (attention group, n = 15), or received no further instructions (control group, n = 17). To improve imitation with imagery or attention | Both attention and motor imagery can increase the accuracy of imitation and have implications for motor learning and rehabilitation. |
| Sheahan et al. 2018 [26] | HS (n = 58). (36 females; 25.0 ± 4.1 years). | Group 1: Follow through (n = 8), Group 2: Planning only (n = 8), Group 3: MI (n = 16), Group 4: No motor imagery (n = 16), Group 5: Motor imagery no fixation (n = 8). Motor gestures acquisition | Results showed that simply imagining different future movements could enable the learning and expression of multiple motor skills executed over the same physical states. When subjects performed the gesture and only imagined the follow-through, substantial learning occurred. |
| Dana & Gozalzadeh, 2017 [27] | Young male HS (n = 36) (15 to 18 years). | Internal MI plus physical practice (n = 12) External MI plus physical practice (n = 12) CG: no-imagery, mental math exercise plus physical practice (n = 12). The performance accuracy of the groups on the serve, forehand, and backhand strokes was measured. | Results showed significant increases in the performance accuracy of all three tennis strokes in all three groups, but serve accuracy in the internal imagery group and forehand accuracy in the external imagery group showed greater improvements, while backhand accuracy was similarly improved in all three groups. |
| Kim et al. 2017 [28] | HS (n = 40), novices. | Four groups: Action observation training (n = 10), Motor imagery training (n = 10), Physical practice (n = 10) and no practice (n = 10). Golf putting performance. | Results showed that the accuracy of the putting performance were improved over time through the two types of cognitive training (AO and MI training). |
| Gonzalez-Rosa et al. 2014 [29] | HS (n = 30), non-athletes, right-handed volunteers (17 females, 13 males, mean age 22.9 + 2.3 years). | Three groups: AO watched a video of the task (n = 9), MI had to imagine it (n = 12), and CG with a distracting computation task (n = 9). Early learning of a complex four limb, hand-foot coordination task, and kinematic analysis. | AO showed better learning compared with MI, and also elicited a stronger activity of the sensorimotor cortex during training, resulting in a lower amount of cortical activation during task execution. During AO, subjects appear to process and collect sensory and motor information relevant to action in an effective and efficient manner, which allowed them to apply a series of decision making strategies appropriate to defining which movement sequence to perform, and activating control processes such as feed forward control during motor execution. |
| Hidalgo-Pérez et al. 2015 [30] | HS (n = 40) 24 men and 16 women aged from 18 to 65 years. | Group 1: MI plus motor control exercise (n = 20), Group 2: motor control exercise in isolation (n = 20). Sensorimotor function of the craniocervical region and the cervical kinesthetic sense. | Combining MI with the motor control exercise produced statistically significant changes in sensorimotor function variables of the craniocervical region. Cervical kinesthetic sense was not significantly different between both groups. |
| Ingram et al. 2016 [31] | HS (n = 102) | Four groups: MI or PP tested in either perceptual (altering the sensory cue) or motor (switching the hand) transfer conditions (n = 60). CG (n = 42) that did not perform a transfer condition. Perceptual and motor learning through reaction time. | Results suggested that MI-based training relies on both perceptual and motor learning, while PP-based training relied more on motor processes. |
| Nishizawa & Kimura, 2017 [32] | HS females (n = 45) mean age 20.4 + 1.7 years). | Three groups: Model- and self-observation (n = 15), model-observation (n = 15), and self-observation (n = 15). Motor gesture learning through the acquisition of correct sports movement. | Observation combining model and self-observation exerted a positive effect on short-term motor gesture learning. |
| Kawasaki et al. 2018 [33] | Elderly HS (n = 36) aged 60 years or older (7 women and 29 men, mean age = 70.5 ± 6.19 years). | Three groups: Unskilled or skilled model observation groups (n = 12, respectively), or the CG (n = 12). Ball rotation performance (ball rotation speed). | Results indicated that the time taken for early phase learning of a finger coordination skill was improved when an unskilled model, rather than a skilled model, was used for AO combined with MI training. |
| Kraeutner et al. 2016 [34] | HS (n = 64) right-handed participants (42 female, 22.1 ± 5.3 years). | Two groups: MI in isolation (n = 31) Physical practice (n = 33) Implicit sequence learning task. | The magnitude of the learning did not differ between groups. It is suggested that MI and physical practice are equally effective in facilitating implicit sequence learning. |
| Kraeutner et al. 2017 [35] | HS (n = 72) Right-handed subjects (49 females, 23.8 ± 7.2 years) | Four conditions of MI-based practice: 4 training blocks with a high (4-High) or low (4-Low) sequence to noise ratio, or 2 training blocks with a high (2-High) or low (2-Low) sequence to noise ratio. Implicit sequence learning task. | Results showed that the extent to which implicit sequence learning occurs through MI is impacted by manipulations to entire training time and the sequence to noise ratio. In addition, results showed that the extent of implicit sequence learning occurring through MI is a function of exposure, indicating that like physical practice, the cognitive mechanisms of MI-based implicit sequence learning rely on the formation of stimulus response associations. |
| Lagravinese et al. 2016 [36] | HS (n = 25) | (AO) training: subjects were exposed to the observation of a video showing finger tapping movements executed at 3 Hz, a frequency higher than the spontaneous one (2 Hz) for four consecutive days. The changes in motor performance and motor resonance. | Results showed that multiple sessions of AO training induced a shift of the speed of execution of finger tapping movements toward the observed one and a change in motor resonance. |
| Lei et al. 2016 [37] | HS (n = 47) right-handed individuals (23 men, 17 women), aged from 18 to 30 years. | Five conditions: - AO, in which the subjects watched a video of a model who adapted to a novel visuomotor rotation - Proprioceptive training, in which the subject’s arm was moved passively to target locations that were associated with desired trajectories - Combined training, in which the subjects watched the video of a model during a half of the session and experienced passive movements during the other half - Active training, in which the subjects adapted actively to the rotation - A control condition, in which the subjects did not perform any task. | Results showed an improvement in visuomotor adaptation following the action observation, as compared with the adaptation performed by the individuals who were naïve to the given visuomotor rotation |
| Salfi et al. 2019 [38] | HS (n = 39) (aged 24.9 ± 3.0 years; range, 20–34; 18 males). | MI and Targeted memory reactivation. Four conditions: - MI in isolation - MI with an incompatible sound stimulation - AO - Auditory targeted memory reactivation during AO To assess the influence on performance on a sequential finger tapping task of an auditory targeted memory reactivation during MI practice. | The combination of MI and targeted memory reactivation showed the largest early performance improvement, as indexed by the combined measure of speed and accuracy |
| Sobierajewicz et al. 2016 [39] | HS (n = 24) 6 males and 18 females range 21 to 28 years. | After an informative cue, a response sequence had either to be executed, imagined, or withheld. The learning of a fine hand motor gesture. | Both physical condition and MI condition improved the response time and accuracy although the effect of motor learning by motor imagery was smaller than the effect of physical practice |
| Items | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | 9 | 10 | 11 | Total | |
| Cabral-Sequeira et al. 2016 [20] | 1 | 1 | 0 | 1 | 0 | 0 | 0 | 1 | 0 | 1 | 1 | 5 |
| Kumar et al. 2016 [21] | 1 | 1 | 1 | 1 | 0 | 0 | 1 | 1 | 0 | 1 | 1 | 7 |
| Keynen et al. 2018 [22] | 1 | 1 | 1 | 1 | 0 | 0 | 0 | 1 | 0 | 0 | 1 | 5 |
| La Touche et al. 2019 [9] | 1 | 1 | 1 | 1 | 1 | 0 | 1 | 1 | 0 | 1 | 1 | 8 |
| Moukarzel et al. 2019 [23] | 1 | 1 | 0 | 1 | 1 | 0 | 1 | 1 | 0 | 1 | 1 | 7 |
| Cuenca-Martínez et al. 2019 [24] | 1 | 1 | 1 | 1 | 1 | 0 | 1 | 1 | 0 | 1 | 1 | 8 |
| Bek et al. 2016 [25] | 1 | 1 | 0 | 1 | 0 | 0 | 0 | 1 | 0 | 1 | 1 | 5 |
| Sheahan et al. 2018 [26] | 1 | 0 | 0 | 1 | 0 | 0 | 0 | 1 | 0 | 1 | 1 | 4 |
| Dana & Gozalzadeh 2017 [27] | 1 | 0 | 0 | 0 | 0 | 0 | 0 | 1 | 0 | 1 | 1 | 3 |
| Kim et al. 2017 [28] | 1 | 1 | 0 | 1 | 0 | 0 | 0 | 1 | 0 | 1 | 1 | 5 |
| González-Rosa et al. 2014 [29] | 1 | 1 | 0 | 1 | 0 | 0 | 0 | 1 | 0 | 1 | 1 | 5 |
| Hidalgo-Pérez et al. 2015 [30] | 1 | 1 | 1 | 1 | 0 | 0 | 1 | 1 | 0 | 1 | 1 | 7 |
| Ingram et al. 2016 [31] | 1 | 1 | 0 | 1 | 0 | 0 | 0 | 1 | 0 | 1 | 1 | 5 |
| Nishizawa & Kimura, 2017 [32] | 1 | 1 | 0 | 1 | 0 | 0 | 0 | 1 | 0 | 1 | 0 | 4 |
| Kawasaki et al. 2018 [33] | 1 | 1 | 1 | 1 | 0 | 0 | 0 | 1 | 0 | 1 | 1 | 6 |
| Kraeutner et al. 2016 [34] | 1 | 1 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 1 | 1 | 4 |
| Kraeutner et al. 2017 [35] | 1 | 1 | 0 | 1 | 0 | 0 | 0 | 0 | 0 | 1 | 1 | 4 |
| Lagravinese et al. 2016 [36] | 1 | 0 | 0 | 1 | 1 | 0 | 0 | 1 | 0 | 1 | 1 | 5 |
| Lei et al. 2016 [37] | 1 | 0 | 0 | 1 | 0 | 0 | 0 | 1 | 0 | 1 | 0 | 3 |
| Salfi et al. 2019 [38] | 1 | 0 | 0 | 1 | 0 | 0 | 0 | 1 | 0 | 1 | 1 | 4 |
| Sobierajewicz et al. 2016 [39] | 1 | 0 | 0 | 1 | 0 | 0 | 0 | 0 | 0 | 1 | 1 | 3 |
| Domain | Variables | Influence |
|---|---|---|
| Physical MI *** AO * | - Levels of physical activity | - Greater physical activity levels might generate greater facility in constructing the movement due to the experience, development, and elaboration of habitual motor schemes. |
| - Perceived of mental fatigue | - The presence of high fatigue levels can affect attention, thereby limiting the brain’s construction of movement. | |
| - Disturbances in sensorimotor integration | - The presence of somatosensory disturbances can generate aberrant sensorimotor schemes that could affect the movement’s construction, thereby leading to a decreased ability to generate motor images. | |
| Cognitive–Evaluator MI *** AO * | - Understanding motor gestures and verbal instructions | - Understanding movements that are not physically elaborated can improve the planning phases of movement because emotional and cognitive limitations can be reduced. |
| - Context | - The development of the movement in family and specific contexts could facilitate imagination and observation. | |
| - Functioning of the working memory | - Better functioning of the working memory could increase the ability to collect the provided information and its subsequent consolidation into long-term memory, thereby facilitating the motor learning process. | |
| - Self-efficacy levels | - Greater self-perception of the ability to generate motor images could enhance the brain’s ability to construct motor images. | |
| - Attention levels | - Maintaining attention could facilitate the mental construction of movements and the total effort dedicated to that construction. | |
| - Expectations | - Expectations of the effects of movement representation techniques might influence the efficiency of the motor learning process. | |
| - Perception of difficulty | - Greater perception of the difficulty could lead to a reduced ability to generate motor representation and thereby worsen motor learning. | |
| Motivational–Emotional MI *** AO *** | - Motivation (reasons, intention, and desires) | - Higher motivation levels could lead directly to a better predisposition towards the learning process and, therefore, on the effects of movement representation techniques. |
| - Fear of movement | - Higher kinesiophobia levels can lead to an interruption of the motion representation process, thereby impairing the motor learning process. | |
| Direct modulation MI *** AO * | - Ability to create motor images | - The effectiveness of MI might depend on the ability to create motor images. This aspect can be influenced by other domains. |
| - Synchronization | - Greater time congruence between physical practice and motion representation could facilitate the motor learning process. | |
| - Activity of the autonomous nervous system | - Greater neurovegetative activity could indicate higher neurophysiological activity of the sensoriomotor cortical–subcortical networks, indicating greater effort dedicated to the task, greater attention, and less fatigue, thereby favoring motor learning. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cuenca-Martínez, F.; Suso-Martí, L.; León-Hernández, J.V.; La Touche, R. The Role of Movement Representation Techniques in the Motor Learning Process: A Neurophysiological Hypothesis and a Narrative Review. Brain Sci. 2020, 10, 27. https://doi.org/10.3390/brainsci10010027
Cuenca-Martínez F, Suso-Martí L, León-Hernández JV, La Touche R. The Role of Movement Representation Techniques in the Motor Learning Process: A Neurophysiological Hypothesis and a Narrative Review. Brain Sciences. 2020; 10(1):27. https://doi.org/10.3390/brainsci10010027
Chicago/Turabian StyleCuenca-Martínez, Ferran, Luis Suso-Martí, Jose Vicente León-Hernández, and Roy La Touche. 2020. "The Role of Movement Representation Techniques in the Motor Learning Process: A Neurophysiological Hypothesis and a Narrative Review" Brain Sciences 10, no. 1: 27. https://doi.org/10.3390/brainsci10010027
APA StyleCuenca-Martínez, F., Suso-Martí, L., León-Hernández, J. V., & La Touche, R. (2020). The Role of Movement Representation Techniques in the Motor Learning Process: A Neurophysiological Hypothesis and a Narrative Review. Brain Sciences, 10(1), 27. https://doi.org/10.3390/brainsci10010027