Interplay between Gating and Block of Ligand-Gated Ion Channels

Abstract
:1. Introduction
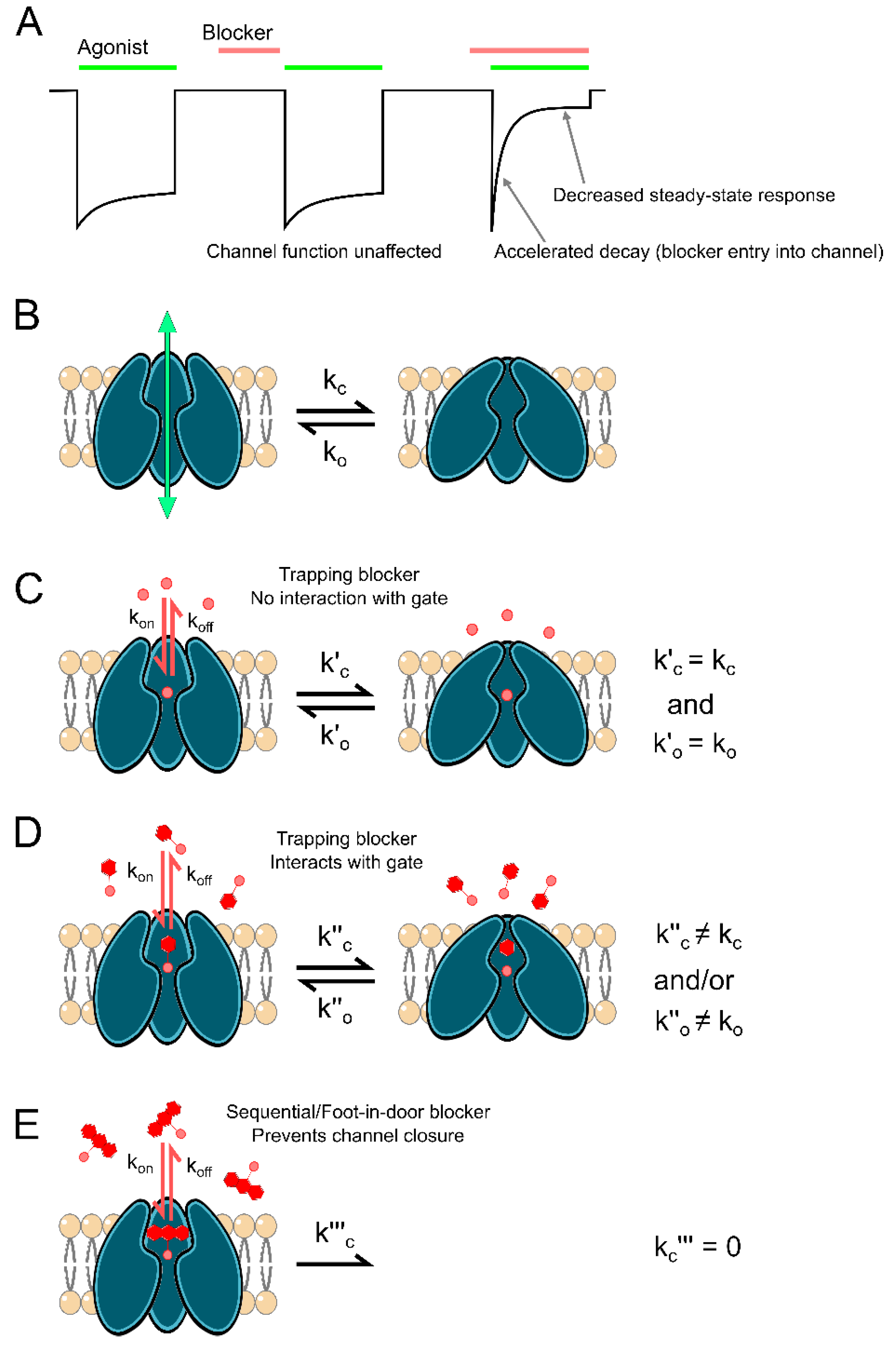
2. Reciprocal Interactions between Channel Block and Channel Gating
- Alter agonist binding and/or unbinding kinetics;
- Stabilize channel open states;
- Stabilize channel closed states.
3. Nicotinic Acetylcholine Receptors
Channel Block of nAChRs
4. Ionotropic Glutamate Receptors
5. Characteristics of AMPA and Kainate Receptor Block
5.1. Channel Block of AMPAR and KAR is Regulated by Channel Gating
5.2. Effects of Auxiliary Proteins on Gating of AMPARs and KARs Modulates Block by Endogenous Polyamines
5.3. Effects of Polyamine Block on Gating Transitions of AMPARs and KARs
6. Characteristics of NMDAR Channel Block
6.1. Sequential Blockers of NMDARs Prevent Channel Closure and Agonist Dissociation

{kind=link}
{kind=link}
| Compound | Structure | Type of Blocker | Effects on Gating |
|---|---|---|---|
| Magnesium |  | Unclear—due to fast unblocking kinetics, trapping of Mg2+ has not been directly demonstrated. | None [13,202]. |
| 9-aminoacridine |  | Sequential [7,201]. | Stabilizes open state [7,201]. Prevents agonist dissociation [7,201]. |
| IEM-1754 |  | Depolarized potentials: sequential [173]. Strongly negative potentials: trapping [173]. | Depolarized potentials: Stabilizes open state [173]. |
| Amantadine |  | Partial trapping [12,13]. | Accelerates channel closure of native NMDARs and GluN1/2B receptors [8]. |
| Memantine |  | Partial trapping [8,16,197,203,204]. | Slows GluN1/2A receptor recovery from Ca2+-dependent desensitization [3]. |
| Ketamine |  | Trapping [204]. | Accelerates GluN1/2B receptor recovery from desensitization [3]. |
6.2. Trapping Channel Blockers Modulate NMDAR State Transitions
6.3. Channel Block by Mg2+ Does Not Appear to Affect NMDAR State Transitions
7. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Katz, B.; Thesleff, S. A study of the desensitization produced by acetylcholine at the motor end-plate. J. Physiol. 1957, 138, 63–80. [Google Scholar] [CrossRef] [PubMed]
- Mayer, M.L.; Westbrook, G.L. The action of N-methyl-D-aspartic acid on mouse spinal neurones in culture. J. Physiol. 1985, 361, 65–90. [Google Scholar] [CrossRef] [PubMed]
- Glasgow, N.G.; Povysheva, N.V.; Azofeifa, A.M.; Johnson, J.W. Memantine and ketamine differentially alter NMDA receptor desensitization. J. Neurosci. 2017, 37, 9686–9704. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Legendre, P.; Rosenmund, C.; Westbrook, G.L. Inactivation of NMDA channels in cultured hippocampal neurons by intracellular calcium. J. Neurosci. 1993, 13, 674–684. [Google Scholar] [CrossRef]
- Hille, B. Ion Channels of Excitable Membranes, 3rd ed.; Sinauer Associates Is an Imprint of Oxford University Press: Sunderland, MA, USA, 2001. [Google Scholar]
- Yuan, H.; Low, C.-M.; Moody, O.A.; Jenkins, A.; Traynelis, S.F. Ionotropic GABA and glutamate receptor mutations and human neurologic diseases. Mol. Pharmacol. 2015, 88, 203–217. [Google Scholar] [CrossRef]
- Benveniste, M.; Mayer, M.L. Trapping of glutamate and glycine during open channel block of rat hippocampal neuron NMDA receptors by 9-aminoacridine. J. Physiol. 1995, 483 Pt 2, 367–384. [Google Scholar] [CrossRef] [Green Version]
- Blanpied, T.A.; Clarke, R.J.; Johnson, J.W. Amantadine inhibits NMDA receptors by accelerating channel closure during channel block. J. Neurosci. 2005, 25, 3312–3322. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Heidmann, T.; Changeux, J.P. Characterization of the transient agonist-triggered state of the acetylcholine receptor rapidly labeled by the noncompetitive blocker [3H]chlorpromazine: Additional evidence for the open channel conformation. Biochemistry 1986, 25, 6109–6113. [Google Scholar] [CrossRef] [PubMed]
- Purohit, Y.; Grosman, C. Block of muscle nicotinic receptors by choline suggests that the activation and desensitization gates act as distinct molecular entities. J. Gen. Physiol. 2006, 127, 703–717. [Google Scholar] [CrossRef] [PubMed]
- Courtney, K.R. Mechanism of frequency-dependent inhibition of sodium currents in frog myelinated nerve by the lidocaine derivative GEA. J. Pharmacol. Exp. Ther. 1975, 195, 225–236. [Google Scholar]
- Blanpied, T.A.; Boeckman, F.A.; Aizenman, E.; Johnson, J.W. Trapping channel block of NMDA-activated responses by amantadine and memantine. J. Neurophysiol. 1997, 77, 309–323. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sobolevsky, A.I.; Yelshansky, M.V. The trapping block of NMDA receptor channels in acutely isolated rat hippocampal neurones. J. Physiol. 2000, 526 Pt 3, 493–506. [Google Scholar] [CrossRef]
- Mealing, G.A.; Lanthorn, T.H.; Small, D.L.; Murray, R.J.; Mattes, K.C.; Comas, T.M.; Morley, P. Structural modifications to an N-methyl-D-aspartate receptor antagonist result in large differences in trapping block. J. Pharmacol. Exp. Ther. 2001, 297, 906–914. [Google Scholar] [PubMed]
- Bolshakov, K.V.; Gmiro, V.E.; Tikhonov, D.B.; Magazanik, L.G. Determinants of trapping block of N-methyl-d-aspartate receptor channels. J. Neurochem. 2003, 87, 56–65. [Google Scholar] [CrossRef]
- Kotermanski, S.E.; Wood, J.T.; Johnson, J.W. Memantine binding to a superficial site on NMDA receptors contributes to partial trapping. J. Physiol. 2009, 587, 4589–4604. [Google Scholar] [CrossRef] [PubMed]
- Sobolevsky, A.I.; Koshelev, S.G.; Khodorov, B.I. Probing of NMDA channels with fast blockers. J. Neurosci. 1999, 19, 10611–10626. [Google Scholar] [CrossRef] [PubMed]
- Armstrong, C.M. Interaction of tetraethylammonium ion derivatives with the potassium channels of giant axons. J. Gen. Physiol. 1971, 58, 413–437. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ruff, R.L. A quantitative analysis of local anaesthetic alteration of miniature end-plate currents and end-plate current fluctuations. J. Physiol. 1977, 264, 89–124. [Google Scholar] [CrossRef]
- Neher, E.; Steinbach, J.H. Local anaesthetics transiently block currents through single acetylcholine-receptor channels. J. Physiol. 1978, 277, 153–176. [Google Scholar] [CrossRef]
- Song, X.; Jensen, M.Ø.; Jogini, V.; Stein, R.A.; Lee, C.-H.; Mchaourab, H.S.; Shaw, D.E.; Gouaux, E. Mechanism of NMDA receptor channel block by MK-801 and memantine. Nature 2018, 556, 515–519. [Google Scholar] [CrossRef]
- Langley, J.N. On the reaction of cells and of nerve-endings to certain poisons, chiefly as regards the reaction of striated muscle to nicotine and to curari. J. Physiol. 1905, 33, 374–413. [Google Scholar] [CrossRef] [PubMed]
- Noda, M.; Takahashi, H.; Tanabe, T.; Toyosato, M.; Kikyotani, S.; Furutani, Y.; Hirose, T.; Takashima, H.; Inayama, S.; Miyata, T.; et al. Structural homology of Torpedo californica acetylcholine receptor subunits. Nature 1983, 302, 528–532. [Google Scholar] [CrossRef] [PubMed]
- Devillers-Thiery, A.; Giraudat, J.; Bentaboulet, M.; Changeux, J.P. Complete mRNA coding sequence of the acetylcholine binding alpha-subunit of Torpedo marmorata acetylcholine receptor: A model for the transmembrane organization of the polypeptide chain. Proc. Natl. Acad. Sci. USA 1983, 80, 2067–2071. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Claudio, T.; Ballivet, M.; Patrick, J.; Heinemann, S. Nucleotide and deduced amino acid sequences of Torpedo californica acetylcholine receptor gamma subunit. Proc. Natl. Acad. Sci. USA 1983, 80, 1111–1115. [Google Scholar] [CrossRef] [Green Version]
- Cartaud, J.; Benedetti, E.L.; Cohen, J.B.; Meunier, J.C.; Changeux, J.P. Presence of a lattice structure in membrane fragments rich in nicotinic receptor protein from the electric organ of Torpedo marmorata. FEBS Lett. 1973, 33, 109–113. [Google Scholar] [CrossRef] [Green Version]
- Miledi, R.; Molinoff, P.; Potter, L.T. Isolation of the cholinergic receptor protein of Torpedo electric tissue. Nature 1971, 229, 554–557. [Google Scholar] [CrossRef]
- Karlin, A.; Prives, J.; Deal, W.; Winnik, M. Affinity labeling of the acetylcholine receptor in the electroplax. J. Mol. Biol. 1971, 61, 175–188. [Google Scholar] [CrossRef]
- Changeux, J.P.; Kasai, M.; Lee, C.Y. Use of a snake venom toxin to characterize the cholinergic receptor protein. Proc. Natl. Acad. Sci. USA 1970, 67, 1241–1247. [Google Scholar] [CrossRef] [Green Version]
- Changeux, J.-P. The nicotinic acetylcholine receptor: The founding father of the pentameric ligand-gated ion channel superfamily. J. Biol. Chem. 2012, 287, 40207–40215. [Google Scholar] [CrossRef] [Green Version]
- Miyazawa, A.; Fujiyoshi, Y.; Unwin, N. Structure and gating mechanism of the acetylcholine receptor pore. Nature 2003, 423, 949–955. [Google Scholar] [CrossRef]
- Colquhoun, D.; Hawkes, A.G. Relaxation and fluctuations of membrane currents that flow through drug-operated channels. Proc. R. Soc. Lond. B Biol. Sci. 1977, 199, 231–262. [Google Scholar] [CrossRef] [PubMed]
- Neher, E.; Sakmann, B. Single-channel currents recorded from membrane of denervated frog muscle fibres. Nature 1976, 260, 799–802. [Google Scholar] [CrossRef] [PubMed]
- Hamill, O.P.; Marty, A.; Neher, E.; Sakmann, B.; Sigworth, F.J. Improved patch-clamp techniques for high-resolution current recording from cells and cell-free membrane patches. Pflugers Arch. 1981, 391, 85–100. [Google Scholar] [CrossRef] [PubMed]
- Caillé, S.; Guillem, K.; Cador, M.; Manzoni, O.; Georges, F. Voluntary nicotine consumption triggers in vivo potentiation of cortical excitatory drives to midbrain dopaminergic neurons. J. Neurosci. 2009, 29, 10410–10415. [Google Scholar] [CrossRef] [PubMed]
- Mao, D.; Gallagher, K.; McGehee, D.S. Nicotine potentiation of excitatory inputs to ventral tegmental area dopamine neurons. J. Neurosci. 2011, 31, 6710–6720. [Google Scholar] [CrossRef]
- Levin, E.D. Nicotinic receptor subtypes and cognitive function. J. Neurobiol. 2002, 53, 633–640. [Google Scholar] [CrossRef] [Green Version]
- Dani, J.A. Neuronal nicotinic acetylcholine receptor structure and function and response to nicotine. Int. Rev. Neurobiol. 2015, 124, 3–19. [Google Scholar] [CrossRef] [Green Version]
- Ji, D.; Lape, R.; Dani, J.A. Timing and location of nicotinic activity enhances or depresses hippocampal synaptic plasticity. Neuron 2001, 31, 131–141. [Google Scholar] [CrossRef] [Green Version]
- McKay, B.E.; Placzek, A.N.; Dani, J.A. Regulation of synaptic transmission and plasticity by neuronal nicotinic acetylcholine receptors. Biochem. Pharmacol. 2007, 74, 1120–1133. [Google Scholar] [CrossRef] [Green Version]
- Bertrand, S.; Weiland, S.; Berkovic, S.F.; Steinlein, O.K.; Bertrand, D. Properties of neuronal nicotinic acetylcholine receptor mutants from humans suffering from autosomal dominant nocturnal frontal lobe epilepsy. Br. J. Pharmacol. 1998, 125, 751–760. [Google Scholar] [CrossRef] [Green Version]
- Vallés, A.S.; Borroni, M.V.; Barrantes, F.J. Targeting brain α7 nicotinic acetylcholine receptors in Alzheimer’s disease: Rationale and current status. CNS Drugs 2014, 28, 975–987. [Google Scholar] [CrossRef] [PubMed]
- Yakel, J.L. Cholinergic receptors: Functional role of nicotinic ACh receptors in brain circuits and disease. Pflugers Arch. 2013, 465, 441–450. [Google Scholar] [CrossRef] [PubMed]
- Martínez-Hernández, R.; Bernal, S.; Also-Rallo, E.; Alías, L.; Barceló, M.J.; Hereu, M.; Esquerda, J.E.; Tizzano, E.F. Synaptic defects in type I spinal muscular atrophy in human development. J. Pathol. 2013, 229, 49–61. [Google Scholar] [CrossRef]
- Steinlein, O.K.; Bertrand, D. Neuronal nicotinic acetylcholine receptors: From the genetic analysis to neurological diseases. Biochem. Pharmacol. 2008, 76, 1175–1183. [Google Scholar] [CrossRef]
- Leonard, S.; Mexal, S.; Freedman, R. Smoking, genetics and schizophrenia: Evidence for self medication. J. Dual Diagn. 2007, 3, 43–59. [Google Scholar] [CrossRef] [Green Version]
- Lindstrom, J. Autoimmune diseases involving nicotinic receptors. J. Neurobiol. 2002, 53, 656–665. [Google Scholar] [CrossRef]
- Tarr, T.B.; Malick, W.; Liang, M.; Valdomir, G.; Frasso, M.; Lacomis, D.; Reddel, S.W.; Garcia-Ocano, A.; Wipf, P.; Meriney, S.D. Evaluation of a novel calcium channel agonist for therapeutic potential in Lambert-Eaton myasthenic syndrome. J. Neurosci. 2013, 33, 10559–10567. [Google Scholar] [CrossRef] [Green Version]
- Tarr, T.B.; Wipf, P.; Meriney, S.D. Synaptic Pathophysiology and Treatment of Lambert-Eaton Myasthenic Syndrome. Mol. Neurobiol. 2015, 52, 456–463. [Google Scholar] [CrossRef] [Green Version]
- Thompson, A.J.; Lester, H.A.; Lummis, S.C.R. The structural basis of function in Cys-loop receptors. Q. Rev. Biophys. 2010, 43, 449–499. [Google Scholar] [CrossRef] [Green Version]
- Séguéla, P.; Wadiche, J.; Dineley-Miller, K.; Dani, J.A.; Patrick, J.W. Molecular cloning, functional properties, and distribution of rat brain alpha 7: A nicotinic cation channel highly permeable to calcium. J. Neurosci. 1993, 13, 596–604. [Google Scholar] [CrossRef] [Green Version]
- Cooper, E.; Couturier, S.; Ballivet, M. Pentameric structure and subunit stoichiometry of a neuronal nicotinic acetylcholine receptor. Nature 1991, 350, 235–238. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wada, E.; Wada, K.; Boulter, J.; Deneris, E.; Heinemann, S.; Patrick, J.; Swanson, L.W. Distribution of alpha 2, alpha 3, alpha 4, and beta 2 neuronal nicotinic receptor subunit mRNAs in the central nervous system: A hybridization histochemical study in the rat. J. Comp. Neurol. 1989, 284, 314–335. [Google Scholar] [CrossRef] [PubMed]
- Palma, E.; Bertrand, S.; Binzoni, T.; Bertrand, D. Neuronal nicotinic alpha 7 receptor expressed in Xenopus oocytes presents five putative binding sites for methyllycaconitine. J. Physiol. 1996, 491 Pt 1, 151–161. [Google Scholar] [CrossRef]
- Taly, A.; Corringer, P.-J.; Guedin, D.; Lestage, P.; Changeux, J.-P. Nicotinic receptors: Allosteric transitions and therapeutic targets in the nervous system. Nat. Rev. Drug Discov. 2009, 8, 733–750. [Google Scholar] [CrossRef]
- Fasoli, F.; Gotti, C. Structure of neuronal nicotinic receptors. Curr. Top. Behav. Neurosci. 2015, 23, 1–17. [Google Scholar] [CrossRef]
- Vernino, S.; Amador, M.; Luetje, C.W.; Patrick, J.; Dani, J.A. Calcium modulation and high calcium permeability of neuronal nicotinic acetylcholine receptors. Neuron 1992, 8, 127–134. [Google Scholar] [CrossRef]
- Dani, J.A.; Eisenman, G. Monovalent and divalent cation permeation in acetylcholine receptor channels. Ion transport related to structure. J. Gen. Physiol. 1987, 89, 959–983. [Google Scholar] [CrossRef]
- Auerbach, A. Agonist activation of a nicotinic acetylcholine receptor. Neuropharmacology 2015, 96, 150–156. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dale, H. Chemical transmission of the effects of nerve impulses. Br. Med. J. 1934, 1, 835–841. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chang, C.C.; Lee, C.Y. Isolation of neurotoxins from the venom of bungarus multicinctus and their modes of neuromuscular blocking action. Arch. Int. Pharm. Ther. 1963, 144, 241–257. [Google Scholar]
- Löscher, W.; Rogawski, M.A. How theories evolved concerning the mechanism of action of barbiturates. Epilepsia 2012, 53 (Suppl. 8), 12–25. [Google Scholar] [CrossRef] [PubMed]
- Steinbach, A.B. Alteration by xylocaine (lidocaine) and its derivatives of the time course of the end plate potential. J. Gen. Physiol. 1968, 52, 144–161. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Steinbach, A.B. A kinetic model for the action of xylocaine on receptors for acetylcholine. J. Gen. Physiol. 1968, 52, 162–180. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Adams, P.R. Voltage jump analysis of procaine action at frog end-plate. J. Physiol. 1977, 268, 291–318. [Google Scholar] [CrossRef] [Green Version]
- Adams, P.R. Drug blockade of open end-plate channels. J. Physiol. 1976, 260, 531–552. [Google Scholar] [CrossRef]
- Adams, P.R. A model for the procaine end-plate current. J. Physiol. 1975, 246, 61P–63P. [Google Scholar] [PubMed]
- Auerbach, A.; Akk, G. Desensitization of mouse nicotinic acetylcholine receptor channels. A two-gate mechanism. J. Gen. Physiol. 1998, 112, 181–197. [Google Scholar] [CrossRef] [Green Version]
- Lape, R.; Krashia, P.; Colquhoun, D.; Sivilotti, L.G. Agonist and blocking actions of choline and tetramethylammonium on human muscle acetylcholine receptors. J. Physiol. 2009, 587, 5045–5072. [Google Scholar] [CrossRef]
- Chiara, D.C.; Hamouda, A.K.; Ziebell, M.R.; Mejia, L.A.; Garcia, G.; Cohen, J.B. [(3)H]chlorpromazine photolabeling of the torpedo nicotinic acetylcholine receptor identifies two state-dependent binding sites in the ion channel. Biochemistry 2009, 48, 10066–10077. [Google Scholar] [CrossRef] [Green Version]
- Giraudat, J.; Dennis, M.; Heidmann, T.; Chang, J.Y.; Changeux, J.P. Structure of the high-affinity binding site for noncompetitive blockers of the acetylcholine receptor: Serine-262 of the delta subunit is labeled by [3H]chlorpromazine. Proc. Natl. Acad. Sci. USA 1986, 83, 2719–2723. [Google Scholar] [CrossRef] [Green Version]
- Revah, F.; Galzi, J.L.; Giraudat, J.; Haumont, P.Y.; Lederer, F.; Changeux, J.P. The noncompetitive blocker [3H]chlorpromazine labels three amino acids of the acetylcholine receptor gamma subunit: Implications for the alpha-helical organization of regions MII and for the structure of the ion channel. Proc. Natl. Acad. Sci. USA 1990, 87, 4675–4679. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gielen, M.; Corringer, P.-J. The dual-gate model for pentameric ligand-gated ion channels activation and desensitization. J. Physiol. 2018, 596, 1873–1902. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Paoletti, P.; Bellone, C.; Zhou, Q. NMDA receptor subunit diversity: Impact on receptor properties, synaptic plasticity and disease. Nat. Rev. Neurosci. 2013, 14, 383–400. [Google Scholar] [CrossRef] [PubMed]
- Traynelis, S.F.; Wollmuth, L.P.; McBain, C.J.; Menniti, F.S.; Vance, K.M.; Ogden, K.K.; Hansen, K.B.; Yuan, H.; Myers, S.J.; Dingledine, R. Glutamate receptor ion channels: Structure, regulation, and function. Pharmacol. Rev. 2010, 62, 405–496. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Burnashev, N.; Szepetowski, P. NMDA receptor subunit mutations in neurodevelopmental disorders. Curr. Opin. Pharmacol. 2015, 20, 73–82. [Google Scholar] [CrossRef] [PubMed]
- Salpietro, V.; Dixon, C.L.; Guo, H.; Bello, O.D.; Vandrovcova, J.; Efthymiou, S.; Maroofian, R.; Heimer, G.; Burglen, L.; Valence, S.; et al. AMPA receptor GluA2 subunit defects are a cause of neurodevelopmental disorders. Nat. Commun. 2019, 10, 3094. [Google Scholar] [CrossRef] [Green Version]
- Lee, E.-J.; Choi, S.Y.; Kim, E. NMDA receptor dysfunction in autism spectrum disorders. Curr. Opin. Pharmacol. 2015, 20, 8–13. [Google Scholar] [CrossRef]
- Zorumski, C.F.; Olney, J.W. Excitotoxic neuronal damage and neuropsychiatric disorders. Pharmacol. Ther. 1993, 59, 145–162. [Google Scholar] [CrossRef]
- Lau, C.G.; Zukin, R.S. NMDA receptor trafficking in synaptic plasticity and neuropsychiatric disorders. Nat. Rev. Neurosci. 2007, 8, 413–426. [Google Scholar] [CrossRef]
- Javitt, D.C. Glutamate as a therapeutic target in psychiatric disorders. Mol. Psychiatry 2004, 9, 984–997. [Google Scholar] [CrossRef] [Green Version]
- Bowie, D. Ionotropic glutamate receptors & CNS disorders. CNS Neurol. Disord. Drug Targets 2008, 7, 129–143. [Google Scholar] [PubMed] [Green Version]
- Benamer, N.; Marti, F.; Lujan, R.; Hepp, R.; Aubier, T.G.; Dupin, A.A.M.; Frébourg, G.; Pons, S.; Maskos, U.; Faure, P.; et al. GluD1, linked to schizophrenia, controls the burst firing of dopamine neurons. Mol. Psychiatry 2018, 23, 691–700. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ady, V.; Perroy, J.; Tricoire, L.; Piochon, C.; Dadak, S.; Chen, X.; Dusart, I.; Fagni, L.; Lambolez, B.; Levenes, C. Type 1 metabotropic glutamate receptors (mGlu1) trigger the gating of GluD2 delta glutamate receptors. EMBO Rep. 2014, 15, 103–109. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gantz, S.C.; Moussawi, K.; Hake, H.S. Delta glutamate receptor conductance drives excitation of mouse dorsal raphe neurons. eLife 2020, 9, e56054. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Araki, K.; Meguro, H.; Kushiya, E.; Takayama, C.; Inoue, Y.; Mishina, M. Selective expression of the glutamate receptor channel delta 2 subunit in cerebellar Purkinje cells. Biochem. Biophys. Res. Commun. 1993, 197, 1267–1276. [Google Scholar] [CrossRef] [PubMed]
- Lomeli, H.; Sprengel, R.; Laurie, D.J.; Köhr, G.; Herb, A.; Seeburg, P.H.; Wisden, W. The rat delta-1 and delta-2 subunits extend the excitatory amino acid receptor family. FEBS Lett. 1993, 315, 318–322. [Google Scholar] [CrossRef]
- Yamazaki, M.; Araki, K.; Shibata, A.; Mishina, M. Molecular cloning of a cDNA encoding a novel member of the mouse glutamate receptor channel family. Biochem. Biophys. Res. Commun. 1992, 183, 886–892. [Google Scholar] [CrossRef]
- Orth, A.; Tapken, D.; Hollmann, M. The delta subfamily of glutamate receptors: Characterization of receptor chimeras and mutants. Eur. J. Neurosci. 2013, 37, 1620–1630. [Google Scholar] [CrossRef]
- Premkumar, L.S.; Auerbach, A. Identification of a high affinity divalent cation binding site near the entrance of the NMDA receptor channel. Neuron 1996, 16, 869–880. [Google Scholar] [CrossRef] [Green Version]
- Hume, R.I.; Dingledine, R.; Heinemann, S.F. Identification of a site in glutamate receptor subunits that controls calcium permeability. Science 1991, 253, 1028–1031. [Google Scholar] [CrossRef]
- Sommer, B.; Köhler, M.; Sprengel, R.; Seeburg, P.H. RNA editing in brain controls a determinant of ion flow in glutamate-gated channels. Cell 1991, 67, 11–19. [Google Scholar] [CrossRef]
- Burnashev, N.; Schoepfer, R.; Monyer, H.; Ruppersberg, J.P.; Günther, W.; Seeburg, P.H.; Sakmann, B. Control by asparagine residues of calcium permeability and magnesium blockade in the NMDA receptor. Science 1992, 257, 1415–1419. [Google Scholar] [CrossRef] [PubMed]
- Twomey, E.C.; Yelshanskaya, M.V.; Grassucci, R.A.; Frank, J.; Sobolevsky, A.I. Channel opening and gating mechanism in AMPA-subtype glutamate receptors. Nature 2017, 549, 60–65. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Twomey, E.C.; Sobolevsky, A.I. Structural mechanisms of gating in ionotropic glutamate receptors. Biochemistry 2018, 57, 267–276. [Google Scholar] [CrossRef]
- Twomey, E.C.; Yelshanskaya, M.V.; Vassilevski, A.A.; Sobolevsky, A.I. Mechanisms of Channel Block in Calcium-Permeable AMPA Receptors. Neuron 2018, 99, 956–968.e4. [Google Scholar] [CrossRef] [Green Version]
- Nakagawa, T. Structures of the AMPA receptor in complex with its auxiliary subunit cornichon. Science 2019, 366, 1259–1263. [Google Scholar] [CrossRef]
- Chou, T.-H.; Tajima, N.; Romero-Hernandez, A.; Furukawa, H. Structural basis of functional transitions in mammalian NMDA receptors. Cell 2020, 182, 357–371.e13. [Google Scholar] [CrossRef]
- Ferrer-Montiel, A.V.; Merino, J.M.; Planells-Cases, R.; Sun, W.; Montal, M. Structural determinants of the blocker binding site in glutamate and NMDA receptor channels. Neuropharmacology 1998, 37, 139–147. [Google Scholar] [CrossRef]
- Fedele, L.; Newcombe, J.; Topf, M.; Gibb, A.; Harvey, R.J.; Smart, T.G. Disease-associated missense mutations in GluN2B subunit alter NMDA receptor ligand binding and ion channel properties. Nat. Commun. 2018, 9, 957. [Google Scholar] [CrossRef]
- Mori, H.; Masaki, H.; Yamakura, T.; Mishina, M. Identification by mutagenesis of a Mg(2+)-block site of the NMDA receptor channel. Nature 1992, 358, 673–675. [Google Scholar] [CrossRef]
- Lemke, J.R.; Hendrickx, R.; Geider, K.; Laube, B.; Schwake, M.; Harvey, R.J.; James, V.M.; Pepler, A.; Steiner, I.; Hörtnagel, K.; et al. GRIN2B mutations in West syndrome and intellectual disability with focal epilepsy. Ann. Neurol. 2014, 75, 147–154. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mesbahi-Vasey, S.; Veras, L.; Yonkunas, M.; Johnson, J.W.; Kurnikova, M.G. All atom NMDA receptor transmembrane domain model development and simulations in lipid bilayers and water. PLoS ONE 2017, 12, e0177686. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Trott, O.; Olson, A.J. AutoDock Vina: Improving the speed and accuracy of docking with a new scoring function, efficient optimization, and multithreading. J. Comput. Chem. 2010, 31, 455–461. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Humphrey, W.; Dalke, A.; Schulten, K. VMD: Visual molecular dynamics. J. Mol. Graph. 1996, 14, 33–38. [Google Scholar] [CrossRef]
- Smith, T.C.; Howe, J.R. Concentration-dependent substate behavior of native AMPA receptors. Nat. Neurosci. 2000, 3, 992–997. [Google Scholar] [CrossRef]
- Jin, R.; Banke, T.G.; Mayer, M.L.; Traynelis, S.F.; Gouaux, E. Structural basis for partial agonist action at ionotropic glutamate receptors. Nat. Neurosci. 2003, 6, 803–810. [Google Scholar] [CrossRef]
- Rosenmund, C.; Stern-Bach, Y.; Stevens, C.F. The tetrameric structure of a glutamate receptor channel. Science 1998, 280, 1596–1599. [Google Scholar] [CrossRef] [Green Version]
- Trussell, L.O.; Fischbach, G.D. Glutamate receptor desensitization and its role in synaptic transmission. Neuron 1989, 3, 209–218. [Google Scholar] [CrossRef]
- Higuchi, M.; Single, F.N.; Köhler, M.; Sommer, B.; Sprengel, R.; Seeburg, P.H. RNA editing of AMPA receptor subunit GluR-B: A base-paired intron-exon structure determines position and efficiency. Cell 1993, 75, 1361–1370. [Google Scholar] [CrossRef]
- Rosenthal, J.J.C.; Seeburg, P.H. A-to-I RNA editing: Effects on proteins key to neural excitability. Neuron 2012, 74, 432–439. [Google Scholar] [CrossRef] [Green Version]
- Melcher, T.; Maas, S.; Higuchi, M.; Keller, W.; Seeburg, P.H. Editing of alpha-amino-3-hydroxy-5-methylisoxazole-4-propionic acid receptor GluR-B pre-mRNA in vitro reveals site-selective adenosine to inosine conversion. J. Biol. Chem. 1995, 270, 8566–8570. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hollmann, M.; Hartley, M.; Heinemann, S. Ca2+ permeability of KA-AMPA—Gated glutamate receptor channels depends on subunit composition. Science 1991, 252, 851–853. [Google Scholar] [CrossRef] [PubMed]
- Burnashev, N.; Monyer, H.; Seeburg, P.H.; Sakmann, B. Divalent ion permeability of AMPA receptor channels is dominated by the edited form of a single subunit. Neuron 1992, 8, 189–198. [Google Scholar] [CrossRef]
- Bowie, D.; Mayer, M.L. Inward rectification of both AMPA and kainate subtype glutamate receptors generated by polyamine-mediated ion channel block. Neuron 1995, 15, 453–462. [Google Scholar] [CrossRef] [Green Version]
- Kamboj, S.K.; Swanson, G.T.; Cull-Candy, S.G. Intracellular spermine confers rectification on rat calcium-permeable AMPA and kainate receptors. J. Physiol. 1995, 486 Pt 2, 297–303. [Google Scholar] [CrossRef] [Green Version]
- Wilding, T.J.; Zhou, Y.; Huettner, J.E. Q/R site editing controls kainate receptor inhibition by membrane fatty acids. J. Neurosci. 2005, 25, 9470–9478. [Google Scholar] [CrossRef]
- Wilding, T.J.; Chen, K.; Huettner, J.E. Fatty acid modulation and polyamine block of GluK2 kainate receptors analyzed by scanning mutagenesis. J. Gen. Physiol. 2010, 136, 339–352. [Google Scholar] [CrossRef] [Green Version]
- Bowie, D.; Lange, G.D.; Mayer, M.L. Activity-dependent modulation of glutamate receptors by polyamines. J. Neurosci. 1998, 18, 8175–8185. [Google Scholar] [CrossRef] [Green Version]
- Rozov, A.; Zilberter, Y.; Wollmuth, L.P.; Burnashev, N. Facilitation of currents through rat Ca2+-permeable AMPA receptor channels by activity-dependent relief from polyamine block. J. Physiol. 1998, 511 Pt 2, 361–377. [Google Scholar] [CrossRef]
- Brown, P.M.G.E.; Aurousseau, M.R.P.; Musgaard, M.; Biggin, P.C.; Bowie, D. Kainate receptor pore-forming and auxiliary subunits regulate channel block by a novel mechanism. J. Physiol. 2016, 594, 1821–1840. [Google Scholar] [CrossRef]
- Brown, P.M.G.E.; McGuire, H.; Bowie, D. Stargazin and cornichon-3 relieve polyamine block of AMPA receptors by enhancing blocker permeation. J. Gen. Physiol. 2018, 150, 67–82. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Burnashev, N.; Zhou, Z.; Neher, E.; Sakmann, B. Fractional calcium currents through recombinant GluR channels of the NMDA, AMPA and kainate receptor subtypes. J. Physiol. 1995, 485 Pt 2, 403–418. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, L.; Chetkovich, D.M.; Petralia, R.S.; Sweeney, N.T.; Kawasaki, Y.; Wenthold, R.J.; Bredt, D.S.; Nicoll, R.A. Stargazin regulates synaptic targeting of AMPA receptors by two distinct mechanisms. Nature 2000, 408, 936–943. [Google Scholar] [CrossRef] [PubMed]
- McGee, T.P.; Bats, C.; Farrant, M.; Cull-Candy, S.G. Auxiliary Subunit GSG1L Acts to Suppress Calcium-Permeable AMPA Receptor Function. J. Neurosci. 2015, 35, 16171–16179. [Google Scholar] [CrossRef]
- Wang, R.; Walker, C.S.; Brockie, P.J.; Francis, M.M.; Mellem, J.E.; Madsen, D.M.; Maricq, A.V. Evolutionary conserved role for TARPs in the gating of glutamate receptors and tuning of synaptic function. Neuron 2008, 59, 997–1008. [Google Scholar] [CrossRef] [Green Version]
- Zhang, W.; St-Gelais, F.; Grabner, C.P.; Trinidad, J.C.; Sumioka, A.; Morimoto-Tomita, M.; Kim, K.S.; Straub, C.; Burlingame, A.L.; Howe, J.R.; et al. A transmembrane accessory subunit that modulates kainate-type glutamate receptors. Neuron 2009, 61, 385–396. [Google Scholar] [CrossRef] [Green Version]
- Copits, B.A.; Robbins, J.S.; Frausto, S.; Swanson, G.T. Synaptic targeting and functional modulation of GluK1 kainate receptors by the auxiliary neuropilin and tolloid-like (NETO) proteins. J. Neurosci. 2011, 31, 7334–7340. [Google Scholar] [CrossRef]
- Straub, C.; Hunt, D.L.; Yamasaki, M.; Kim, K.S.; Watanabe, M.; Castillo, P.E.; Tomita, S. Distinct functions of kainate receptors in the brain are determined by the auxiliary subunit Neto1. Nat. Neurosci. 2011, 14, 866–873. [Google Scholar] [CrossRef]
- Tang, M.; Pelkey, K.A.; Ng, D.; Ivakine, E.; McBain, C.J.; Salter, M.W.; McInnes, R.R. Neto1 is an auxiliary subunit of native synaptic kainate receptors. J. Neurosci. 2011, 31, 10009–10018. [Google Scholar] [CrossRef] [Green Version]
- Tomita, S.; Castillo, P.E. Neto1 and Neto2: Auxiliary subunits that determine key properties of native kainate receptors. J. Physiol. 2012, 590, 2217–2223. [Google Scholar] [CrossRef] [Green Version]
- Jackson, A.C.; Nicoll, R.A. The expanding social network of ionotropic glutamate receptors: TARPs and other transmembrane auxiliary subunits. Neuron 2011, 70, 178–199. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Haering, S.C.; Tapken, D.; Pahl, S.; Hollmann, M. Auxiliary subunits: Shepherding AMPA receptors to the plasma membrane. Membranes 2014, 4, 469–490. [Google Scholar] [CrossRef] [PubMed]
- Greger, I.H.; Watson, J.F.; Cull-Candy, S.G. Structural and Functional Architecture of AMPA-Type Glutamate Receptors and Their Auxiliary Proteins. Neuron 2017, 94, 713–730. [Google Scholar] [CrossRef] [PubMed]
- Howe, J.R. Modulation of non-NMDA receptor gating by auxiliary subunits. J. Physiol. 2015, 593, 61–72. [Google Scholar] [CrossRef] [Green Version]
- Copits, B.A.; Swanson, G.T. Dancing partners at the synapse: Auxiliary subunits that shape kainate receptor function. Nat. Rev. Neurosci. 2012, 13, 675–686. [Google Scholar] [CrossRef]
- Bissen, D.; Foss, F.; Acker-Palmer, A. AMPA receptors and their minions: Auxiliary proteins in AMPA receptor trafficking. Cell Mol. Life Sci. 2019, 76, 2133–2169. [Google Scholar] [CrossRef] [Green Version]
- Bowie, D. Polyamine-mediated channel block of ionotropic glutamate receptors and its regulation by auxiliary proteins. J. Biol. Chem. 2018, 293, 18789–18802. [Google Scholar] [CrossRef] [Green Version]
- Klaassen, R.V.; Stroeder, J.; Coussen, F.; Hafner, A.-S.; Petersen, J.D.; Renancio, C.; Schmitz, L.J.; Normand, E.; Lodder, J.C.; Rotaru, D.C.; et al. Shisa6 traps AMPA receptors at postsynaptic sites and prevents their desensitization during synaptic activity. Nat. Commun. 2016, 7, 10682. [Google Scholar] [CrossRef] [Green Version]
- Soto, D.; Coombs, I.D.; Gratacòs-Batlle, E.; Farrant, M.; Cull-Candy, S.G. Molecular mechanisms contributing to TARP regulation of channel conductance and polyamine block of calcium-permeable AMPA receptors. J. Neurosci. 2014, 34, 11673–11683. [Google Scholar] [CrossRef] [Green Version]
- Ben-Yaacov, A.; Gillor, M.; Haham, T.; Parsai, A.; Qneibi, M.; Stern-Bach, Y. Molecular mechanism of AMPA receptor modulation by tarp/stargazin. Neuron 2017, 93, 1126–1137.e4. [Google Scholar] [CrossRef] [Green Version]
- Coombs, I.D.; Soto, D.; Zonouzi, M.; Renzi, M.; Shelley, C.; Farrant, M.; Cull-Candy, S.G. Cornichons modify channel properties of recombinant and glial AMPA receptors. J. Neurosci. 2012, 32, 9796–9804. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Coombs, I.D.; MacLean, D.M.; Jayaraman, V.; Farrant, M.; Cull-Candy, S.G. Dual Effects of TARP γ-2 on Glutamate Efficacy Can Account for AMPA Receptor Autoinactivation. Cell Rep. 2017, 20, 1123–1135. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, S.; Zhao, Y.; Wang, Y.; Shekhar, M.; Tajkhorshid, E.; Gouaux, E. Activation and Desensitization Mechanism of AMPA Receptor-TARP Complex by Cryo-EM. Cell 2017, 170, 1234–1246.e14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Twomey, E.C.; Yelshanskaya, M.V.; Grassucci, R.A.; Frank, J.; Sobolevsky, A.I. Elucidation of AMPA receptor-stargazin complexes by cryo-electron microscopy. Science 2016, 353, 83–86. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jackson, A.C.; Milstein, A.D.; Soto, D.; Farrant, M.; Cull-Candy, S.G.; Nicoll, R.A. Probing TARP modulation of AMPA receptor conductance with polyamine toxins. J. Neurosci. 2011, 31, 7511–7520. [Google Scholar] [CrossRef]
- Soto, D.; Coombs, I.D.; Kelly, L.; Farrant, M.; Cull-Candy, S.G. Stargazin attenuates intracellular polyamine block of calcium-permeable AMPA receptors. Nat. Neurosci. 2007, 10, 1260–1267. [Google Scholar] [CrossRef]
- Fisher, J.L.; Mott, D.D. The auxiliary subunits Neto1 and Neto2 reduce voltage-dependent inhibition of recombinant kainate receptors. J. Neurosci. 2012, 32, 12928–12933. [Google Scholar] [CrossRef] [Green Version]
- Shelley, C.; Farrant, M.; Cull-Candy, S.G. TARP-associated AMPA receptors display an increased maximum channel conductance and multiple kinetically distinct open states. J. Physiol. 2012, 590, 5723–5738. [Google Scholar] [CrossRef]
- Rozov, A.; Zakharova, Y.; Vazetdinova, A.; Valiullina-Rakhmatullina, F. The Role of Polyamine-Dependent Facilitation of Calcium Permeable AMPARs in Short-Term Synaptic Enhancement. Front. Cell Neurosci. 2018, 12, 345. [Google Scholar] [CrossRef]
- Baukrowitz, T.; Yellen, G. Use-dependent blockers and exit rate of the last ion from the multi-ion pore of a K+ channel. Science 1996, 271, 653–656. [Google Scholar] [CrossRef] [Green Version]
- Weiss, J.H. Ca permeable AMPA channels in diseases of the nervous system. Front. Mol. Neurosci. 2011, 4, 42. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wright, A.; Vissel, B. The essential role of AMPA receptor GluR2 subunit RNA editing in the normal and diseased brain. Front. Mol. Neurosci. 2012, 5, 34. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lerma, J.; Marques, J.M. Kainate receptors in health and disease. Neuron 2013, 80, 292–311. [Google Scholar] [CrossRef] [Green Version]
- Deng, W.; Rosenberg, P.A.; Volpe, J.J.; Jensen, F.E. Calcium-permeable AMPA/kainate receptors mediate toxicity and preconditioning by oxygen-glucose deprivation in oligodendrocyte precursors. Proc. Natl. Acad. Sci. USA 2003, 100, 6801–6806. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Johnson, J.W.; Ascher, P. Glycine potentiates the NMDA response in cultured mouse brain neurons. Nature 1987, 325, 529–531. [Google Scholar] [CrossRef]
- Nowak, L.; Bregestovski, P.; Ascher, P.; Herbet, A.; Prochiantz, A. Magnesium gates glutamate-activated channels in mouse central neurones. Nature 1984, 307, 462–465. [Google Scholar] [CrossRef]
- Mayer, M.L.; Westbrook, G.L.; Guthrie, P.B. Voltage-dependent block by Mg2+ of NMDA responses in spinal cord neurones. Nature 1984, 309, 261–263. [Google Scholar] [CrossRef]
- Mayer, M.L.; MacDermott, A.B.; Westbrook, G.L.; Smith, S.J.; Barker, J.L. Agonist- and voltage-gated calcium entry in cultured mouse spinal cord neurons under voltage clamp measured using arsenazo III. J. Neurosci. 1987, 7, 3230–3244. [Google Scholar] [CrossRef] [Green Version]
- Vicini, S.; Wang, J.F.; Li, J.H.; Zhu, W.J.; Wang, Y.H.; Luo, J.H.; Wolfe, B.B.; Grayson, D.R. Functional and pharmacological differences between recombinant N-methyl-D-aspartate receptors. J. Neurophysiol. 1998, 79, 555–566. [Google Scholar] [CrossRef] [Green Version]
- Wyllie, D.J.; Béhé, P.; Colquhoun, D. Single-channel activations and concentration jumps: Comparison of recombinant NR1a/NR2A and NR1a/NR2D NMDA receptors. J. Physiol. 1998, 510 Pt 1, 1–18. [Google Scholar] [CrossRef]
- Malenka, R.C.; Bear, M.F. LTP and LTD: An embarrassment of riches. Neuron 2004, 44, 5–21. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Akgül, G.; McBain, C.J. Diverse roles for ionotropic glutamate receptors on inhibitory interneurons in developing and adult brain. J. Physiol. 2016, 594, 5471–5490. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sheng, M.; Cummings, J.; Roldan, L.A.; Jan, Y.N.; Jan, L.Y. Changing subunit composition of heteromeric NMDA receptors during development of rat cortex. Nature 1994, 368, 144–147. [Google Scholar] [CrossRef] [PubMed]
- Grand, T.; Gerges, S.A.; David, M.; Diana, M.A.; Paoletti, P. Unmasking GluN1/GluN3A excitatory glycine NMDA receptors. Nat. Commun. 2018, 9, 4769. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mothet, J.P.; Parent, A.T.; Wolosker, H.; Brady, R.O.; Linden, D.J.; Ferris, C.D.; Rogawski, M.A.; Snyder, S.H. D-serine is an endogenous ligand for the glycine site of the N-methyl-D-aspartate receptor. Proc. Natl. Acad. Sci. USA 2000, 97, 4926–4931. [Google Scholar] [CrossRef] [Green Version]
- Benveniste, M.; Mayer, M.L. Kinetic analysis of antagonist action at N-methyl-D-aspartic acid receptors. Two binding sites each for glutamate and glycine. Biophys. J. 1991, 59, 560–573. [Google Scholar] [CrossRef] [Green Version]
- Clements, J.D.; Westbrook, G.L. Activation kinetics reveal the number of glutamate and glycine binding sites on the N-methyl-D-aspartate receptor. Neuron 1991, 7, 605–613. [Google Scholar] [CrossRef]
- Schorge, S.; Elenes, S.; Colquhoun, D. Maximum likelihood fitting of single channel NMDA activity with a mechanism composed of independent dimers of subunits. J. Physiol. 2005, 569, 395–418. [Google Scholar] [CrossRef]
- Gilling, K.E.; Jatzke, C.; Hechenberger, M.; Parsons, C.G. Potency, voltage-dependency, agonist concentration-dependency, blocking kinetics and partial untrapping of the uncompetitive N-methyl-D-aspartate (NMDA) channel blocker memantine at human NMDA (GluN1/GluN2A) receptors. Neuropharmacology 2009, 56, 866–875. [Google Scholar] [CrossRef]
- Parsons, C.G.; Quack, G.; Bresink, I.; Baran, L.; Przegalinski, E.; Kostowski, W.; Krzascik, P.; Hartmann, S.; Danysz, W. Comparison of the potency, kinetics and voltage-dependency of a series of uncompetitive NMDA receptor antagonists in vitro with anticonvulsive and motor impairment activity in vivo. Neuropharmacology 1995, 34, 1239–1258. [Google Scholar] [CrossRef]
- MacDonald, J.F.; Bartlett, M.C.; Mody, I.; Pahapill, P.; Reynolds, J.N.; Salter, M.W.; Schneiderman, J.H.; Pennefather, P.S. Actions of ketamine, phencyclidine and MK-801 on NMDA receptor currents in cultured mouse hippocampal neurones. J. Physiol. 1991, 432, 483–508. [Google Scholar] [CrossRef] [PubMed]
- Antonov, S.M.; Johnson, J.W. Voltage-dependent interaction of open-channel blocking molecules with gating of NMDA receptors in rat cortical neurons. J. Physiol. 1996, 493 Pt 2, 425–445. [Google Scholar] [CrossRef] [PubMed]
- Krystal, J.H.; Karper, L.P.; Seibyl, J.P.; Freeman, G.K.; Delaney, R.; Bremner, J.D.; Heninger, G.R.; Bowers, M.B.; Charney, D.S. Subanesthetic effects of the noncompetitive NMDA antagonist, ketamine, in humans. Psychotomimetic, perceptual, cognitive, and neuroendocrine responses. Arch. Gen. Psychiatry 1994, 51, 199–214. [Google Scholar] [CrossRef] [PubMed]
- Persson, J. Ketamine in pain management. CNS Neurosci. Ther. 2013, 19, 396–402. [Google Scholar] [CrossRef]
- Zhou, H.-Y.; Chen, S.-R.; Pan, H.-L. Targeting N-methyl-D-aspartate receptors for treatment of neuropathic pain. Expert Rev. Clin. Pharmacol. 2011, 4, 379–388. [Google Scholar] [CrossRef] [Green Version]
- Kafi, H.; Salamzadeh, J.; Beladimoghadam, N.; Sistanizad, M.; Kouchek, M. Study of the neuroprotective effects of memantine in patients with mild to moderate ischemic stroke. Iran. J. Pharm. Res. 2014, 13, 591–598. [Google Scholar]
- Danysz, W.; Parsons, C.G. Alzheimer’s disease, β-amyloid, glutamate, NMDA receptors and memantine—Searching for the connections. Br. J. Pharmacol. 2012, 167, 324–352. [Google Scholar] [CrossRef] [Green Version]
- Parsons, C.G.; Danysz, W.; Quack, G. Memantine is a clinically well tolerated N-methyl-D-aspartate (NMDA) receptor antagonist—A review of preclinical data. Neuropharmacology 1999, 38, 735–767. [Google Scholar] [CrossRef]
- Abdallah, C.G.; Averill, L.A.; Krystal, J.H. Ketamine as a promising prototype for a new generation of rapid-acting antidepressants. Ann. N. Y. Acad. Sci. 2015, 1344, 66–77. [Google Scholar] [CrossRef] [Green Version]
- Kong, M.; Ba, M.; Ren, C.; Yu, L.; Dong, S.; Yu, G.; Liang, H. An updated meta-analysis of amantadine for treating dyskinesia in Parkinson’s disease. Oncotarget 2017, 8, 57316–57326. [Google Scholar] [CrossRef] [Green Version]
- Nair, A.S.; Sahoo, R.K. Efficacy of memantine hydrochloride in neuropathic pain. Indian J. Palliat. Care 2019, 25, 161–162. [Google Scholar] [CrossRef]
- Kashiwagi, K.; Masuko, T.; Nguyen, C.D.; Kuno, T.; Tanaka, I.; Igarashi, K.; Williams, K. Channel blockers acting at N-methyl-D-aspartate receptors: Differential effects of mutations in the vestibule and ion channel pore. Mol. Pharmacol. 2002, 61, 533–545. [Google Scholar] [CrossRef] [Green Version]
- Miller, O.H.; Yang, L.; Wang, C.-C.; Hargroder, E.A.; Zhang, Y.; Delpire, E.; Hall, B.J. GluN2B-containing NMDA receptors regulate depression-like behavior and are critical for the rapid antidepressant actions of ketamine. eLife 2014, 3, e03581. [Google Scholar] [CrossRef]
- Noppers, I.; Niesters, M.; Aarts, L.; Smith, T.; Sarton, E.; Dahan, A. Ketamine for the treatment of chronic non-cancer pain. Expert Opin. Pharmacother. 2010, 11, 2417–2429. [Google Scholar] [CrossRef]
- Amidfar, M.; Réus, G.Z.; Quevedo, J.; Kim, Y.-K. The role of memantine in the treatment of major depressive disorder: Clinical efficacy and mechanisms of action. Eur. J. Pharmacol. 2018, 827, 103–111. [Google Scholar] [CrossRef]
- Gideons, E.S.; Kavalali, E.T.; Monteggia, L.M. Mechanisms underlying differential effectiveness of memantine and ketamine in rapid antidepressant responses. Proc. Natl. Acad. Sci. USA 2014, 111, 8649–8654. [Google Scholar] [CrossRef] [Green Version]
- Chen, H.-S.V.; Lipton, S.A. The chemical biology of clinically tolerated NMDA receptor antagonists. J. Neurochem. 2006, 97, 1611–1626. [Google Scholar] [CrossRef]
- Parsons, C.G.; Stöffler, A.; Danysz, W. Memantine: A NMDA receptor antagonist that improves memory by restoration of homeostasis in the glutamatergic system—Too little activation is bad, too much is even worse. Neuropharmacology 2007, 53, 699–723. [Google Scholar] [CrossRef]
- Lipton, S.A. Paradigm shift in neuroprotection by NMDA receptor blockade: Memantine and beyond. Nat. Rev. Drug Discov. 2006, 5, 160–170. [Google Scholar] [CrossRef]
- Johnson, J.W.; Qian, A. Interaction between channel blockers and channel gating of NMDA receptors. Biol. Membr. 2002, 19, 110–115. [Google Scholar]
- Sobolevsky, A.I.; Koshelev, S.G.; Khodorov, B.I. Interaction of memantine and amantadine with agonist-unbound NMDA-receptor channels in acutely isolated rat hippocampal neurons. J. Physiol. 1998, 512 Pt 1, 47–60. [Google Scholar] [CrossRef]
- Dilmore, J.G.; Johnson, J.W. Open channel block and alteration of N-methyl-D-aspartic acid receptor gating by an analog of phencyclidine. Biophys. J. 1998, 75, 1801–1816. [Google Scholar] [CrossRef] [Green Version]
- Wright, J.M.; Nowak, L.M. Effects of low doses of bicuculline on N-methyl-D-aspartate single-channel kinetics are not evident in whole-cell currents. Mol. Pharmacol. 1992, 41, 900–907. [Google Scholar]
- Sobolevsky, A.I. Quantitative analysis of tetrapentylammonium-induced blockade of open N-methyl-D-aspartate channels. Biophys. J. 2000, 79, 1324–1335. [Google Scholar] [CrossRef] [Green Version]
- Antonov, S.M.; Johnson, J.W.; Lukomskaya, N.Y.; Potapyeva, N.N.; Gmiro, V.E.; Magazanik, L.G. Novel adamantane derivatives act as blockers of open ligand-gated channels and as anticonvulsants. Mol. Pharmacol. 1995, 47, 558–567. [Google Scholar]
- Chen, H.S.; Lipton, S.A. Mechanism of memantine block of NMDA-activated channels in rat retinal ganglion cells: Uncompetitive antagonism. J. Physiol. 1997, 499 Pt 1, 27–46. [Google Scholar] [CrossRef]
- Costa, A.C.; Albuquerque, E.X. Dynamics of the actions of tetrahydro-9-aminoacridine and 9-aminoacridine on glutamatergic currents: Concentration-jump studies in cultured rat hippocampal neurons. J. Pharmacol. Exp. Ther. 1994, 268, 503–514. [Google Scholar]
- Vorobjev, V.S.; Sharonova, I.N. Tetrahydroaminoacridine blocks and prolongs NMDA receptor-mediated responses in a voltage-dependent manner. Eur. J. Pharmacol. 1994, 253, 1–8. [Google Scholar] [CrossRef]
- Li-Smerin, Y.; Johnson, J.W. Effects of intracellular Mg2+ on channel gating and steady-state responses of the NMDA receptor in cultured rat neurons. J. Physiol. 1996, 491 Pt 1, 137–150. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Koshelev, S.G.; Khodorov, B.I. Blockade of open NMDA channel by tetrabutylammonium, 9-aminoacridine and tacrine prevents channels closing and desensitization. Membr. Cell Biol. C/C Biol. Membr. 1995, 9, 93–110. [Google Scholar]
- Qian, A.; Antonov, S.M.; Johnson, J.W. Modulation by permeant ions of Mg2+ inhibition of NMDA-activated whole-cell currents in rat cortical neurons. J. Physiol. 2002, 538, 65–77. [Google Scholar] [CrossRef] [Green Version]
- Kotermanski, S.E.; Johnson, J.W. Mg2+ imparts NMDA receptor subtype selectivity to the Alzheimer’s drug memantine. J. Neurosci. 2009, 29, 2774–2779. [Google Scholar] [CrossRef]
- Mealing, G.A.; Lanthorn, T.H.; Murray, C.L.; Small, D.L.; Morley, P. Differences in degree of trapping of low-affinity uncompetitive N-methyl-D-aspartic acid receptor antagonists with similar kinetics of block. J. Pharmacol. Exp. Ther. 1999, 288, 204–210. [Google Scholar]
- Antonov, S.M.; Gmiro, V.E.; Johnson, J.W. Binding sites for permeant ions in the channel of NMDA receptors and their effects on channel block. Nat. Neurosci. 1998, 1, 451–461. [Google Scholar] [CrossRef]
- Qian, A.; Johnson, J.W. Channel gating of NMDA receptors. Physiol. Behav. 2002, 77, 577–582. [Google Scholar] [CrossRef]
- Tajima, N.; Karakas, E.; Grant, T.; Simorowski, N.; Diaz-Avalos, R.; Grigorieff, N. Activation of NMDA receptors and the mechanism of inhibition by ifenprodil. Nature 2016, 534, 63–68. [Google Scholar] [CrossRef] [Green Version]
- Glasgow, N.G.; Wilcox, M.R.; Johnson, J.W. Effects of Mg2+ on recovery of NMDA receptors from inhibition by memantine and ketamine reveal properties of a second site. Neuropharmacology 2018, 137, 344–358. [Google Scholar] [CrossRef]
- Wilcox, M.R.; Glasgow, N.G.; Mesbahi-Vasey, S.; Nigam, A.; Phillips, M.B.; Turcu, A.L.; Narangoda, C.; Kurnikova, M.G.; Vazquez, S.; Johnson, J.W. A Hydrophobic Path Allows Drug Access to the NMDA Receptor Channel. Program No. 370.16.; 2018 Neuroscience Meeting Planner; Society for Neuroscience: San Diego, CA, USA, 2018. [Google Scholar]
- Sobolevsky, A.; Koshelev, S. Two blocking sites of amino-adamantane derivatives in open N-methyl-D-aspartate channels. Biophys. J. 1998, 74, 1305–1319. [Google Scholar] [CrossRef] [Green Version]
- Maki, B.A.; Aman, T.K.; Amico-Ruvio, S.A.; Kussius, C.L.; Popescu, G.K. C-terminal domains of N-methyl-D-aspartic acid receptor modulate unitary channel conductance and gating. J. Biol. Chem. 2012, 287, 36071–36080. [Google Scholar] [CrossRef] [Green Version]
- Krupp, J.J.; Vissel, B.; Heinemann, S.F.; Westbrook, G.L. N-terminal domains in the NR2 subunit control desensitization of NMDA receptors. Neuron 1998, 20, 317–327. [Google Scholar] [CrossRef] [Green Version]
- Krupp, J.J.; Vissel, B.; Thomas, C.G.; Heinemann, S.F.; Westbrook, G.L. Calcineurin acts via the C-terminus of NR2A to modulate desensitization of NMDA receptors. Neuropharmacology 2002, 42, 593–602. [Google Scholar] [CrossRef]
- Villarroel, A.; Regalado, M.P.; Lerma, J. Glycine-independent NMDA receptor desensitization: Localization of structural determinants. Neuron 1998, 20, 329–339. [Google Scholar] [CrossRef]
- Ehlers, M.D.; Zhang, S.; Bernhadt, J.P.; Huganir, R.L. Inactivation of NMDA receptors by direct interaction of calmodulin with the NR1 subunit. Cell 1996, 84, 745–755. [Google Scholar] [CrossRef] [Green Version]
- Rothman, S.M.; Olney, J.W. Excitotoxicity and the NMDA receptor—Still lethal after eight years. Trends Neurosci. 1995, 18, 57–58. [Google Scholar] [CrossRef]
- Okamoto, S.; Pouladi, M.A.; Talantova, M.; Yao, D.; Xia, P.; Ehrnhoefer, D.E.; Zaidi, R.; Clemente, A.; Kaul, M.; Graham, R.K.; et al. Balance between synaptic versus extrasynaptic NMDA receptor activity influences inclusions and neurotoxicity of mutant huntingtin. Nat. Med. 2009, 15, 1407–1413. [Google Scholar] [CrossRef] [Green Version]
- Hardingham, G.E.; Bading, H. Synaptic versus extrasynaptic NMDA receptor signalling: Implications for neurodegenerative disorders. Nat. Rev. Neurosci. 2010, 11, 682–696. [Google Scholar] [CrossRef] [Green Version]
- Ascher, P.; Nowak, L. The role of divalent cations in the N-methyl-D-aspartate responses of mouse central neurones in culture. J. Physiol. 1988, 399, 247–266. [Google Scholar] [CrossRef]
- Clarke, R.J.; Johnson, J.W. NMDA receptor NR2 subunit dependence of the slow component of magnesium unblock. J. Neurosci. 2006, 26, 5825–5834. [Google Scholar] [CrossRef]
- Clarke, R.J.; Glasgow, N.G.; Johnson, J.W. Mechanistic and structural determinants of NMDA receptor voltage-dependent gating and slow Mg2+ unblock. J. Neurosci. 2013, 33, 4140–4150. [Google Scholar] [CrossRef] [Green Version]
- Vargas-Caballero, M.; Robinson, H.P.C. A slow fraction of Mg2+ unblock of NMDA receptors limits their contribution to spike generation in cortical pyramidal neurons. J. Neurophysiol. 2003, 89, 2778–2783. [Google Scholar] [CrossRef]
- Spruston, N.; Jonas, P.; Sakmann, B. Dendritic glutamate receptor channels in rat hippocampal CA3 and CA1 pyramidal neurons. J. Physiol. 1995, 482 Pt 2, 325–352. [Google Scholar] [CrossRef]
- Kampa, B.M.; Clements, J.; Jonas, P.; Stuart, G.J. Kinetics of Mg2+ unblock of NMDA receptors: Implications for spike-timing dependent synaptic plasticity. J. Physiol. 2004, 556, 337–345. [Google Scholar] [CrossRef]
- Clarke, R.J.; Johnson, J.W. Voltage-dependent gating of NR1/2B NMDA receptors. J. Physiol. 2008, 586, 5727–5741. [Google Scholar] [CrossRef]


Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Phillips, M.B.; Nigam, A.; Johnson, J.W. Interplay between Gating and Block of Ligand-Gated Ion Channels. Brain Sci. 2020, 10, 928. https://doi.org/10.3390/brainsci10120928
Phillips MB, Nigam A, Johnson JW. Interplay between Gating and Block of Ligand-Gated Ion Channels. Brain Sciences. 2020; 10(12):928. https://doi.org/10.3390/brainsci10120928
Chicago/Turabian StylePhillips, Matthew B., Aparna Nigam, and Jon W. Johnson. 2020. "Interplay between Gating and Block of Ligand-Gated Ion Channels" Brain Sciences 10, no. 12: 928. https://doi.org/10.3390/brainsci10120928
APA StylePhillips, M. B., Nigam, A., & Johnson, J. W. (2020). Interplay between Gating and Block of Ligand-Gated Ion Channels. Brain Sciences, 10(12), 928. https://doi.org/10.3390/brainsci10120928