Functional and Dysfunctional Neuroplasticity in Learning to Cope with Stress

 ,
, 
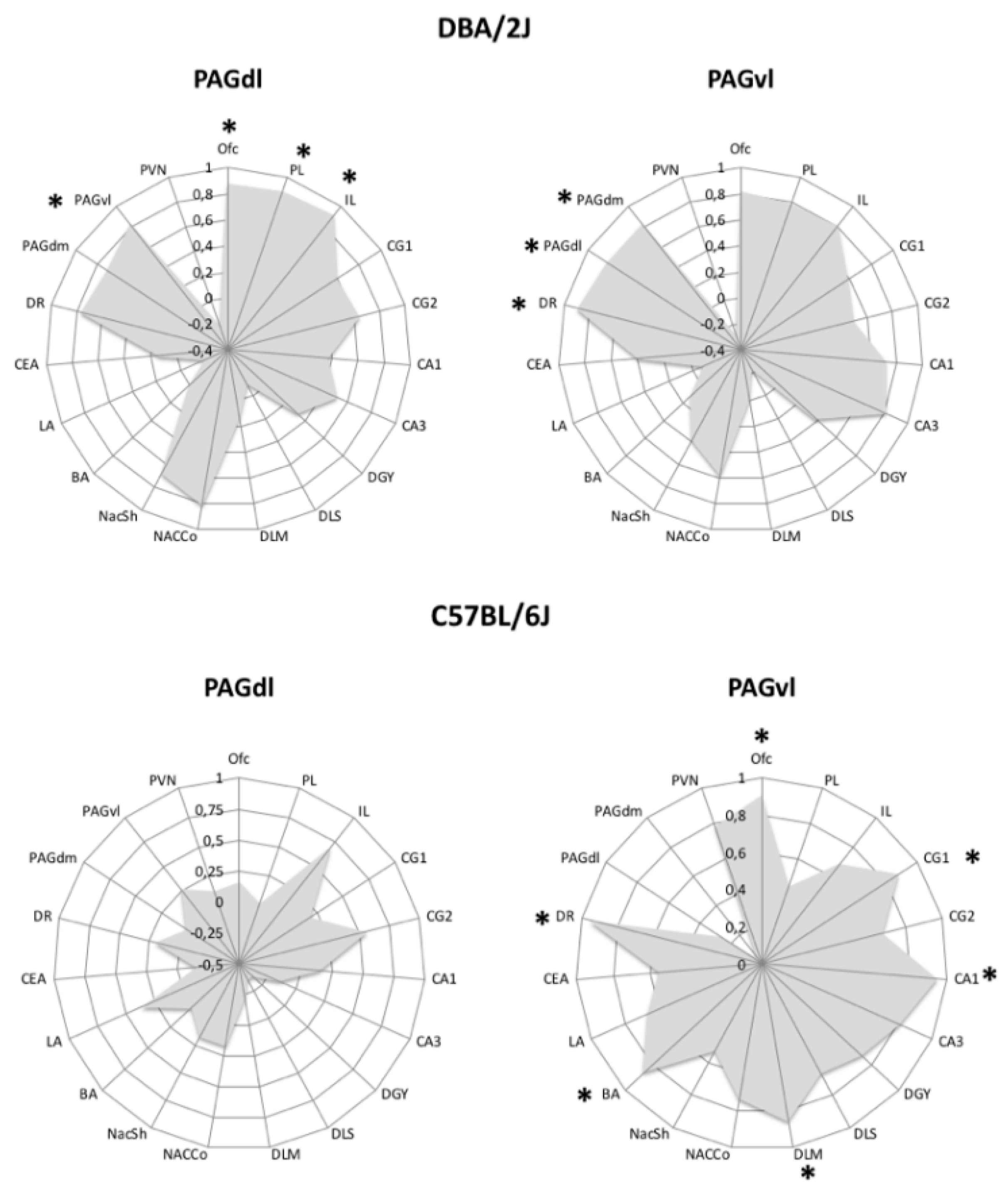{kind=link}
Abstract
:1. Introduction
2. Adaptive Coping and Functional Neuroplasticity
3. Maladaptive Coping
4. Stress-Induced Dysfunctional Neuroplasticity and Psychopathology
5. Conclusions
Conflicts of Interest
References
- Schwabe, L. Memory under stress: From single systems to network changes. Eur. J. Neurosci. 2016, 45, 478–489. [Google Scholar] [CrossRef] [PubMed]
- Morris, S.E.; Rumsey, J.M.; Cuthbert, B.N. Rethinking mental disorders: The role of learning and brain plasticity. Restor. Neurol. Neurosci. 2014, 32, 5–23. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cabib, S.; Paolo, C.; Colelli, V. Learning to cope with stress: Psychobiological mechanisms of stress resilience. Rev. Neurosci. 2012, 23, 659–672. [Google Scholar] [CrossRef] [PubMed]
- Gillan, C.; Robbins, T.; Sahakian, B.; Heuvel, O.A.V.D.; Van Wingen, G. The role of habit in compulsivity. Eur. Neuropsychopharmacol. 2015, 26, 828–840. [Google Scholar] [CrossRef] [PubMed]
- Robbins, T. Limbic-Striatal Memory Systems and Drug Addiction. Neurobiol. Learn. Mem. 2002, 78, 625–636. [Google Scholar] [CrossRef] [PubMed]
- Cantone, M.; Bramanti, A.; Lanza, G.; Pennisi, M.; Bramanti, P.; Pennisi, G.; Bella, R. Cortical Plasticity in Depression: A Neurochemical Perspective From Transcranial Magnetic Stimulation. ASN Neuro 2017, 9. [Google Scholar] [CrossRef] [Green Version]
- Hays, S.A.; Rennaker, R.L.; Kilgard, M. Targeting Plasticity with Vagus Nerve Stimulation to Treat Neurological Disease; Elsevie: Amsterdam, The Netherlands, 2013; Volume 207, pp. 275–299. [Google Scholar]
- Pedroarena-Leal, N.; Heidemeyer, L.; Trenado, C.; Ruge, D. Human Depotentiation following Induction of Spike Timing Dependent Plasticity. Biomedicines 2018, 6, 71. [Google Scholar] [CrossRef] [Green Version]
- Peña, D.F.; Engineer, N.D.; McIntyre, C.K. Rapid remission of conditioned fear expression with extinction training paired with vagus nerve stimulation. Boil. Psychiatry 2012, 73, 1071–1077. [Google Scholar] [CrossRef] [Green Version]
- Réus, G.Z.; De Moura, A.B.; Borba, L.A.; Abelaira, H.M.; Quevedo, J. Strategies for Treatment-Resistant Depression: Lessons Learned from Animal Models. Mol. Neuropsychiatry 2019, 5, 178–189. [Google Scholar] [CrossRef]
- Folkman, S.; Lazarus, R.S.; Gruen, R.J.; DeLongis, A. Appraisal, coping, health status, and psychological symptoms. J. Personal. Soc. Psychol. 1986, 50, 571–579. [Google Scholar] [CrossRef]
- Lazarus, R.S. Toward better research on stress and coping. Am. Psychol. 2000, 55, 665–673. [Google Scholar] [CrossRef] [PubMed]
- Koolhaas, J.; Korte, S.M.; De Boer, S.; Van Der Vegt, B.; Van Reenen, C.; Hopster, H.; De Jong, I.; Ruis, M.; Blokhuis, H. Coping styles in animals: Current status in behavior and stress-physiology. Neurosci. Biobehav. Rev. 1999, 23, 925–935. [Google Scholar] [CrossRef]
- Taylor, S.E.; Stanton, A.L. Coping Resources, Coping Processes, and Mental Health. Annu. Rev. Clin. Psychol. 2007, 3, 377–401. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cabib, S.; Puglisi-Allegra, S. The mesoaccumbens dopamine in coping with stress. Neurosci. Biobehav. Rev. 2012, 36, 79–89. [Google Scholar] [CrossRef]
- McEwen, B.S.; Gianaros, P.J. Stress- and allostasis-induced brain plasticity. Annu. Rev. Med. 2011, 62, 431–445. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McEwen, B.S. Central effects of stress hormones in health and disease: Understanding the protective and damaging effects of stress and stress mediators. Eur. J. Pharmacol. 2008, 583, 174–185. [Google Scholar] [CrossRef] [Green Version]
- Drugan, R.C.; Eren, S.; Hazi, A.; Silva, J.; Christianson, J.P.; Kent, S. Impact of water temperature and stressor controllability on swim stress-induced changes in body temperature, serum corticosterone, and immobility in rats. Pharmacol. Biochem. Behav. 2005, 82, 397–403. [Google Scholar] [CrossRef]
- Cabib, S.; Puglisi-Allegra, S. Opposite responses of mesolimbic dopamine system to controllable and uncontrollable aversive experiences. J. Neurosci. 1994, 14, 3333–3340. [Google Scholar] [CrossRef] [Green Version]
- Maier, S.F.; Watkins, L.R. Stressor controllability and learned helplessness: The roles of the dorsal raphe nucleus, serotonin, and corticotropin-releasing factor. Neurosci. Biobehav. Rev. 2005, 29, 829–841. [Google Scholar] [CrossRef]
- Maier, S.F.; Watkins, L.R. Role of the medial prefrontal cortex in coping and resilience. Brain Res. 2010, 1355, 52–60. [Google Scholar] [CrossRef] [Green Version]
- Maia, T. Two-factor theory, the actor—Critic model, and conditioned avoidance. Learn. Behav. 2010, 38, 50–67. [Google Scholar] [CrossRef] [PubMed]
- LeDoux, J.E.; Moscarello, J.; Sears, R.; Campese, V. The birth, death and resurrection of avoidance: A reconceptualization of a troubled paradigm. Mol. Psychiatry 2016, 22, 24–36. [Google Scholar] [CrossRef] [PubMed]
- Moscarello, J.; Maren, S. Flexibility in the face of fear: Hippocampal-prefrontal regulation of fear and avoidance. Curr. Opin. Behav. Sci. 2017, 19, 44–49. [Google Scholar] [CrossRef] [PubMed]
- Wendler, E.; Gaspar, J.C.; Ferreira, T.; Barbiero, J.K.; Andreatini, R.; Vital, M.A.; Blaha, C.; Winn, P.; Da Cunha, C. The roles of the nucleus accumbens core, dorsomedial striatum, and dorsolateral striatum in learning: Performance and extinction of Pavlovian fear-conditioned responses and instrumental avoidance responses. Neurobiol. Learn. Mem. 2014, 109, 27–36. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Diehl, M.M.; Bravo-Rivera, C.; Quirk, G.J. The study of active avoidance: A platform for discussion. Neurosci. Biobehav. Rev. 2019, 107, 229–237. [Google Scholar] [CrossRef]
- Tremblay, L.; Worbe, Y.; Thobois, S.; Sgambato-Faure, V.; Féger, J. Selective dysfunction of basal ganglia subterritories: From movement to behavioral disorders. Mov. Disord. 2015, 30, 1155–1170. [Google Scholar] [CrossRef]
- Martínez-Rivera, F.J.; Bravo-Rivera, C.; Velázquez-Díaz, C.D.; Montesinos-Cartagena, M.; Quirk, G.J. Prefrontal circuits signaling active avoidance retrieval and extinction. Psychopharmacology 2018, 236, 399–406. [Google Scholar] [CrossRef]
- Gröger, N.; Mannewitz, A.; Bock, J.; Becker, S.; Guttmann, K.; Poeggel, G.; Braun, K. Infant avoidance training alters cellular activation patterns in prefronto-limbic circuits during adult avoidance learning: II. Cellular imaging of neurons expressing the activity-regulated cytoskeleton-associated protein (Arc/Arg3.1). Brain Struct. Funct. 2017, 223, 713–725. [Google Scholar] [CrossRef]
- Rosas-Vidal, L.; Lozada-Miranda, V.; Cantres-Rosario, Y.; Vega-Medina, A.; Melendez, L.; Quirk, G.J.; Lozada, V. Alteration of BDNF in the medial prefrontal cortex and the ventral hippocampus impairs extinction of avoidance. Neuropsychopharmacology 2018, 43, 2636–2644. [Google Scholar] [CrossRef] [Green Version]
- Ramirez, F.; Moscarello, J.; LeDoux, J.E.; Sears, R.M. Active avoidance requires a serial basal amygdala to nucleus accumbens shell circuit. J. Neurosci. 2015, 35, 3470–3477. [Google Scholar] [CrossRef] [Green Version]
- Porsolt, R.D.; Le Pichon, M.; Jalfre, M. Depression: A new animal model sensitive to antidepressant treatments. Nature 1977, 266, 730–732. [Google Scholar] [CrossRef] [PubMed]
- Mitchell, J.B.; Meaney, M.J. Effects of corticosterone on response consolidation and retrieval in the forced swim test. Behav. Neurosci. 1991, 105, 798–803. [Google Scholar] [CrossRef] [PubMed]
- Colelli, V.; Campus, P.; Conversi, D.; Orsini, C.; Cabib, C. Either the dorsal hippocampus or the dorsolateral striatum is selectively involved in consolidation of forced swim-induced immobility depending on genetic background. Neurobiol. Learn. Mem. 2014, 111, 49–55. [Google Scholar] [CrossRef]
- Reul, J.M. Making Memories of Stressful Events: A Journey Along Epigenetic, Gene Transcription, and Signaling Pathways. Front. Psychol. 2014, 5, 5. [Google Scholar] [CrossRef]
- Andolina, D.; Maran, D.; Valzania, A.; Conversi, D.; Puglisi-Allegra, S. Prefrontal/Amygdalar System Determines Stress Coping Behavior Through 5-HT/GABA Connection. Neuropsychopharmacology 2013, 38, 2057–2067. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Campus, P.; Colelli, V.; Orsini, C.; Sarra, D.; Cabib, S. Evidence for the involvement of extinction-associated inhibitory learning in the forced swimming test. Behav. Brain Res. 2015, 278, 348–355. [Google Scholar] [CrossRef]
- De Kloet, E.; Molendijk, M.L. Coping with the Forced Swim Stressor: Towards Understanding an Adaptive Mechanism. Neural Plast. 2016, 2016, 1–13. [Google Scholar] [CrossRef] [Green Version]
- Molendijk, M.L.; De Kloet, E. Coping with the forced swim stressor: Current state-of-the-art. Behav. Brain Res. 2019, 364, 1–10. [Google Scholar] [CrossRef]
- West, A. Neurobehavioral studies of forced swimming: The role of learning and memory in the forced swim test. Prog. Neuro-Psychopharmacol. Boil. Psychiatry 1990, 14, 863–877. [Google Scholar] [CrossRef]
- De Kloet, E.R.; Molendijk, M.; Meijer, O.; Sb, J.; Eb, E.; Rt, L.; Rm, A.; Sa, R.-M.; Rt, L.; Ns, N.; et al. F1000Prime recommendation of Prefrontal-bed nucleus circuit modulation of a passive coping response set. F1000-Post-Publ. Peer Rev. Biomed. Lit. 2019, 39, 1405–1419. [Google Scholar]
- Warden, M.R.; Selimbeyoglu, A.; Mirzabekov, J.J.; Lo, M.; Thompson, K.R.; Kim, S.-Y.; Adhikari, A.; Tye, K.M.; Frank, L.M.; Deisseroth, K. A prefrontal cortex–brainstem neuronal projection that controls response to behavioural challenge. Nature 2012, 492, 428–432. [Google Scholar] [CrossRef]
- Andolina, D.; Puglisi-Allegra, S.; Ventura, R. Strain-dependent differences in corticolimbic processing of aversive or rewarding stimuli. Front. Syst. Neurosci. 2015, 8, 207. [Google Scholar] [CrossRef] [Green Version]
- Latagliata, E.C.; Puglisi-Allegra, S.; Ventura, R.; Cabib, S. Norepinephrine in the Medial Pre-frontal Cortex Supports Accumbens Shell Responses to a Novel Palatable Food in Food-Restricted Mice Only. Front. Behav. Neurosci. 2018, 12, 7. [Google Scholar] [CrossRef] [PubMed]
- Pascucci, T.; Ventura, R.; Latagliata, E.C.; Cabib, S.; Puglisi-Allegra, S. The Medial Prefrontal Cortex Determines the Accumbens Dopamine Response to Stress through the Opposing Influences of Norepinephrine and Dopamine. Cereb. Cortex 2007, 17, 2796–2804. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ventura, R.; Cabib, S.; Puglisi-Allegra, S. Opposite genotype-dependent mesocorticolimbic dopamine response to stress. Neuroscience 2001, 104, 627–631. [Google Scholar] [CrossRef]
- Collins, A.; Hill, L.E.; Chandramohan, Y.; Whitcomb, D.; Droste, S.K.; Reul, J.M. Exercise Improves Cognitive Responses to Psychological Stress through Enhancement of Epigenetic Mechanisms and Gene Expression in the Dentate Gyrus. PLoS ONE 2009, 4, e4330. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chandramohan, Y.; Droste, S.K.; Arthur, J.S.C.; Reul, J.M. The forced swimming-induced behavioural immobility response involves histone H3 phospho-acetylation and c-Fos induction in dentate gyrus granule neurons via activation of theN-methyl-d-aspartate/extracellular signal-regulated kinase/mitogen- and stress-activated kinase signalling pathway. Eur. J. Neurosci. 2008, 27, 2701–2713. [Google Scholar]
- De Kloet, E.; De Kock, S.; Schild, V.; Veldhuis, H.D. Antiglucocorticoid RU 38486 Attenuates Retention of a Behaviour and Disinhibits the Hypothalamic-Pituitary Adrenal Axis at Different Brain Sites. Neuroendocrinology 1988, 47, 109–115. [Google Scholar] [CrossRef]
- Campus, P.; Maiolati, M.; Orsini, C.; Cabib, S. Altered consolidation of extinction-like inhibitory learning in genotype-specific dysfunctional coping fostered by chronic stress in mice. Behav. Brain Res. 2016, 315, 23–35. [Google Scholar] [CrossRef]
- Barker, J.M.; Taylor, J.R.; Chandler, L.J. A unifying model of the role of the infralimbic cortex in extinction and habits. Learn. Mem. 2014, 21, 441–448. [Google Scholar] [CrossRef] [Green Version]
- Huston, J.; Silva, M.A.D.S.; Komorowski, M.; Schulz, D.; Topic, B. Animal models of extinction-induced depression: Loss of reward and its consequences. Neurosci. Biobehav. Rev. 2013, 37, 2059–2070. [Google Scholar] [CrossRef] [PubMed]
- Maier, S.F.; Seligman, M.E.P. Learned helplessness at fifty: Insights from neuroscience. Psychol. Rev. 2016, 123, 349–367. [Google Scholar] [CrossRef] [PubMed]
- Amat, J.; Aleksejev, R.M.; Paul, E.; Watkins, L.R.; Maier, S.F. Behavioral control over shock blocks behavioral and neurochemical effects of later social defeat. Neuroscience 2009, 165, 1031–1038. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Szklarczyk, K.; Korostynski, M.; Golda, S.; Solecki, W.; Przewlocki, R. Genotype-dependent consequences of traumatic stress in four inbred mouse strains. Genes Brain Behav. 2012, 11, 977–985. [Google Scholar] [CrossRef]
- Giorgi, O.; Corda, M.G.; Fernández-Teruel, A. A Genetic Model of Impulsivity, Vulnerability to Drug Abuse and Schizophrenia-Relevant Symptoms with Translational Potential: The Roman High- vs. Low-Avoidance Rats. Front. Behav. Neurosci. 2019, 13, 145. [Google Scholar] [CrossRef]
- Serra, M.P.; Poddighe, L.; Boi, M.; Sanna, F.; Piludu, M.A.; Sanna, F.; Corda, M.G.; Giorgi, O.; Quartu, M. Effect of Acute Stress on the Expression of BDNF, trkB, and PSA-NCAM in the Hippocampus of the Roman Rats: A Genetic Model of Vulnerability/Resistance to Stress-Induced Depression. Int. J. Mol. Sci. 2018, 19, 3745. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Giorgi, O.; Piras, G.; Corda, M.G. The psychogenetically selected Roman high- and low-avoidance rat lines: A model to study the individual vulnerability to drug addiction. Neurosci. Biobehav. Rev. 2007, 31, 148–163. [Google Scholar] [CrossRef]
- Giorgi, O.; Piras, G.; Lecca, D.; Corda, M. Differential activation of dopamine release in the nucleus accumbens core and shell after acute or repeated amphetamine injections: A comparative study in the Roman high- and low-avoidance rat lines. Neuroscience 2005, 135, 987–998. [Google Scholar] [CrossRef]
- Coppens, C.M.; De Boer, S.F.; Koolhaas, J.M. Coping styles and behavioural flexibility: Towards underlying mechanisms. Philos. Trans. R. Soc. B Boil. Sci. 2010, 365, 4021–4028. [Google Scholar] [CrossRef]
- Coppens, C.M.; De Boer, S.F.; Steimer, T.; Koolhaas, J.M. Correlated Behavioral Traits in Rats of the Roman Selection Lines. Behav. Genet. 2013, 43, 220–226. [Google Scholar] [CrossRef] [Green Version]
- De Boer, S.F.; Buwalda, B.; Koolhaas, J.M. Untangling the neurobiology of coping styles in rodents: Towards neural mechanisms underlying individual differences in disease susceptibility. Neurosci. Biobehav. Rev. 2017, 74, 401–422. [Google Scholar] [CrossRef] [PubMed]
- Ventura, R.; Cabib, S.; Puglisi-Allegra, S. Genetic susceptibility of mesocortical dopamine to stress determines liability to inhibition of mesoaccumbens dopamine and to behavioral ’despair’ in a mouse model of depression. Neuroscience 2002, 115, 999–1007. [Google Scholar] [CrossRef]
- Bovet, D.; Bovet-Nitti, F.; Oliverio, A. Genetic Aspects of Learning and Memory in Mice. Science 1969, 163, 139–149. [Google Scholar] [CrossRef] [PubMed]
- Falls, W.A.; Carlson, S.; Turner, J.G.; Willott, J.F. Fear-potentiated startle in two strains of inbred mice. Behav. Neurosci. 1997, 111, 855–861. [Google Scholar] [CrossRef]
- Brennan, F.X. Genetic differences in leverpress escape/avoidance conditioning in seven mouse strains. Genes Brain Behav. 2004, 3, 110–114. [Google Scholar] [CrossRef] [Green Version]
- Andolina, D.; Maran, D.; Viscomi, M.T.; Puglisi-Allegra, S. Strain-Dependent Variations in Stress Coping Behavior Are Mediated by a 5-HT/GABA Interaction within the Prefrontal Corticolimbic System. Int. J. Neuropsychopharmacol. 2014, 18, pyu074. [Google Scholar] [CrossRef] [Green Version]
- Popova, N.K.; Naumenko, V.S.; Tibeikina, M.A.; Kulikov, A.V. Serotonin transporter, 5-HT1Areceptor, and behavior in DBA/2J mice in comparison with four inbred mouse strains. J. Neurosci. Res. 2009, 87, 3649–3657. [Google Scholar] [CrossRef]
- Santarnecchi, E.; Sprugnoli, G.; Tatti, E.; Mencarelli, L.; Neri, F.; Momi, D.; Di Lorenzo, G.; Pascual-Leone, A.; Rossi, S.; Rossi, A. Brain functional connectivity correlates of coping styles. Cogn. Affect. Behav. Neurosci. 2018, 18, 495–508. [Google Scholar] [CrossRef]
- Shanks, N.; Anisman, H. Stressor-provoked behavioral changes in six strains of mice. Behav. Neurosci. 1988, 102, 894–905. [Google Scholar] [CrossRef]
- Alcaro, A.; Cabib, S.; Ventura, R.; Puglisi-Allegra, S. Genotype- and experience-dependent susceptibility to depressive-like responses in the forced-swimming test. Psychopharmacology 2002, 164, 138–143. [Google Scholar] [CrossRef]
- Stramecki, F.; Kotowicz, K.; Piotrowski, P.; Beszłej, J.A.; Rymaszewska, J.; Samochowiec, J.; Samochowiec, A.; Moustafa, A.; Jabłoński, M.; Podwalski, P.; et al. Coping styles and symptomatic manifestation of first-episode psychosis: Focus on cognitive performance. Psychiatry Res. 2019, 272, 246–251. [Google Scholar] [CrossRef] [PubMed]
- Misiak, B.; Kotowicz, K.; Loska, O.; Stramecki, F.; Beszłej, J.A.; Samochowiec, J.; Jabłoński, M.; Podwalski, P.; Waszczuk, K.; Wroński, M.; et al. Decreased use of active coping styles contributes to elevated allostatic load index in first-episode psychosis. Psychoneuroendocrinology 2018, 96, 166–172. [Google Scholar] [CrossRef] [PubMed]
- Fortgang, R.; Hultman, C.M.; Cannon, T.D. Coping Styles in Twins Discordant for Schizophrenia, Bipolar Disorder, and Depression. Clin. Psychol. Sci. 2015, 4, 216–228. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bridi, K.P.B.; Loredo-Souza, A.C.M.; Fijtman, A.; Moreno, M.V.; Kauer-Sant’Anna, M.; Ceresér, K.M.M.; Kunz, M. Differences in coping strategies in adult patients with bipolar disorder and their first-degree relatives in comparison to healthy controls. Trends Psychiatry Psychother. 2018, 40, 318–325. [Google Scholar] [CrossRef] [Green Version]
- Conoscenti, M.A.; Fanselow, M.S. Dissociation in Effective Treatment and Behavioral Phenotype Between Stress-Enhanced Fear Learning and Learned Helplessness. Front. Behav. Neurosci. 2019, 13, 104. [Google Scholar] [CrossRef]
- Cabib, S.; Orsini, C.; Allegra, S.P. Animal models of liability to post-traumatic stress disorder. Behav. Pharmacol. 2019, 30, 122–129. [Google Scholar] [CrossRef]
- Bravo-Rivera, C.; Roman-Ortiz, C.; Montesinos-Cartagena, M.; Quirk, G.J. Persistent active avoidance correlates with activity in prelimbic cortex and ventral striatum. Front. Behav. Neurosci. 2015, 9, 184. [Google Scholar] [CrossRef] [Green Version]
- Gillan, C.; Morein-Zamir, S.; Urcelay, G.; Sule, A.; Voon, V.; Apergis-Schoute, A.M.; Fineberg, N.A.; Sahakian, B.; Robbins, T. Enhanced avoidance habits in obsessive-compulsive disorder. Boil. Psychiatry 2013, 75, 631–638. [Google Scholar] [CrossRef] [Green Version]
- Dalley, J.W.; Everitt, B.J.; Robbins, T. Impulsivity, Compulsivity, and Top-Down Cognitive Control. Neuron 2011, 69, 680–694. [Google Scholar] [CrossRef] [Green Version]
- Gelegen, C.; Collier, D.A.; Campbell, I.C.; Oppelaar, H.; Heuvel, J.V.D.; Adan, R.A.; Kas, M.J.H. Difference in susceptibility to activity-based anorexia in two inbred strains of mice. Eur. Neuropsychopharmacol. 2007, 17, 199–205. [Google Scholar] [CrossRef]
- Pjetri, E.; De Haas, R.; De Jong, S.; Gelegen, C.; Oppelaar, H.; Verhagen, L.A.W.; Eijkemans, M.J.C.; Adan, R.A.; Olivier, B.; Kas, M.J. Identifying Predictors of Activity Based Anorexia Susceptibility in Diverse Genetic Rodent Populations. PLoS ONE 2012, 7, e50453. [Google Scholar] [CrossRef] [Green Version]
- Cabib, S. Abolition and Reversal of Strain Differences in Behavioral Responses to Drugs of Abuse After a Brief Experience. Science 2000, 289, 463–465. [Google Scholar] [CrossRef]
- Badiani, A.; Cabib, S.; Puglisi-Allegra, S. Chronic stress induces strain-dependent sensitization to the behavioral effects of amphetamine in the mouse. Pharmacol. Biochem. Behav. 1992, 43, 53–60. [Google Scholar] [CrossRef]
- Steketee, J.D.; Kalivas, P.W. Drug wanting: Behavioral sensitization and relapse to drug-seeking behavior. Pharmacol. Rev. 2011, 63, 348–365. [Google Scholar] [CrossRef]
- Robinson, T.E.; Berridge, K. Addiction. Annu. Rev. Psychol. 2003, 54, 25–53. [Google Scholar] [CrossRef]
- Flory, J.D.; Yehuda, R. Comorbidity between post-traumatic stress disorder and major depressive disorder: Alternative explanations and treatment considerations. Dialogues Clin. Neurosci. 2015, 17, 141–150. [Google Scholar]
- Roberts, N.P.; Roberts, P.A.; Jones, N.; Bisson, J.I. Psychological interventions for post-traumatic stress disorder and comorbid substance use disorder: A systematic review and meta-analysis. Clin. Psychol. Rev. 2015, 38, 25–38. [Google Scholar] [CrossRef]
- Danzi, B.A.; La Greca, A.M. Genetic pathways to posttraumatic stress disorder and depression in children: Investigation of catechol-O-methyltransferase (COMT) Val158Met using different PTSD diagnostic models. J. Psychiatr. Res. 2018, 102, 81–86. [Google Scholar] [CrossRef] [PubMed]
- Brewerton, T.D. Food addiction as a proxy for eating disorder and obesity severity, trauma history, PTSD symptoms, and comorbidity. Eat. Weight. Disord.-Stud. Anorexia, Bulim. Obes. 2017, 22, 241–247. [Google Scholar] [CrossRef] [PubMed]
- Murphy, D.L.; Moya, P.R.; Fox, M.A.; Rubenstein, L.M.; Wendland, J.R.; Timpano, K.R. Anxiety and affective disorder comorbidity related to serotonin and other neurotransmitter systems: Obsessive–compulsive disorder as an example of overlapping clinical and genetic heterogeneity. Philos. Trans. R. Soc. B Boil. Sci. 2013, 368. [Google Scholar] [CrossRef] [PubMed]
- Cabib, S.; Algeri, S.; Perego, C.; Puglisi-Allegra, S. Behavioral and biochemical changes monitored in two inbred strains of mice during exploration of an unfamiliar environment. Physiol. Behav. 1990, 47, 749–753. [Google Scholar] [CrossRef]
- Cabib, S.; Castellano, C.; Patacchioli, F.R.; Cigliana, G.; Angelucci, L.; Puglisi-Allegra, S. Opposite strain-dependent effects of post-training corticosterone in a passive avoidance task in mice: Role of dopamine. Brain Res. 1996, 729, 110–118. [Google Scholar] [CrossRef]
- Jones, B.C. Contribution of Sex and Genetics to Neuroendocrine Adaptation to Stress in Mice. Psychoneuroendocrinology 1998, 23, 505–517. [Google Scholar] [CrossRef]
- McNamara, R.K.; Lenox, R.H. Acute restraint stress reduces protein kinase C gamma in the hippocampus of C57BL/6 but not DBA/2 mice. Neurosci. Lett. 2004, 368, 293–296. [Google Scholar] [CrossRef]
- Terenina, E.E.; Cavigelli, S.; Mormede, P.; Zhao, W.; Parks, C.; Lu, L.; Jones, B.C.; Mulligan, M.K. Genetic Factors Mediate the Impact of Chronic Stress and Subsequent Response to Novel Acute Stress. Front. Mol. Neurosci. 2019, 13, 438. [Google Scholar] [CrossRef] [Green Version]
- Mozhui, K.; Karlsson, R.-M.; Kash, T.L.; Ihne, J.; Norcross, M.; Patel, S.; Farrell, M.R.; Hill, E.E.; Graybeal, C.; Martin, K.P.; et al. Strain differences in stress responsivity are associated with divergent amygdala gene expression and glutamate-mediated neuronal excitability. J. Neurosci. 2010, 30, 5357–5367. [Google Scholar] [CrossRef] [Green Version]
- De Jong, I.; Steenbergen, P.; De Kloet, E. Strain differences in the effects of adrenalectomy on the midbrain dopamine system: Implication for behavioral sensitization to cocaine. Neuroscience 2008, 153, 594–604. [Google Scholar] [CrossRef]
- De Jong, I.; Steenbergen, P.J.; De Kloet, E. Behavioral sensitization to cocaine: Cooperation between glucocorticoids and epinephrine. Psychopharmacology 2009, 204, 693–703. [Google Scholar] [CrossRef] [Green Version]
- Campus, P.; Canterini, S.; Orsini, C.; Fiorenza, M.T.; Puglisi-Allegra, S.; Cabib, S. Stress-Induced Reduction of Dorsal Striatal D2 Dopamine Receptors Prevents Retention of a Newly Acquired Adaptive Coping Strategy. Front. Pharmacol. 2017, 8, 621. [Google Scholar] [CrossRef] [Green Version]
- Nicholson, A.A.; Friston, K.J.; Zeidman, P.; Harricharan, S.; McKinnon, M.C.; Densmore, M.; Neufeld, R.W.; Théberge, J.; Corrigan, F.; Jetly, R.; et al. Dynamic causal modeling in PTSD and its dissociative subtype: Bottom-up versus top-down processing within fear and emotion regulation circuitry. Hum. Brain Mapp. 2017, 38, 5551–5561. [Google Scholar] [CrossRef] [Green Version]
- Harricharan, S.; Rabellino, D.; Frewen, P.A.; Densmore, M.; Théberge, J.; McKinnon, M.C.; Schore, A.N.; Lanius, R.A. fMRI functional connectivity of the periaqueductal gray in PTSD and its dissociative subtype. Brain Behav. 2016, 6, e00579. [Google Scholar] [CrossRef] [PubMed]
- Zangrossi, H.; Viana, M.B.; Zanoveli, J.M.; Bueno, C.; Nogueira, R.L.; Graeff, F. Serotonergic regulation of inhibitory avoidance and one-way escape in the rat elevated T-maze. Neurosci. Biobehav. Rev. 2001, 25, 637–645. [Google Scholar] [CrossRef]
- Keay, K.A.; Bandler, R. Parallel circuits mediating distinct emotional coping reactions to different types of stress. Neurosci. Biobehav. Rev. 2001, 25, 669–678. [Google Scholar] [CrossRef]

© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cabib, S.; Campus, P.; Conversi, D.; Orsini, C.; Puglisi-Allegra, S. Functional and Dysfunctional Neuroplasticity in Learning to Cope with Stress. Brain Sci. 2020, 10, 127. https://doi.org/10.3390/brainsci10020127
Cabib S, Campus P, Conversi D, Orsini C, Puglisi-Allegra S. Functional and Dysfunctional Neuroplasticity in Learning to Cope with Stress. Brain Sciences. 2020; 10(2):127. https://doi.org/10.3390/brainsci10020127
Chicago/Turabian StyleCabib, Simona, Paolo Campus, David Conversi, Cristina Orsini, and Stefano Puglisi-Allegra. 2020. "Functional and Dysfunctional Neuroplasticity in Learning to Cope with Stress" Brain Sciences 10, no. 2: 127. https://doi.org/10.3390/brainsci10020127
APA StyleCabib, S., Campus, P., Conversi, D., Orsini, C., & Puglisi-Allegra, S. (2020). Functional and Dysfunctional Neuroplasticity in Learning to Cope with Stress. Brain Sciences, 10(2), 127. https://doi.org/10.3390/brainsci10020127