Leg Dominance Effects on Postural Control When Performing Challenging Balance Exercises



Abstract
:1. Introduction
2. Materials and Methods
2.1. Participants
2.2. Experimental Equipment
2.3. Experimental Procedures
2.4. Data Analysis
2.4.1. Kinematic Data Pre-processing
2.4.2. Principal Component Analysis
2.5. Statistical Analysis
3. Results
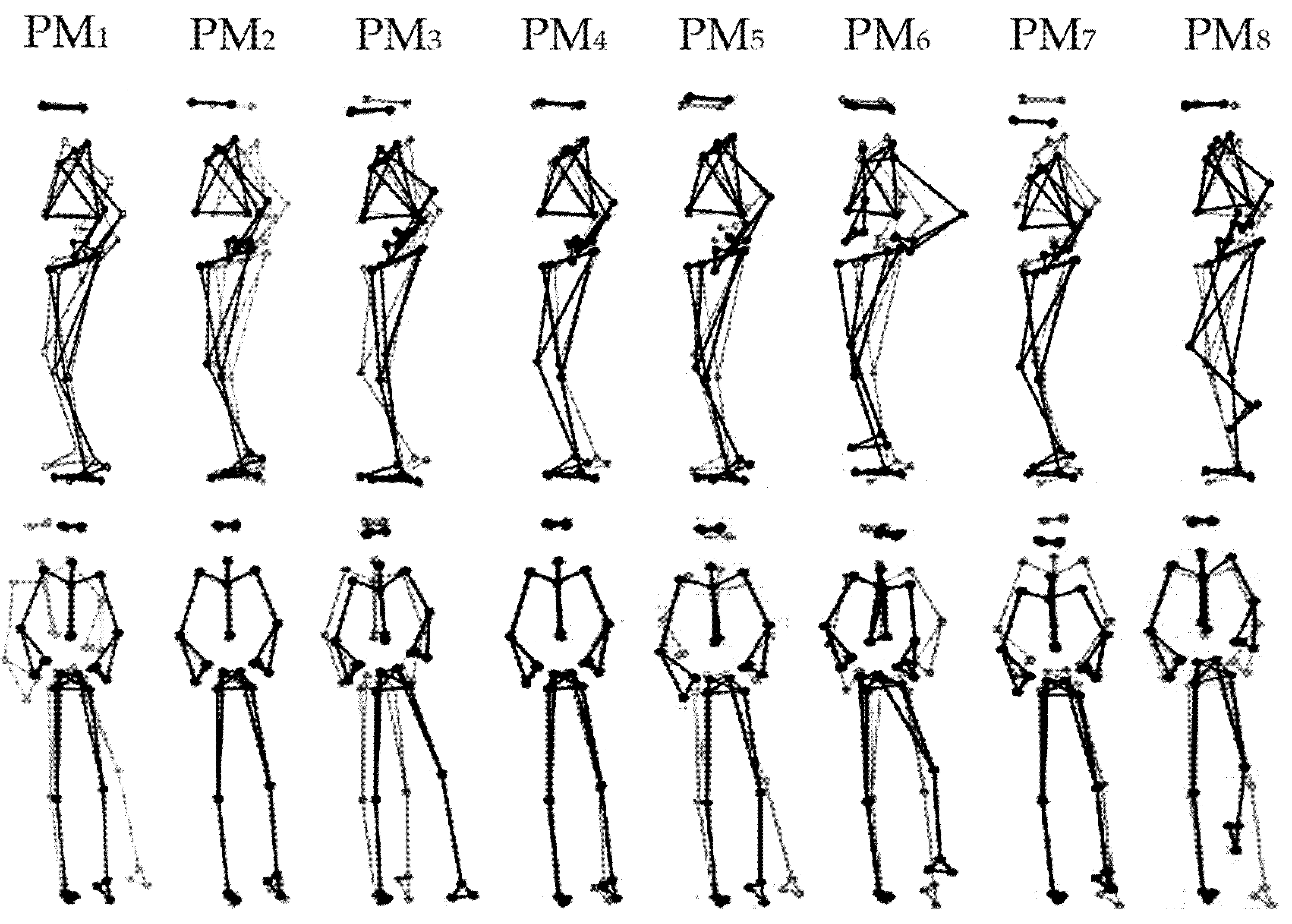
3.1. Characteristics of the First Eight Principal Movements (PM1–8)
3.2. Leg-Dominance Effects on the Composition of Single-Leg Balancing Movements
3.3. Leg-Dominance Effects on the Control of Movement Components
3.4. Sex Effects and Interactions between Sex and Leg Dominance
4. Discussion
Limitations
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Sainburg, R.L. Handedness: Differential specializations for control of trajectory and position. Exerc. Sport Sci. Rev. 2005, 33, 206–213. [Google Scholar] [CrossRef]
- Kapreli, E.; Athanasopoulos, S.; Papathanasiou, M.; Van Hecke, P.; Strimpakos, N.; Gouliamos, A.; Peeters, R.; Sunaert, S. Lateralization of brain activity during lower limb joints movement. An fMRI study. Neuroimage 2006, 32, 1709–1721. [Google Scholar] [CrossRef]
- Schaffer, J.E.; Sainburg, R.L. Interlimb differences in coordination of unsupported reaching movements. Neuroscience 2017, 350, 54–64. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yadav, V.; Sainburg, R.L. Handedness can be explained by a serial hybrid control scheme. Neuroscience 2014, 278, 385–396. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reuter, E.-M.; Cunnington, R.; Mattingley, J.B.; Riek, S.; Carroll, T.J. Feedforward compensation for novel dynamics depends on force field orientation but is similar for the left and right arms. J. Neurophysiol. 2016, 116, 2260–2271. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stockinger, C.; Thürer, B.; Focke, A.; Stein, T. Intermanual transfer characteristics of dynamic learning: direction, coordinate frame, and consolidation of interlimb generalization. J. Neurophysiol. 2015, 114, 3166–3176. [Google Scholar] [CrossRef] [Green Version]
- Velotta, J.; Weyer, J.; Ramirez, A.; Winstead, J.; Bahamonde, R. Relationship between leg dominance tests and type of task. Port. J. Sport Sci. 2011, 11, 1035–1038. [Google Scholar]
- Huurnink, A.; Fransz, D.P.; Kingma, I.; Hupperets, M.D.W.; van Dieën, J.H. The effect of leg preference on postural stability in healthy athletes. J. Biomech. 2014, 47, 308–312. [Google Scholar] [CrossRef] [Green Version]
- Tran, U.S.; Voracek, M. Footedness is associated with self-reported sporting performance and motor abilities in the general population. Front. Psychol. 2016, 7. [Google Scholar] [CrossRef]
- Van Melick, N.; Meddeler, B.M.; Hoogeboom, T.J.; Nijhuis-van der Sanden, M.W.G.G.; van Cingel, R.E.H.H. How to determine leg dominance: The agreement between self-reported and observed performance in healthy adults. PLoS ONE 2017, 12, e0189876. [Google Scholar] [CrossRef] [Green Version]
- Promsri, A.; Haid, T.; Federolf, P. How does lower limb dominance influence postural control movements during single leg stance? Hum. Mov. Sci. 2018, 58, 165–174. [Google Scholar] [CrossRef] [PubMed]
- Brophy, R.; Silvers, H.J.; Gonzales, T.; Mandelbaum, B.R.; Holly, M.; Silvers, J.; Mandelbaum, B.R. Gender influences: The role of leg dominance in ACL injury among soccer players. Br. J. Sports Med. 2010, 44, 694–697. [Google Scholar] [CrossRef] [PubMed]
- Ruedl, G.; Webhofer, M.; Helle, K.; Strobl, M.; Schranz, A.; Fink, C.; Gatterer, H.; Burtscher, M. Leg Dominance Is a Risk Factor for Noncontact Anterior Cruciate Ligament Injuries in Female Recreational Skiers. Am. J. Sports Med. 2012, 40, 1269–1273. [Google Scholar] [CrossRef] [PubMed]
- Steidl-Müller, L.; Hildebrandt, C.; Müller, E.; Fink, C.; Raschner, C. Limb symmetry index in competitive alpine ski racers: Reference values and injury risk identification according to age-related performance levels. J. Sport Heal. Sci. 2018, 7, 405–415. [Google Scholar] [CrossRef]
- Murphy, D.F.; Connolly, D.A.J.; Beynnon, B.D. Risk factors for lower extremity injury: A review of the literature. Br. J. Sports Med. 2003, 37, 13–29. [Google Scholar] [CrossRef] [Green Version]
- Hoffman, M.; Schrader, J.; Applegate, T.; Koceja, D. Unilateral postural control of the functionally dominant and nondominant extremities of healthy subjects. J. Athl. Train. 1998, 33, 319–322. [Google Scholar]
- Teixeira, L.A.; de Oliveira, D.L.; Romano, R.G.; Correa, S.C. Leg preference and interlateral asymmetry of balance stability in soccer players. Res. Q. Exerc. Sport 2011, 82, 21–27. [Google Scholar] [CrossRef]
- King, A.C.; Wang, Z. Asymmetrical stabilization and mobilization exploited during static single leg stance and goal directed kicking. Hum. Mov. Sci. 2017, 54, 182–190. [Google Scholar] [CrossRef]
- Yoshida, T.; Ikemiyagi, F.; Ikemiyagi, Y.; Tanaka, T.; Yamamoto, M.; Suzuki, M. The dominant foot affects the postural control mechanism: examination by body tracking test. Acta Otolaryngol. 2014, 134, 1146–1150. [Google Scholar] [CrossRef]
- Nowicka, A.; Tacikowski, P. Transcallosal transfer of information and functional asymmetry of the human brain. Laterality 2011, 16, 35–74. [Google Scholar] [CrossRef]
- Hellige, J.B.; Laeng, B.; Michimata, C. Processing Asymmetries in the Visual System. In The Two Halves of the Brain; The MIT Press: Cambridge, MA, USA, 2013; pp. 379–416. [Google Scholar]
- Federolf, P.A. A novel approach to study human posture control: “Principal movements” obtained from a principal component analysis of kinematic marker data. J. Biomech. 2016, 49, 364–370. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Horak, F.B.; Nashner, L.M. Central programming of postural movements: adaptation to altered support-surface configurations. J. Neurophysiol. 1986, 55, 1369–1381. [Google Scholar] [CrossRef] [PubMed]
- Federolf, P.; Roos, L.; Nigg, B.M. Analysis of the multi-segmental postural movement strategies utilized in bipedal, tandem and one-leg stance as quantified by a principal component decomposition of marker coordinates. J. Biomech. 2013, 46, 2626–2633. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Todorov, E.; Jordan, M.I. Optimal feedback control as a theory of motor coordination. Nat. Neurosci. 2002, 5, 1226–1235. [Google Scholar] [CrossRef]
- Todorov, E.; Jordan, M.I. A minimal intervention principle for coordinated movement. Adv. Neural Inf. Process. Syst. 2003, 15, 27–34. [Google Scholar]
- Todorov, E. Optimality principles in sensorimotor control. Nat. Neurosci. 2004, 7, 907–915. [Google Scholar] [CrossRef] [Green Version]
- Bernstein, N.A. The co-ordination and regulation of movements: Conclusions towards the Study of Motor Co-ordination. Biodyn. Locomot. 1967, 104–113. [Google Scholar]
- Scott, S.H. Optimal feedback control and the neural basis of volitional motor control. Nat Rev Neurosci. 2004, 5, 532–546. [Google Scholar] [CrossRef]
- Diedrichsen, J.; Shadmehr, R.; Ivry, R.B. The coordination of movement: optimal feedback control and beyond. Trends Cogn. Sci. 2010, 14, 31–39. [Google Scholar] [CrossRef] [Green Version]
- Wachholz, F.; Tiribello, F.; Promsri, A.; Federolf, P. Should the Minimal Intervention Principle Be Considered When Investigating Dual-Tasking Effects on Postural Control? Brain Sci. 2020, 10, 1. [Google Scholar] [CrossRef] [Green Version]
- Haid, T.H.; Doix, A.-C.M.; Nigg, B.M.; Federolf, P.A. Age effects in postural control analyzed via a principal component analysis of kinematic data and interpreted in relation to predictions of the optimal feedback control theory. Front. Aging Neurosci. 2018, 10, 22. [Google Scholar] [CrossRef] [Green Version]
- Haid, T.; Federolf, P. The Effect of Cognitive Resource Competition Due to Dual-Tasking on the Irregularity and Control of Postural Movement Components. Entropy 2019, 21, 70. [Google Scholar] [CrossRef] [Green Version]
- Wachholz, F.; Kockum, T.; Haid, T.; Federolf, P. Changed Temporal Structure of Neuromuscular Control, Rather Than Changed Intersegment Coordination, Explains Altered Stabilographic Regularity After a Moderate Perturbation of the Postural Control System. Entropy 2019, 21, 614. [Google Scholar] [CrossRef] [Green Version]
- Promsri, A.; Longo, A.; Haid, T.; Doix, A.-C.M.; Federolf, P. Leg Dominance as a Risk Factor for Lower-Limb Injuries in Downhill Skiers—A Pilot Study into Possible Mechanisms. Int. J. Environ. Res. Public Health 2019, 16, 3399. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Waddington, G.; Seward, H.; Wrigley, T.; Lacey, N.; Adams, R. Comparing wobble board and jump-landing training effects on knee and ankle movement discrimination. J. Sci. Med. Sport 2000, 3, 449–459. [Google Scholar] [CrossRef]
- Cimadoro, G.; Paizis, C.; Alberti, G.; Babault, N. Effects of different unstable supports on EMG activity and balance. Neurosci. Lett. 2013, 548, 228–232. [Google Scholar] [CrossRef]
- De Ridder, R.; Willems, T.; De Mits, S.; Vanrenterghem, J.; Roosen, P. Foot orientation affects muscle activation levels of ankle stabilizers in a single-legged balance board protocol. Hum. Mov. Sci. 2014, 33, 419–431. [Google Scholar] [CrossRef]
- Clark, V.M.; Burden, A.M. A 4-week wobble board exercise programme improved muscle onset latency and perceived stability in individuals with a functionally unstable ankle. Phys. Ther. Sport 2005, 6, 181–187. [Google Scholar] [CrossRef]
- Promsri, A.; Haid, T.; Federolf, P. Complexity, Composition, and Control of Bipedal Balancing Movements as the Postural Control System Adapts to Unstable Support Surfaces or Altered Feet Positions. Neuroscience 2020, 430, 113–124. [Google Scholar] [CrossRef]
- Merriaux, P.; Dupuis, Y.; Boutteau, R.; Vasseur, P.; Savatier, X. A study of vicon system positioning performance. Sensors 2017, 17, 1591. [Google Scholar] [CrossRef]
- Federolf, P.A. A Novel Approach to Solve the “Missing Marker Problem” in Marker-Based Motion Analysis That Exploits the Segment Coordination Patterns in Multi-Limb Motion Data. PLoS ONE 2013, 8, e78689. [Google Scholar] [CrossRef] [Green Version]
- Gløersen, Ø.; Federolf, P. Predicting missing marker trajectories in human motion data using marker intercorrelations. PLoS ONE 2016, 11, e0152616. [Google Scholar] [CrossRef]
- Daffertshofer, A.; Lamoth, C.J.C.; Meijer, O.G.; Beek, P.J. PCA in studying coordination and variability: A tutorial. Clin. Biomech. 2004, 19, 415–428. [Google Scholar] [CrossRef]
- Troje, N.F. Decomposing biological motion: A framework for analysis and synthesis of human gait patterns. J. Vis. 2002, 2, 371–387. [Google Scholar] [CrossRef]
- Verrel, J.; Lövdén, M.; Schellenbach, M.; Schaefer, S.; Lindenberger, U. Interacting effects of cognitive load and adult age on the regularity of whole-body motion during treadmill walking. Psychol. Aging 2009, 24, 75–81. [Google Scholar] [CrossRef] [Green Version]
- Haid, T.H.; Zago, M.; Promsri, A.; Doix, A.-C.M.; Federolf, P.A. PManalyzer: A Software Facilitating the Study of Sensorimotor Control of Whole-Body Movements. Front. Neuroinform. 2019, 13, 24. [Google Scholar] [CrossRef] [Green Version]
- De Leva, P. Adjustments to Zatsiorsky-Seluyanov’s segment inertia parameters. J. Biomech. 1996, 29, 1223–1230. [Google Scholar] [CrossRef]
- Gløersen, Ø.; Myklebust, H.; Hallén, J.; Federolf, P. Technique analysis in elite athletes using principal component analysis. J. Sports Sci. 2018, 36, 229–237. [Google Scholar] [CrossRef] [PubMed]
- Nashner, L.M.; McCollum, G. The organization of human postural movements: A formal basis and experimental synthesis. Behav. Brain Sci. 1985, 8, 135–172. [Google Scholar] [CrossRef]
- Gatev, P.; Thomas, S.; Kepple, T.; Hallett, M. Feedforward ankle strategy of balance during quiet stance in adults. J. Physiol. 1999, 514, 915–928. [Google Scholar] [CrossRef] [PubMed]
- Winter, D.A. Human balance and posture control during standing and walking. Gait Posture 1995, 3, 193–214. [Google Scholar] [CrossRef]
- Longo, A.; Haid, T.; Meulenbroek, R.; Federolf, P. Biomechanics in posture space: Properties and relevance of principal accelerations for characterizing movement control. J. Biomech. 2019, 82, 397–403. [Google Scholar] [CrossRef] [PubMed]
- Winter, D.A.; Sidwall, H.G.; Hobson, D.A. Measurement and reduction of noise in kinematics of locomotion. J. Biomech. 1974, 7, 157–159. [Google Scholar] [CrossRef]
- Holm, S. A simple sequentially rejective Bonferroni test procedure. Scand. J. Stat. 1979, 6, 65–70. [Google Scholar]
- Amrhein, V.; Greenland, S.; McShane, B. Scientists rise up against statistical significance. Nature 2019, 567, 305–307. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cohen, J. Eta-Squared and Partial Eta-Squared in Fixed Factor Anova Designs. Educ. Psychol. Meas. 1973, 33, 107–112. [Google Scholar] [CrossRef]
- Van Dieën, J.H.; van Leeuwen, M.; Faber, G.S. Learning to balance on one leg: Motor strategy and sensory weighting. J. Neurophysiol. 2015, 114, 2967–2982. [Google Scholar] [CrossRef]
- Proske, U. What is the role of muscle receptors in proprioception? Muscl. Nerve 2005, 31, 780–787. [Google Scholar] [CrossRef]
- Proske, U.; Gandevia, S.C. The proprioceptive senses: their roles in signaling body shape, body position and movement, and muscle force. Physiol. Rev. 2012, 92, 1651–1697. [Google Scholar] [CrossRef]
- Horslen, B.C.; Murnaghan, C.D.; Inglis, J.T.; Chua, R.; Carpenter, M.G. Effects of postural threat on spinal stretch reflexes: Evidence for increased muscle spindle sensitivity? J. Neurophysiol. 2013, 110, 899–906. [Google Scholar] [CrossRef] [Green Version]
- Davis, J.R.; Horslen, B.C.; Nishikawa, K.; Fukushima, K.; Chua, R.; Inglis, J.T.; Carpenter, M.G. Human proprioceptive adaptations during states of height-induced fear and anxiety. J. Neurophysiol. 2011, 106, 3082–3090. [Google Scholar] [CrossRef] [PubMed]
- Sharman, M.J.; Cresswell, A.G.; Riek, S. Proprioceptive neuromuscular facilitation stretching: Mechanisms and clinical implications. Sport. Med. 2006, 36, 929–939. [Google Scholar] [CrossRef] [PubMed]
- Moreira, R.; Lial, L.; Teles Monteiro, M.G.; Aragão, A.; Santos David, L.; Coertjens, M.; Silva-Júnior, F.L.; Dias, G.; Velasques, B.; Ribeiro, P.; et al. Diagonal movement of the upper limb produces greater adaptive plasticity than sagittal plane flexion in the shoulder. Neurosci. Lett. 2017, 643, 8–15. [Google Scholar] [CrossRef] [PubMed]
- Kibler, W.B.; Press, J.; Sciascia, A. The role of core stability in athletic function. Sport. Med. 2006, 36, 189–198. [Google Scholar] [CrossRef] [PubMed]
- Akuthota, V.; Ferreiro, A.; Moore, T.; Fredericson, M. Core stability exercise principles. Curr. Sports Med. Rep. 2008, 7, 39–44. [Google Scholar] [CrossRef] [PubMed]
- Ham, Y.W.; Kim, D.M.; Baek, J.Y.; Lee, D.C.; Sung, P.S. Kinematic analyses of trunk stability in one leg standing for individuals with recurrent low back pain. J. Electromyogr. Kinesiol. 2010, 20, 1134–1140. [Google Scholar] [CrossRef] [PubMed]
- Leetun, D.T.; Ireland, M.L.; Willson, J.D.; Ballantyne, B.T.; Davis, I.M.C. Core stability measures as risk factors for lower extremity injury in athletes. Med. Sci. Sports Exerc. 2004, 36, 926–934. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McGrath, T.M.; Waddington, G.; Scarvell, J.M.; Ball, N.B.; Creer, R.; Woods, K.; Smith, D. The effect of limb dominance on lower limb functional performance—A systematic review. J. Sports Sci. 2016, 34, 289–302. [Google Scholar] [CrossRef]
- Ocklenburg, S.; Beste, C.; Güntürkün, O. Handedness: A neurogenetic shift of perspective. Neurosci. Biobehav. Rev. 2013, 37, 2788–2793. [Google Scholar] [CrossRef]
- Frayer, D.W.; Lozano, M.; Bermúdez de Castro, J.M.; Carbonell, E.; Arsuaga, J.L.; Radovčić, J.; Fiore, I.; Bondioli, L. More than 500,000 years of right-handedness in Europe. Laterality 2012, 17, 51–69. [Google Scholar] [CrossRef]


{kind=link}
| Male (n = 14) | Female (n = 12) | p-Value | |
|---|---|---|---|
| Age (y) | 25.8 ± 2.9 | 24.6 ± 5.3 | 0.478 |
| Weight (kg) | 77.5 ± 10.8 | 62.6 ± 4.9 | <0.001 * |
| Height (cm) | 180.0 ± 7.2 | 169.2 ± 4.3 | <0.001 * |
| Body mass index (kg/m2) | 23.9 ± 2.8 | 21.9 ± 2.3 | 0.059 |
| Physical-activity participation (h/wk) | 8.1 ± 5.5 | 8.8 ± 4.5 | 0.418 |
| PM | rVAR [%] | Main Plane of Motion | Main Movements |
|---|---|---|---|
| 1 | 44.2 ± 11.3 | Frontal (Mediolateral) | Hip strategy coupled with ankle supination/pronation and knee flexion/extension of the stance leg, and hip adduction/abduction and flexion/extension of the lifted leg |
| 2 | 26.0 ± 13.2 | Sagittal (Anteroposterior) | Ankle strategy |
| 3 | 7.6 ± 5.7 | Diagonal (Anterolateral) | Hip strategy coupled with ankle supination/pronation of the stance leg, and the combination of hip abduction/adduction and knee flexion/extension of the lifted leg |
| 4 | 7.0 ± 3.9 | Frontal (Mediolateral) | Hip strategy coupled with knee flexion/extension and ankle supination/pronation of the stance leg, and knee flexion/extension of the lifted leg |
| 5 | 3.2 ± 3.0 | Frontal (Mediolateral) | Hip strategy coupled with ankle supination/pronation of the stance leg, and the combination of hip flexion/extension and abduction/adduction of the lifted leg |
| 6 | 2.5 ± 1.6 | Diagonal (Oblique- transverse) | Upper-body oblique rotation to the lifted leg coupled with hip flexion/extension of the lifted leg |
| 7 | 2.0 ± 2.5 | Vertical | Breathing movement patterns: upward/downward moving of the shoulders coupled with knee flexion/extension of the stance leg, and the combination of hip and knee flexion/extension of the lifted leg |
| 8 | 1.6 ± 1.2 | Vertical | Hip and knee flexion/extension of the lifted leg coupled with slightly upward/downward moving of the shoulder, and ankle dorsiflexion/plantarflexion of the stance leg |
| PM | rVAR (%) | ||||
|---|---|---|---|---|---|
| ND | DO | p-Value | Effect Size | Observed Power | |
| 1 a | 41.1 ± 13.2 | 47.2 ± 11.5 | 0.061 | ηp² = 0.139 | 0.472 |
| 2 a | 29.3 ± 12.7 | 22.8 ± 13.5 | 0.046 | ηp² = 0.156 | 0.524 |
| 3 b | 7.8 ± 7.2 | 7.3 ± 5.8 | 0.970 | r = 0.007 | 0.050 |
| 4 b | 6.9 ± 5.2 | 7.1 ± 3.9 | 0.949 | r = 0.012 | 0.050 |
| 5 b | 3.1 ± 2.0 | 3.3 ± 3.1 | 0.949 | r = 0.012 | 0.050 |
| 6 b | 2.2 ± 1.3 | 2.7 ± 1.6 | 0.269 | r = 0.216 | 0.185 |
| 7 b | 1.7 ± 1.6 | 2.3 ± 2.6 | 0.151 | r = 0.281 | 0.281 |
| 8 b | 1.5 ± 0.9 | 1.7 ± 1.2 | 0.568 | r = 0.111 | 0.085 |
| k | Nk (Counts of PAk(t)-zero Crossings during 60 s) | ||||
| ND | DO | p-Value | Effect Size (ηp2) | Observed Power | |
| 1 | 441 ± 74 | 433 ± 67 | 0.642 | 0.009 | 0.074 |
| 2 | 527 ± 72 | 511 ± 74 | 0.204 | 0.066 | 0.241 |
| 3 | 516 ± 73 | 464 ± 75 | 0.005 * | 0.283 | 0.839 |
| 4 | 531 ± 57 | 510 ± 68 | 0.144 | 0.087 | 0.305 |
| 5 | 531 ± 55 | 532 ± 56 | 0.832 | 0.002 | 0.055 |
| 6 | 565 ± 72 | 543 ± 72 | 0.016 | 0.218 | 0.700 |
| 7 | 509 ± 40 | 503 ± 45 | 0.444 | 0.025 | 0.116 |
| 8 | 519 ± 56 | 515 ± 53 | 0.632 | 0.010 | 0.075 |
| k | σk (milliseconds) | ||||
| ND | DO | p-Value | Effect Size (ηp2) | Observed Power | |
| 1 | 108.4 ± 28.9 | 113.6 ± 28.9 | 0.434 | 0.026 | 0.119 |
| 2 | 84.6 ± 18.4 | 86.7 ± 19.4 | 0.522 | 0.017 | 0.096 |
| 3 | 85.3 ± 18.3 | 96.3 ± 21.2 | 0.035 | 0.172 | 0.573 |
| 4 | 71.2 ± 15.6 | 74.9 ± 20.0 | 0.345 | 0.037 | 0.152 |
| 5 | 72.1 ± 13.6 | 72.4 ± 13.3 | 0.900 | 0.001 | 0.052 |
| 6 | 68.0 ± 14.2 | 74.6 ± 17.2 | 0.025 | 0.192 | 0.630 |
| 7 | 52.7 ± 8.4 | 53.6 ± 10.4 | 0.657 | 0.008 | 0.072 |
| 8 | 55.1 ± 7.5 | 56.1 ± 8.1 | 0.492 | 0.020 | 0.103 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Promsri, A.; Haid, T.; Werner, I.; Federolf, P. Leg Dominance Effects on Postural Control When Performing Challenging Balance Exercises. Brain Sci. 2020, 10, 128. https://doi.org/10.3390/brainsci10030128
Promsri A, Haid T, Werner I, Federolf P. Leg Dominance Effects on Postural Control When Performing Challenging Balance Exercises. Brain Sciences. 2020; 10(3):128. https://doi.org/10.3390/brainsci10030128
Chicago/Turabian StylePromsri, Arunee, Thomas Haid, Inge Werner, and Peter Federolf. 2020. "Leg Dominance Effects on Postural Control When Performing Challenging Balance Exercises" Brain Sciences 10, no. 3: 128. https://doi.org/10.3390/brainsci10030128
APA StylePromsri, A., Haid, T., Werner, I., & Federolf, P. (2020). Leg Dominance Effects on Postural Control When Performing Challenging Balance Exercises. Brain Sciences, 10(3), 128. https://doi.org/10.3390/brainsci10030128