Increased Resting State Triple Network Functional Connectivity in Undergraduate Problematic Cannabis Users: A Preliminary EEG Coherence Study

 , , , and
, , , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Participants
2.2. Self-Report Measures
2.3. EEG Data Acquisition and Functional Connectivity Analysis
2.4. Statistical Analysis
3. Results
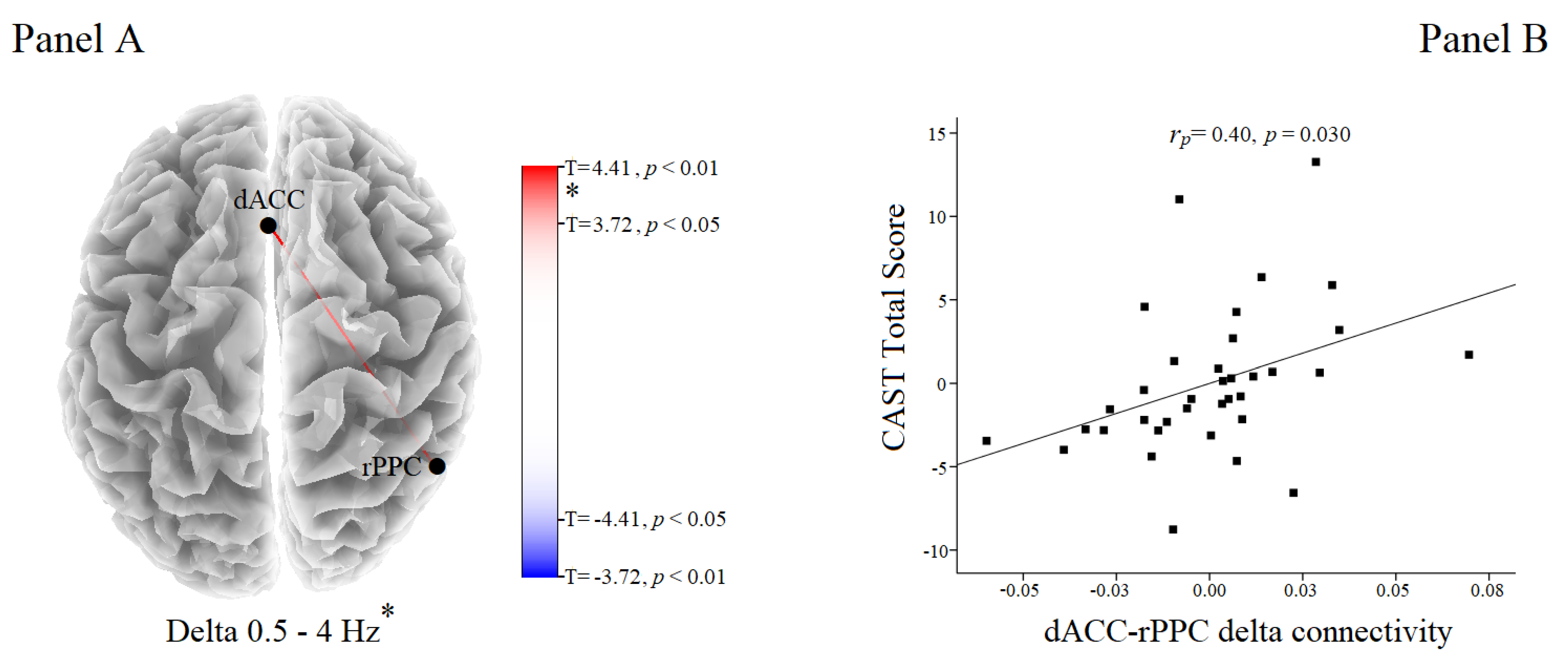
Functional Connectivity Results
4. Discussion
5. Conclusions
Author Contributions
Conflicts of Interest
Abbreviations
| eLORETA | exact Low Resolution Electromagnetic Tomography software |
| MNI | Montreal Neurological Institute |
| DMN | default mode network |
| mPFC | medial prefrontal cortex |
| PCC | posterior cingulate cortex |
| SN | salience network |
| dACC | dorsal anterior cingulate cortex |
| AI | anterior insula |
| CEN | central executive network |
| dlPFC | dorsolateral prefrontal cortex |
| PPC | posterior parietal cortex |
References
- European Monitoring Centre for Drugs and Drug Addiction. European Drug Report 2019: Trends and Developments; Publications Office of the European Union: Luxembourg, Belgium, 2019. [Google Scholar]
- Hasin, D.S.; Kerridge, B.T.; Saha, T.D.; Huang, B.; Pickering, R.; Smith, S.M.; Jung, J.; Zhang, H.; Grant, B.F. Prevalence and correlates of dsm-5 cannabis use disorder, 2012–2013: Findings from the national epidemiologic survey on alcohol and related conditions-iii. Am. J. Psychiatry 2016, 173, 588–599. [Google Scholar] [CrossRef] [PubMed]
- Yanes, J.A.; Riedel, M.C.; Ray, K.L.; Kirkland, A.E.; Bird, R.T.; Boeving, E.R.; Reid, M.A.; Gonzalez, R.; Robinson, J.L.; Laird, A.R.; et al. Neuroimaging meta-analysis of cannabis use studies reveals convergent functional alterations in brain regions supporting cognitive control and reward processing. J. Psychopharmacol. 2018, 32, 283–295. [Google Scholar] [CrossRef] [PubMed]
- Hirst, R.A.; Lambert, D.G.; Notcutt, W.G. Pharmacology and potential therapeutic uses of cannabis. Br. J. Anaesth. 1998, 81, 77–84. [Google Scholar] [CrossRef] [PubMed]
- Calabrese, E.J.; Rubio-Casillas, A. Biphasic effects of thc in memory and cognition. Eur. J. Clin. Invest. 2018, 48, e12920. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zehra, A.; Burns, J.; Liu, C.K.; Manza, P.; Wiers, C.E.; Volkow, N.D.; Wang, G.J. Cannabis addiction and the brain: A review. J. Neuroimmune Pharmacol. 2018, 13, 438–452. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bloomfield, M.A.P.; Hindocha, C.; Green, S.F.; Wall, M.B.; Lees, R.; Petrilli, K.; Costello, H.; Ogunbiyi, M.O.; Bossong, M.G.; Freeman, T.P. The neuropsychopharmacology of cannabis: A review of human imaging studies. Pharmacol. Ther. 2019, 195, 132–161. [Google Scholar] [CrossRef]
- Nader, D.A.; Sanchez, Z.M. Effects of regular cannabis use on neurocognition, brain structure, and function: A systematic review of findings in adults. Am. J. Drug Alcohol Abus. 2018, 44, 4–18. [Google Scholar] [CrossRef]
- Moreno-Alcazar, A.; Gonzalvo, B.; Canales-Rodriguez, E.J.; Blanco, L.; Bachiller, D.; Romaguera, A.; Monte-Rubio, G.C.; Roncero, C.; McKenna, P.J.; Pomarol-Clotet, E. Larger gray matter volume in the basal ganglia of heavy cannabis users detected by voxel-based morphometry and subcortical volumetric analysis. Front. Psychiatry 2018, 9, 175. [Google Scholar] [CrossRef] [Green Version]
- Shevorykin, A.; Ruglass, L.M.; Melara, R.D. Frontal alpha asymmetry and inhibitory control among individuals with cannabis use disorders. Brain Sci. 2019, 9, 219. [Google Scholar] [CrossRef] [Green Version]
- Asmaro, D.; Carolan, P.L.; Liotti, M. Electrophysiological evidence of early attentional bias to drug-related pictures in chronic cannabis users. Addict. Behav. 2014, 39, 114–121. [Google Scholar] [CrossRef]
- Ruglass, L.M.; Shevorykin, A.; Dambreville, N.; Melara, R.D. Neural and behavioral correlates of attentional bias to cannabis cues among adults with cannabis use disorders. Psychol. Addict. Behav. 2019, 33, 69–80. [Google Scholar] [CrossRef] [PubMed]
- Prashad, S.; Dedrick, E.S.; Filbey, F.M. Cannabis users exhibit increased cortical activation during resting state compared to non-users. Neuroimage 2018, 179, 176–186. [Google Scholar] [CrossRef] [PubMed]
- Struve, F.A.; Patrick, G.; Straumanis, J.J.; Fitz-Gerald, M.J.; Manno, J. Possible eeg sequelae of very long duration marihuana use: Pilot findings from topographic quantitative eeg analyses of subjects with 15 to 24 years of cumulative daily exposure to thc. Clin. Electroencephalogr. 1998, 29, 31–36. [Google Scholar] [CrossRef] [PubMed]
- Struve, F.A.; Straumanis, J.J.; Patrick, G.; Leavitt, J.; Manno, J.E.; Manno, B.R. Topographic quantitative eeg sequelae of chronic marihuana use: A replication using medically and psychiatrically screened normal subjects. Drug Alcohol Depend. 1999, 56, 167–179. [Google Scholar] [CrossRef]
- Fingelkurts, A.A.; Kivisaari, R.; Autti, T.; Borisov, S.; Puuskari, V.; Jokela, O.; Kahkonen, S. Increased local and decreased remote functional connectivity at eeg alpha and beta frequency bands in opioid-dependent patients. Psychopharmacology 2006, 188, 42–52. [Google Scholar] [CrossRef] [PubMed]
- Fingelkurts, A.A.; Kivisaari, R.; Autti, T.; Borisov, S.; Puuskari, V.; Jokela, O.; Kahkonen, S. Opioid withdrawal results in an increased local and remote functional connectivity at eeg alpha and beta frequency bands. Neurosci. Res. 2007, 58, 40–49. [Google Scholar] [CrossRef]
- Zilverstand, A.; Huang, A.S.; Alia-Klein, N.; Goldstein, R.Z. Neuroimaging impaired response inhibition and salience attribution in human drug addiction: A systematic review. Neuron 2018, 98, 886–903. [Google Scholar] [CrossRef] [Green Version]
- Bechara, A. Decision making, impulse control and loss of willpower to resist drugs: A neurocognitive perspective. Nat. Neurosci. 2005, 8, 1458–1463. [Google Scholar] [CrossRef]
- Menon, V. Large-Scale brain networks and psychopathology: A unifying triple network model. Trends Cogn. Sci. 2011, 15, 483–506. [Google Scholar] [CrossRef]
- Bressler, S.L.; Menon, V. Large-Scale brain networks in cognition: Emerging methods and principles. Trends Cogn. Sci. 2010, 14, 277–290. [Google Scholar] [CrossRef]
- Menon, B. Towards a new model of understanding—The triple network, psychopathology and the structure of the mind. Med. Hypotheses 2019, 133, 109385. [Google Scholar] [CrossRef] [PubMed]
- Andrews-Hanna, J.R.; Smallwood, J.; Spreng, R.N. The default network and self-generated thought: Component processes, dynamic control, and clinical relevance. Ann. N. Y. Acad. Sci. 2014, 1316, 29–52. [Google Scholar] [CrossRef] [PubMed]
- Andrews-Hanna, J.R. The brain’s default network and its adaptive role in internal mentation. Neuroscientist 2012, 18, 251–270. [Google Scholar] [CrossRef] [PubMed]
- Goulden, N.; Khusnulina, A.; Davis, N.J.; Bracewell, R.M.; Bokde, A.L.; McNulty, J.P.; Mullins, P.G. The salience network is responsible for switching between the default mode network and the central executive network: Replication from dcm. Neuroimage 2014, 99, 180–190. [Google Scholar] [CrossRef] [PubMed]
- Sridharan, D.; Levitin, D.J.; Menon, V. A critical role for the right fronto-insular cortex in switching between central-executive and default-mode networks. Proc. Natl. Acad. Sci. USA 2008, 105, 12569–12574. [Google Scholar] [CrossRef] [Green Version]
- Seeley, W.W.; Menon, V.; Schatzberg, A.F.; Keller, J.; Glover, G.H.; Kenna, H.; Reiss, A.L.; Greicius, M.D. Dissociable intrinsic connectivity networks for salience processing and executive control. J. Neurosci. 2007, 27, 2349–2356. [Google Scholar] [CrossRef]
- Menon, V.; Uddin, L.Q. Saliency, switching, attention and control: A network model of insula function. Brain Struct. Funct. 2010, 214, 655–667. [Google Scholar] [CrossRef] [Green Version]
- Sutherland, M.T.; McHugh, M.J.; Pariyadath, V.; Stein, E.A. Resting state functional connectivity in addiction: Lessons learned and a road ahead. Neuroimage 2012, 62, 2281–2295. [Google Scholar] [CrossRef] [Green Version]
- Lerman, C.; Gu, H.; Loughead, J.; Ruparel, K.; Yang, Y.; Stein, E.A. Large-Scale brain network coupling predicts acute nicotine abstinence effects on craving and cognitive function. JAMA Psychiatry 2014, 71, 523–530. [Google Scholar] [CrossRef]
- Liang, X.; He, Y.; Salmeron, B.J.; Gu, H.; Stein, E.A.; Yang, Y. Interactions between the salience and default-mode networks are disrupted in cocaine addiction. J. Neurosci. 2015, 35, 8081–8090. [Google Scholar] [CrossRef] [Green Version]
- Geng, X.; Hu, Y.; Gu, H.; Salmeron, B.J.; Adinoff, B.; Stein, E.A.; Yang, Y. Salience and default mode network dysregulation in chronic cocaine users predict treatment outcome. Brain 2017, 140, 1513–1524. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, Q.; Liu, J.; Wang, W.; Wang, Y.; Li, W.; Chen, J.; Zhu, J.; Yan, X.; Li, Y.; Li, Z.; et al. Disrupted coupling of large-scale networks is associated with relapse behaviour in heroin-dependent men. J. Psychiatry Neurosci. 2018, 43, 48–57. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wall, M.B.; Pope, R.; Freeman, T.P.; Kowalczyk, O.S.; Demetriou, L.; Mokrysz, C.; Hindocha, C.; Lawn, W.; Bloomfield, M.A.; Freeman, A.M.; et al. Dissociable effects of cannabis with and without cannabidiol on the human brain’s resting-state functional connectivity. J. Psychopharmacol. 2019, 33, 822–830. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Neuner, I.; Arrubla, J.; Werner, C.J.; Hitz, K.; Boers, F.; Kawohl, W.; Shah, N.J. The default mode network and eeg regional spectral power: A simultaneous fmri-eeg study. PLoS ONE 2014, 9, e88214. [Google Scholar] [CrossRef] [Green Version]
- Thatcher, R.W.; North, D.M.; Biver, C.J. Loreta eeg phase reset of the default mode network. Front. Hum. Neurosci. 2014, 8, 529. [Google Scholar] [CrossRef]
- Srinivasan, R.; Winter, W.R.; Ding, J.; Nunez, P.L. Eeg and meg coherence: Measures of functional connectivity at distinct spatial scales of neocortical dynamics. J. Neurosci. Methods 2007, 166, 41–52. [Google Scholar] [CrossRef] [Green Version]
- Whitton, A.E.; Deccy, S.; Ironside, M.L.; Kumar, P.; Beltzer, M.; Pizzagalli, D.A. Electroencephalography source functional connectivity reveals abnormal high-frequency communication among large-scale functional networks in depression. Biol. Psychiatry Cogn. Neurosci. Neuroimaging 2018, 3, 50–58. [Google Scholar] [CrossRef]
- Legleye, S.; Karila, L.; Beck, F.; Reynaud, M. Validation of the cast, a general population cannabis abuse screening test. J. Subst. Use 2007, 17, 233–242. [Google Scholar] [CrossRef]
- Bastiani, L.; Siciliano, V.; Curzio, O.; Luppi, C.; Gori, M.; Grassi, M.; Molinaro, S. Optimal scaling of the cast and of sds scale in a national sample of adolescents. Addict. Behav. 2013, 38, 2060–2067. [Google Scholar] [CrossRef]
- Ewing, J.A. Detecting alcoholism. The cage questionnaire. JAMA 1984, 252, 1905–1907. [Google Scholar] [CrossRef]
- Klaghofer, R.; Brähler, E. Konstruktion und teststatistische prüfung einer kurzform der scl-90-r (construction and test statistical evaluation of a short version of the scl-90–r). Z. Klin. Psychol. Psychiatr. Psychother. 2001, 49, 115–124. [Google Scholar]
- Imperatori, C.; Farina, B.; Valenti, E.M.; Di Poce, A.; D’Ari, S.; De Rossi, E.; Murgia, C.; Carbone, G.A.; Massullo, C.; Della Marca, G. Is resting state frontal alpha connectivity asymmetry a useful index to assess depressive symptoms? A preliminary investigation in a sample of university students. J. Affect. Disord. 2019, 257, 152–159. [Google Scholar] [CrossRef] [PubMed]
- Adenzato, M.; Imperatori, C.; Ardito, R.B.; Valenti, E.M.; Marca, G.D.; D’Ari, S.; Palmiero, L.; Penso, J.S.; Farina, B. Activating attachment memories affects default mode network in a non-clinical sample with perceived dysfunctional parenting: An eeg functional connectivity study. Behav. Brain Res. 2019, 372, 112059. [Google Scholar] [CrossRef] [PubMed]
- Imperatori, C.; Farina, B.; Adenzato, M.; Valenti, E.M.; Murgia, C.; Marca, G.D.; Brunetti, R.; Fontana, E.; Ardito, R.B. Default mode network alterations in individuals with high-trait-anxiety: An eeg functional connectivity study. J. Affect. Disord. 2019, 246, 611–618. [Google Scholar] [CrossRef]
- Imperatori, C.; Massullo, C.; Carbone, G.A.; Farina, B.; Colmegna, F.; Riboldi, I.; Giacomo, E.D.; Clerici, M.; Dakanalis, A. Electroencephalographic (eeg) alterations in young women with high subclinical eating pathology levels: A quantitative eeg study. Eat. Weight Disord. 2019. [Google Scholar] [CrossRef]
- Casajuana, C.; Lopez-Pelayo, H.; Balcells, M.M.; Miquel, L.; Colom, J.; Gual, A. Definitions of risky and problematic cannabis use: A systematic review. Subst. Use Misuse 2016, 51, 1760–1770. [Google Scholar] [CrossRef]
- Legleye, S.; Kraus, L.; Piontek, D.; Phan, O.; Jouanne, C. Validation of the cannabis abuse screening test in a sample of cannabis inpatients. Eur. Addict. Res. 2012, 18, 193–200. [Google Scholar] [CrossRef]
- Legleye, S.; Piontek, D.; Kraus, L. Psychometric properties of the cannabis abuse screening test (cast) in a french sample of adolescents. Drug Alcohol Depend. 2011, 113, 229–235. [Google Scholar] [CrossRef]
- Legleye, S.; Eslami, A.; Bougeard, S. Assessing the structure of the cast (cannabis abuse screening test) in 13 European countries using multigroup analyses. Int. J. Methods Psychiatr. Res. 2017, 26. [Google Scholar] [CrossRef]
- Cantillano, V.; Del Villar, P.; Contreras, L.; Martinez, D.; Zuzulich, M.S.; Ramirez, C.; Pons, C.; Bashford, J. Psychometric properties of the spanish version of the cannabis use problems identification test among Chilean University students: A validation study. Drug Alcohol Depend. 2017, 170, 32–36. [Google Scholar] [CrossRef]
- Mayfield, D.; McLeod, G.; Hall, P. The cage questionnaire: Validation of a new alcoholism screening instrument. Am. J. Psychiatry 1974, 131, 1121–1123. [Google Scholar] [PubMed]
- Dhalla, S.; Kopec, J.A. The cage questionnaire for alcohol misuse: A review of reliability and validity studies. Clin. Invest. Med. 2007, 30, 33–41. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Agabio, R.; Marras, P.; Gessa, G.L.; Carpiniello, B. Alcohol use disorders, and at-risk drinking in patients affected by a mood disorder, in Cagliari, Italy: Sensitivity and specificity of different questionnaires. Alcohol Alcohol. 2007, 42, 575–581. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Derogatis, L. The Scl-90-R Manual; Clinical Psychometric Research Unit, Johns Hopkins University School of Medicine: Baltimore, MD, USA, 1977. [Google Scholar]
- Petrowski, K.; Schmalbach, B.; Kliem, S.; Hinz, A.; Brahler, E. Symptom-checklist-k-9: Norm values and factorial structure in a representative German sample. PLoS ONE 2019, 14, e0213490. [Google Scholar] [CrossRef] [Green Version]
- Imperatori, C.; Bianciardi, E.; Niolu, C.; Fabbricatore, M.; Gentileschi, P.; Di Lorenzo, G.; Siracusano, A.; Innamorati, M. The Symptom-Checklist-K-9 (SCL-K-9) discriminates between overweight/obese patients with and without significant binge eating pathology: Psychometric Properties of an Italian version. Nutrients 2020, 12, 647. [Google Scholar] [CrossRef] [Green Version]
- Imperatori, C.; Brunetti, R.; Farina, B.; Speranza, A.M.; Losurdo, A.; Testani, E.; Contardi, A.; Della Marca, G. Modification of eeg power spectra and eeg connectivity in autobiographical memory: A sloreta study. Cogn. Process. 2014, 15, 351–361. [Google Scholar] [CrossRef]
- Imperatori, C.; Farina, B.; Brunetti, R.; Gnoni, V.; Testani, E.; Quintiliani, M.I.; Del Gatto, C.; Indraccolo, A.; Contardi, A.; Speranza, A.M.; et al. Modifications of eeg power spectra in mesial temporal lobe during n-back tasks of increasing difficulty. A sloreta study. Front. Hum. Neurosci. 2013, 7, 109. [Google Scholar] [CrossRef] [Green Version]
- Imperatori, C.; Farina, B.; Quintiliani, M.I.; Onofri, A.; Castelli Gattinara, P.; Lepore, M.; Gnoni, V.; Mazzucchi, E.; Contardi, A.; Della Marca, G. Aberrant eeg functional connectivity and eeg power spectra in resting state post-traumatic stress disorder: A sloreta study. Biol. Psychol. 2014, 102, 10–17. [Google Scholar] [CrossRef]
- Imperatori, C.; Della Marca, G.; Amoroso, N.; Maestoso, G.; Valenti, E.M.; Massullo, C.; Carbone, G.A.; Contardi, A.; Farina, B. Alpha/Theta neurofeedback increases mentalization and default mode network connectivity in a non-clinical sample. Brain Topogr. 2017, 30, 822–831. [Google Scholar] [CrossRef]
- Canuet, L.; Tellado, I.; Couceiro, V.; Fraile, C.; Fernandez-Novoa, L.; Ishii, R.; Takeda, M.; Cacabelos, R. Resting-State network disruption and apoe genotype in Alzheimer’s disease: A lagged functional connectivity study. PLoS ONE 2012, 7, e46289. [Google Scholar] [CrossRef]
- Hata, M.; Kazui, H.; Tanaka, T.; Ishii, R.; Canuet, L.; Pascual-Marqui, R.D.; Aoki, Y.; Ikeda, S.; Kanemoto, H.; Yoshiyama, K.; et al. Functional connectivity assessed by resting state eeg correlates with cognitive decline of Alzheimer’s disease—An eloreta study. Clin. Neurophysiol. 2016, 127, 1269–1278. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kitaura, Y.; Nishida, K.; Yoshimura, M.; Mii, H.; Katsura, K.; Ueda, S.; Ikeda, S.; Pascual-Marqui, R.D.; Ishii, R.; Kinoshita, T. Functional localization and effective connectivity of cortical theta and alpha oscillatory activity during an attention task. Clin. Neurophysiol. Pract. 2017, 2, 193–200. [Google Scholar] [CrossRef] [PubMed]
- Di Lorenzo, G.; Daverio, A.; Ferrentino, F.; Santarnecchi, E.; Ciabattini, F.; Monaco, L.; Lisi, G.; Barone, Y.; Di Lorenzo, C.; Niolu, C.; et al. Altered resting-state eeg source functional connectivity in schizophrenia: The effect of illness duration. Front. Hum. Neurosci. 2015, 9, 234. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Imperatori, C.; Fabbricatore, M.; Innamorati, M.; Farina, B.; Quintiliani, M.I.; Lamis, D.A.; Mazzucchi, E.; Contardi, A.; Vollono, C.; Della Marca, G. Modification of eeg functional connectivity and eeg power spectra in overweight and obese patients with food addiction: An eloreta study. Brain Imaging Behav. 2015, 9, 703–716. [Google Scholar] [CrossRef] [PubMed]
- Pascual-Marqui, R.D.; Lehmann, D.; Koukkou, M.; Kochi, K.; Anderer, P.; Saletu, B.; Tanaka, H.; Hirata, K.; John, E.R.; Prichep, L.; et al. Assessing interactions in the brain with exact low-resolution electromagnetic tomography. Philos. Trans. A Math. Phys. Eng. Sci. 2011, 369, 3768–3784. [Google Scholar] [CrossRef]
- Fuchs, M.; Kastner, J.; Wagner, M.; Hawes, S.; Ebersole, J.S. A standardized boundary element method volume conductor model. Clin. Neurophysiol. 2002, 113, 702–712. [Google Scholar] [CrossRef] [Green Version]
- Mazziotta, J.; Toga, A.; Evans, A.; Fox, P.; Lancaster, J.; Zilles, K.; Woods, R.; Paus, T.; Simpson, G.; Pike, B.; et al. A probabilistic atlas and reference system for the human brain: International consortium for brain mapping (ICBM). Philos. Trans. R. Soc. Lond. B Biol. Sci. 2001, 356, 1293–1322. [Google Scholar] [CrossRef]
- Jurcak, V.; Tsuzuki, D.; Dan, I. 10/20, 10/10, and 10/5 systems revisited: Their validity as relative head-surface-based positioning systems. Neuroimage 2007, 34, 1600–1611. [Google Scholar] [CrossRef]
- Oostenveld, R.; Praamstra, P. The five percent electrode system for high-resolution eeg and erp measurements. Clin. Neurophysiol. 2001, 112, 713–719. [Google Scholar] [CrossRef]
- Lancaster, J.L.; Woldorff, M.G.; Parsons, L.M.; Liotti, M.; Freitas, C.S.; Rainey, L.; Kochunov, P.V.; Nickerson, D.; Mikiten, S.A.; Fox, P.T. Automated talairach atlas labels for functional brain mapping. Hum. Brain Mapp. 2000, 10, 120–131. [Google Scholar] [CrossRef]
- Hata, M.; Hayashi, N.; Ishii, R.; Canuet, L.; Pascual-Marqui, R.D.; Aoki, Y.; Ikeda, S.; Sakamoto, T.; Iwata, M.; Kimura, K.; et al. Short-Term meditation modulates eeg activity in subjects with post-traumatic residual disabilities. Clin. Neurophysiol. Pract. 2019, 4, 30–36. [Google Scholar] [CrossRef] [PubMed]
- Jatoi, M.A.; Kamel, N.; Malik, A.S.; Faye, I. Eeg based brain source localization comparison of sloreta and eloreta. Australas. Phys. Eng. Sci. Med. 2014, 37, 713–721. [Google Scholar] [CrossRef] [PubMed]
- Takahashi, H.; Rissling, A.J.; Pascual-Marqui, R.; Kirihara, K.; Pela, M.; Sprock, J.; Braff, D.L.; Light, G.A. Neural substrates of normal and impaired preattentive sensory discrimination in large cohorts of nonpsychiatric subjects and schizophrenia patients as indexed by mmn and p3a change detection responses. Neuroimage 2013, 66, 594–603. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Michel, C.M.; Brunet, D. Eeg source imaging: A practical review of the analysis steps. Front. Neurol. 2019, 10, 325. [Google Scholar] [CrossRef] [Green Version]
- De Ridder, D.; Vanneste, S.; Kovacs, S.; Sunaert, S.; Dom, G. Transient alcohol craving suppression by rtms of dorsal anterior cingulate: An fmri and loreta eeg study. Neurosci. Lett. 2011, 496, 5–10. [Google Scholar] [CrossRef]
- Huang, Y.; Mohan, A.; De Ridder, D.; Sunaert, S.; Vanneste, S. The neural correlates of the unified percept of alcohol-related craving: A fmri and eeg study. Sci. Rep. 2018, 8, 923. [Google Scholar] [CrossRef]
- Kirino, E. Three-Dimensional stereotactic surface projection in the statistical analysis of single photon emission computed tomography data for distinguishing between alzheimer’s disease and depression. World J. Psychiatry 2017, 7, 121–127. [Google Scholar] [CrossRef]
- Muller, T.J.; Federspiel, A.; Horn, H.; Lovblad, K.; Lehmann, C.; Dierks, T.; Strik, W.K. The neurophysiological time pattern of illusionary visual perceptual transitions: A simultaneous eeg and fmri study. Int. J. Psychophysiol. 2005, 55, 299–312. [Google Scholar] [CrossRef]
- Horacek, J.; Brunovsky, M.; Novak, T.; Skrdlantova, L.; Klirova, M.; Bubenikova-Valesova, V.; Krajca, V.; Tislerova, B.; Kopecek, M.; Spaniel, F.; et al. Effect of low-frequency rtms on electromagnetic tomography (loreta) and regional brain metabolism (pet) in schizophrenia patients with auditory hallucinations. Neuropsychobiology 2007, 55, 132–142. [Google Scholar] [CrossRef]
- Pizzagalli, D.A.; Oakes, T.R.; Fox, A.S.; Chung, M.K.; Larson, C.L.; Abercrombie, H.C.; Schaefer, S.M.; Benca, R.M.; Davidson, R.J. Functional but not structural subgenual prefrontal cortex abnormalities in melancholia. Mol. Psychiatry 2004, 9, 393–405. [Google Scholar] [CrossRef] [Green Version]
- Zumsteg, D.; Wennberg, R.A.; Treyer, V.; Buck, A.; Wieser, H.G. H2(15)o or 13nh3 pet and electromagnetic tomography (loreta) during partial status epilepticus. Neurology 2005, 65, 1657–1660. [Google Scholar] [CrossRef] [PubMed]
- Canuet, L.; Ishii, R.; Pascual-Marqui, R.D.; Iwase, M.; Kurimoto, R.; Aoki, Y.; Ikeda, S.; Takahashi, H.; Nakahachi, T.; Takeda, M. Resting-State eeg source localization and functional connectivity in schizophrenia-like psychosis of epilepsy. PLoS ONE 2011, 6, e27863. [Google Scholar] [CrossRef] [Green Version]
- Todder, D.; Levine, J.; Abujumah, A.; Mater, M.; Cohen, H.; Kaplan, Z. The quantitative electroencephalogram and the low-resolution electrical tomographic analysis in posttraumatic stress disorder. Clin. EEG Neurosci. 2012, 43, 48–53. [Google Scholar] [CrossRef] [PubMed]
- Toppi, J.; Borghini, G.; Petti, M.; He, E.J.; De Giusti, V.; He, B.; Astolfi, L.; Babiloni, F. Investigating cooperative behavior in ecological settings: An eeg hyperscanning study. PLoS ONE 2016, 11, e0154236. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pascual-Marqui, R.D. Coherence and phase synchronization: Generalization to pairs of multivariate time series, and removal of zero-lag contributions. arXiv 2007, arXiv:0706.1776v3. Available online: http://arxiv.org/pdf/0706.1776 (accessed on 12 July 2007).
- Olbrich, S.; Trankner, A.; Chittka, T.; Hegerl, U.; Schonknecht, P. Functional connectivity in major depression: Increased phase synchronization between frontal cortical eeg-source estimates. Psychiatry Res. 2014, 222, 91–99. [Google Scholar] [CrossRef]
- Palva, J.M.; Wang, S.H.; Palva, S.; Zhigalov, A.; Monto, S.; Brookes, M.J.; Schoffelen, J.M.; Jerbi, K. Ghost interactions in meg/eeg source space: A note of caution on inter-areal coupling measures. Neuroimage 2018, 173, 632–643. [Google Scholar] [CrossRef]
- Pagani, M.; Di Lorenzo, G.; Verardo, A.R.; Nicolais, G.; Monaco, L.; Lauretti, G.; Russo, R.; Niolu, C.; Ammaniti, M.; Fernandez, I.; et al. Neurobiological correlates of emdr monitoring—An eeg study. PLoS ONE 2012, 7, e45753. [Google Scholar] [CrossRef] [Green Version]
- Takamiya, A.; Hirano, J.; Yamagata, B.; Takei, S.; Kishimoto, T.; Mimura, M. Electroconvulsive therapy modulates resting-state eeg oscillatory pattern and phase synchronization in nodes of the default mode network in patients with depressive disorder. Front. Hum. Neurosci. 2019, 13, 1. [Google Scholar] [CrossRef] [Green Version]
- Ramyead, A.; Kometer, M.; Studerus, E.; Koranyi, S.; Ittig, S.; Gschwandtner, U.; Fuhr, P.; Riecher-Rossler, A. Aberrant current source-density and lagged phase synchronization of neural oscillations as markers for emerging psychosis. Schizophr. Bull. 2015, 41, 919–929. [Google Scholar] [CrossRef]
- Pascual-Marqui, R.D.; Michel, C.M.; Lehmann, D. Low resolution electromagnetic tomography: A new method for localizing electrical activity in the brain. Int. J. Psychophysiol. 1994, 18, 49–65. [Google Scholar] [CrossRef]
- Pascual-Marqui, R.D.; Michel, C.M.; Lehmann, D. Segmentation of brain electrical activity into microstates: Model estimation and validation. IEEE Trans. Biomed. Eng. 1995, 42, 658–665. [Google Scholar] [CrossRef] [PubMed]
- Nichols, T.E.; Holmes, A.P. Nonparametric permutation tests for functional neuroimaging: A primer with examples. Hum. Brain Mapp. 2002, 15, 1–25. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Winkler, A.M.; Webster, M.A.; Brooks, J.C.; Tracey, I.; Smith, S.M.; Nichols, T.E. Non-Parametric combination and related permutation tests for neuroimaging. Hum. Brain Mapp. 2016, 37, 1486–1511. [Google Scholar] [CrossRef] [Green Version]
- Cohen, J. Statistical Power Analysis for the Behavioral Sciences, 2nd ed.; Erlbaum: Hillsdale, NJ, USA, 1988. [Google Scholar]
- Cocchi, L.; Zalesky, A.; Fornito, A.; Mattingley, J.B. Dynamic cooperation and competition between brain systems during cognitive control. Trends Cogn. Sci. 2013, 17, 493–501. [Google Scholar] [CrossRef]
- Elton, A.; Gao, W. Divergent task-dependent functional connectivity of executive control and salience networks. Cortex 2014, 51, 56–66. [Google Scholar] [CrossRef]
- Heilbronner, S.R.; Hayden, B.Y. Dorsal anterior cingulate cortex: A bottom-up view. Annu. Rev. Neurosci. 2016, 39, 149–170. [Google Scholar] [CrossRef] [Green Version]
- Kuhn, S.; Gallinat, J. Common biology of craving across legal and illegal drugs—A quantitative meta-analysis of cue-reactivity brain response. Eur. J. Neurosci. 2011, 33, 1318–1326. [Google Scholar] [CrossRef]
- Sestieri, C.; Shulman, G.L.; Corbetta, M. The contribution of the human posterior parietal cortex to episodic memory. Nat. Rev. Neurosci. 2017, 18, 183–192. [Google Scholar] [CrossRef] [Green Version]
- Knyazev, G.G. Motivation, emotion, and their inhibitory control mirrored in brain oscillations. Neurosci. Biobehav. Rev. 2007, 31, 377–395. [Google Scholar] [CrossRef]
- Knyazev, G.G. Eeg delta oscillations as a correlate of basic homeostatic and motivational processes. Neurosci. Biobehav. Rev. 2012, 36, 677–695. [Google Scholar] [CrossRef] [PubMed]
- Reid, M.S.; Prichep, L.S.; Ciplet, D.; O’Leary, S.; Tom, M.; Howard, B.; Rotrosen, J.; John, E.R. Quantitative electroencephalographic studies of cue-induced cocaine craving. Clin. Electroencephalogr. 2003, 34, 110–123. [Google Scholar] [CrossRef]
- Reid, M.S.; Flammino, F.; Howard, B.; Nilsen, D.; Prichep, L.S. Topographic imaging of quantitative eeg in response to smoked cocaine self-administration in humans. Neuropsychopharmacology 2006, 31, 872–884. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, X.; Ma, R.; Pang, L.; Lv, W.; Xie, Y.; Chen, Y.; Zhang, P.; Chen, J.; Wu, Q.; Cui, G.; et al. Delta coherence in resting-state eeg predicts the reduction in cigarette craving after hypnotic aversion suggestions. Sci. Rep. 2017, 7, 2430. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Parvaz, M.A.; Alia-Klein, N.; Woicik, P.A.; Volkow, N.D.; Goldstein, R.Z. Neuroimaging for drug addiction and related behaviors. Rev. Neurosci. 2011, 22, 609–624. [Google Scholar] [CrossRef] [Green Version]
- Satre, D.D.; Bahorik, A.; Zaman, T.; Ramo, D. Psychiatric disorders and comorbid cannabis use: How common is it and what is the clinical impact? J. Clin. Psychiatry 2018, 79. [Google Scholar] [CrossRef] [Green Version]



{kind=link}
{kind=link}
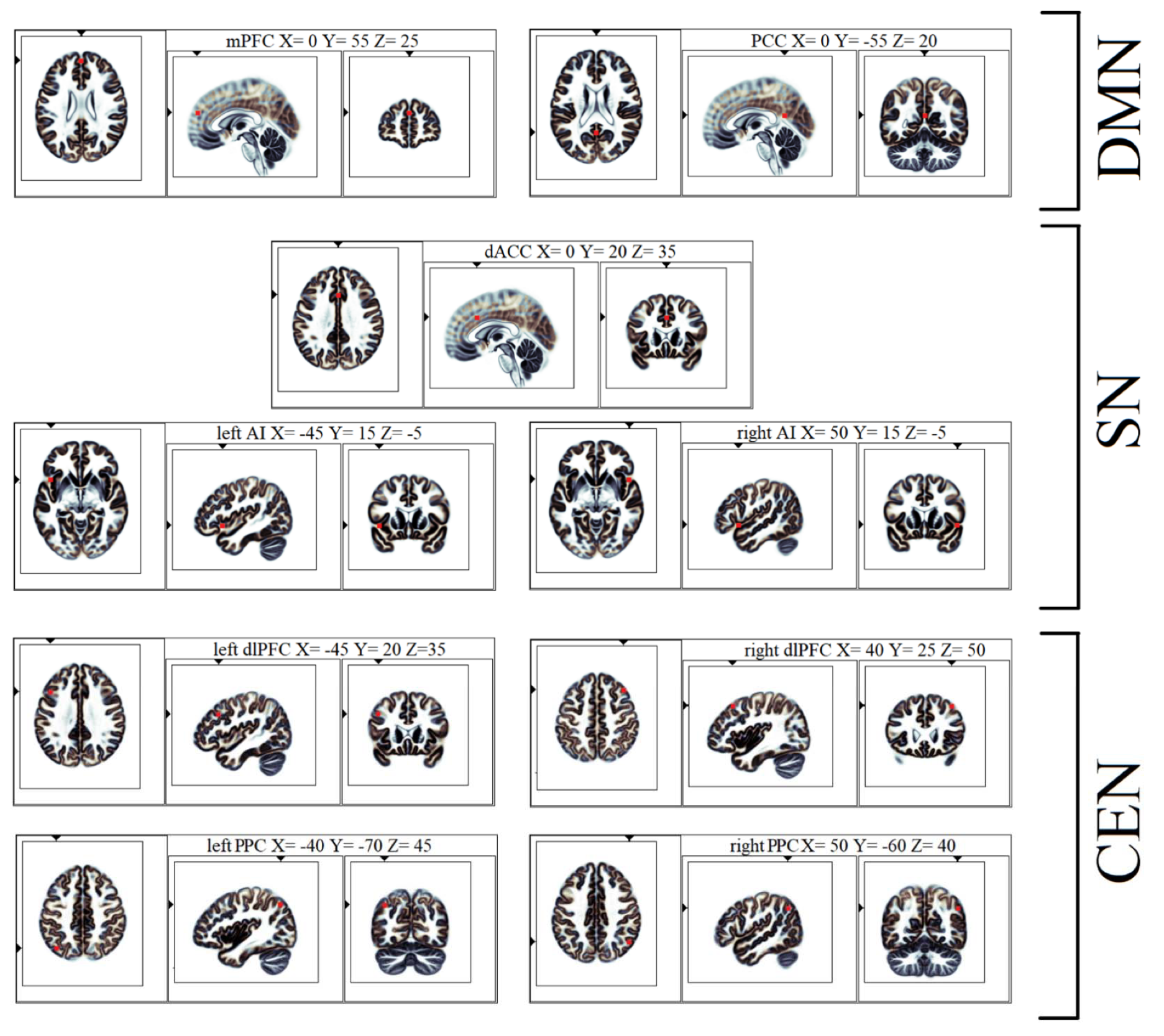
| Brain Network | Anatomical Structure | eLORETA MNI Coordinates 1 eLORETA Talairach Coordinates 1 | ||
|---|---|---|---|---|
| x | y | z | ||
| DMN | mPFC | 0 | 55 | 25 |
| 0 | 54 | 20 | ||
| PCC | 0 | −55 | 20 | |
| 0 | −52 | 21 | ||
| SN | dACC | 0 | 20 | 35 |
| 0 | 21 | 31 | ||
| Left AI | −45 | 15 | −5 | |
| −45 | 14 | −5 | ||
| Right AI | 50 | 15 | −5 | |
| 50 | 14 | −5 | ||
| CEN | Left dlPFC | −45 | 20 | 35 |
| −45 | 21 | 31 | ||
| Right dlPFC | 40 | 25 | 50 | |
| 40 | 27 | 45 | ||
| Left PPC | −40 | −70 | 45 | |
| −40 | −66 | 45 | ||
| Right PPC | 50 | −60 | 40 | |
| 50 | −56 | 40 | ||
| PCU (N = 12) | Non-PCU (N = 24) | test | p | |
|---|---|---|---|---|
| Variables | ||||
| Age–M (SD) | 23.33 ± 3.47 | 21.21 ± 2.70 | Z-test = 1.06 | 0.211 |
| Educational level (years)–M ± SD | 16.42 ± 1.51 | 15.54 ± 1.50 | Z-test = 0.83 | 0.504 |
| Men–N (%) | 7 (58.3%) | 9 (37.5%) | χ21 = 1.41 | 0.236 |
| Tobacco use in the last 6 months–N (%) | 8 (66.7%) | 7 (29.2%) | χ21 = 4.63 | 0.031 |
| CAST–M (SD) | 10.25 ± 4.31 | 0.00 ± 0.00 | - | - |
| CAGE–M (SD) | 0.67 ± 1.07 | 0.04 ± 0.20 | Z-test = 0.82 | 0.504 |
| CAGE ≥ 2–N (%) | 3 (25%) | 0 (0%) | χ21 = 6.55 | 0.011 |
| SCL-K-9–M (SD) | 1.22 ± 0.97 | 0.73 ± 0.44 | Z-test = 0.94 | 0.336 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Imperatori, C.; Massullo, C.; Carbone, G.A.; Panno, A.; Giacchini, M.; Capriotti, C.; Lucarini, E.; Ramella Zampa, B.; Murillo-Rodríguez, E.; Machado, S.; et al. Increased Resting State Triple Network Functional Connectivity in Undergraduate Problematic Cannabis Users: A Preliminary EEG Coherence Study. Brain Sci. 2020, 10, 136. https://doi.org/10.3390/brainsci10030136
Imperatori C, Massullo C, Carbone GA, Panno A, Giacchini M, Capriotti C, Lucarini E, Ramella Zampa B, Murillo-Rodríguez E, Machado S, et al. Increased Resting State Triple Network Functional Connectivity in Undergraduate Problematic Cannabis Users: A Preliminary EEG Coherence Study. Brain Sciences. 2020; 10(3):136. https://doi.org/10.3390/brainsci10030136
Chicago/Turabian StyleImperatori, Claudio, Chiara Massullo, Giuseppe Alessio Carbone, Angelo Panno, Marta Giacchini, Cristina Capriotti, Elisa Lucarini, Benedetta Ramella Zampa, Eric Murillo-Rodríguez, Sérgio Machado, and et al. 2020. "Increased Resting State Triple Network Functional Connectivity in Undergraduate Problematic Cannabis Users: A Preliminary EEG Coherence Study" Brain Sciences 10, no. 3: 136. https://doi.org/10.3390/brainsci10030136
APA StyleImperatori, C., Massullo, C., Carbone, G. A., Panno, A., Giacchini, M., Capriotti, C., Lucarini, E., Ramella Zampa, B., Murillo-Rodríguez, E., Machado, S., & Farina, B. (2020). Increased Resting State Triple Network Functional Connectivity in Undergraduate Problematic Cannabis Users: A Preliminary EEG Coherence Study. Brain Sciences, 10(3), 136. https://doi.org/10.3390/brainsci10030136