The Brain Differentially Prepares Inner and Overt Speech Production: Electrophysiological and Vascular Evidence


Abstract
:1. Introduction
1.1. Inner Versus Overt Speech
1.2. Speech Preparation Versus Execution
1.3. The Present Study
2. Materials and Methods
2.1. Participants
2.2. Material
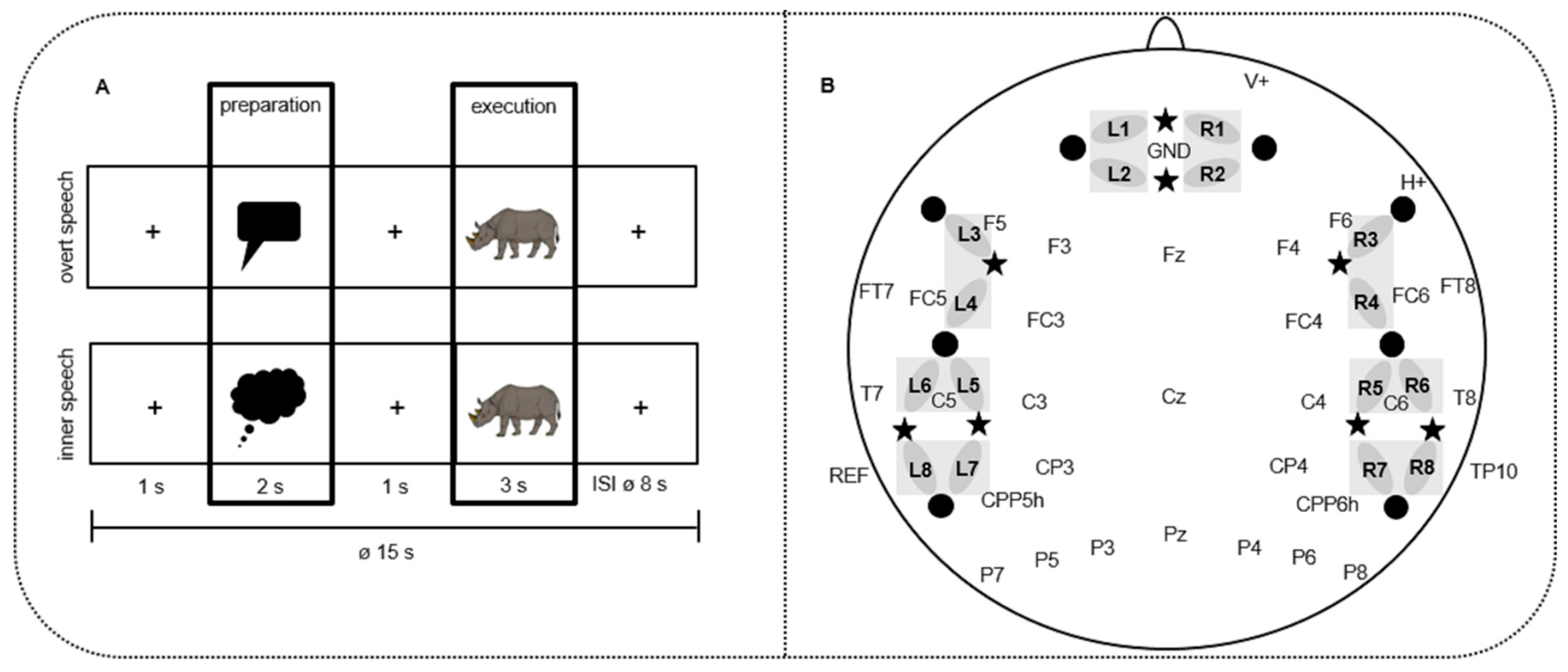
2.3. Tasks and Procedure
2.4. NIRS/EEG Data Recordings
2.4.1. fNIRS Data Recording
2.4.2. EEG Data Recording
2.5. Data Analyses
2.5.1. fNIRS Data Analyses
2.5.2. EEG Data Analyses
3. Results
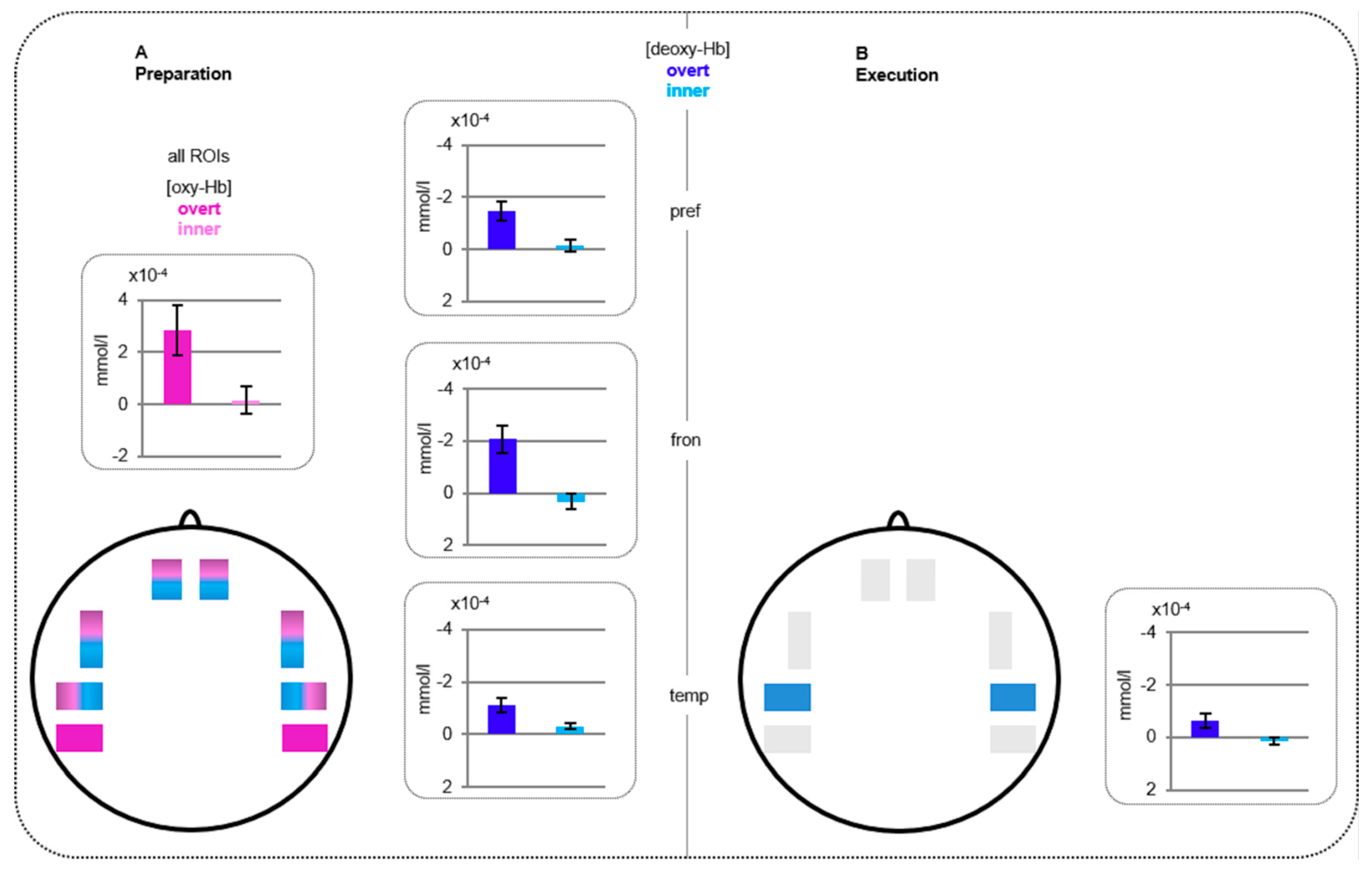
3.1. fNIRS Results
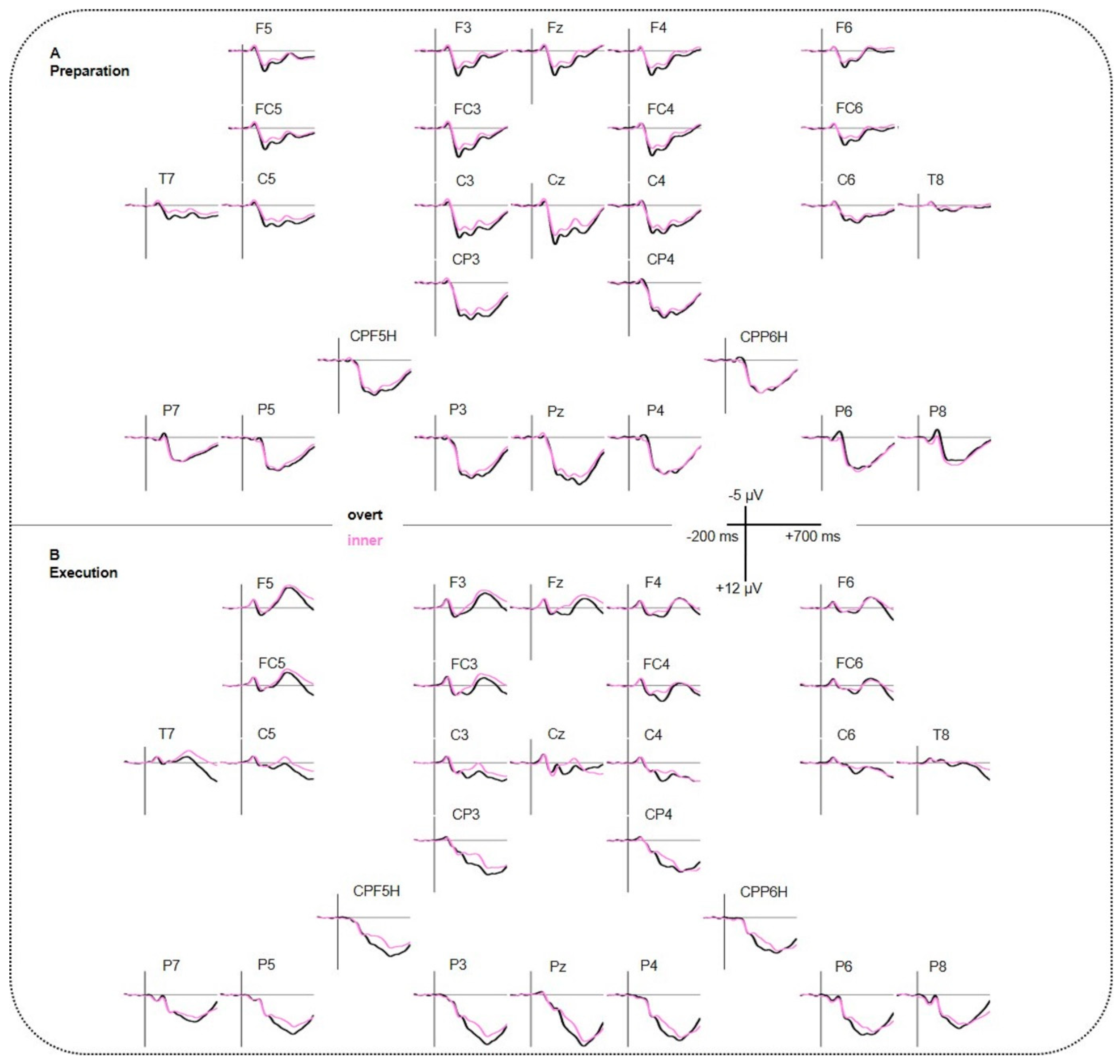
3.2. EEG Results
4. Discussion
4.1. Brain Areas Underlying Inner and Overt Speech in the Speech Execution Phase
4.2. Timing Characteristics of Inner and Overt Speech in the Speech Execution Phase
4.3. Inner and Overt Speech in the Preparation Versus Execution Phase
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Alderson-Day, B.; Fernyhough, C. Inner Speech: Development, Cognitive Functions, Phenomenology, and Neurobiology. Psychol. Bull. 2015, 141, 931–965. [Google Scholar] [CrossRef] [Green Version]
- Carota, F.; Posada, A.; Harquel, S.; Delpuech, C.; Bertrand, O.; Sirigu, A. Neural dynamics of the intention to speak. Cereb. Cortex 2010, 20, 1891–1897. [Google Scholar] [CrossRef] [Green Version]
- Gehrig, J.; Wibral, M.; Arnold, C.; Kell, C.A. Setting Up the Speech Production Network: How Oscillations Contribute to Lateralized Information Routing. Front. Psychol. 2012, 3, 169. [Google Scholar] [CrossRef] [Green Version]
- Hickok, G.; Houde, J.; Rong, F. Sensorimotor Integration in Speech Processing: Computational Basis and Neural Organization. Neuron 2011, 69, 407–422. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Perrone-Bertolotti, M.; Rapin, L.; Lachaux, J.P.; Baciu, M.; Lœvenbruck, H. What is that little voice inside my head? Inner speech phenomenology, its role in cognitive performance, and its relation to self-monitoring. Behav. Brain Res. 2014, 261, 220–239. [Google Scholar] [CrossRef] [PubMed]
- Indefrey, P. The Spatial and Temporal Signatures of Word Production Components: A Critical Update. Front. Psychol. 2011, 2, 255. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hickok, G. Computational neuroanatomy of speech production. Nat. Rev. Neurosci. 2012, 13, 135–145. [Google Scholar] [CrossRef]
- Rossi, S.; Telkemeyer, S.; Wartenburger, I.; Obrig, H. Shedding light on words and sentences: Near-infrared spectroscopy in language research. Brain Lang. 2012, 121, 152–163. [Google Scholar] [CrossRef]
- Vygotsky. Thought and Language; The MIT Press: Cambridge, MA, USA, 1986. [Google Scholar]
- Watson, J.B. Psychology as the behaviorist views it. Psychol. Rev. 1913, 20, 158–177. [Google Scholar] [CrossRef] [Green Version]
- Oppenheim, G.M.; Dell, G.S. Inner speech slips exhibit lexical bias, but not the phonemic similarity effect. Cognition 2008, 106, 528–537. [Google Scholar] [CrossRef] [Green Version]
- Dell, G.S.; Repka, R.J. Errors in inner speech. In Experimental Slips and Human Error: Exploring the Architecture of Volition; Cognition and Language; Plenum Press: New York, NY, USA, 1992; pp. 237–262. ISBN 978-0-306-43866-0. [Google Scholar]
- Abramson, M.; Goldinger, S.D. What the reader’s eye tells the mind’s ear: Silent reading activates inner speech. Percept. Psychophys. 1997, 59, 1059–1068. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brocklehurst, P.H.; Corley, M. Investigating the inner speech of people who stutter: Evidence for (and against) the covert repair hypothesis. J. Commun. Disord. 2011, 44, 246–260. [Google Scholar] [CrossRef] [Green Version]
- Corley, M.; Brocklehurst, P.H.; Moat, H.S. Error biases in inner and overt speech: Evidence from tongue twisters. J. Exp. Psychol. Learn. Mem. Cogn. 2011, 37, 162–175. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Levelt, W.J.M. Speaking: From Intention to Articulation; The MIT Press: Cambridge, MA, USA, 1989; ISBN 978-0-262-12137-8. [Google Scholar]
- Indefrey, P.; Levelt, W.J.M. The spatial and temporal signatures of word production components. Cognition 2004, 92, 101–144. [Google Scholar] [CrossRef] [Green Version]
- Indefrey, P.; Levelt, W.J.M. The neural correlates of language production. In The New Cognitive Neurosciences, 2nd ed.; Gazzaniga, M.S., Ed.; The MIT Press: Cambridge, MA, USA, 2000; pp. 845–865. ISBN 978-0-262-07195-6. [Google Scholar]
- Levelt, W.J.M. Monitoring and self-repair in speech. Cognition 1983, 14, 41–104. [Google Scholar] [CrossRef] [Green Version]
- Levelt, W.J.M. The ability to speak: From intentions to spoken words. Eur. Rev. 1995, 3, 13–23. [Google Scholar] [CrossRef] [Green Version]
- Levelt, W.J.; Roelofs, A.; Meyer, A.S. A theory of lexical access in speech production. Behav. Brain Sci. 1999, 22, 1–38; Discussion 38–75. [Google Scholar] [CrossRef] [Green Version]
- Kielar, A.; Milman, L.; Bonakdarpour, B.; Thompson, C.K. Neural correlates of covert and overt production of tense and agreement morphology: Evidence from fMRI. J. Neurolinguist. 2011, 24, 183–201. [Google Scholar] [CrossRef] [Green Version]
- Palmer, E.D.; Rosen, H.J.; Ojemann, J.G.; Buckner, R.L.; Kelley, W.M.; Petersen, S.E. An event-related fMRI study of overt and covert word stem completion. Neuroimage 2001, 14, 182–193. [Google Scholar] [CrossRef] [Green Version]
- Rosen, H.J.; Ojemann, J.G.; Ollinger, J.M.; Petersen, S.E. Comparison of Brain Activation during Word Retrieval Done Silently and Aloud Using fMRI. Brain Cogn. 2000, 42, 201–217. [Google Scholar] [CrossRef] [Green Version]
- Shuster, L.I.; Lemieux, S.K. An fMRI investigation of covertly and overtly produced mono- and multisyllabic words. Brain Lang. 2005, 93, 20–31. [Google Scholar] [CrossRef] [PubMed]
- Huang, J.; Carr, T.H.; Cao, Y. Comparing cortical activations for silent and overt speech using event-related fMRI. Hum. Brain Mapp. 2002, 15, 39–53. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Borowsky, R.; Owen, W.J.; Wile, T.L.; Friesen, C.K.; Martin, J.L.; Sarty, G.E. Neuroimaging of Language Processes: fMRI of Silent and Overt Lexical Processing and the Promise of Multiple Process Imaging in Single Brain Studies. Can. Assoc. Radiol. J. 2005, 56, 204–213. [Google Scholar]
- Moriai-Izawa, A.; Dan, H.; Dan, I.; Sano, T.; Oguro, K.; Yokota, H.; Tsuzuki, D.; Watanabe, E. Multichannel fNIRS assessment of overt and covert confrontation naming. Brain Lang. 2012, 121, 185–193. [Google Scholar] [CrossRef] [PubMed]
- Bookheimer, S.Y.; Zeffiro, T.A.; Blaxton, T.; Gaillard, W.; Theodore, W. Regional cerebral blood flow during object naming and word reading. Hum. Brain Mapp. 1995, 3, 93–106. [Google Scholar] [CrossRef]
- Basho, S.; Palmer, E.D.; Rubio, M.A.; Wulfeck, B.; Müller, R.-A. Effects of generation mode in fMRI adaptations of semantic fluency: Paced production and overt speech. Neuropsychologia 2007, 45, 1697–1706. [Google Scholar] [CrossRef] [Green Version]
- Kell, C.A.; Morillon, B.; Kouneiher, F.; Giraud, A.-L. Lateralization of speech production starts in sensory cortices--a possible sensory origin of cerebral left dominance for speech. Cereb. Cortex 2011, 21, 932–937. [Google Scholar] [CrossRef]
- Kell, C.A. Funktionelle Bildgebung physiologischer und pathologischer Sprachproduktion. Nervenarzt 2014, 85, 701–707. [Google Scholar] [CrossRef]
- Jurado, M.B.; Rosselli, M. The elusive nature of executive functions: A review of our current understanding. Neuropsychol. Rev. 2007, 17, 213–233. [Google Scholar] [CrossRef]
- Garon, N.; Bryson, S.E.; Smith, I.M. Executive function in preschoolers: A review using an integrative framework. Psychol. Bull. 2008, 134, 31–60. [Google Scholar] [CrossRef] [Green Version]
- Miller, E.K.; Cohen, J.D. An integrative theory of prefrontal cortex function. Annu. Rev. Neurosci. 2001, 24, 167–202. [Google Scholar] [CrossRef]
- Baddeley, A.D.; Hitch, G. Working Memory. In Psychology of Learning and Motivation; Bower, G.H., Ed.; Academic Press: Cambridge, MA, USA, 1974; Volume 8, pp. 47–89. [Google Scholar]
- Norman, D.A.; Shallice, T. Attention to Action. In Consciousness and Self-Regulation: Advances in Research and Theory; Davidson, R.J., Schwartz, G.E., Shapiro, D., Eds.; Springer: Boston, MA, USA, 1986; Volume 4, pp. 1–18. ISBN 978-1-4757-0629-1. [Google Scholar]
- Anderson, V.; Northam, E.; Wrennall, J.; Northam, E.; Wrennall, J. Developmental Neuropsychology: A Clinical Approach; Routledge: Abingdon, UK, 2018; ISBN 978-0-203-79912-3. [Google Scholar]
- Hobson, P.; Leeds, L. Executive functioning in older people. Rev. Clin. Gerontol. 2001, 11, 361–372. [Google Scholar] [CrossRef]
- Piguet, O.; Grayson, D.A.; Broe, G.A.; Tate, R.L.; Bennett, H.P.; Lye, T.C.; Creasey, H.; Ridley, L. Normal aging and executive functions in “old-old” community dwellers: Poor performance is not an inevitable outcome. Int. Psychogeriatr. 2002, 14, 139–159. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Elliott, R. Executive functions and their disorders. Br. Med. Bull. 2003, 65, 49–59. [Google Scholar] [CrossRef] [PubMed]
- Miyake, A.; Friedman, N.P.; Emerson, M.J.; Witzki, A.H.; Howerter, A.; Wager, T.D. The unity and diversity of executive functions and their contributions to complex “Frontal Lobe” tasks: A latent variable analysis. Cogn. Psychol. 2000, 41, 49–100. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Funahashi, S. Neuronal mechanisms of executive control by the prefrontal cortex. Neurosci. Res. 2001, 39, 147–165. [Google Scholar] [CrossRef]
- Obrig, H.; Villringer, A. Beyond the visible--imaging the human brain with light. J. Cereb. Blood Flow Metab. 2003, 23, 1–18. [Google Scholar] [CrossRef] [Green Version]
- Wallois, F.; Mahmoudzadeh, M.; Patil, A.; Grebe, R. Usefulness of simultaneous EEG-NIRS recording in language studies. Brain Lang. 2012, 121, 110–123. [Google Scholar] [CrossRef]
- De Zubicaray, G.I.; Wilson, S.J.; McMahon, K.L.; Muthiah, S. The semantic interference effect in the picture-word paradigm: An event-related fMRI study employing overt responses. Hum. Brain Mapp. 2001, 14, 218–227. [Google Scholar] [CrossRef]
- Lloyd-Fox, S.; Blasi, A.; Elwell, C.E. Illuminating the developing brain: The past, present and future of functional near infrared spectroscopy. Neurosci. Biobehav. Rev. 2010, 34, 269–284. [Google Scholar] [CrossRef]
- Costa, A.; Strijkers, K.; Martin, C.; Thierry, G. The time course of word retrieval revealed by event-related brain potentials during overt speech. Proc. Natl. Acad. Sci. USA 2009, 106, 21442–21446. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hull, R.; Bortfeld, H.; Koons, S. Near-Infrared Spectroscopy and Cortical Responses to Speech Production. Open Neuroimaging J. 2009, 3, 26–30. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Strijkers, K.; Costa, A.; Thierry, G. Tracking lexical access in speech production: Electrophysiological correlates of word frequency and cognate effects. Cereb. Cortex 2010, 20, 912–928. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rossion, B.; Pourtois, G. Revisiting Snodgrass and Vanderwart’s object pictorial set: The role of surface detail in basic-level object recognition. Perception 2004, 33, 217–236. [Google Scholar] [CrossRef] [Green Version]
- Oldfield, R.C. The assessment and analysis of handedness: The Edinburgh inventory. Neuropsychologia 1971, 9, 97–113. [Google Scholar] [CrossRef]
- Snodgrass, J.G.; Vanderwart, M. A standardized set of 260 pictures: Norms for name agreement, image agreement, familiarity, and visual complexity. J. Exp. Psychol. Hum. Learn. Mem. 1980, 6, 174–215. [Google Scholar] [CrossRef]
- Schröder, A.; Kauschke, C.; DeBleser, R. Messungen des Erwerbsalters für konkrete Nomina. Neurolinguitik 2003, 17, 83–114. [Google Scholar]
- Sharbrough, F.; Chatrian, G.E.; Lesser, R.; Luders, H.; Nuwer, M.; Picton, T.W. American Electroencephalographic Society guidelines for standard electrode position nomenclature. J. Clin. Neurophysiol. 1991, 8, 200–202. [Google Scholar]
- Cope, M.; Delpy, D.T.; Reynolds, E.O.; Wray, S.; Wyatt, J.; van der Zee, P. Methods of quantitating cerebral near infrared spectroscopy data. Adv. Exp. Med. Biol. 1988, 222, 183–189. [Google Scholar]
- Boynton, G.M.; Engel, S.A.; Glover, G.H.; Heeger, D.J. Linear Systems Analysis of Functional Magnetic Resonance Imaging in Human V1. J. Neurosci. 1996, 16, 4207–4221. [Google Scholar] [CrossRef]
- Greenhouse, S.W.; Geisser, S. On methods in the analysis of profile data. Psychometrika 1959, 24, 95–112. [Google Scholar] [CrossRef]
- Gratton, G.; Coles, M.; Donchin, E. A new method for off-line removal of ocular artifact. Electroencephalogr. Clin. Neurophysiol. 1983, 468–484. [Google Scholar] [CrossRef]
- Baus, C.; Sebanz, N.; Fuente, V.; de la Branzi, F.M.; Martin, C.; Costa, A. On predicting others’ words: Electrophysiological evidence of prediction in speech production. Cognition 2014, 133, 395–407. [Google Scholar] [CrossRef] [PubMed]
- Eulitz, C.; Hauk, O.; Cohen, R. Electroencephalographic activity over temporal brain areas during phonological encoding in picture naming. Clin. Neurophysiol. 2000, 111, 2088–2097. [Google Scholar] [CrossRef]
- Sakai, K.; Passingham, R.E. Prefrontal set activity predicts rule-specific neural processing during subsequent cognitive performance. J. Neurosci. 2006, 26, 1211–1218. [Google Scholar] [CrossRef]
- McEvoy, L.K.; Pellouchoud, E.; Smith, M.E.; Gevins, A. Neurophysiological signals of working memory in normal aging. Brain Res. Cogn. Brain Res. 2001, 11, 363–376. [Google Scholar] [CrossRef]
- Allison, T.; Begleiter, A.; McCarthy, G.; Roessler, E.; Nobre, A.C.; Spencer, D.D. Electrophysiological studies of color processing in human visual cortex. Electroencephalogr. Clin. Neurophysiol. Potentials Sect. 1993, 88, 343–355. [Google Scholar] [CrossRef]
- Hillyard, S.A.; Münte, T.F. Selective attention to color and location: An analysis with event-related brain potentials. Percept. Psychophys. 1984, 36, 185–198. [Google Scholar] [CrossRef] [Green Version]
- Anllo-Vento, L.; Hillyard, S.A. Selective attention to the color and direction of moving stimuli: Electrophysiological correlates of hierarchical feature selection. Percept. Psychophys. 1996, 58, 191–206. [Google Scholar] [CrossRef] [Green Version]
- Gajewski, P.D.; Falkenstein, M. Effects of task complexity on ERP components in Go/Nogo tasks. Int. J. Psychophysiol. 2013, 87, 273–278. [Google Scholar] [CrossRef]
- Kirmizi-Alsan, E.; Bayraktaroglu, Z.; Gurvit, H.; Keskin, Y.H.; Emre, M.; Demiralp, T. Comparative analysis of event-related potentials during Go/NoGo and CPT: Decomposition of electrophysiological markers of response inhibition and sustained attention. Brain Res. 2006, 1104, 114–128. [Google Scholar] [CrossRef] [PubMed]
- Hanslmayr, S.; Pastötter, B.; Bäuml, K.-H.; Gruber, S.; Wimber, M.; Klimesch, W. The electrophysiological dynamics of interference during the Stroop task. J. Cogn. Neurosci. 2008, 20, 215–225. [Google Scholar] [CrossRef] [PubMed]
- Shang, M.; Debruille, J.B. N400 processes inhibit inappropriately activated representations: Adding a piece of evidence from a high-repetition design. Neuropsychologia 2013, 51, 1989–1997. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rodriguez-Fornells, A.; Schmitt, B.M.; Kutas, M.; Münte, T.F. Electrophysiological estimates of the time course of semantic and phonological encoding during listening and naming. Neuropsychologia 2002, 40, 778–787. [Google Scholar] [CrossRef] [Green Version]
- Oppenheim, G.M.; Dell, G.S. Motor movement matters: The flexible abstractness of inner speech. Mem. Cogn. 2010, 38, 1147–1160. [Google Scholar] [CrossRef]
- Levelt, W.J.M.; Schriefers, H.; Vorberg, D.; Meyer, A.S.; Pechmann, T.; Havinga, J. The time course of lexical access in speech production: A study of picture naming. Psychol. Rev. 1991, 98, 122–142. [Google Scholar] [CrossRef]
- Dell, G.S.; O’Seaghdha, P.G. Stages of lexical access in language production. Cognition 1992, 42, 287–314. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Effect | 100–200 ms | 200–300 ms | 300–500 ms | 500–600 ms |
|---|---|---|---|---|
| Lateral ROIs | ||||
| cond | ns | (1,44):4.35/0.043 | (1,44):14.35/<0.001 | ns |
| cond*phase | ns | ns | ns | ns |
| cond*region | (5,22):14.98/<0.001 | (5,22):9.83/<0.0001 | (5,22): 6.11/0.003 | ns |
| cond*phase*region | ns | (5,22):10.05/<0.001 | (5,22):6.86/<0.001 | ns |
| cond*hemi | ns | ns | ns | ns |
| cond*phase*hemi | ns | ns | ns | ns |
| cond*region*hemi | ns | ns | (5,22): 3.69/0.012 | (5,22):7.91/<0.001 |
| cond*phase*region*hemi | ns | ns | ns | ns |
| Midline ROIs | ||||
| cond | ns | (1,44):8.15/0.007 | (1,44):28.68/<0.0001 | ns |
| cond*phase | ns | ns | ns | ns |
| cond*elecs | (2,88):4.03/0.027 | ns | ns | ns |
| cond*phase*elecs | ns | (2,88):4.38/0.033 | ns | (2,88):4.35/0.026 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Stephan, F.; Saalbach, H.; Rossi, S. The Brain Differentially Prepares Inner and Overt Speech Production: Electrophysiological and Vascular Evidence. Brain Sci. 2020, 10, 148. https://doi.org/10.3390/brainsci10030148
Stephan F, Saalbach H, Rossi S. The Brain Differentially Prepares Inner and Overt Speech Production: Electrophysiological and Vascular Evidence. Brain Sciences. 2020; 10(3):148. https://doi.org/10.3390/brainsci10030148
Chicago/Turabian StyleStephan, Franziska, Henrik Saalbach, and Sonja Rossi. 2020. "The Brain Differentially Prepares Inner and Overt Speech Production: Electrophysiological and Vascular Evidence" Brain Sciences 10, no. 3: 148. https://doi.org/10.3390/brainsci10030148
APA StyleStephan, F., Saalbach, H., & Rossi, S. (2020). The Brain Differentially Prepares Inner and Overt Speech Production: Electrophysiological and Vascular Evidence. Brain Sciences, 10(3), 148. https://doi.org/10.3390/brainsci10030148