Modulation of Fibers to Motor Cortex during Thalamic DBS in Tourette Patients Correlates with Tic Reduction

 ,
,  and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Patients and Study Design
2.2. Stereotactic Surgery
2.3. Fiber Tracking
2.4. Statistical Analysis
3. Results
3.1. Demographic and Clinical Characteristics
3.2. Neuroimaging Analysis
3.2.1. Generation of Fiber Tracts and Cortical Projections
3.2.2. Parcellation of Seed Regions
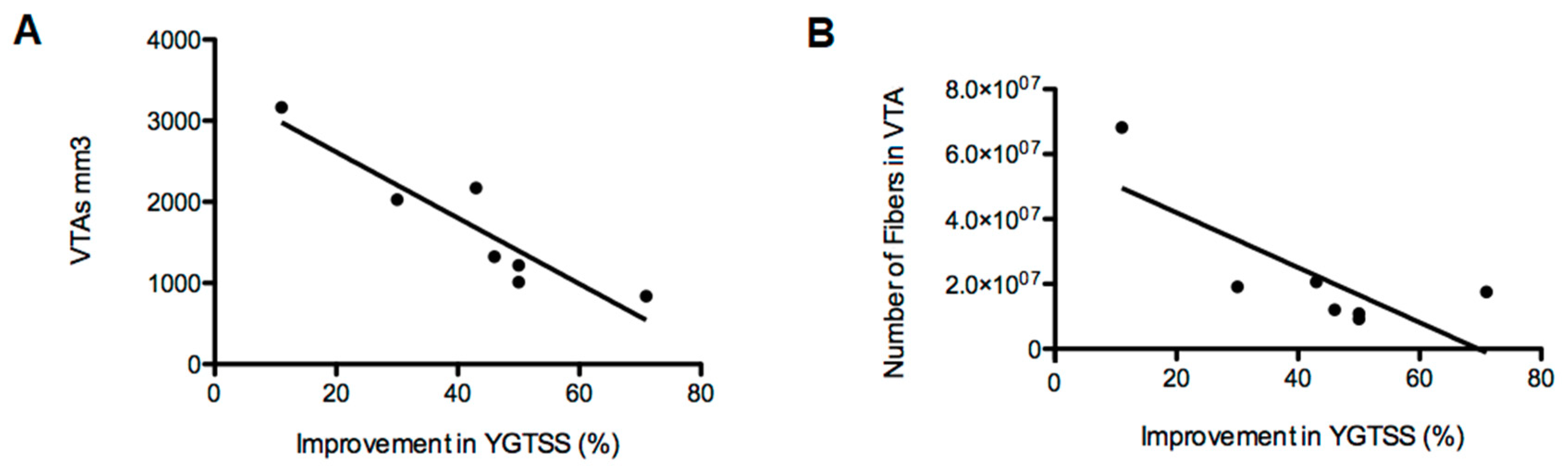
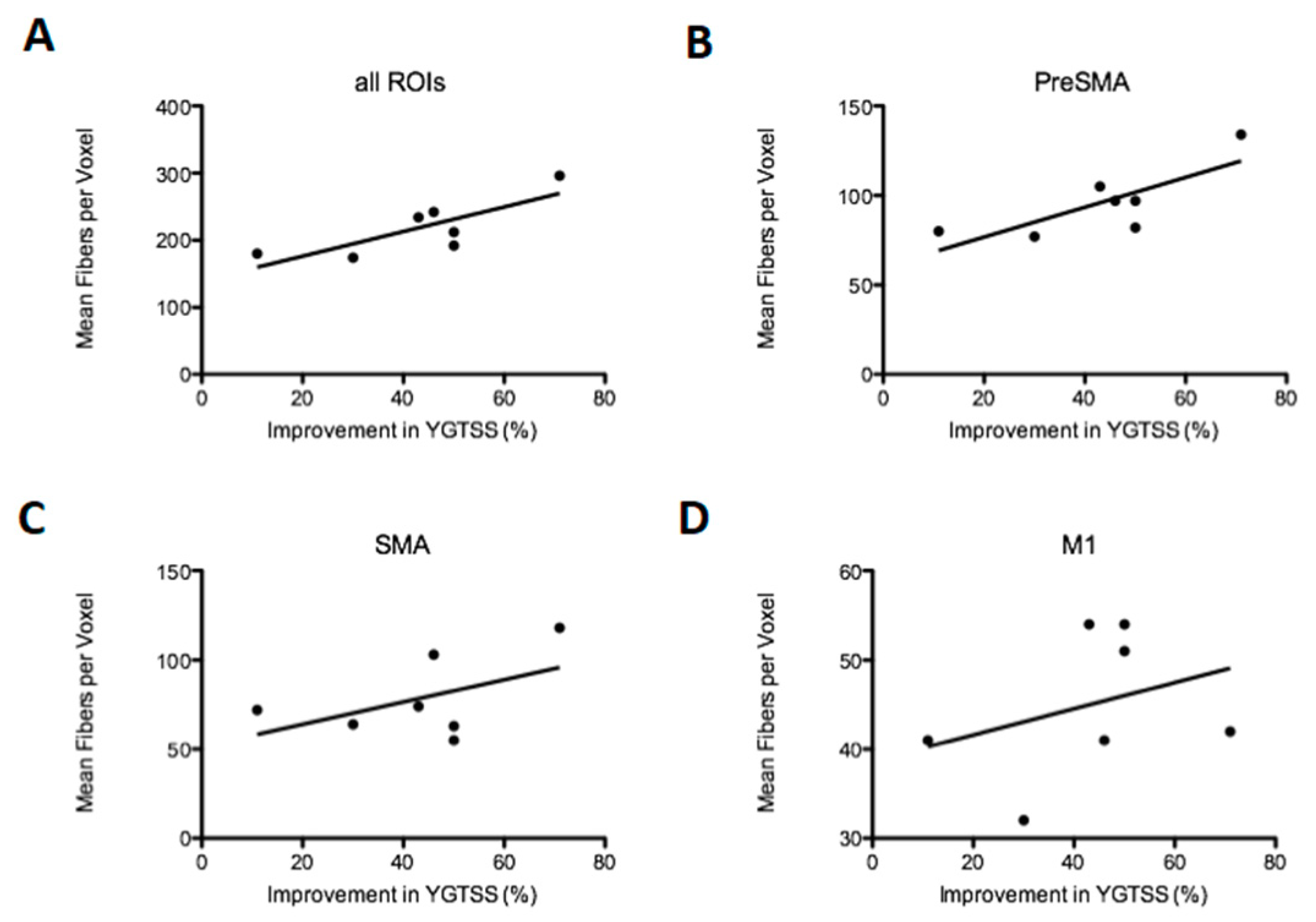
3.2.3. Density of Fibers in the Motor Cortex
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Appendix A


References
- American Psychiatric Association. Diagnostic and Statistical Manual of Mental Disorders, 5th ed.; American Psychiatric Association Publishing: Arlington, VA, USA, 2013. [Google Scholar]
- Heise, K.-F.; Steven, B.; Liuzzi, G.; Thomalla, G.; Jonas, M.; Müller-Vahl, K.; Sauseng, P.; Münchau, A.; Gerloff, C.; Hummel, F.C. Altered modulation of intracortical excitability during movement preparation in Gilles de la Tourette syndrome. Brain 2009, 133, 580–590. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ganos, C.; Roessner, V.; Münchau, A. The functional anatomy of Gilles de la Tourette syndrome. Neurosci. Biobehav. Rev. 2013, 37, 1050–1062. [Google Scholar] [CrossRef] [PubMed]
- Worbe, Y.; Malherbe, C.; Hartmann, A.; Pélégrini-Issac, M.; Messé, A.; Vidailhet, M.; Lehéricy, S.; Benali, H. Functional immaturity of cortico-basal ganglia networks in Gilles de la Tourette syndrome. Brain 2012, 135, 1937–1946. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cheng, B.; Braass, H.; Ganos, C.; Treszl, A.; Biermann-Ruben, K.; Hummel, F.C.; Müller-Vahl, K.; Schnitzler, A.; Gerloff, C.; Münchau, A.; et al. Altered intrahemispheric structural connectivity in Gilles de la Tourette syndrome. NeuroImage Clin. 2013, 4, 174–181. [Google Scholar] [CrossRef] [Green Version]
- Makki, M.I.; Govindan, R.M.; Wilson, B.J.; Behen, M.E.; Chugani, H.T. Altered Fronto-Striato-Thalamic Connectivity in Children with Tourette Syndrome Assessed with Diffusion Tensor MRI and Probabilistic Fiber Tracking. J. Child Neurol. 2009, 24, 669–678. [Google Scholar] [CrossRef]
- Götz, T.; Siebner, H.R.; Jonas, M.; Bäumer, T.; Biermann-Ruben, K.; Hummel, F.; Gerloff, C.; Müller-Vahl, K.; Schnitzler, A.; Orth, M.; et al. Structural changes in the somatosensory system correlate with tic severity in Gilles de la Tourette syndrome. Brain 2009, 132, 765–777. [Google Scholar] [CrossRef] [Green Version]
- Worbe, Y.; Marrakchi-Kacem, L.; LeComte, S.; Valabregue, R.; Poupon, F.; Guevara, P.; Tucholka, A.; Mangin, J.-F.; Vidailhet, M.; Lehericy, S.; et al. Altered structural connectivity of cortico-striato-pallido-thalamic networks in Gilles de la Tourette syndrome. Brain 2014, 138, 472–482. [Google Scholar] [CrossRef] [Green Version]
- Leckman, J.F.; Zhang, H.; Vitale, A.; Lahnin, F.; Lynch, K.; Bondi, C.; Kim, Y.-S.; Peterson, B.S. Course of tic severity in Tourette syndrome: The first two decades. Pediatrics 1998, 102, 14–19. [Google Scholar] [CrossRef]
- Jankovic, J.; Kurlan, R. Tourette syndrome: Evolving concepts. Mov. Disord. 2011, 26, 1149–1156. [Google Scholar] [CrossRef]
- Andrade, P.; Visser-Vandewalle, V. DBS in Tourette syndrome: Where are we standing now? J. Neural Transm. 2016, 123, 791–796. [Google Scholar] [CrossRef]
- A Johnson, K.; Fletcher, P.T.; Servello, D.; Bona, A.; Porta, M.; Ostrem, J.L.; Bardinet, E.; Welter, M.-L.; Lozano, A.M.; Baldermann, J.C.; et al. Image-based analysis and long-term clinical outcomes of deep brain stimulation for Tourette syndrome: A multisite study. J. Neurol. Neurosurg. Psychiatry 2019, 90, 1078–1090. [Google Scholar] [CrossRef] [PubMed]
- Baldermann, J.; Schüller, T.; Huys, D.; Becker, I.; Timmermann, L.; Jessen, F.; Visser-Vandewalle, V.; Kuhn, J.; Information, P.E.K.F.C. Deep Brain Stimulation for Tourette-Syndrome: A Systematic Review and Meta-Analysis. Brain Stimul. 2016, 9, 296–304. [Google Scholar] [CrossRef]
- Schrock, L.E.; Mink, J.W.; Woods, D.W.; Porta, M.; Servello, D.; Visser-Vandewalle, V.; Silburn, P.A.; Foltynie, T.; Walker, H.C.; Shahed-Jimenez, J.; et al. Tourette Syndrome Association International Deep Brain Stimulation (DBS) Database and Registry Study Group. Tourette syndrome deep brain stimulation: A review and updated recommendations. Mov. Disord. 2015, 30, 448–471. [Google Scholar] [CrossRef] [PubMed]
- Henderson, J.M. “Connectomic surgery”: Diffusion tensor imaging (DTI) tractography as a targeting modality for surgical modulation of neural networks. Front. Integr. Neurosci. 2012, 6, 15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hartmann, C.J.; Lujan, J.L.; Chaturvedi, A.; Goodman, W.K.; Okun, M.S.; McIntyre, C.C.; Haq, I.U. Tractography Activation Patterns in Dorsolateral Prefrontal Cortex Suggest Better Clinical Responses in OCD DBS. Front. Mol. Neurosci. 2016, 9, 1104. [Google Scholar] [CrossRef] [Green Version]
- Vanegas-Arroyave, N.; Lauro, P.M.; Huang, L.; Hallett, M.; Horovitz, S.G.; Zaghloul, K.; Lungu, C. Tractography patterns of subthalamic nucleus deep brain stimulation. Brain 2016, 139, 1200–1210. [Google Scholar] [CrossRef] [Green Version]
- Morishita, T.; Higuchi, M.-A.; Kobayashi, H.; Abe, H.; Higashi, T.; Inoue, T. A retrospective evaluation of thalamic targeting for tremor deep brain stimulation using high-resolution anatomical imaging with supplementary fiber tractography. J. Neurol. Sci. 2019, 398, 148–156. [Google Scholar] [CrossRef]
- Mayka, M.A.; Corcos, D.M.; Leurgans, S.E.; Vaillancourt, D.E. Three-dimensional locations and boundaries of motor and premotor cortices as defined by functional brain imaging: A meta-analysis. NeuroImage 2006, 31, 1453–1474. [Google Scholar] [CrossRef] [Green Version]
- Tustison, N.J.; Avants, B.B.; Gee, J.C. Learning image-based spatial transformations via convolutional neural networks: A review. Magn. Reson. Imaging 2019, 64, 142–153. [Google Scholar] [CrossRef]
- Yushkevich, P.A.; Piven, J.; Hazlett, H.C.; Smith, R.G.; Ho, S.; Gee, J.C.; Gerig, G. User-guided 3D active contour segmentation of anatomical structures: Significantly improved efficiency and reliability. NeuroImage 2006, 31, 1116–1128. [Google Scholar] [CrossRef] [Green Version]
- Horn, A.; A Kühn, A. Lead-DBS: A toolbox for deep brain stimulation electrode localizations and visualizations. NeuroImage 2015, 107, 127–135. [Google Scholar] [CrossRef]
- Dembek, T.A.; Barbe, M.T.; Åström, M.; Hoevels, M.; Visser-Vandewalle, V.; Fink, G.R.; Timmermann, L. Probabilistic mapping of deep brain stimulation effects in essential tremor. NeuroImage Clin. 2016, 13, 164–173. [Google Scholar] [CrossRef] [PubMed]
- Andersson, J.L.; Skare, S.; Ashburner, J. How to correct susceptibility distortions in spin-echo echo-planar images: Application to diffusion tensor imaging. NeuroImage 2003, 20, 870–888. [Google Scholar] [CrossRef]
- Andersson, J.L.; Sotiropoulos, S.N. An integrated approach to correction for off-resonance effects and subject movement in diffusion MR imaging. NeuroImage 2016, 125, 1063–1078. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Andersson, J.L.R.; Sotiropoulos, S.N. Non-parametric representation and prediction of single- and multi-shell diffusion-weighted MRI data using Gaussian processes. NeuroImage 2015, 122, 166–176. [Google Scholar] [CrossRef] [Green Version]
- Smith, S.M.; Jenkinson, M.; Woolrich, M.W.; Beckmann, C.F.; Behrens, T.E.; Johansen-Berg, H.; Bannister, P.R.; De Luca, M.; Drobnjak, I.; Flitney, D.E.; et al. Advances in functional and structural MR image analysis and implementation as FSL. NeuroImage 2004, 23, S208–S219. [Google Scholar] [CrossRef] [Green Version]
- Behrens, T.E.; Woolrich, M.; Jenkinson, M.; Johansen-Berg, H.; Nunes, R.G.; Clare, S.; Matthews, P.M.; Brady, J.; Smith, S.M. Characterization and propagation of uncertainty in diffusion-weighted MR imaging. Magn. Reson. Med. 2003, 50, 1077–1088. [Google Scholar] [CrossRef]
- Behrens, T.E.; Johansen-Berg, H.; Jbabdi, S.; Rushworth, M.F.; Woolrich, M. Probabilistic diffusion tractography with multiple fibre orientations: What can we gain? NeuroImage 2007, 34, 144–155. [Google Scholar] [CrossRef]
- Behrens, T.E.J.; Johansen-Berg, H.; Woolrich, M.W.; Smith, S.; Wheeler-Kingshott, C.A.G.; A Boulby, P.; Barker, G.J.; Sillery, E.L.; Sheehan, K.; Ciccarelli, O.; et al. Non-invasive mapping of connections between human thalamus and cortex using diffusion imaging. Nat. Neurosci. 2003, 6, 750–757. [Google Scholar] [CrossRef]
- Polyanska, L.; Critchley, H.D.; Rae, C.L. Centrality of prefrontal and motor preparation cortices to Tourette Syndrome revealed by meta-analysis of task-based neuroimaging studies. NeuroImage Clin. 2017, 16, 257–267. [Google Scholar] [CrossRef]
- Zapparoli, L.; Porta, M.; Paulesu, E. The anarchic brain in action. Curr. Opin. Neurol. 2015, 28, 604–611. [Google Scholar] [CrossRef]
- Nachev, P.; Wydell, H.; O’Neill, K.; Husain, M.; Kennard, C. The role of the pre-supplementary motor area in the control of action. NeuroImage 2007, 36, T155–T163. [Google Scholar] [CrossRef] [Green Version]
- Cavanna, A.E.; Black, K.J.; Hallett, M.; Voon, V. Neurobiology of the Premonitory Urge in Tourette’s Syndrome: Pathophysiology and Treatment Implications. J. Neuropsychiatry Clin. Neurosci. 2017, 29, 95–104. [Google Scholar] [CrossRef] [Green Version]
- Hassler, R.; Dieckmann, G. Stereotaxic treatment of tics and inarticulate cries or coprolalia considered as motor obsessional phenomena in Gilles de la Tourette’s disease. Rev. Neurol. 1970, 123, 89–100. [Google Scholar] [PubMed]
- Vandewalle, V.; Van Der Linden, C.; Groenewegen, H.J.; Caemaert, J. Stereotactic treatment of Gilles de la Tourette syndrome by high frequency stimulation of thalamus. Lancet 1999, 353, 724. [Google Scholar] [CrossRef]
- Ackermans, L.; Duits, A.; Van Der Linden, C.; Tijssen, M.; Schruers, K.; Temel, Y.; Kleijer, M.; Nederveen, P.; Bruggeman, R.; Tromp, S.; et al. Double-blind clinical trial of thalamic stimulation in patients with Tourette syndrome. Brain 2011, 134, 832–844. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brito, M.; Teixeira, M.J.; Mendes, M.M.; França, C.; Iglesio, R.; Barbosa, E.R.; Cury, R.G. Exploring the clinical outcomes after deep brain stimulation in Tourette syndrome. J. Neurol. Sci. 2019, 402, 48–51. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Pat. Number | Gender | Age | YGTSS Preop | YGTSS Postop | Improvement (%) | Active Contacts |
| Pat. 1 | Male | 22 | 78 | 22 | 71.8 | 0 and 1 |
| Pat. 2 | Male | 32 | 89 | 44 | 50.6 | 2 and 3 |
| Pat. 3 | Female | 26 | 95 | 47 | 50.5 | 1 and 2 |
| Pat. 4 | Male | 37 | 81 | 43 | 46.9 | 2 and 3 |
| Pat. 5 | Male | 20 | 87 | 49 | 43.7 | 2 and 3 |
| Pat. 6 | Male | 46 | 100 | 70 | 30.0 | 2 and 3 |
| Pat. 7 | Male | 27 | 100 | 89 | 11.0 | 1 and 2 |
| Pat. Number | Freq. Hz | Pulse Width µs | Ampl. V | Comorbidity | Medication | |
| Pat. 1 | 130 | 90 | 3.2 | None | Aripiprazole 10 mg, Citalopram 30 mg | |
| Pat. 2 | 130 | 90 | 3.3 | None | Tiapride 800 mg, Quetiapine 150 mg | |
| Pat. 3 | 100 | 90 | 3.0 | Depression | Escitalopram 10 mg, THC 5 mg, Risperidone 4 mg, Quetiapine 25 mg | |
| Pat. 4 | 110 | 90 | 4.1 | Opiate dependence (in substitution) | Methadone 28 mL | |
| Pat. 5 | 100 | 150 | 5.2 | None | None | |
| Pat. 6 | 80 | 150 | 5.0 | Depression, OCD | Aripiprazole 5 mg, Pipamperone 40 mg, Quetiapine 200 mg | |
| Pat. 7 | 80 | 300 | 4.7 | Impulse control disorder, OCD, anxiety disorder | Clonazepam 1 mg, Pregabaline 300 mg, Risperidone 2 mg, THC 15 mg, Venlafaxine 225 mg | |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Andrade, P.; Heiden, P.; Hoevels, M.; Schlamann, M.; Baldermann, J.C.; Huys, D.; Visser-Vandewalle, V. Modulation of Fibers to Motor Cortex during Thalamic DBS in Tourette Patients Correlates with Tic Reduction. Brain Sci. 2020, 10, 302. https://doi.org/10.3390/brainsci10050302
Andrade P, Heiden P, Hoevels M, Schlamann M, Baldermann JC, Huys D, Visser-Vandewalle V. Modulation of Fibers to Motor Cortex during Thalamic DBS in Tourette Patients Correlates with Tic Reduction. Brain Sciences. 2020; 10(5):302. https://doi.org/10.3390/brainsci10050302
Chicago/Turabian StyleAndrade, Pablo, Petra Heiden, Moritz Hoevels, Marc Schlamann, Juan C. Baldermann, Daniel Huys, and Veerle Visser-Vandewalle. 2020. "Modulation of Fibers to Motor Cortex during Thalamic DBS in Tourette Patients Correlates with Tic Reduction" Brain Sciences 10, no. 5: 302. https://doi.org/10.3390/brainsci10050302
APA StyleAndrade, P., Heiden, P., Hoevels, M., Schlamann, M., Baldermann, J. C., Huys, D., & Visser-Vandewalle, V. (2020). Modulation of Fibers to Motor Cortex during Thalamic DBS in Tourette Patients Correlates with Tic Reduction. Brain Sciences, 10(5), 302. https://doi.org/10.3390/brainsci10050302