Effects of Transcranial Magnetic Stimulation Therapy on Evoked and Induced Gamma Oscillations in Children with Autism Spectrum Disorder

 ,
,  ,
, 

Abstract
:1. Introduction
2. Methods and Materials
2.1. Subjects
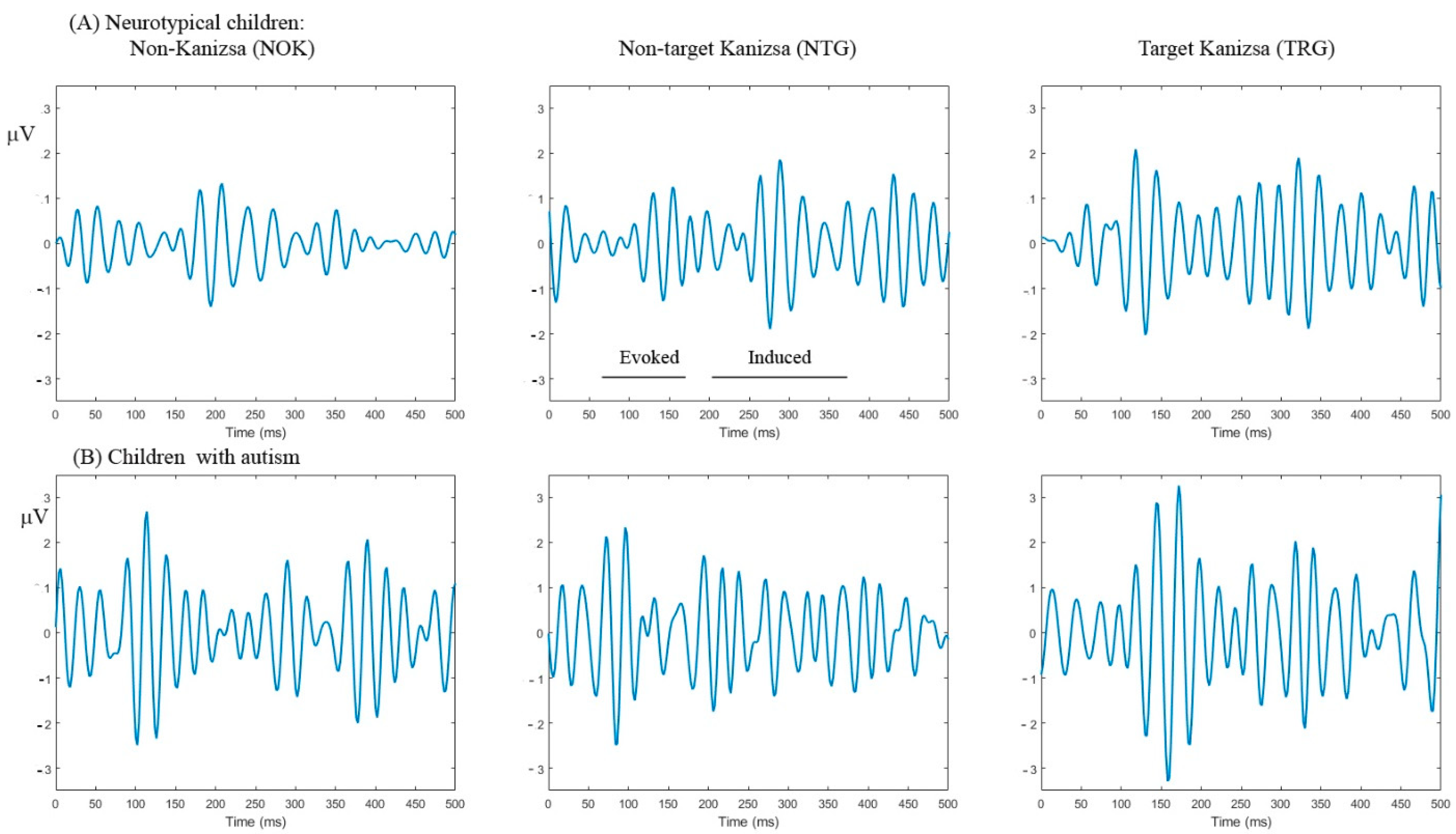
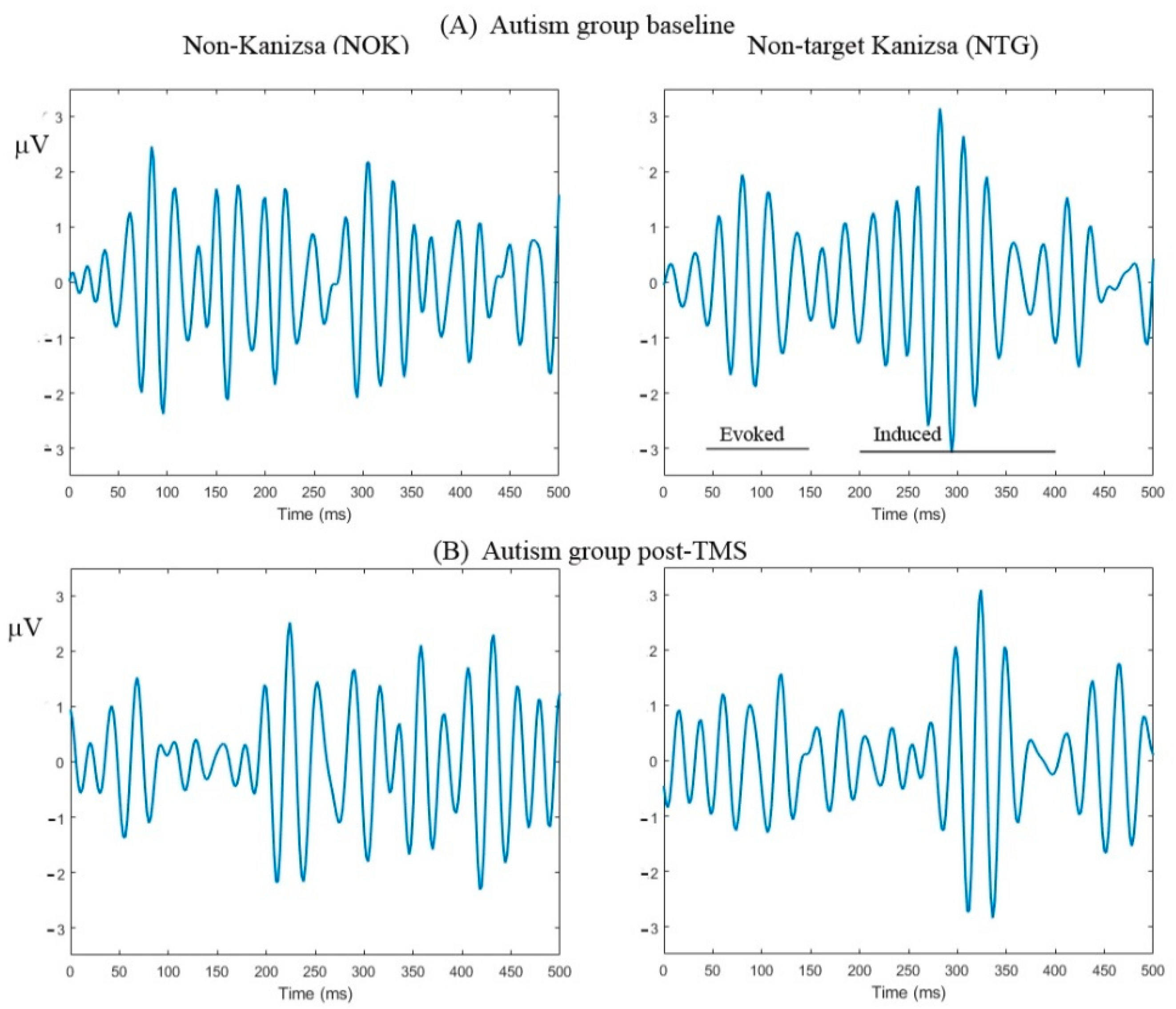
2.2. Experimental Task: Visual Oddball with Illusory Kanizsa Figures
2.3. Event-Related Gamma Oscillations Recording
2.4. EEG Analysis
Method Description
2.5. Transcranial Magnetic Stimulation
2.6. Behavioral and Social Functioning Evaluation
2.7. Statistical Analysis
3. Results
3.1. Behavioral Responses (Reaction Time and Accuracy)
3.2. Behavior Evaluations Post-TMS
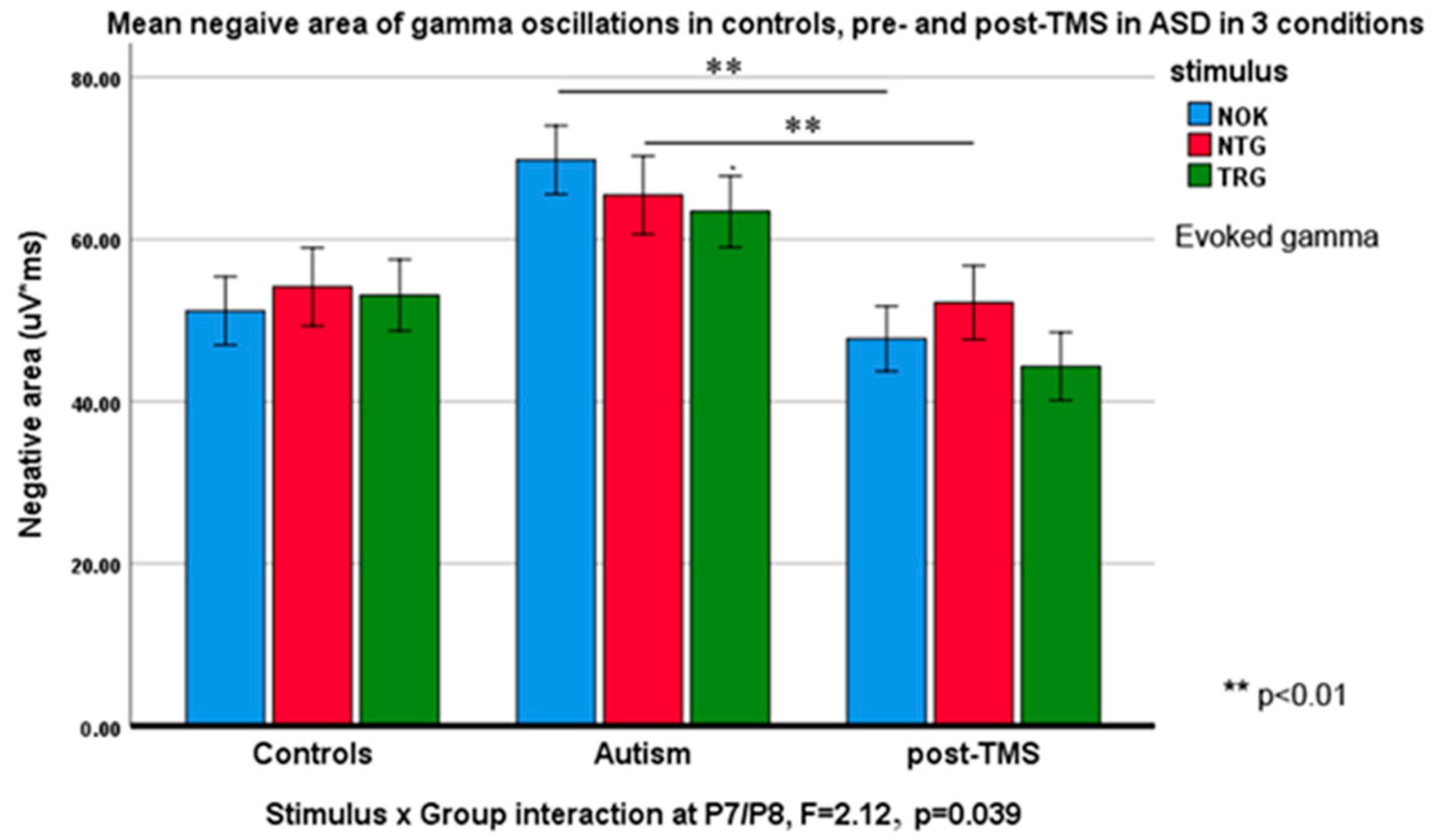
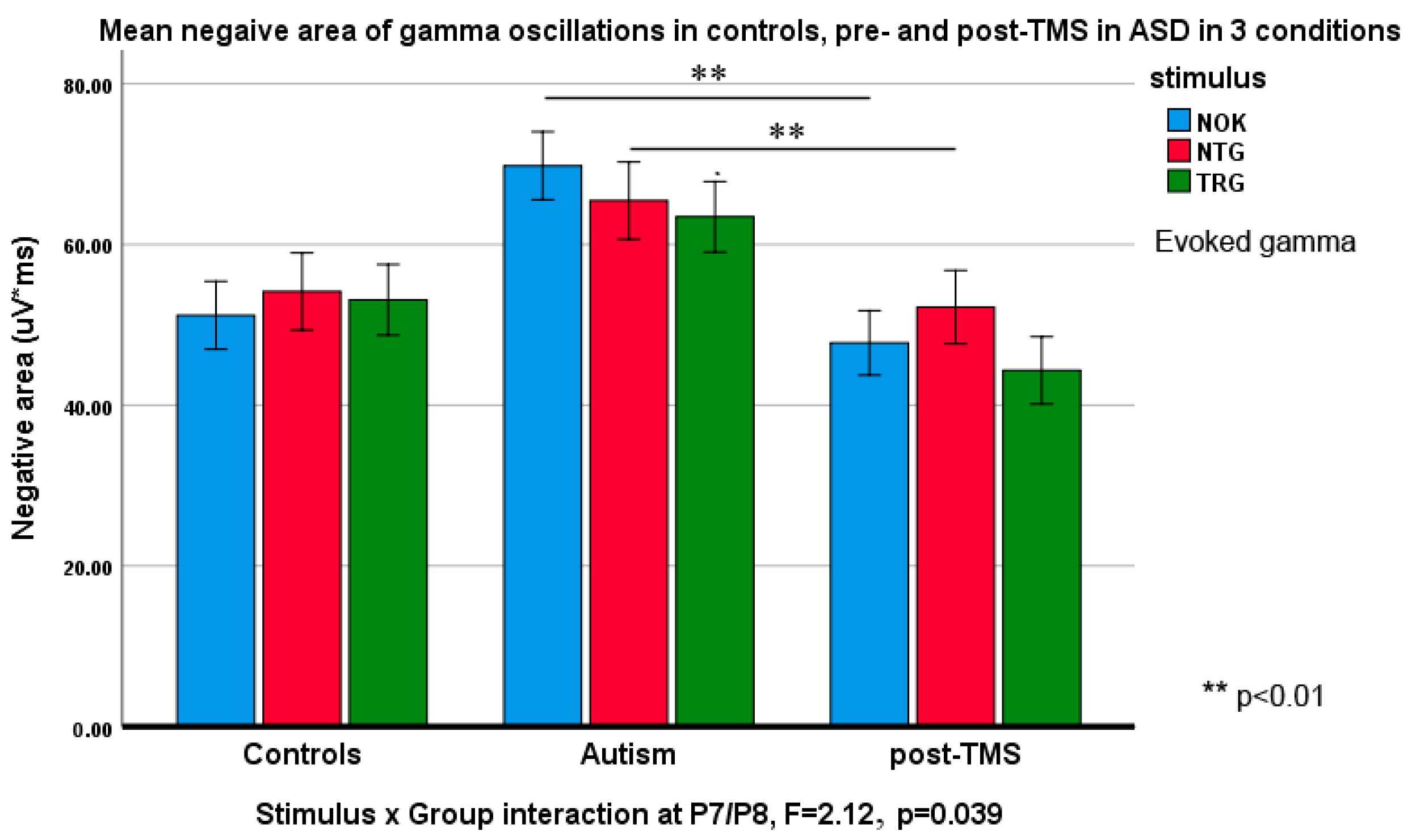
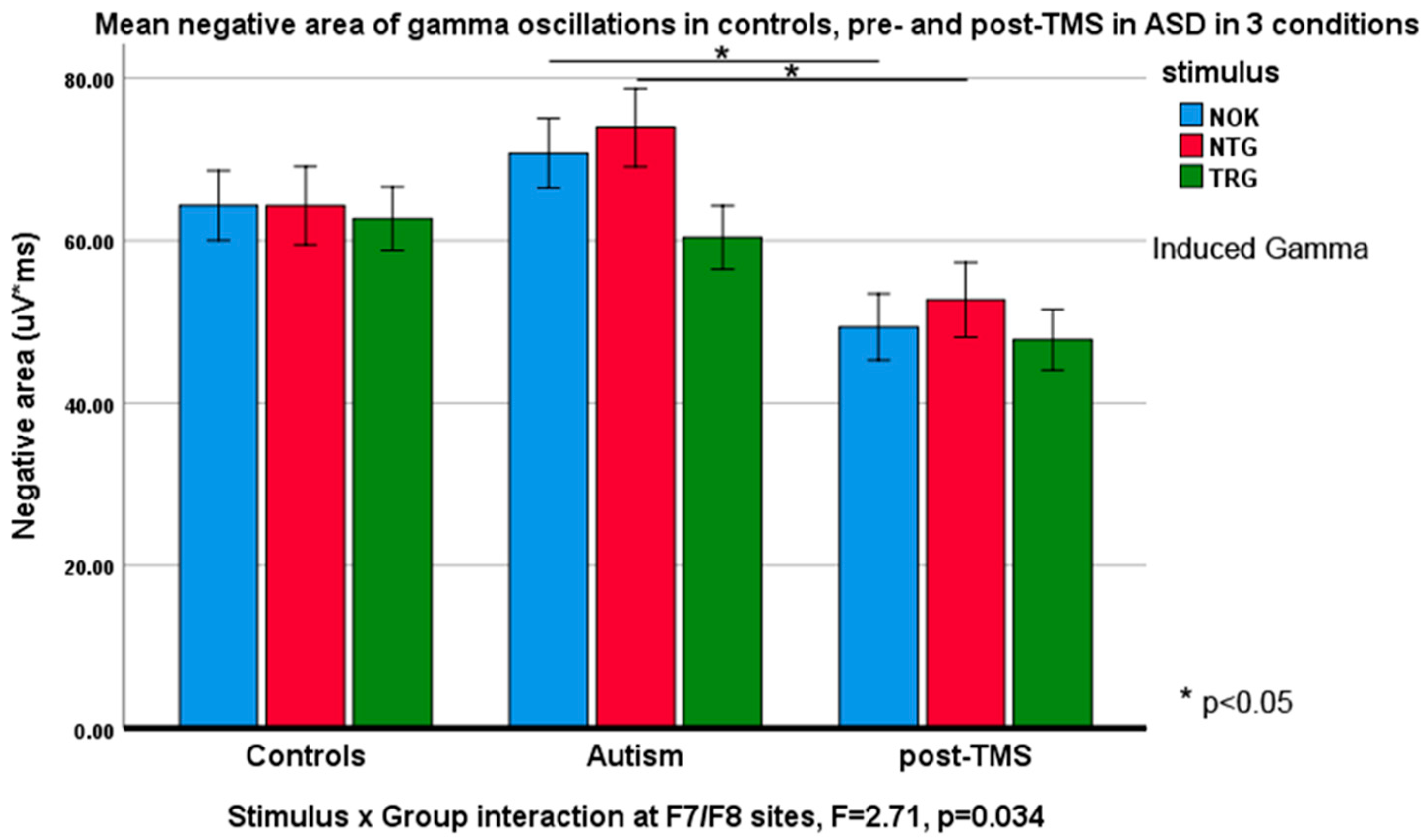
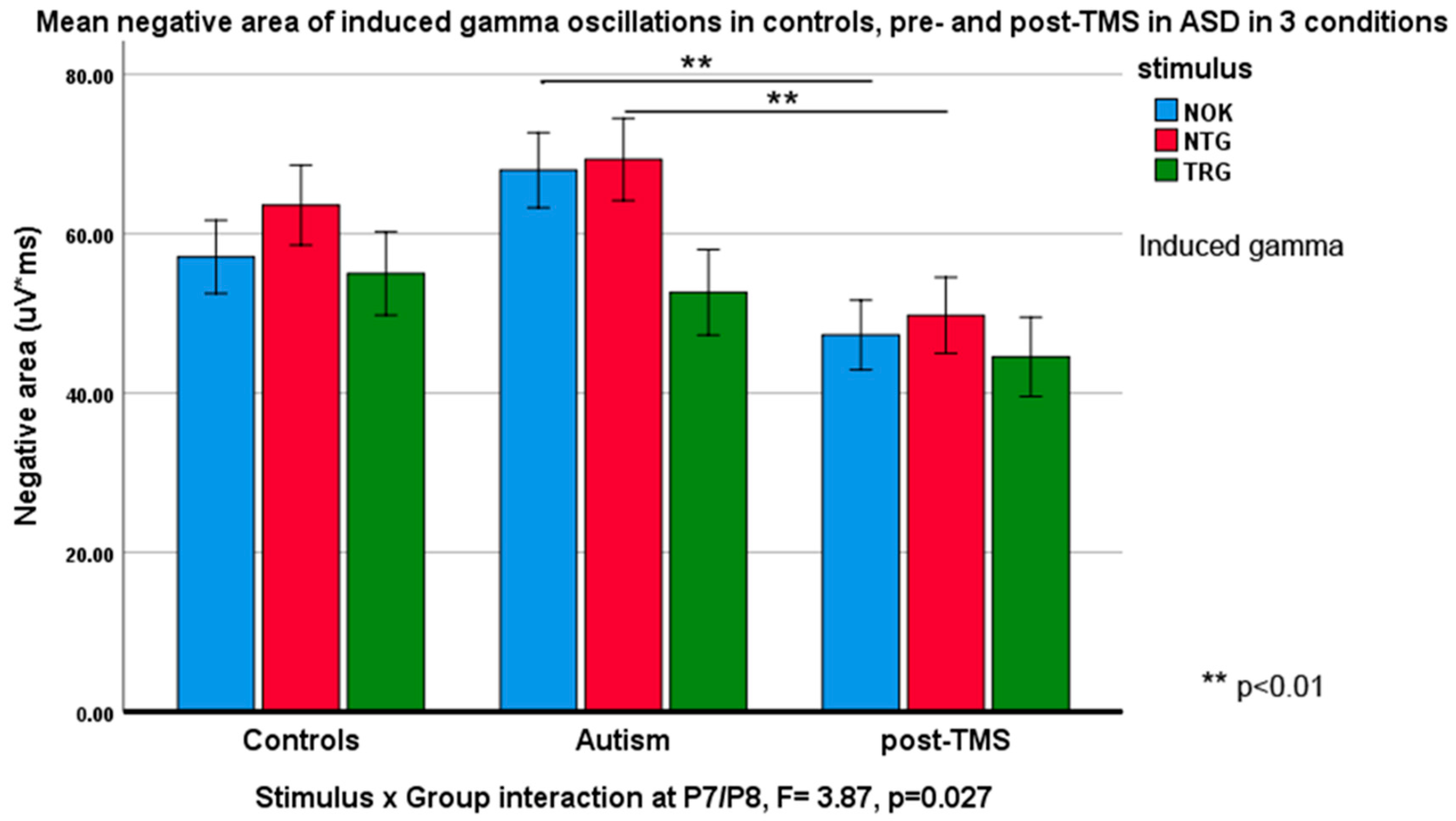
3.3. Evoked and Induced Gamma Oscillations
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Buzsáki, G.; Wang, X.J. Mechanisms of gamma oscillations. Annu. Rev. Neurosci. 2012, 35, 203–225. [Google Scholar] [CrossRef] [Green Version]
- Snijders, T.M.; Milivojevic, B.; Kemner, C. Atypical excitation-inhibition balance in autism captured by the gamma response to contextual modulation. Neuroimage Clin. 2013, 3, 65–72. [Google Scholar] [CrossRef] [Green Version]
- Steullet, P.; Cabungcal, J.-H.; Coyle, J.; Didriksen, M.; Gill, K.; Grace, A.A.; Hensch, T.K.; LaMantia, A.-S.; Lindemann, L.; Maynard, T.M.; et al. Oxidative stress-driven parvalbumin interneuron impairment as a common mechanism in models of schizophrenia. Mol. Psychiatry 2017, 22, 936–943. [Google Scholar] [CrossRef]
- Hashemi, E.; Ariza, J.; Rogers, H.; Noctor, S.C.; Martinez-Cerdeno, V. The number of parvalbumin-expressing interneurons is decreased in the prefrontal cortex in autism. Cereb. Cortex 2017, 27, 1931–1943. [Google Scholar] [PubMed] [Green Version]
- Lee, E.; Lee, J.; Kim, E. Excitation/inhibition imbalance in animal models of autism spectrum disorders. Biol. Psychiatry 2017, 81, 838–847. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saunders, J.A.; Tatard-Leitman, V.M.; Suh, J.; Billingslea, E.N.; Roberts, T.P.; Siegel, S.J. Knockout of NMDA receptors in parvalbumin interneurons recreates autism-like phenotypes. Autism Res. 2013, 6, 69–77. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wöhr, M.; Orduz, D.; Gregory, P.; Moreno, H.; Khan, U.; Vörckel, K.J.; Wolfer, D.P.; Welzl, H.; Gall, D.; Schiffmann, S.N.; et al. Lack of parvalbumin in mice leads to behavioral deficits relevant to all human autism core symptoms and related neural morphofunctional abnormalities. Transl. Psychiatry 2015, 5, e525. [Google Scholar] [CrossRef]
- Selimbeyoglu, A.; Kim, C.K.; Inoue, M.; Lee, S.Y.; Hong, A.S.O.; Kauvar, I.; Ramakrishnan, C.; Fenno, L.E.; Davidson, T.J.; Wright, M.; et al. Modulation of prefrontal cortex excitation/inhibition balance rescues social behavior in CNTNAP2-deficient mice. Sci. Transl. Med. 2017, 9, eaah6733. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mukherjee, A.; Carvalho, F.; Eliez, S.; Caroni, P. Long-lasting rescue of network and cognitive dysfunction in a genetic schizophrenia model. Cell 2019, 178, 1387–1402. [Google Scholar] [CrossRef]
- Sokhadze, E.M.; El-Baz, A.; Baruth, J.; Mathai, G.; Sears, L.; Casanova, M.F. Effects of low frequency repetitive transcranial magnetic stimulation (rTMS) on gamma frequency oscillations and event-related potentials during processing of illusory figures in autism. J. Autism Dev. Disord. 2009, 39, 619–634. [Google Scholar] [CrossRef]
- Rojas, D.C.; Wilson, L.B. Gamma-band abnormalities as markers of autism spectrum disorders. Biomark Med. 2014, 8, 353–368. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Casanova, M.F.; Baruth, J.; El-Baz, A.S.; Sokhadze, G.E.; Hensley, M.; Sokhadze, E.S. Evoked and induced gamma frequency oscillations in autism. In Imaging the Brain in Autism; Casanova, M.F., El-Baz, A.S., Suri, J.S., Eds.; Springer: New York, NY, USA, 2013; pp. 87–106. [Google Scholar]
- Rippon, G. Gamma abnormalities in autism spectrum disorders. In Autism Imaging and Devices; Casanova, M.F., El-Baz, A., Suri, J.S., Eds.; CRC Press, Taylor and Francis Group: Boca Raton, FL, USA, 2017; pp. 457–496. [Google Scholar]
- Başar, E.; Tülay, E.; Güntekin, B. Multiple gamma oscillations in the brain: A new strategy to differentiate functional correlates and P300 dynamics. Int. J. Psychophysiol. 2015, 95, 406–420. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Galambos, R.A. Comparison of certain gamma band 40 Hz brain rhythms in cat and man. In Induced Rhythms in the Brain; Basar, E., Bullock, T.H., Eds.; Birkhauser: Boston, MA, USA, 1992; pp. 201–216. [Google Scholar]
- Sheer, D.E. Focused arousal and 40 Hz EEG. In The Neuropsychology of Learning Disorders; Knight, R.M., Baker, D.J., Eds.; University Park Press: Baltimore, MD, USA, 1976. [Google Scholar]
- Kahana, M.J. The cognitive correlates of human brain oscillations. J. Neurosci. 2006, 26, 1669–1672. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kaiser, J.; Lutzenberger, W. Induced gamma-band activity and human brain function. Neuroscientist 2003, 9, 475–484. [Google Scholar] [CrossRef]
- Tallon-Baudry, C.; Bertrand, O. Oscillatory gamma activity in humans and its role in object representation. Trends Cogn. Sci. 1999, 3, 151–162. [Google Scholar] [CrossRef]
- Tallon-Baudry, C.; Bertrand, O.; Henaff, M.A.; Isnard, J.; Fischer, C. Attention modulates gamma-band oscillations differently in the human lateral occipital cortex and fusiform gyrus. Cereb. Cortex 2005, 15, 654–662. [Google Scholar] [CrossRef] [Green Version]
- Brock, J.; Brown, C.C.; Boucher, J.; Rippon, G. The temporal binding deficit hypothesis of autism. Dev. Psychopathol. 2002, 14, 209–224. [Google Scholar] [CrossRef] [Green Version]
- Brown, C.; Gruber, T.; Boucher, J.; Rippon, G.; Brock, J. Gamma abnormalities during perception of illusory figures in autism. Cortex 2005, 41, 364–376. [Google Scholar] [CrossRef]
- Rippon, G.; Brock, J.; Brown, C.; Boucher, J. Disordered connectivity in the autistic brain: Challenges for the “new psychophysiology”. Int. J. Psychophysiol. 2007, 63, 164–172. [Google Scholar] [CrossRef]
- Maxwell, C.R.; Villalobos, M.E.; Schultz, R.T.; Herpertz-Dahlmann, B.; Konrad, K.; Kohls, G. Atypical laterality of resting gamma oscillations in autism spectrum disorders. J. Autism Dev. Disord. 2015, 45, 292–297. [Google Scholar] [CrossRef]
- Kanizsa, G. Subjective contours. Sci. Am. 1976, 234, 48–52. [Google Scholar] [CrossRef] [PubMed]
- Baruth, J.M.; Casanova, M.F.; El-Baz, A.; Horrell, T.; Mathai, G.; Sears, L.; Sokhadze, E. Low-frequency repetitive transcranial magnetic stimulation (rTMS) modulates evoked-gamma frequency oscillations in autism spectrum disorder (ASD). J. Neurother 2010, 14, 179–194. [Google Scholar] [CrossRef]
- Baruth, J.M.; Williams, E.L.; Sokhadze, E.M.; El-Baz, A.S.; Casanova, M.F. Beneficial effects of repetitive transcranial magnetic stimulation (rTMS) on behavioral outcome measures in autism spectrum disorder. Autism Sci. Dig. 2011, 1, 52–57. [Google Scholar]
- Casanova, M.F.; Baruth, J.M.; El-Baz, A.; Tasman, A.; Sears, L.; Sokhadze, E. Repetitive transcranial magnetic stimulation (rTMS) modulates event-related potential (ERP) indices of attention in autism. Transl. Neurosci. 2012, 3, 170–180. [Google Scholar] [CrossRef] [PubMed]
- Sokhadze, E.M.; El-Baz, A.S.; Sears, L.L.; Opris, I.; Casanova, M.F. rTMS neuromodulation improves electrocortical functional measures of information processing and behavioral responses in autism. Front. Syst. Neurosci. 2014, 8, 134. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Casanova, M.F.; Shaban, M.; Ghazal, M.; Sokhadze, E.; Casanova, E.L.; El-Baz, A.S. Ringing decay of gamma oscillations and transcranial magnetic stimulation therapy in autism spectrum disorder. Appl. Psychophysiol. Biofeedback 2020. under review. [Google Scholar]
- Silver, M.L.; Tiedemann, D. Dynamic Geotechnical Testing; ASTM International: West Conshohocken, PA, USA, 1978. [Google Scholar]
- Attallah, B.V.; Scanziani, M. Instantaneous modulation of gamma oscillation frequency by balancing excitation with inhibition. Neuron 2009, 62, 566–577. [Google Scholar] [CrossRef] [Green Version]
- American Psychiatric Association. Diagnostic and Statistical Manual of Mental Disorders (DSM-IV), 4th ed.; American Psychiatric Publishing, Inc.: Washington, DC, USA, 2000. [Google Scholar]
- American Psychiatric Association. Diagnostic and Statistical Manual of Mental Disorders (DSM-5), 5th ed.; American Psychiatric Publishing, Inc.: Washington, DC, USA, 2013. [Google Scholar]
- LeCouteur, A.; Lord, C.; Rutter, M. The Autism Diagnostic Interview—Revised (ADI-R); Western Psychological Services: Los Angeles, CA, USA, 2003. [Google Scholar]
- Wechsler, D. Wechsler Intelligence Scale for Children (WISC-IV), 4th ed.; Harcourt Assessment, Inc.: San Antonio, TX, USA, 2003. [Google Scholar]
- Wechsler, D. Wechsler Abbreviated Scale of Intelligence (WASI); Harcourt Assessment, Inc.: San Antonio, TX, USA, 1999. [Google Scholar]
- First, M.B.; Spitzer, R.L.; Gibbon, M.; Williams, J.B.W. Structured Clinical Interview for DSM-IV-TR Axis I Disorders, Research Version (SCID-I/NP); Biometrics Research, New York State Psychiatric Institute: New York, NY, USA, 2002. [Google Scholar]
- Luu, P.; Tucker, D.M.; Englander, R.; Lockfeld, A.; Lutsep, H.; Oken, B. Localizing acute stroke-related EEG changes: Assessing the effects of spatial undersampling. J. Clin. Neurophysiol. 2001, 18, 302–317. [Google Scholar] [CrossRef]
- Sokhadze, E.M.; Baruth, J.M.; Sears, L.; Sokhadze, G.E.; El-Baz, A.; Casanova, M.F. Prefrontal neuromodulation using rTMS improves error monitoring and correction function in autism. Appl. Psychophysiol. Biofeedback 2012, 37, 91–102. [Google Scholar] [CrossRef]
- Sokhadze, E.M.; Lamina, E.V.; Casanova, E.L.; Kelly, D.P.; Opris, I.; Tasman, A.; Casanova, M.F. Exploratory study of rTMS neuromodulation effects on electrocortical functional measures of performance in an oddball test and behavioral symptoms in autism. Front. Syst. Neurosci. 2018, 12, 20. [Google Scholar] [CrossRef] [Green Version]
- Mir-Moghtadaei, A.; Caballero, R.; Fried, P.; Fox, M.D.; Lee, K.; Giacobbe, P.; Daskalakis, Z.J.; Blumberger, D.M.; Downar, J. Concordance between BeamF3 and MRI-neuronavigated target sites for repetitive transcranial magnetic stimulation of the left dorsolateral prefrontal cortex. Brain Stimul. 2015, 8, 965–973. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pommier, B.; Vassal, F.; Boutet, C.; Jeannin, S.; Peyron, R.; Faillenot, I. Easy methods to make the neuronavigated targeting of DLPFC accurate and routinely accessible for rTMS. Neurophysiol. Clin. 2017, 7, 35–46. [Google Scholar] [CrossRef] [PubMed]
- Pascual-Leone, A.; Walsh, V.; Rothwell, J. Transcranial magnetic stimulation in cognitive neuroscience--virtual lesion, chronometry, and functional connectivity. Curr. Opin. Neurobiol. 2000, 10, 232–237. [Google Scholar] [CrossRef]
- Oberman, L.M.; Enticott, P.G.; Casanova, M.F.; Rotenberg, A.; Pascual-Leone, A.; McCracken, J.T.; TMS in ASD Consensus Group. Transcranial magnetic stimulation in autism spectrum disorder: Challenges, promise, and roadmap for future research. Autism Res. 2016, 9, 184–203. [Google Scholar] [CrossRef]
- Wassermann, E.M.; Lisanby, S.H. Therapeutic application of repetitive transcranial magnetic stimulation: A review. Clin. Neurophysiol. 2001, 112, 1367–1377. [Google Scholar] [CrossRef]
- Wassermann, E.M.; Zimmermann, T. Transcranial magnetic stimulation: Therapeutic promises and scientific gaps. Pharm. Ther. 2012, 133, 98–107. [Google Scholar] [CrossRef] [Green Version]
- Maeda, F.; Keenan, J.P.; Tormos, J.M.; Topka, H.; Pascual-Leone, A. Modulation of corticospinal excitability by repetitive transcranial magnetic stimulation. Clin. Neurophysiol. 2000, 111, 800–805. [Google Scholar] [CrossRef]
- Aman, M.G.; Singh, N.N. Aberrant Behavior Checklist-Community. Supplementary Manual; Slosson Educational Publications: East Aurora, NY, USA, 1994. [Google Scholar]
- Bodfish, J.W.; Symons, F.J.; Lewis, J. Repetitive Behavior Scale (Western Carolina Center Research Reports); Western Carolina Center: Morganton, NC, USA, 1999. [Google Scholar]
- Jovanov, E.; Rakovic, D.; Radivojevic, V.; Kusic, D. Band Power Envelope Analysis: A New Method in Quantitative EEG. In Proceedings of the 17th International Conference of the Engineering in Medicine and Biology Society, Montréal, QC, Canada, 20–23 September 1995. [Google Scholar]
- Clochon, P.; Fontbonne, J.; Lebrun, N.; Etevenon, P. A new method for quantifying EEG event-related desynchronization: Amplitude envelope analysis. Electroencephalogr. Clin. Neurophysiol. 1996, 98, 126–129. [Google Scholar] [CrossRef]
- Draganova, R.; Popivanov, D. Assessment of EEG frequency dynamics using complex demodulation. Physiol. Res. 1999, 48, 157–165. [Google Scholar]
- Baron-Cohen, S.; Wheelwright, S.; Skinner, R.; Martin, J.; Clubley, E. The autism-spectrum quotient (AQ): Evidence from Asperger syndrome/high-functioning autism, males and females, scientists and mathematicians. J. Autism Dev. Disord. 2001, 31, 5–17. [Google Scholar] [CrossRef]
- Cooper, N.R.; Simpson, A.; Till, A.; Simmons, K.; Puzzo, I. Beta event-related desynchronization as an index of individual differences in processing human facial expression: Further investigations of autistic traits in typically developing adults. Front. Hum. Neurosci. 2013, 7, 159. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Trambaiolli, L.R.; Falk, T.H.; Fraga, F.J.; Anghinah, R.; Lorena, A.C. EEG spectro-temporal modulation energy: A new feature for automated diagnosis of Alzheimer’s disease. Conf. Proc. IEEE Eng. Med. Biol. Soc. 2011, 2011, 3828–3831. [Google Scholar] [PubMed] [Green Version]
- Griskova, I.; Ruksenas, O.; Dapsys, K.; Herpertz, S.; Hoppner, J. The effects of 10 Hz repetitive transcranial magnetic stimulation on resting EEG power spectrum in healthy subjects. Neurosci. Lett. 2007, 419, 162–167. [Google Scholar] [CrossRef] [PubMed]
- Gabard-Durnam, L.J.; Wilkinson, C.; Kapur, K.; Tager-Flusberg, H.; Levin, A.R.; Nelson, C.A. Longitudinal EEG power in the first postnatal year differentiates autism outcomes. Nat. Commun. 2019, 10, 4188. [Google Scholar] [CrossRef]
- Gutfreund, Y.; Yarom, Y.; Segev, I. Subthreshold oscillations and resonant frequency in guinea-pig cortical neurons: Physiology and modelling. J. Physiol. 1995, 483, 621–640. [Google Scholar] [CrossRef] [PubMed]
- Baars, B.J.; Gage, N.M. Cognition, Brain, and Consciousness, 2nd ed.; Academic Press: Cambridge, MA, USA, 2010. [Google Scholar]
- Arnsten, A.F.T. The neurobiology of thought: The groundbreaking discoveries of Patricia Goldman-Rakic 1937–2003. Cereb. Cortex 2013, 23, 2269–2281. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Curtis, C.E.; D’Esposito, M. Persistent activity in the prefrontal cortex during working memory. Trends Cogn. Sci. 2003, 7, 415–423. [Google Scholar] [CrossRef] [Green Version]
- Panerai, S.; Tasca, D.; Ferri, R.; D’Arrigo, V.G.; Elia, M. Executive functions and adaptive behavior in autism spectrum disorder with and without intellectual disability. Psychiatry J. 2014, 2014, 941809. [Google Scholar] [CrossRef] [Green Version]
- Hill, E.L. Executive dysfunction in autism. Trends Cogn. Sci. 2004, 8, 26–32. [Google Scholar] [CrossRef] [Green Version]
- Moser, D.J.; Jorge, R.E.; Manes, F.; Paradiso, S.; Benjamin, M.L.; Robinson, R.G. Improved executive functioning following repetitive transcranial magnetic stimulation. Neurology 2002, 58, 1288–1290. [Google Scholar] [CrossRef]
- Palaus, M.; Marron, E.M.; Viejo-Sobera, R.; Redolar-Ripoll, D. Cognitive Enhancement by Means of TMS: Memory and Executive Functions. In Proceedings of the 5th Conference of the European Societies of Neuropsychology, Tampere, Finland, 9–11 September 2015. [Google Scholar]
- Hauer, L.; Sellner, J.; Brigo, F.; Trinka, E.; Sebastianelli, L.; Saltuari, L.; Versace, V.; Höller, Y.; Nardone, R. Effects of repetitive transcranial magnetic stimulation over prefrontal cortex on attention in psychiatric disorders: A systematic review. J. Clin. Med. 2019, 8, 416. [Google Scholar] [CrossRef] [Green Version]
- Sokhadze, E.; Baruth, J.; El-Baz, A.; Horrell, T.; Sokhadze, G.; Carroll, T.; Tasman, A.; Sears, L.; Casanova, M.F. Impaired error monitoring and correction function in autism. J. Neurother 2010, 14, 79–95. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- van Es, M.W.J.; Schoffelen, J.M. Stimulus-induced gamma power predicts the amplitude of the subsequent visual evoked response. Neuroimage 2019, 186, 703–712. [Google Scholar] [CrossRef]
- Brunel, N.; Wang, X.J. What determines the frequency of fast network oscillations with irregular neural discharges? I. Synaptic dynamics and excitation-inhibition balance. J. Neurophysiol. 2003, 90, 415–430. [Google Scholar] [CrossRef] [PubMed]
- Edden, R.A.E.; Muthukumaraswamy, S.D.; Freeman, T.C.A.; Singh, K.D. Orientation discrimination performance is predicted by GABA concentration and gamma oscillation frequency in human primary visual cortex. J. Neurosci. 2009, 29, 15721–15726. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Muthukumaraswamy, S.D.; Edden, R.A.E.; Jones, D.K.; Swettenham, J.B.; Singh, K.D. Resting GABA concentration predicts peak gamma frequency and fMRI amplitude in response to visual stimulation in humans. Proc. Natl. Acad. Sci. USA 2009, 106, 8356–8361. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saxena, N.; Muthukumaraswamy, S.D.; Diukova, A.; Singh, K.; Hall, J.; Wise, R. Enhanced stimulus-induced gamma activity in humans during propofol-induced sedation. PLoS ONE 2013, 8, e57685. [Google Scholar] [CrossRef]
- Hannah, R.; Rocchi, L.; Tremblay, S.; Wilson, E.; Rothwell, J.C. Pulse width biases the balance of excitation and inhibition recruited by transcranial magnetic stimulation. Brain Stimul. 2020, 13, 536–538. [Google Scholar] [CrossRef] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Stimulus/EEG Site | CNT | ASD | F(1,36) | p |
|---|---|---|---|---|
| Positive/ascending area, evoked | ||||
| Target at F7 | 50.08 ± 22.61 | 69.22 ± 19.62 | 7.77 | 0.008 ** |
| Non-target at F7 | 44.47 ± 13.13 | 59.84 ± 20.54 | 7.55 | 0.009 ** |
| Non-target at F8 | 48.10 ± 15.77 | 60.57 ± 16.37 | 5.73 | 0.022 * |
| Non-target at P3 | 53.27 ± 19.20 | 73.72 ± 23.48 | 8.64 | 0.006 ** |
| Non-Kanizsa at F7 | 51.56 ± 21.35 | 69.82 ± 27.74 | 5.17 | 0.029 * |
| Non-Kanizsa at P7 | 61.09 ± 19.23 | 75.92 ± 21.13 | 5.12 | 0.030* |
| Negative/descending area, evoked | ||||
| Target at F7 | 55.23 ± 4.43 | 49.51 ± 4.42 | 15.86 | <0.001 *** |
| Non-target at F7 | 53.86 ± 4.55 | 49.21 ± 4.85 | 9.29 | 0.004 ** |
| Non-Kanizsa at F7 | 61.63 ± 22.54 | 47.01 ± 3.12 | 7.84 | 0.008 ** |
| Non-Kanizsa at P7 | 53.06 ± 21.58 | 68.35 ± 24.21 | 4.22 | 0.047 * |
| Negative/descending area, induced | ||||
| Target at P7 | 53.71 ± 22.95 | 70.30 ± 22.90 | 4.97 | 0.032 * |
| Non-target at P7 | 52.96 ± 27.07 | 73.67 ± 26.53 | 5.67 | 0.023 * |
| Stimulus/EEG Site | ASD Pre-TMS | ASD Post-TMS | F(1,36) | p |
|---|---|---|---|---|
| Positive/ascending area, evoked | ||||
| Target at F7 | 67.25 ± 20.79 | 51.67 ± 18.62 | 6.25 | 0.017 * |
| Target at F8 | 74.46 ± 25.45 | 51.26 ± 20.97 | 9.97 | 0.003 ** |
| Target at P7 | 69.22 ± 19.62 | 47.59 ± 19.68 | 12.08 | 0.001 ** |
| Target at P8 | 56.37 ± 15.22 | 44.11 ± 16.69 | 5.82 | 0.021 ** |
| Non-target at P3 | 73.72 ± 23.48 | 53.97 ± 19.95 | 8.27 | 0.007 ** |
| Non-target at P4 | 76.16 ± 21.59 | 56.52 ± 22.92 | 7.74 | 0.008 ** |
| Non-target at P8 | 66.49 ± 22.90 | 45.90 ± 17.67 | 10.25 | 0.003 ** |
| Non-Kanizsa at F7 | 75.81 ± 25.23 | 48.48 ± 16.71 | 16.62 | <0.001 *** |
| Non-Kanizsa at F8 | 75.92 ± 21.13 | 50.91 ± 21.58 | 13.66 | 0.001** |
| Non-Kanizsa at P7 | 69.82 ± 27.74 | 47.01 ± 18.62 | 9.49 | 0.004 ** |
| Non-Kanizsa at P8 | 59.99 ± 25.29 | 44.28 ± 19.43 | 4.91 | 0.033 * |
| Negative/descending area, evoked | ||||
| Target at F8 | 48.76 ± 4.57 | 41.01 ± 15.70 | 4.29 | 0.045 * |
| Non-target at F7 | 51.29 ± 3.70 | 39.68 ± 15.57 | 10.03 | 0.003 ** |
| Non-target at F8 | 54.69 ± 4.79 | 39.87 ± 15.38 | 16.17 | <0.001 *** |
| Non-target at P7 | 52.46 ± 6.38 | 41.45 ± 15.13 | 8.65 | 0.006 ** |
| Non-target at P8 | 52.07 ± 5.63 | 43.60 ± 16.97 | 4.29 | 0.045 * |
| Non-Kanizsa at F7 | 51.34 ± 5.25 | 38.80 ± 14.94 | 12.02 | 0.001 ** |
| Non-Kanizsa at F8 | 51.94 ± 4.85 | 39.48 ± 15.63 | 11.07 | 0.002 ** |
| Non-Kanizsa at P7 | 53.71 ± 5.19 | 40.77 ± 16.00 | 12.02 | 0.001 ** |
| Non-Kanizsa at P8 | 51.13 ± 5.32 | 40.48 ± 15.68 | 7.92 | 0.008 ** |
| Negative/descending area, induced | ||||
| Non-target at P3 | 56.80 ± 20.26 | 42.71 ± 16.14 | 5.98 | 0.019 ** |
| Non-target at P4 | 56.37 ± 19.48 | 43.05 ± 18.45 | 4.93 | 0.032 * |
| Non-target at P7 | 70.30 ± 22.90 | 49.33 ± 19.63 | 9.73 | 0.003 ** |
| Non-target at P8 | 65.66 ± 24.02 | 46.18 ± 22.20 | 7.11 | 0.011* |
| Non-Kanizsa at P3 | 61.07 ± 20.00 | 44.24 ± 20.33 | 6.94 | 0.012 * |
| Non-Kanizsa at P7 | 70.99 ± 22.65 | 46.82 ± 20.50 | 12.23 | 0.001 ** |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Casanova, M.F.; Shaban, M.; Ghazal, M.; El-Baz, A.S.; Casanova, E.L.; Opris, I.; Sokhadze, E.M. Effects of Transcranial Magnetic Stimulation Therapy on Evoked and Induced Gamma Oscillations in Children with Autism Spectrum Disorder. Brain Sci. 2020, 10, 423. https://doi.org/10.3390/brainsci10070423
Casanova MF, Shaban M, Ghazal M, El-Baz AS, Casanova EL, Opris I, Sokhadze EM. Effects of Transcranial Magnetic Stimulation Therapy on Evoked and Induced Gamma Oscillations in Children with Autism Spectrum Disorder. Brain Sciences. 2020; 10(7):423. https://doi.org/10.3390/brainsci10070423
Chicago/Turabian StyleCasanova, Manuel F., Mohamed Shaban, Mohammed Ghazal, Ayman S. El-Baz, Emily L. Casanova, Ioan Opris, and Estate M. Sokhadze. 2020. "Effects of Transcranial Magnetic Stimulation Therapy on Evoked and Induced Gamma Oscillations in Children with Autism Spectrum Disorder" Brain Sciences 10, no. 7: 423. https://doi.org/10.3390/brainsci10070423
APA StyleCasanova, M. F., Shaban, M., Ghazal, M., El-Baz, A. S., Casanova, E. L., Opris, I., & Sokhadze, E. M. (2020). Effects of Transcranial Magnetic Stimulation Therapy on Evoked and Induced Gamma Oscillations in Children with Autism Spectrum Disorder. Brain Sciences, 10(7), 423. https://doi.org/10.3390/brainsci10070423