Effects of Phosphatidylserine Source of Docosahexaenoic Acid on Cerebellar Development in Preterm Pigs



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Experimental Design, Preterm Pig Model, and Tissue Collection
2.2. Immunohistochemistry
2.3. Imaging, Cell Counts, and Statistical Analysis
3. Results
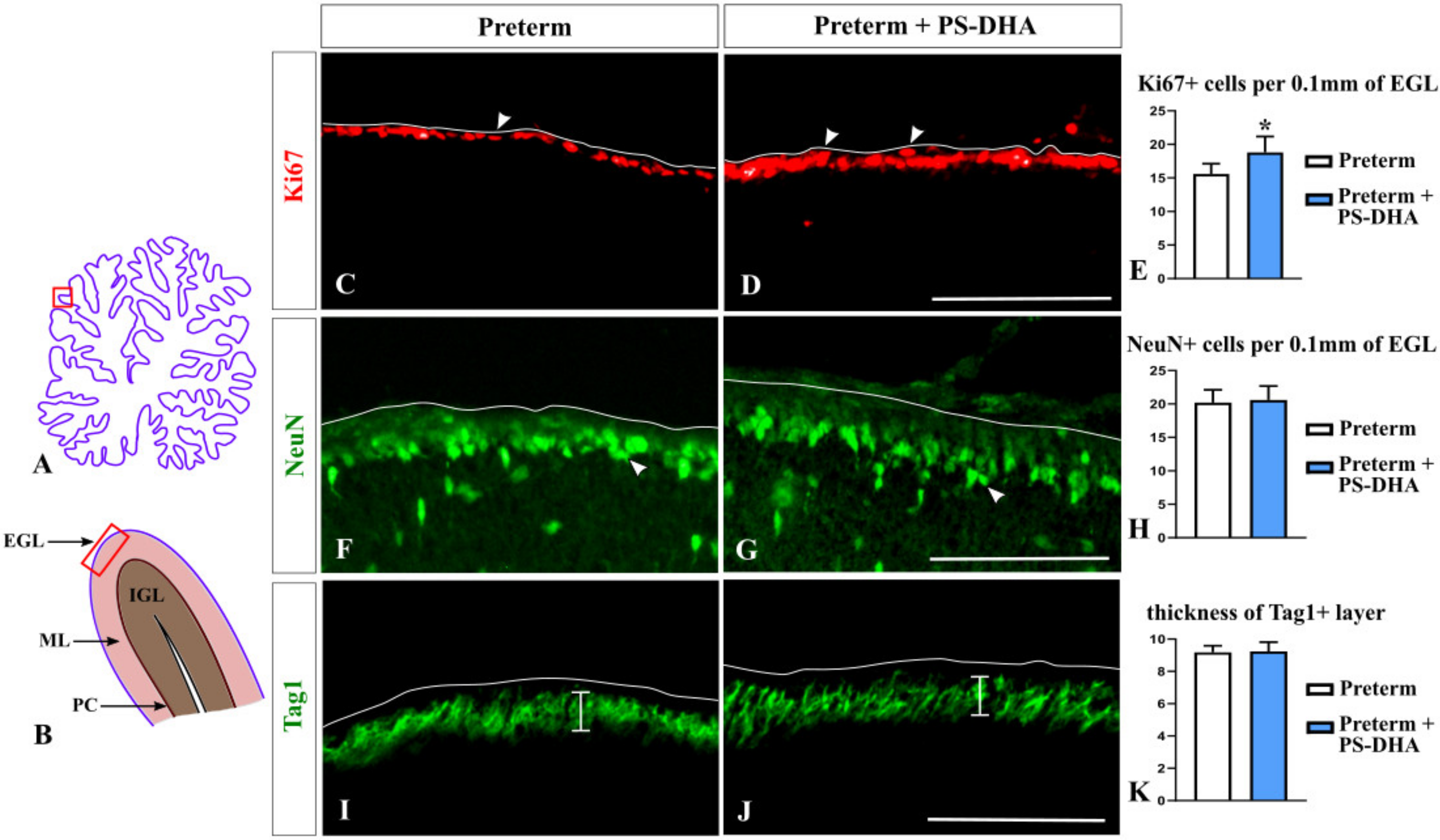
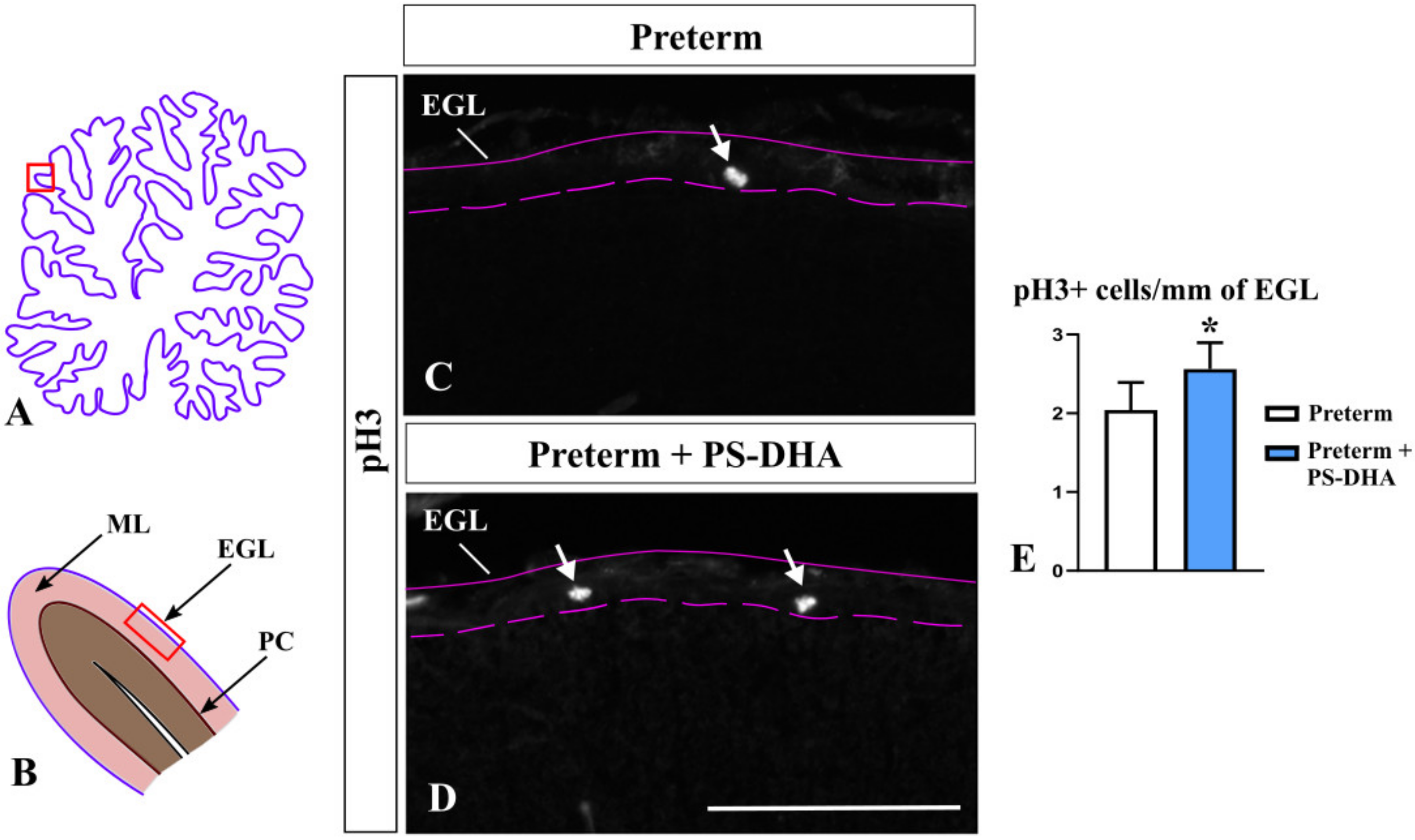
3.1. PS-DHA Promotes Proliferation of Cerebellar Granule Cell Precursors in Pigs Born Preterm
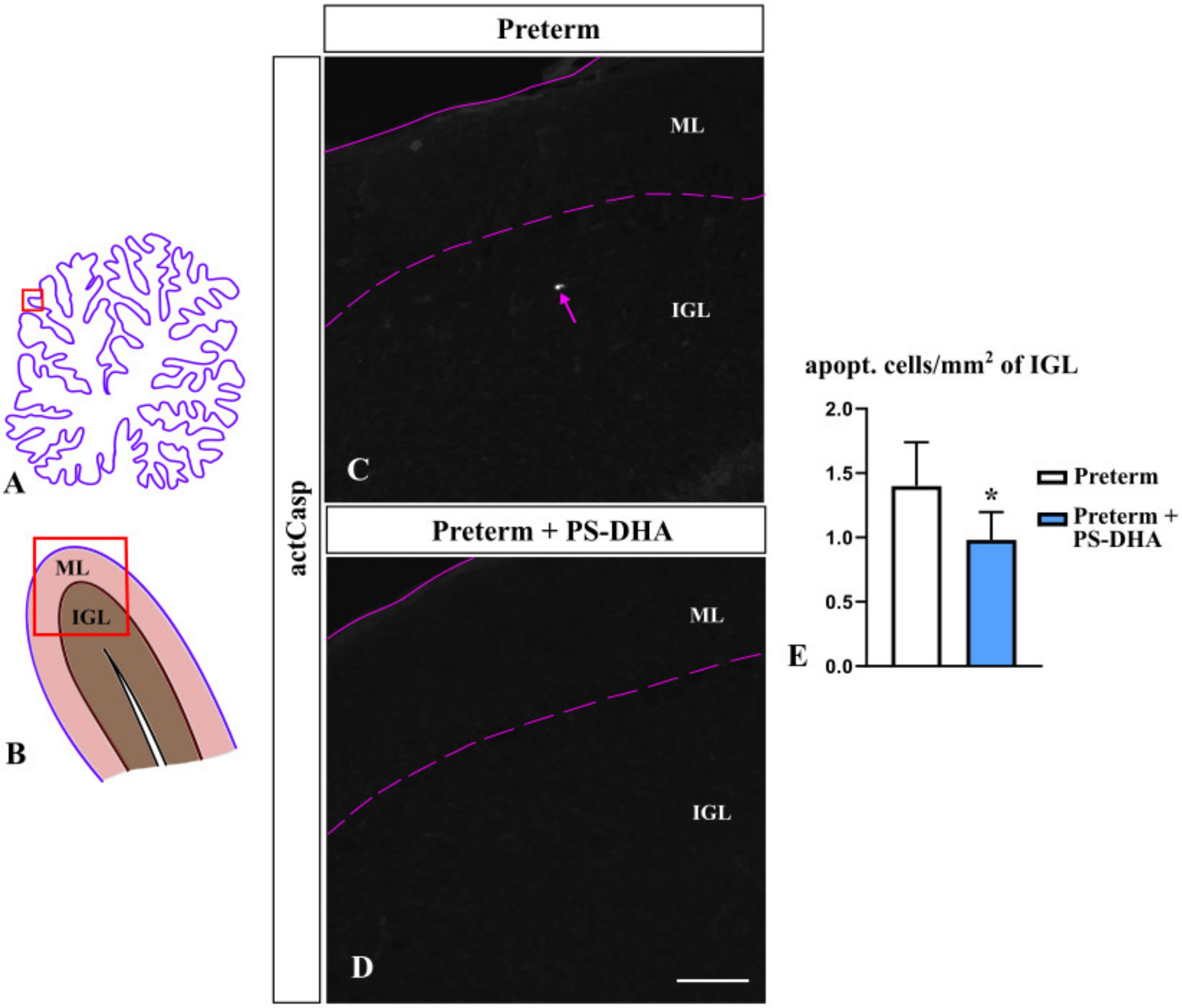
3.2. PS-DHA Promotes Cell Survival in the IGL of Pigs Born Preterm
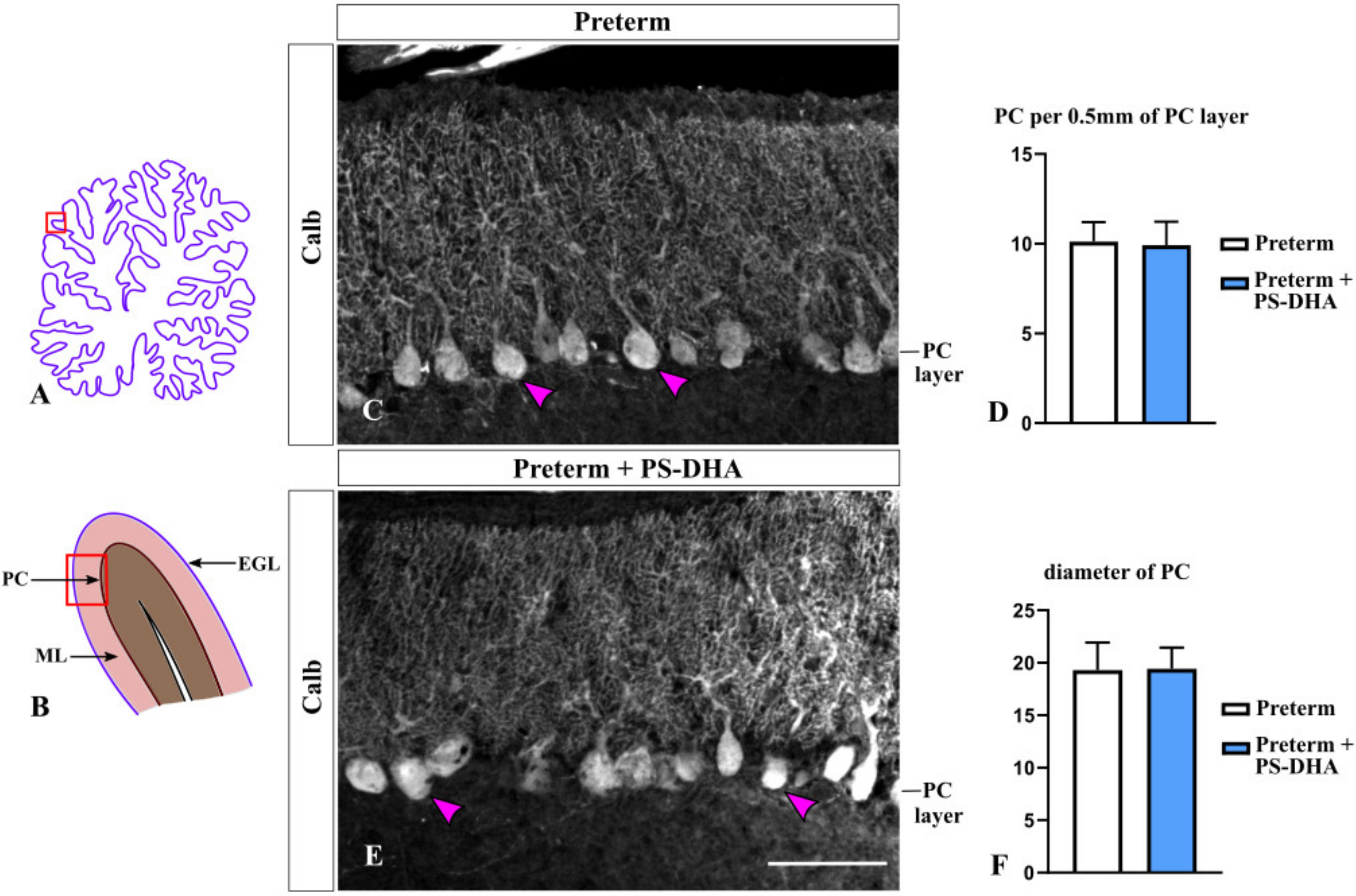
3.3. Purkinje Cells Are Not Affected by PS-DHA Supplementation of Preterm Pigs
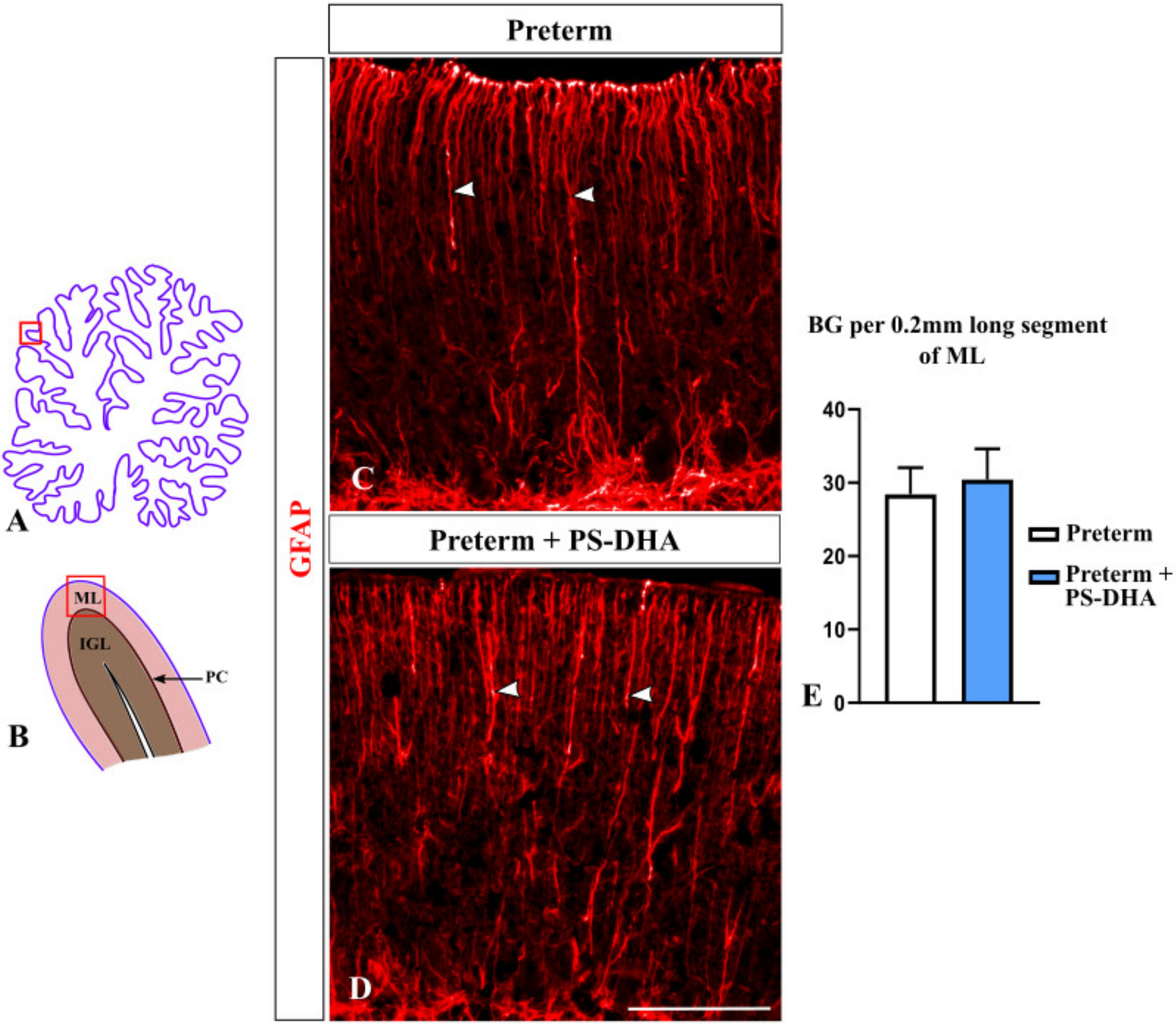
3.4. Bergmann Glia Are Not Affected by PS-DHA Supplementation of Preterm Pigs
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Goldenberg, R.L.; Culhane, J.F.; Iams, J.D.; Romero, R. Epidemiology and causes of preterm birth. Lancet 2008, 371, 75–84. [Google Scholar] [CrossRef]
- Spittle, A.; Orton, J.; Anderson, P.; Boyd, R.; Doyle, L. Early Developmental Intervention Programmes Provided Post Hospital Discharge to Prevent Motor and Cognitive Impairment in Preterm Infants. Cochrane Database Syst. Rev. 2015, 11, CD005495. [Google Scholar] [CrossRef] [PubMed]
- Haldipur, P.; Bharti, U.; Alberti, C.; Sarkar, C.; Gulati, G.; Iyengar, S.; Gressens, P.; Mani, S. Preterm delivery disrupts the developmental program of the cerebellum. PLoS ONE 2011, 6, e23449. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Iskusnykh, I.; Buddington, R.; Chizhikov, V. Preterm birth disrupts cerebellar development by affecting granule cell proliferation program and Bergmann glia. Exp. Neurol. 2018, 306, 209–221. [Google Scholar] [CrossRef]
- Stoodley, C.; MacMore, J.; Makris, N.; Sherman, J.; Schmahmann, J. Location of lesion determines motor vs. cognitive consequences in patients with cerebellar stroke. Neuroimage Clin. 2016, 12, 765–775. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Butts, T.; Green, M.; Wingate, R. Development of the cerebellum: Simple steps to make a ‘little brain’. Development 2014, 141, 4031–4041. [Google Scholar] [CrossRef] [Green Version]
- Basson, M.; Wingate, R. Congenital hypoplasia of the cerebellum: Developmental causes and behavioral consequences. Front. Neuroanat. 2013, 7, 29. [Google Scholar] [CrossRef] [Green Version]
- Haldipur, P.; Dang, D.; Millen, K. Embryology. In Handbook of Clinical Neurology; Elsevier: Amsterdam, The Netherlands, 2018; Volume 154, pp. 29–44. [Google Scholar]
- Chizhikov, V.; Millen, K. Neurogenesis in the cerebellum. In Comprehensive Developmental Neuroscience; Rakic, P., Rubenstein, J., Eds.; Elsevier: Amsterdam, The Netherlands, 2013. [Google Scholar]
- Leto., K.; Arancillo, M.; Becker, E.; Buffo, A.; Chiang, C.; Ding, B.; Dobyns, W.; Dusart, I.; Haldipur, P.; Hatten, M.; et al. Consensus Paper: Cerebellar Development. Cerebellum 2016, 15, 789–828. [Google Scholar] [CrossRef]
- Marzban, H.; Del Bigio, M.; Alizadeh, J.; Ghavami, S.; Zachariah, R.; Rastegar, M. Cellular commitment in the developing cerebellum. Front. Cell Neurosci. 2014, 8, 450. [Google Scholar]
- Wullimann, M.; Mueller, T.; Distel, M.; Babaryka, A.; Grothe, B.; Köster, R. The Long Adventurous Journey of Rhombic Lip Cells in Jawed Vertebrates: A Comparative Developmental Analysis. Front. Neuroanat. 2011, 5, 27. [Google Scholar] [CrossRef] [Green Version]
- Biran, V.; Verney, C.; Ferriero, D. Perinatal cerebellar injury in human and animal models. Neurol Res. Int. 2012, 858929. [Google Scholar] [CrossRef] [PubMed]
- Leung, A.; Li, J. The molecular pathway regulating Bergmann glia and folia generation in the cerebellum. Cerebellum 2018, 17, 42–48. [Google Scholar] [CrossRef] [PubMed]
- Sathyanesan, A.; Kundu, S.; Abbah, J.; Gallo, V. Neonatal brain injury causes cerebellar learning deficits and Purkinje cell dysfunction. Nature Commun. 2018, 9, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Buddington, R.; Chizhikov, V.; Iskusnykh, I.; Sable, H.; Sable, J.; Holloway, Z.; Katzir, T.; van der Merwe, M.; Yakimkova, T.; Buddington, K.; et al. A Phosphatidylserine Source of Docosahexanoic Acid Improves Neurodevelopment and Survival of Preterm Pigs. Nutrients 2018, 10, 637. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lauritzen, L.; Brambilla, P.; Mazzocchi, A.; Harsløf, B.; Ciappolino, V.; Agostoni, C. DHA Effects in Brain Development and Function. Nutrients 2016, 8, 6. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mun, J.; Legette, L.; Ikonte, C.; Mitmesser, S. Choline and DHA in Maternal and Infant Nutrition: Synergistic Implications in Brain and Eye Health. Nutrients 2019, 11, 1125. [Google Scholar] [CrossRef] [Green Version]
- Smith, S.L.; Rouse, C.A. Docosahexaenoic Acid and the Preterm Infant. Matern. Health Neonatol. Perinatol. 2017, 3, 22. [Google Scholar] [CrossRef] [Green Version]
- Tam, E.; Chau, V.; Barkovich, A.; Ferriero, D.; Miller, S.; Rogers, E.; Grunau, R.; Synnes, A.; Xu, D.; Foong, J.; et al. Early postnatal docosahexaenoic acid levels and improved preterm brain development. Pediatr. Res. 2016, 79, 723–730. [Google Scholar] [CrossRef] [Green Version]
- Sabel, K.; Strandvik, B.; Petzold, M.; Lundqvist-Persson, C. Motor, mental and behavioral developments in infancy are associated with fatty acid pattern in breast milk and plasma of premature infants. Prostaglandins Leukot. Essent. Fat. Acids 2012, 86, 183–188. [Google Scholar] [CrossRef]
- Henriksen, C.; Haugholt, K.; Lindgren, M.; Aurvåg, A.; Rønnestad, A.; Grønn, M.; Solberg, R.; Moen, A.; Nakstad, B.; Berge, R.K.; et al. Improved cognitive development among preterm infants attributable to early supplementation of human milk with docosahexaenoic acid and arachidonic acid. Pediatrics 2008, 121, 1137–1145. [Google Scholar] [CrossRef]
- Vance, J.; Steenbergen, R. Metabolism and functions of phosphatidylserine. Prog. Lipid Res. 2005, 44, 207–234. [Google Scholar] [CrossRef] [PubMed]
- Vaisman, N.; Pelled, D. n-3 phosphatidylserine attenuated scopolamine-induced amnesia in middle-aged rats. Prog. Neuro Psychopharmacol. Biol Psychiatry 2009, 33, 952–959. [Google Scholar] [CrossRef] [PubMed]
- Chouinard-Watkins, R.; Lacombe, R.; Metherel, A.; Masoodi, M.; Bazinet, R.P. DHA Esterified to Phosphatidylserine or Phosphatidylcholine is More Efficient at Targeting the Brain than DHA Esterified to Triacylglycerol. Mol. Nutr. Food Res. 2019, 63, e1801224. [Google Scholar] [CrossRef] [PubMed]
- Nakashima, K.; Umeshima, H.; Kengaku, M. Cerebellar Granule Cells Are Predominantly Generated by Terminal Symmetric Divisions of Granule Cell Precursors. Dev. Dyn. 2015, 244, 748–758. [Google Scholar] [CrossRef]
- Wen, J.; Yang, H.; Zhou, B.; Lou, H.; Duan, S. β-Catenin Is Critical for Cerebellar Foliation and Lamination. PLoS ONE 2013, 8, e64451. [Google Scholar] [CrossRef] [Green Version]
- Van Hove, I.; Verslegers, M.; Buyens, T.; Delorme, N.; Lemmens, K.; Stroobants, S.; Gantois, I.; D’Hooge, R.; Moons, L. An Aberrant Cerebellar Development in Mice Lacking Matrix metalloproteinase-3. Mol. Neurobiol. 2012, 45, 17–29. [Google Scholar] [CrossRef]
- Wang, L.; Liu, Y. Signaling pathways in cerebellar granule cells development. Am. J. Stem Cells 2019, 8, 1–6. [Google Scholar]
- Miterko, L.; White, J.; Lin, T.; Brown, A.; O’Donovan, K.; Sillitoe, R. Persistent Motor Dysfunction Despite Homeostatic Rescue of Cerebellar Morphogenesis in the Car8 Waddles Mutant Mouse. Neural Dev. 2019, 14, 6. [Google Scholar] [CrossRef] [Green Version]
- Sillitoe, R.V.; Joyner, A.L. Morphology, molecular codes, and circuitry produce the three-dimensional complexity of the cerebellum. Annu. Rev. Cell Dev. Biol. 2007, 23, 549–577. [Google Scholar] [CrossRef]
- Fernandez, C.; Tatard, V.; Bertrand, N.; Dahmane, N. Differential modulation of Sonic-hedgehog-induced cerebellar granule cell precursor proliferation by the IGF signaling network. Dev. Neurosci. 2010, 32, 59–70. [Google Scholar] [CrossRef]
- Rio, C.; Rieff, H.; Qi, P.; Khurana, T.; Corfas, G. Neuregulin and erbB receptors play a critical role in neuronal migration. Neuron 1997, 19, 39–50. [Google Scholar] [CrossRef] [Green Version]
- Ju, J.; Liu, Q.; Zhang, Y.; Liu, Y.; Jiang, M.; Zhang, L.; He, X.; Peng, C.; Zheng, T.; Lu, Q.; et al. Olig2 regulates Purkinje cell generation in the early developing mouse cerebellum. Sci. Rep. 2016, 6, 30711. [Google Scholar] [CrossRef] [PubMed]
- Bellamy, T.C. Interactions between Purkinje neurones and Bergmann glia. Cerebellum 2006, 5, 116–126. [Google Scholar] [CrossRef] [PubMed]
- Komine, O.; Nagaoka, M.; Watase, K.; Gutmann, D.; Tanigaki, K.; Honjo, T.; Radtke, F.; Saito, T.; Chiba, S.; Tanaka, K. The Monolayer Formation of Bergmann Glial Cells Is Regulated by Notch/RBP-J Signaling. Dev. Biol. 2007, 311, 238–250. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Darmaun, D.; Lapillonne, A.; Simeoni, U.; Picaud, J.C.; Rozé, J.C.; Saliba, E.; Bocquet, A.; Chouraqui, J.; Dupont, C.; Feillet, F.; et al. Parenteral nutrition for preterm infants: Issues and strategy. Arch. Pediatr. 2018, 25, 286–294. [Google Scholar] [CrossRef] [PubMed]
- Hafström, M.; Källén, K.; Serenius, F.; Maršál, K.; Rehn, E.; Drake, H.; Ådén, U.; Farooqi, A.; Thorngren-Jerneck, K.; Strömberg, B. Cerebral Palsy in Extremely Preterm Infants. Pediatrics 2018, 141, e20171433. [Google Scholar] [CrossRef] [Green Version]
- Lean, R.E.; Paul, R.A.; Smyser, T.A.; Smyser, C.D.; Rogers, C.E. Social Adversity and Cognitive, Language, and Motor Development of Very Preterm Children from 2 to 5 Years of Age. J. Pediatr. 2018, 203, 177–184. [Google Scholar] [CrossRef]
- Allotey, J.; Zamora, J.; Cheong-See, F.; Kalidindi, M.; Arroyo-Manzano, D.; Asztalos, E.; van der Post, J.; Mol, B.; Moore, D.; Moore, D.; et al. Cognitive, Motor, Behavioural and Academic Performances of Children Born Preterm: A Meta-Analysis and Systematic Review Involving 64 061 Children. BJOG 2018, 125, 16–25. [Google Scholar] [CrossRef] [Green Version]
- Srinivasan, L.; Allsop, J.; Counsell, S.; Boardman, J.; Edwards, A.; Rutherford, M. Smaller Cerebellar Volumes in Very Preterm Infants at Term-Equivalent Age are Associated with the Presence of Supratentorial Lesions. Am. J. Neuroradiol. 2006, 27, 573–579. [Google Scholar]
- Reeber, S.; Otis, T.; Sillitoe, R. New roles for the cerebellum in health and disease. Front. Syst. Neurosci. 2013, 7, 83. [Google Scholar] [CrossRef] [Green Version]
- Kopecky, B.; Decook, R.; Fritzsch, B. Mutational ataxia resulting from abnormal vestibular acquisition and processing is partially compensated for. Behav. Neurosci. 2012, 126, 301–313. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Glade, M.J.; Smith, K. Phosphatidylserine and the human brain. Nutrition 2015, 31, 781–786. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.Y.; Huang, B.X.; Spector, A.A. Phosphatidylserine in the brain: Metabolism and function. Prog. Lipid Res. 2014, 56, 1–18. [Google Scholar] [CrossRef] [Green Version]
- Weiser, M.J.; Butt, C.M.; Mohajeri, M.H. Docosahexaenoic Acid and Cognition throughout the Lifespan. Nutrients 2016, 8, 99. [Google Scholar] [CrossRef] [PubMed]
- Van der Heijden, M.E.; Sillitoe, R.V. Interactions Between Purkinje Cells and Granule Cells Coordinate the Development of Functional Cerebellar Circuits. Neuroscience 2020. [Google Scholar] [CrossRef]
- Brossard-Racine, M.; du Plessis, A.; Limperopoulos, C. Developmental cerebellar cognitive affective syndrome in ex-preterm survivors following cerebellar injury. Cerebellum 2015, 14, 151–164. [Google Scholar] [CrossRef] [Green Version]
- Wagner, M.; Kim, T.; Savall, J.; Schnitzer, M.; Luo, L. Cerebellar granule cells encode the expectation of reward. Nature 2017, 544, 96–100. [Google Scholar] [CrossRef] [Green Version]
- Roussel, M.; Hatten, M. Cerebellum development and medulloblastoma. In Current Topics in Developmental Biology; Academic Press: Cambridge, MA, USA, 2011; Volume 94, pp. 235–282. [Google Scholar]
- Solecki, D.; Liu, X.; Tomoda, T.; Fang, Y.; Hatten, M. Activated Notch2 signaling inhibits differentiation of cerebellar granule neuron precursors by maintaining proliferation. Neuron 2001, 31, 557–568. [Google Scholar] [CrossRef] [Green Version]
- Schmitz, T.; Scheuer, T.; Bührer, C. Development of the cerebellar white matter is impaired by postnatal hyperoxia and protected by minocycline. Mol. Cell Pediatr. 2014, 1, A6. [Google Scholar] [CrossRef] [Green Version]
- Huun, M.; Garberg, H.; Escobar, J.; Chafer, C.; Vento, M.; Holme, I.; Saugstad, O.; Solberg, R. DHA reduces oxidative stress following hypoxia-ischemia in newborn piglets: A study of lipid peroxidation products in urine and plasma. J. Perinat. Med. 2018, 46, 209–217. [Google Scholar] [CrossRef]





© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chizhikov, D.; Buddington, R.K.; Iskusnykh, I.Y. Effects of Phosphatidylserine Source of Docosahexaenoic Acid on Cerebellar Development in Preterm Pigs. Brain Sci. 2020, 10, 475. https://doi.org/10.3390/brainsci10080475
Chizhikov D, Buddington RK, Iskusnykh IY. Effects of Phosphatidylserine Source of Docosahexaenoic Acid on Cerebellar Development in Preterm Pigs. Brain Sciences. 2020; 10(8):475. https://doi.org/10.3390/brainsci10080475
Chicago/Turabian StyleChizhikov, Daniel, Randal K. Buddington, and Igor Y. Iskusnykh. 2020. "Effects of Phosphatidylserine Source of Docosahexaenoic Acid on Cerebellar Development in Preterm Pigs" Brain Sciences 10, no. 8: 475. https://doi.org/10.3390/brainsci10080475
APA StyleChizhikov, D., Buddington, R. K., & Iskusnykh, I. Y. (2020). Effects of Phosphatidylserine Source of Docosahexaenoic Acid on Cerebellar Development in Preterm Pigs. Brain Sciences, 10(8), 475. https://doi.org/10.3390/brainsci10080475