Frontal Cortical Modulation of Temporal Visual Cross-Modal Re-organization in Adults with Hearing Loss


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Participants and Ethics Statement
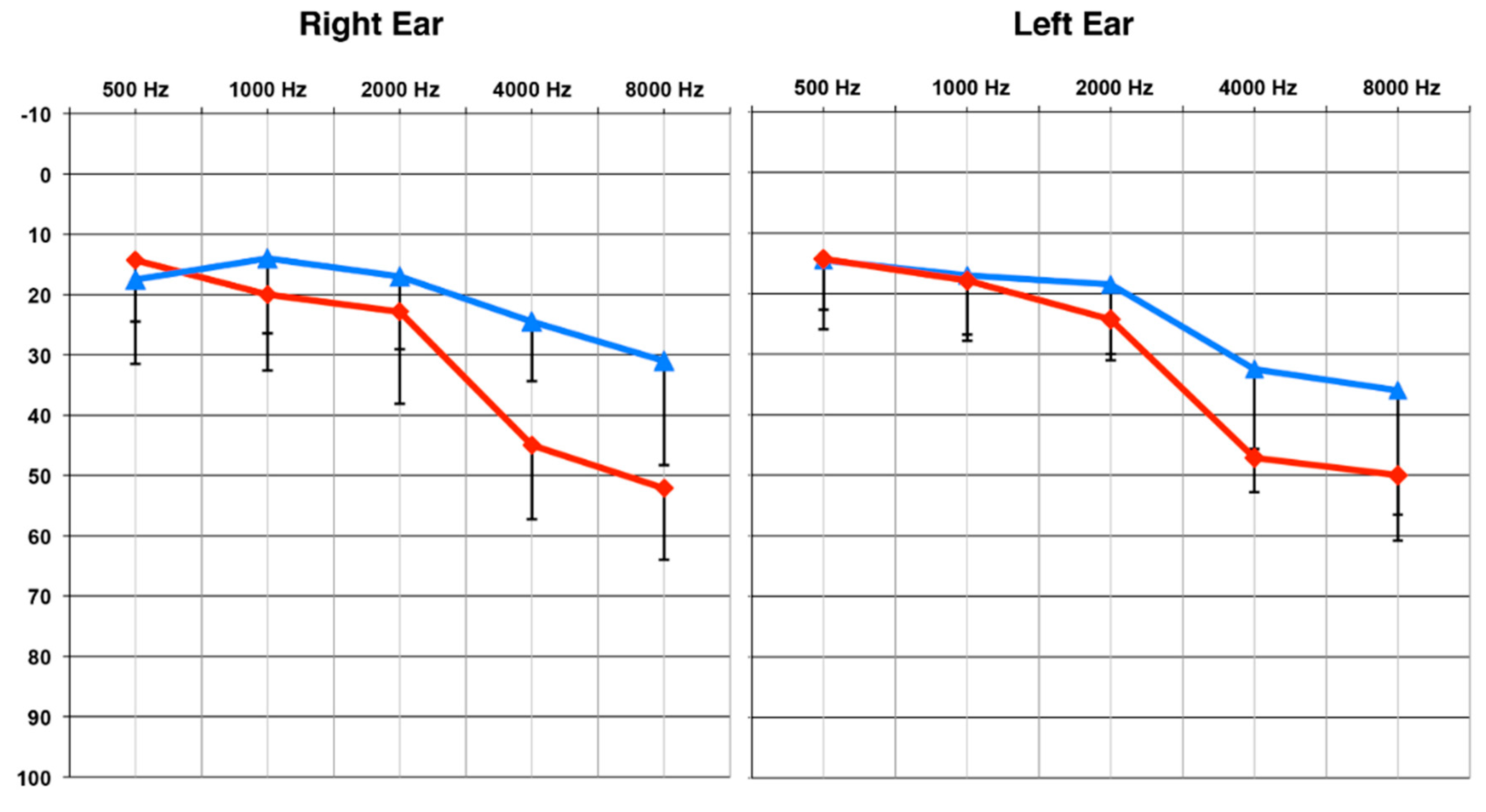
2.2. Auditory Behavioral Testing: Measure of Speech Perception in Noise
2.3. EEG Procedures
2.3.1. Visual Stimuli
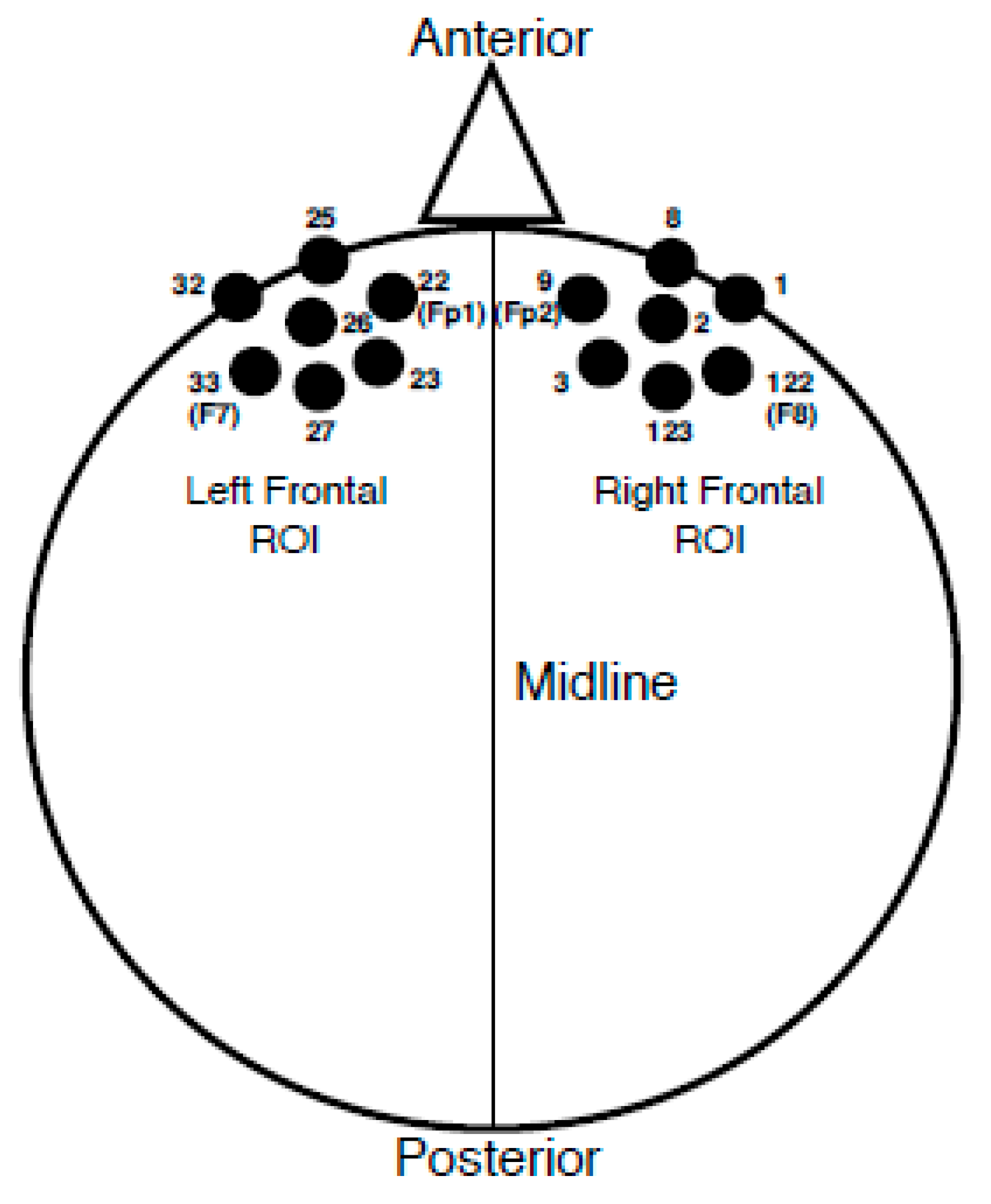
2.3.2. EEG Recording and Analyses
2.4. Current Density Reconstruction
2.5. Statistical Analyses
3. Results
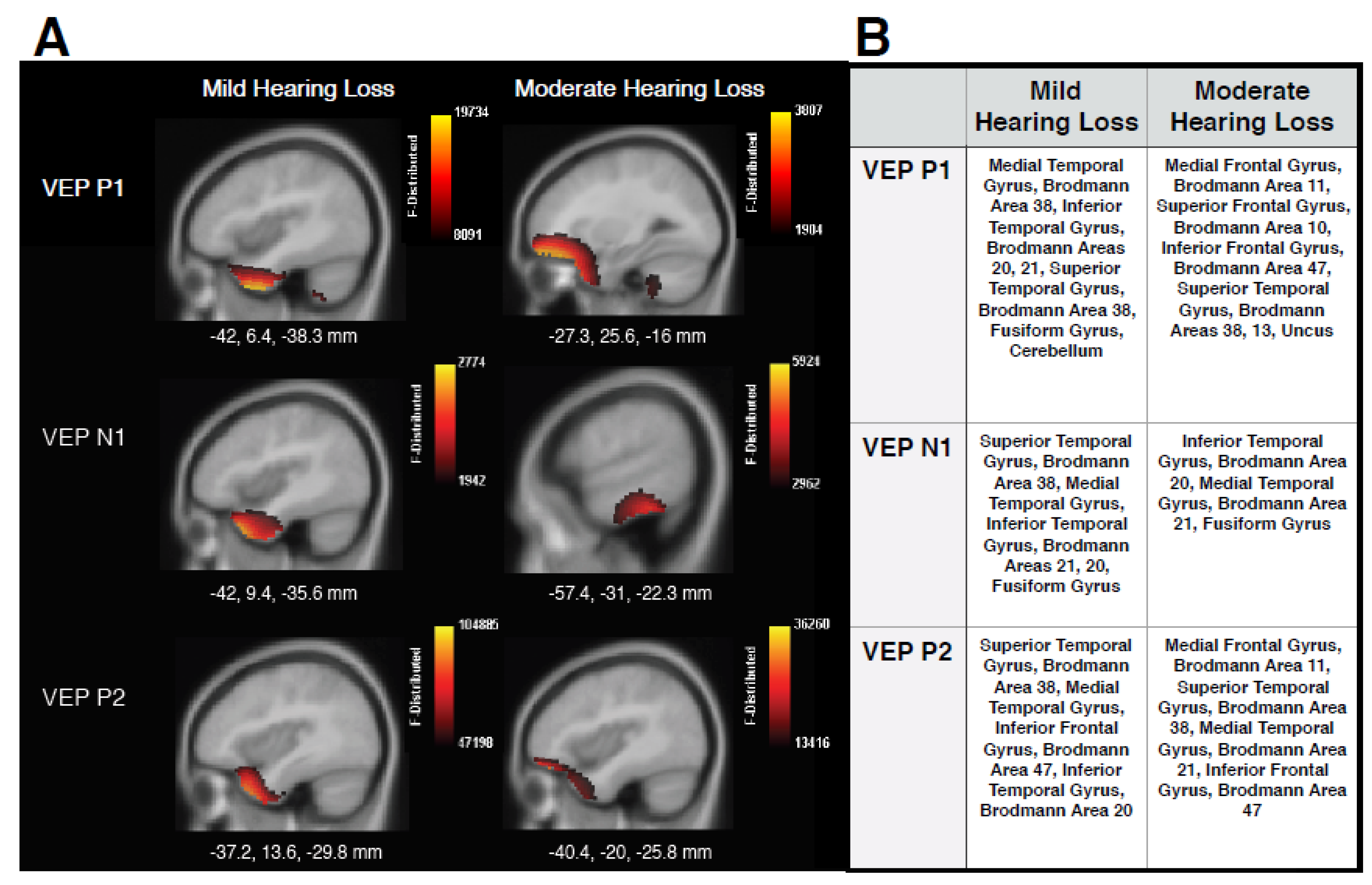
3.1. Current Density Reconstructions
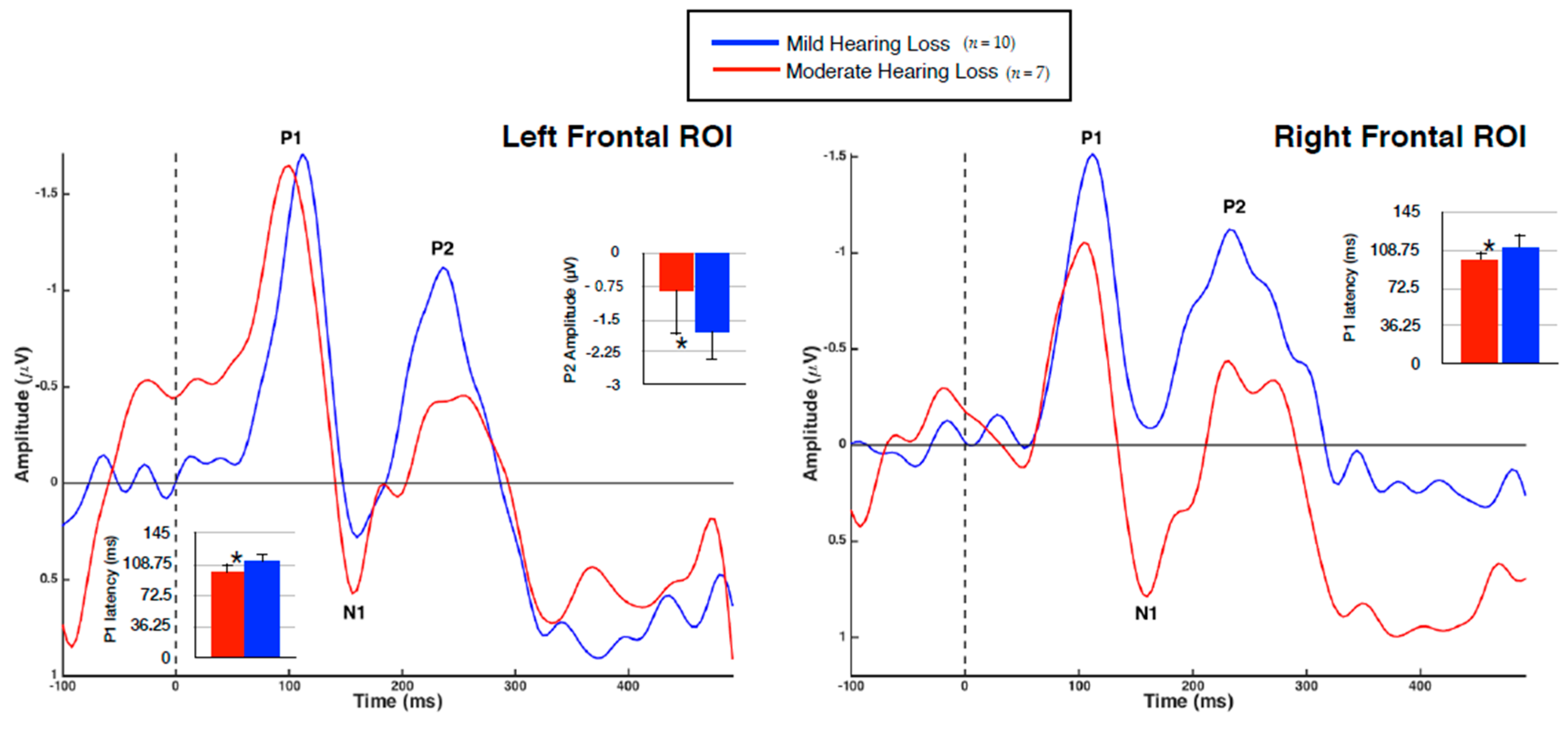
3.2. Visual Evoked Potentials
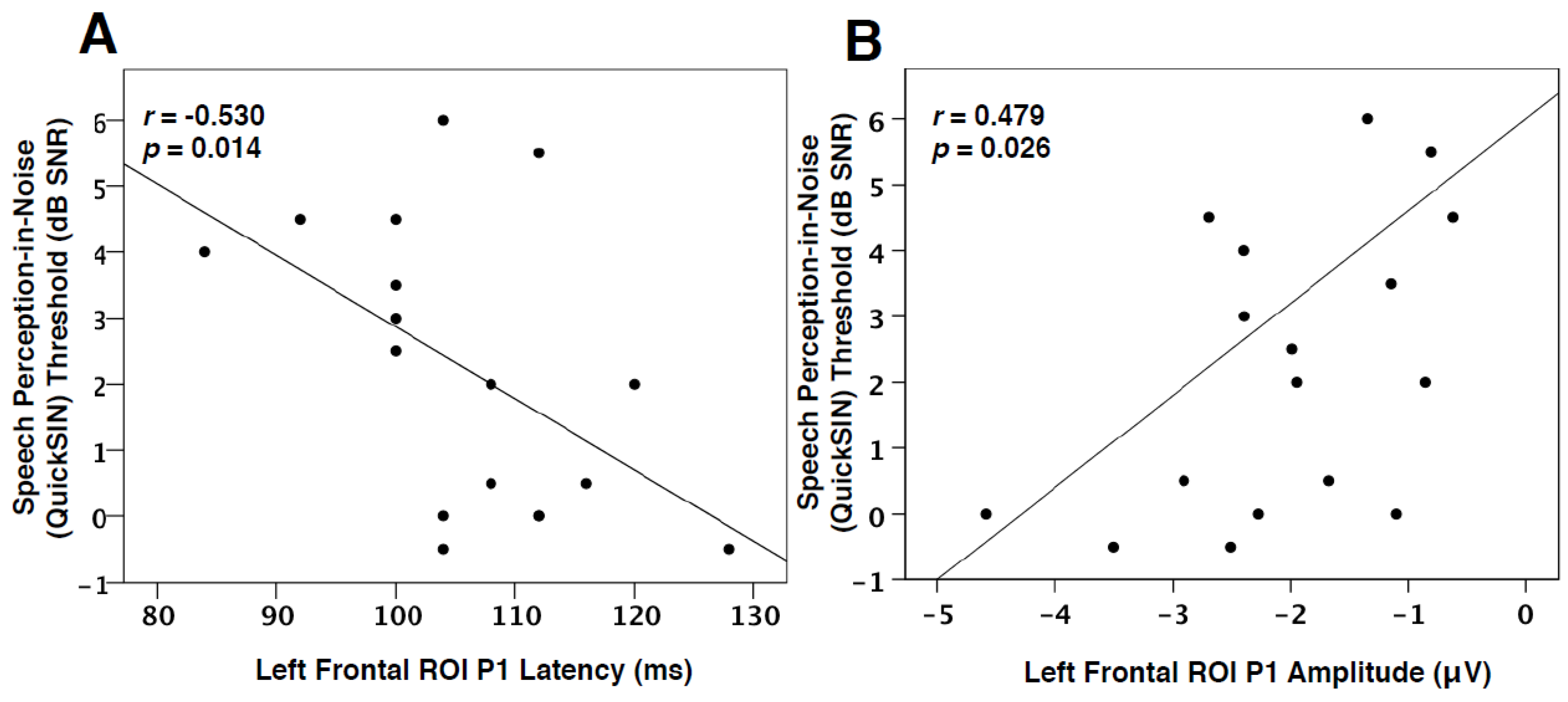
3.3. Speech Perception in Noise and VEPs
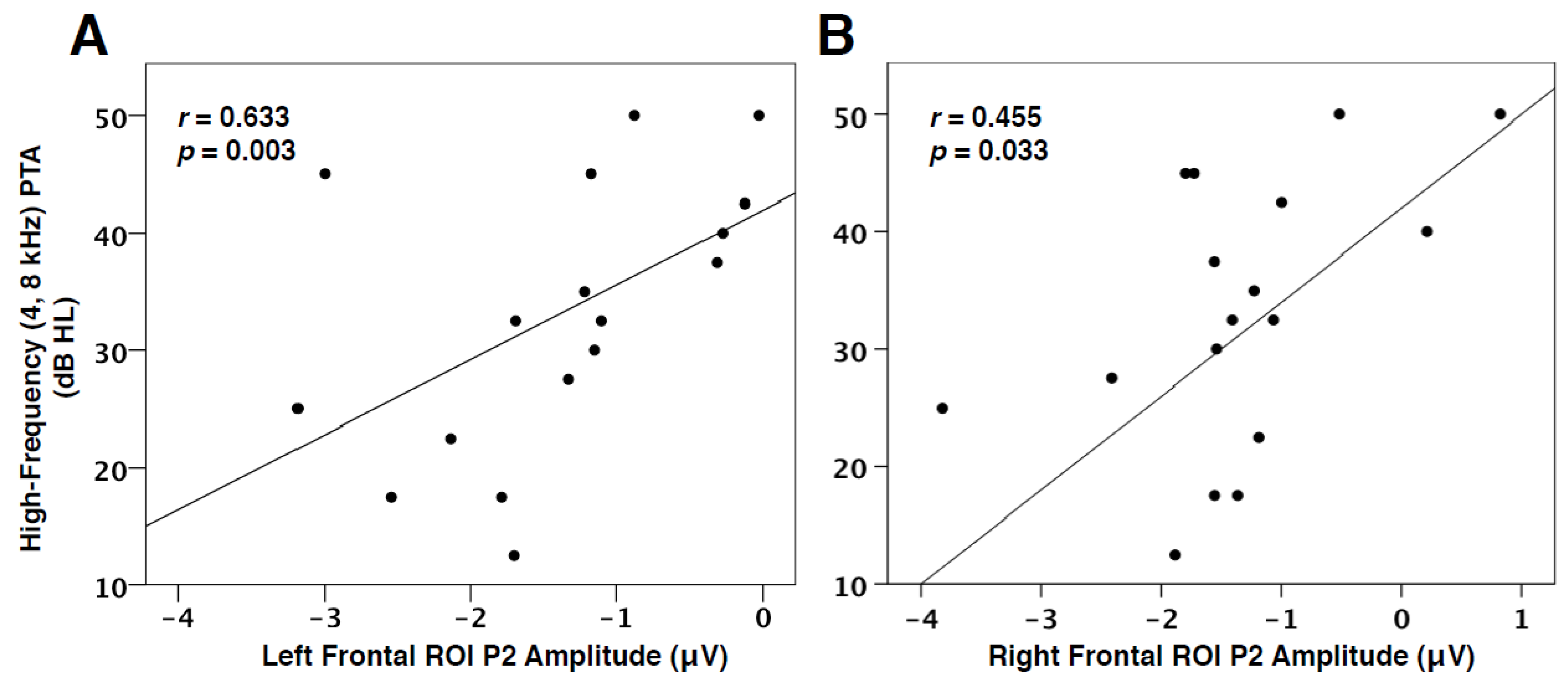
3.4. Hearing Loss and VEP Amplitude
4. Discussion
4.1. Frontal Top-Down Modulation in Hearing Loss
4.2. Evoked Potential Indices of Cortical Plasticity in Hearing Loss
4.3. Visual Cortical Plasticity and Auditory Behavioral Outcomes
5. Summary and Conclusion
Author Contributions
Funding
Conflicts of Interest
References
- Alhanbali, S.; Dawes, P.; Lloyd, S.; Munro, K.J. Self-reported listening-related effort and fatigue in hearing-impaired adults. Ear Hear. 2017, 38, 39–48. [Google Scholar] [CrossRef]
- Campbell, J.; Sharma, A. Cross-modal re-organization in adults with early stage hearing loss. PLoS ONE 2014, 9, e90594. [Google Scholar] [CrossRef] [Green Version]
- Deal, A.; Betz, J.; Yaffe, K.; Harris, T.; Purchase-Helzner, E.; Satterfield, S.; Pratt, S.; Govil, N.; Simonsick, E.M.; Lin, F.R.; et al. Hearing impairment and incident dementia and cognitive decline in older adults: The health ABC study. J. Gerontol. A Biol. Sci. Med. Sci. 2017, 72, 703–709. [Google Scholar] [CrossRef] [PubMed]
- Lin, F.R. Hearing loss and cognition among older adults in the United States. J. Gerontol. A Biol. Sci. Med. Sci. 2011, 66, 1131–1136. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jorgensen, L.E.; Palmer, C.V.; Pratt, S.; Erickson, K.; Moncrieff, D. The effect of decreased audibility on MMSE performance: A measure commonly used for diagnosing dementia. J. Am. Acad. Audiol. 2016, 27, 311–323. [Google Scholar] [CrossRef] [PubMed]
- Bavelier, D.; Hirshorn, E.A. I see where you’re hearing: How cross-modal plasticity may exploit homologous brain structures. Nat. Neurosci. 2010, 13, 1309–1311. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fine, I.; Finney, E.M.; Boynton, G.M.; Dobkins, K.R. Comparing the effects of auditory deprivation and sign language within the auditory and visual cortex. J. Cogn. Neurosci. 2005, 17, 1621–1637. [Google Scholar] [CrossRef]
- Finney, E.M.; Fine, I.; Dobkins, K.R. Visual stimuli activate auditory cortex in the deaf. Nat. Neurosci. 2001, 4, 1171–1173. [Google Scholar] [CrossRef]
- Finney, E.M.; Clementz, B.A.; Hickok, G.; Dobkins, K.R. Visual stimuli activate auditory cortex in deaf subjects: Evidence from MEG. Neuroreport 2003, 14, 1425–1427. [Google Scholar] [CrossRef]
- Lomber, S.G.; Meredith, M.A.; Kral, A. Cross-modal plasticity in specific auditory cortices underlies visual compensations in the deaf. Nat. Neurosci. 2010, 13, 1421–1427. [Google Scholar] [CrossRef]
- Meredith, M.A.; Kryklywy, J.; McMillan, A.J.; Malhotra, S.; Lum-Tai, R.; Lomber, S.G. Crossmodal reorganization in the early deaf switches sensory, but not behavioral roles of auditory cortex. Proc. Natl. Acad. Sci. USA 2011, 108, 8856–8861. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bosworth, R.G.; Dobkins, K.R. The effects of spatial attention on motion processing in deaf signers, hearing signers, and hearing nonsigners. Brain Cogn. 2002, 49, 152–169. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ding, H.; Qin, W.; Liang, M.; Ming, D.; Wan, B.; Li, Q.; Yu, C. Cross-modal activation of auditory regions during visuo-spatial working memory in early deafness. Brain 2015, 138, 2750–2765. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Puschmann, S.; Daeglau, M.; Stropahl, M.; Mirkovic, B.; Rosemann, S.; Thiel, C.M.; Debener, S. Hearing-impaired listeners show increased audiovisual benefit when listening to speech in noise. Neuroimage 2019, 196, 261–268. [Google Scholar] [CrossRef]
- Stropahl, M.; Debener, S. Auditory cross-modal reorganization in cochlear implant users indicates audio-visual integration. NeuroImage Clin. 2017, 16, 514–523. [Google Scholar] [CrossRef]
- Stropahl, M.; Plotz, K.; Schonfeld, R.; Lenarz, T.; Sandmann, P.; Yovel, G.; De Vos, M.; Debener, S. Cross-modal reorganization in cochlear implant users: Auditory cortex contributes to visual face processing. Neuroimage 2015, 121, 159–170. [Google Scholar] [CrossRef]
- Glick, H.; Sharma, A. Cross-modal plasticity in developmental and age-related hearing loss: Clinical Implications. Hear. Res. 2017, 343, 191–201. [Google Scholar] [CrossRef]
- Neville, H.J.; Lawson, D. Attention to central and peripheral visual space in a movement detection task: An event-related potential and behavioral study. II. Congenitally deaf adults. Brain Res. 1987, 405, 268–283. [Google Scholar] [CrossRef]
- Buckley, K.A.; Tobey, E.A. Cross-modal plasticity and speech perception in pre- and postlingually deaf cochlear implant users. Ear Hear. 2011, 32, 2–15. [Google Scholar] [CrossRef]
- Chen, L.-C.; Sandmann, P.; Thorne, J.; Bleichner, M.G.; Debener, S. Cross-modal functional reorganization of visual and auditory cortex in adult cochlear implant users identified with fNIRS. Neural Plast. 2016, 2016, 4382656. [Google Scholar] [CrossRef] [Green Version]
- Doucet, M.E.; Bergeron, F.; Lassonde, M.; Ferron, P.; Lepore, F. Cross-modal reorganization and speech perception in cochlear implant users. Brain 2006, 129, 3376–3383. [Google Scholar] [CrossRef] [PubMed]
- Kim, M.-B.; Shim, H.-Y.; Jin, S.H.; Kang, S.; Woo, J.; Han, J.C.; Lee, J.Y.; Kim, M.; Cho, Y.-S.; Moon, J., II; et al. Cross-modal and intra-modal characteristics of visual function and speech perception performance in postlingually deafened, cochlear implant users. PLoS ONE 2016, 11, e0148466. [Google Scholar] [CrossRef] [PubMed]
- Strelnikov, K.; Rouger, J.; Demonet, J.F.; Lagleyre, S.; Fraysse, B.; Deguine, O.; Barone, P. Visual activity predicts auditory recovery from deafness after adult cochlear implantation. Brain 2013, 136, 3682–3695. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vachon, P.; Voss, P.; Lassonde, M.; Leroux, J.-M.; Mensour, B.; Beaudoin, G.; Bourgouin, P.; Lepore, F. Reorganization of the auditory, visual and multimodal areas in early deaf individuals. Neuroscience 2013, 245, 50–60. [Google Scholar] [CrossRef]
- Giraud, A.L.; Lee, H.J. Predicting cochlear implant outcome from brain organisation in the deaf. Restor. Neurol. Neurosci. 2007, 25, 381–390. [Google Scholar]
- Sandmann, P.; Dillier, N.; Eichele, T.; Meyer, M.; Kegel, A.; Pascual-Marqui, R.D.; Marcar, V.L.; Jäncke, L.; Debener, S. Visual activation of auditory cortex reflects maladaptive plasticity in cochlear implant users. Brain 2012, 135, 555–568. [Google Scholar] [CrossRef] [Green Version]
- Mitchell, T.V.; Maslin, M.T. How vision matters for individuals with hearing loss. Int. J. Audiol. 2007, 46, 500–511. [Google Scholar] [CrossRef]
- Glick, H.A.; Sharma, A. Cortical neuroplasticity and cognitive function in early-stage, mild-moderate hearing loss: Evidence of neurocognitive benefit from hearing aid use. Front. Neurosci. 2020, 14, 93. [Google Scholar] [CrossRef] [Green Version]
- Puschmann, S.; Thiel, C.M. Changed Crossmodal functional connectivity in older adults with hearing loss. Cortex 2017, 86, 109–122. [Google Scholar] [CrossRef]
- Sharma, A.; Glick, H. Cross-modal Re-organization in clinical populations with hearing loss. Brain Sci. 2016, 6, 4. [Google Scholar] [CrossRef]
- Rosemann, S.; Thiel, C.M. Audio-visual speech processing in age-related hearing loss: Stronger integration and increased frontal lobe recruitment. Neuroimage 2018, 175, 425–437. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.; Rajagovindan, R.; Han, S.-M.; Ding, M. Top-down control of alpha oscillations: Sources of control signals and their mechanisms of action. Front. Hum. Neurosci. 2016, 10, 15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Woodhead, Z.V.J.; Barnes, G.R.; Penny, W.; Moran, R.; Teki, S.; Price, C.J.; Leff, A.P. Reading front to back: MEG evidence for early feedback effects during word recognition. Cereb. Cortex 2014, 24, 817–825. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gazzaley, A.; Rissman, J.; Cooney, J.; Rutman, A.; Seibert, T.; Clapp, W.; D’Esposito, M. Functional interactions between prefrontal and visual association cortex contribute to top-down modulation of visual processing. Cereb. Cortex 2007, 17, i125–i135. [Google Scholar] [CrossRef]
- Campbell, J.; Sharma, A. Compensatory changes in cortical resource allocation in adults with hearing loss. Front. Syst. Neurosci. 2013, 7, 71. [Google Scholar] [CrossRef] [Green Version]
- Goman, A.M.; Lin, F.R. Prevalence of hearing loss by severity in the United States. Am. J. Public Health 2016, 106, 1820–1822. [Google Scholar] [CrossRef]
- Mann, H.B.; Whitney, D.R. On a test of whether one of two random variables is stochastically larger than the other. Ann. Math. Stat. 1947, 18, 50–60. [Google Scholar] [CrossRef]
- Killion, M.C.; Niquette, P.A.; Gudmundsen, G.I.; Revit, L.J.; Banerjee, S. Development of a quick speech-in-noise test for measuring signal-to-noise ratio loss in normal-hearing and hearing-impaired listeners. J. Acoust. Soc. Am. 2004, 116, 2395–2405. [Google Scholar] [CrossRef]
- Bertrand, J.-A.; Lassonde, M.; Robert, M.; Nguyen, D.K.; Bertone, A.; Doucet, M.-E.; Bouthillier, A.; Lepore, F. An intracranial event-related potential study on transformational apparent motion. Does its neural processing differ from real motion? Exp. Brain Res. 2012, 216, 145–153. [Google Scholar] [CrossRef]
- Campbell, J.; Sharma, A. Visual cross-modal reorganization in children with cochlear implants. PLoS ONE 2016, 11, e0147793. [Google Scholar] [CrossRef] [Green Version]
- Campbell, J.; Sharma, A. Distinct visual evoked potential morphological patterns for apparent motion processing in school-aged children. Front. Hum. Neurosci. 2016, 10, 277. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Doucet, M.E.; Gosselin, F.; Lassonde, M.; Guillemot, J.P.; Lepore, F. Development of visual evoked potentials to radially modulated concentric patterns. Neuroreport 2005, 16, 1753–1756. [Google Scholar] [CrossRef] [PubMed]
- Delorme, A.; Makeig, S. EEGLAB: An open source toolbox for analysis of single-trial EEG dynamics including independent component analysis. J. Neurosci. Methods 2004, 134, 9–21. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Joyce, C.; Rossion, B. The face-sensitive N170 and VPP components manifest the same brain processes: The effect of reference electrode site. Clin. Neurophysiol. 2005, 116, 2613–2631. [Google Scholar] [CrossRef] [PubMed]
- Debener, S.; Ullsperger, M.; Siegel, M.; Engel, A.K. Single-trial EEG–fMRI reveals the dynamics of cognitive function. Trends Cogn. Sci. 2006, 10, 558–563. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Debener, S.; Hine, J.; Bleeck, S.; Eyles, J. Source localization of auditory evoked potentials after cochlear implantation. Psychophys 2008, 45, 20–24. [Google Scholar] [CrossRef]
- Gilley, P.M.; Sharma, A.; Dorman, M.F. Cortical reorganization in children with cochlear implants. Brain Res. 2008, 1239, 56–65. [Google Scholar] [CrossRef] [Green Version]
- Makeig, S.; Jung, T.P.; Bell, A.J.; Ghahremani, D.; Sejnowski, T.J. Blind separation of auditory event-related brain responses into independent components. Proc. Nat. Acad. Sci. USA 1997, 94, 10979–10984. [Google Scholar] [CrossRef] [Green Version]
- Hine, J.; Debener, S. Late auditory evoked potentials asymmetry revisited. Clin. Neurophysiol. 2007, 118, 1274–1285. [Google Scholar] [CrossRef]
- Makeig, S.; Delorme, A.; Westerfield, M.; Jung, T.P.; Townsend, J.; Courchesne, E.; Sejnowski, T.J. Electroencephalographic brain dynamics following manually responded visual targets. PLoS Biol. 2004, 2, e176. [Google Scholar] [CrossRef] [Green Version]
- Grech, R.; Cassar, T.; Muscat, J.; Camilleri, K.P.; Fabri, S.G.; Zervakis, M.; Vanrumste, B. Review on solving the inverse problem in EEG source analysis. J. Neuroeng. Rehabil. 2008, 5, 25. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pascual-Marqui, R.D. Standardized low-resolution brain electromagnetic tomography (sLORETA): Technical details. Methods Find. Exp. Clin. Pharmacol. 2002, 24 (Suppl. D), 5–12. [Google Scholar]
- Fuchs, M.; Kastner, J.; Wagner, M.; Hawes, S.; Ebersole, J.S. A standardized boundary element method volume conductor model. Clin. Neurophysiol. 2002, 113, 702–712. [Google Scholar] [CrossRef] [Green Version]
- Benjamini, Y.; Hochberg, Y. Controlling the false discovery rate: A practical and powerful approach to multiple testing. J. R. Stat. Soc. Ser. B 1995, 57, 289–300. [Google Scholar] [CrossRef]
- Ding, H.; Ming, D.; Wan, B.; Li, Q.; Qin, W.; Yu, C. Enhanced spontaneous functional connectivity of the superior temporal gyrus in early deafness. Sci. Rep. 2016, 6, 23239. [Google Scholar] [CrossRef] [PubMed]
- Ahmadi, M.; McDevitt, E.A.; Silver, M.A.; Mednick, S.C. Perceptual learning induces changes in early and late visual evoked potentials. Vis. Res. 2018, 152, 101–109. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gonzalez, G.F.; Zaric, G.; Tijms, J.; Bonte, M.; Blomert, L.; Leppanen, P.; van der Molen, M.W. Responsitivity to dyslexia training indexed by the N170 amplitude of the brain potential elicited by word reading. Brain Cogn. 2016, 106, 42–54. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Eisner, F.; McGettigan, C.; Faulkner, A.; Rosen, S.; Scott, S.K. Inferior frontal gyrus activation predicts individual differences in perceptual learning of cochlear-implant simulations. J. Neurosci. 2010, 30, 7179–7186. [Google Scholar] [CrossRef] [Green Version]
- Peelle, J.E. Listening effort: How the cognitive consequences of acoustic challenge are reflected in brain and behavior. Ear Hear. 2018, 39, 204–214. [Google Scholar] [CrossRef]
- Buckner, R.L.; Andrews-Hanna, J.R.; Schacter, D.L. The brain’s default network: Anatomy, function, and relevance to disease. Ann. N.Y. Acad. Sci. 2008, 1124, 1–38. [Google Scholar] [CrossRef] [Green Version]
- Frey, S.; Petrides, M. Orbitofrontal cortex: A key prefrontal region for encoding information. Proc. Natl. Acad. Sci. USA 2000, 97, 8723–8727. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Savage, C.R.; Deckersbach, T.; Heckers, S.; Wagner, A.D.; Schacter, D.L.; Alpert, N.M.; Fischman, A.J.; Rauch, S.L. Prefrontal regions supporting spontaneous and directed application of verbal learning strategies: Evidence from PET. Brain 2001, 124, 219–231. [Google Scholar] [CrossRef] [PubMed]
- Uddin, L.Q.; Kelly, A.M.; Biswal, B.B.; Castellanos, F.X.; Milham, M.P. Functional connectivity of default mode network components: Correlation, anticorrelation, and causality. Hum. Brain Mapp. 2009, 30, 625–637. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bidelman, G.M.; Mahmud, M.S.; Yeasin, M.; Shen, D.; Arnott, S.R.; Alain, C. Age-related hearing loss increases full-brain connectivity while reversing directed signaling within the dorsal-ventral pathway for speech. Brain Struct. Funct. 2019, 224, 2661–2676. [Google Scholar] [CrossRef]
- Price, C.N.; Alain, C.; Bidelman, G.M. Auditory-frontal channeling in α and β bands is altered by age-related hearing loss and relates to speech perception in noise. Neuroscience 2019, 423, 18–28. [Google Scholar] [CrossRef]
- Cardin, V. Effects of aging and adult-onset hearing loss on cortical auditory regions. Front. Neurosci. 2016, 10, 199. [Google Scholar] [CrossRef] [Green Version]
- Peelle, J.E.; Wingfield, A. The neural consequences of age-related hearing loss. Trends Neurosci. 2016, 39, 486–497. [Google Scholar] [CrossRef] [Green Version]
- Gates, G.A.; Anderson, M.L.; McCurry, S.M.; Feeney, M.P.; Larson, E.B. Central auditory dysfunction as a harbinger of Alzheimer dementia. Arch. Otolaryngol. Head Neck Surg. 2011, 137, 390–395. [Google Scholar] [CrossRef] [Green Version]
- Humes, L.E.; Kidd, G.R.; Lentz, J.J. Auditory and cognitive factors underlying individual differences in aided speech-understanding among older adults. Front. Syst. Neurosci. 2013, 7, 55. [Google Scholar] [CrossRef] [Green Version]
- Lin, F.R.; Yaffe, K.; Xia, J.; Xue, Q.-L.; Harris, T.B.; Purchase-Helzner, E.; Satterfield, S.; Ayonayon, H.N.; Ferrucci, L.; Simonsick, E.M. Hearing loss and cognitive decline among older adults. JAMA Intern. Med. 2013, 173, 293–299. [Google Scholar] [CrossRef]
- Harkrider, A.W.; Plyler, P.N.; Hedrick, M.S. Effects of hearing loss and spectral shaping on identification and neural response patterns of stop-consonant stimuli in young adults. Ear Hear. 2009, 30, 31–42. [Google Scholar] [CrossRef] [PubMed]
- Bertoli, S.; Probst, R.; Bodmer, D. Late auditory evoked potentials in elderly long-term hearing-aid users with unilateral or bilateral fittings. Hear. Res. 2011, 280, 58–69. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Peelle, J.E.; Troiani, V.; Grossman, M.; Wingfield, A. Hearing loss in older adults affects neural systems supporting speech comprehension. J. Neurosci. 2011, 31, 12638–12643. [Google Scholar] [CrossRef] [PubMed]
- Wingfield, A.; Peelle, J.E. The effects of hearing loss on neural processing and plasticity. Front. Syst. Neurosci. 2015, 9, 35. [Google Scholar] [CrossRef] [PubMed] [Green Version]






© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Campbell, J.; Sharma, A. Frontal Cortical Modulation of Temporal Visual Cross-Modal Re-organization in Adults with Hearing Loss. Brain Sci. 2020, 10, 498. https://doi.org/10.3390/brainsci10080498
Campbell J, Sharma A. Frontal Cortical Modulation of Temporal Visual Cross-Modal Re-organization in Adults with Hearing Loss. Brain Sciences. 2020; 10(8):498. https://doi.org/10.3390/brainsci10080498
Chicago/Turabian StyleCampbell, Julia, and Anu Sharma. 2020. "Frontal Cortical Modulation of Temporal Visual Cross-Modal Re-organization in Adults with Hearing Loss" Brain Sciences 10, no. 8: 498. https://doi.org/10.3390/brainsci10080498
APA StyleCampbell, J., & Sharma, A. (2020). Frontal Cortical Modulation of Temporal Visual Cross-Modal Re-organization in Adults with Hearing Loss. Brain Sciences, 10(8), 498. https://doi.org/10.3390/brainsci10080498