Effect of Cladribine on Neuronal Apoptosis: New Insight of In Vitro Study in Multiple Sclerosis Therapy


 ,
, 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Design and Patients
2.2. Peripheral Blood Mononuclear Cell Purification
2.3. SH-SY5Y Cell Culture
2.4. Cladribine Treatment
2.5. Western Blot Analysis
2.6. Flow Cytometry on SH-SY5Y
2.7. Statistical Analysis
3. Results
3.1. Cladribine Effects on Lymphocyte Cell Cultures and on Lymphocyte and SH-SY5Y Co-Cultures
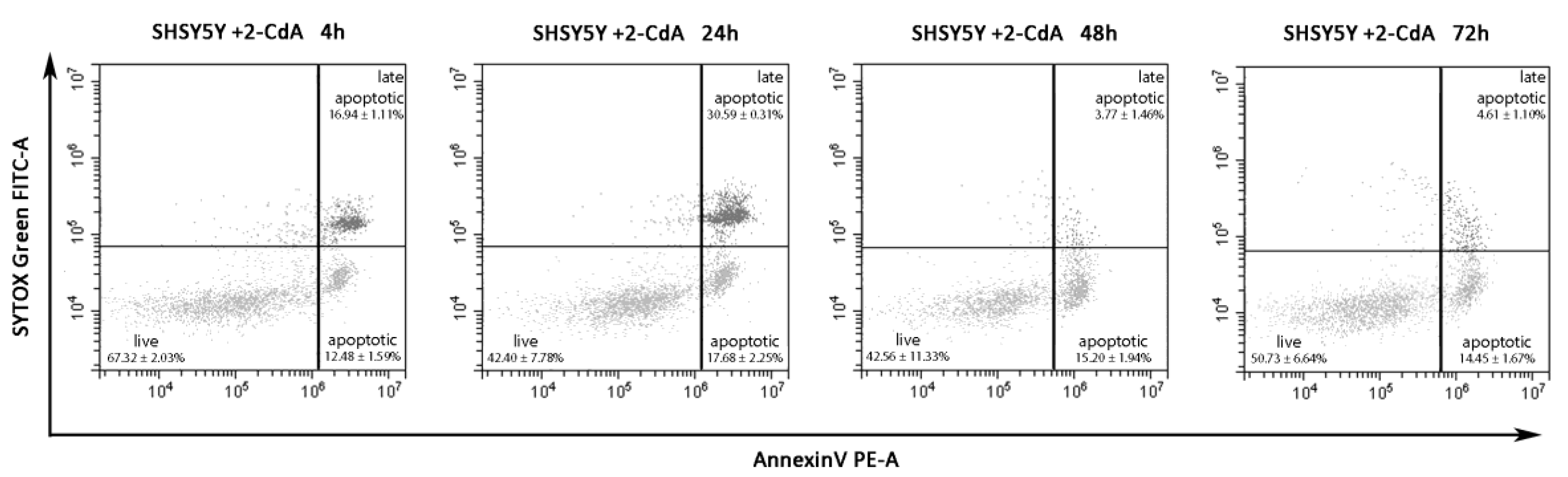
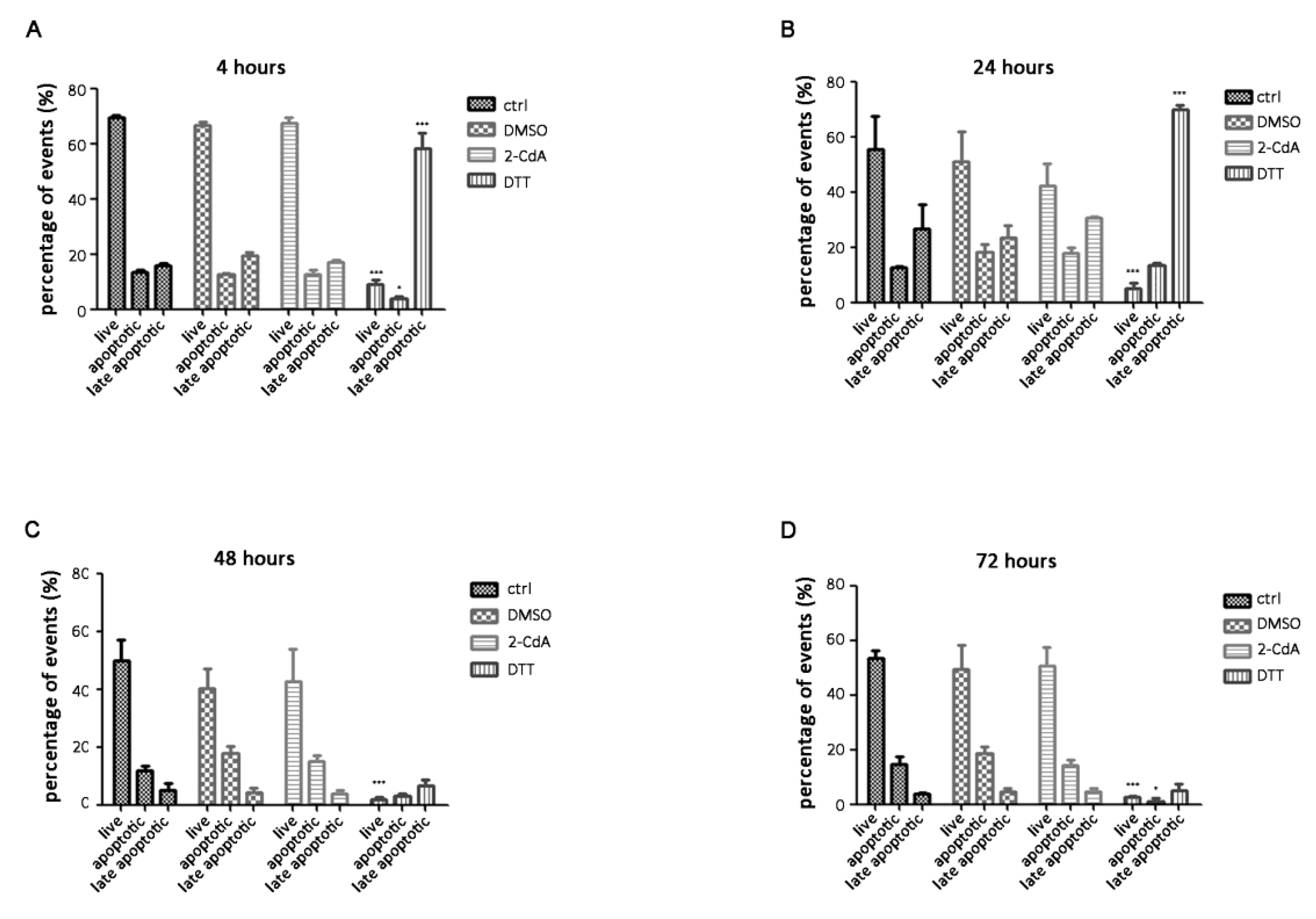
3.2. Annexin V on SH-SY5Y Treated with 2-CdA
4. Discussion and Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Dobson, R.; Giovannoni, G. Multiple sclerosis—A review. Eur. J. Neurol. 2019, 26, 27–40. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chitnis, T. The role of CD4 T cells in the pathogenesis of multiple sclerosis. Int. Rev. Neurobiol. 2007, 79, 43–72. [Google Scholar] [PubMed]
- Raine, C.S. The Dale E. McFarlin Memorial Lecture: The immunology of the multiple sclerosis lesion. Ann. Neurol. 1994, 36, S61–S72. [Google Scholar] [CrossRef] [PubMed]
- Cassan, C.; Liblau, R.S. Immune tolerance and control of CNS autoimmunity: From animal models to MS patients. J. Neurochem. 2007, 100, 883–892. [Google Scholar] [CrossRef] [PubMed]
- Sospedra, M.; Martin, R. Immunology of multiple sclerosis. Annu. Rev. Immunol. 2005, 23, 683–747. [Google Scholar] [CrossRef] [Green Version]
- Chitnis, T.; Najafian, N.; Benou, C.; Salama, A.D.; Grusby, M.J.; Sayegh, M.H.; Khoury, S.J. EVect of targeted disruption of STAT4 and STAT6 on the induction of experimental autoimmune en ephalomyelitis. J. Clin. Investig. 2001, 108, 739–774. [Google Scholar] [CrossRef]
- Häusser-Kinzel, S.; Weber, M.S. The Role of B Cells and Antibodies in Multiple Sclerosis, Neuromyelitis Optica, and Related Disorders. Front. Immunol. 2019, 10, 201. [Google Scholar] [CrossRef]
- Harp, C.T.; Lovett-Racke, A.E.; Racke, M.K.; Frohman, E.M.; Monson, N.L. Impact of myelin-specific antigen presenting B cells on T cell activation in multiple sclerosis. Clin. Immunol. 2008, 128, 382–391. [Google Scholar] [CrossRef]
- Molnarfi, N.; Schulzetopphoff, U.; Weber, M.S.; Patarroyo, J.C.; Prodhomme, T.; Varrindoyer, M.; Shetty, A.; Linington, C.; Slavin, A.J.; Hidalgo, J.; et al. MHC class II-dependent B cell APC function is required for induction of CNS autoimmunity independent of myelin-specific antibodies. J. Exp. Med. 2013, 210, 2921–2937. [Google Scholar] [CrossRef]
- Szczucinski, A.; Losy, J. Chemokines and chemokine receptors in multiple sclerosis. Potential targets for new therapies. Acta Neurol. Scand. 2007, 115, 137–146. [Google Scholar] [CrossRef]
- Ortiz, G.G.; Pacheco-Moisés, F.P.; Macıas-Islas, M.A.; Flores-Alvarado, L.J.; Mireles-Ramırez, M.A.; Gonzàlez-Renovato, E.D.; Hernandez-Navarro, V.E.; Sanchez-Lopez, A.L.; Alatorre-Jiménez, M.A. Role of the Blood-Brain Barrier in Multiple Sclerosis. Arch. Med. Res. 2014, 45, 687–697. [Google Scholar] [CrossRef] [PubMed]
- Lovato, L.; Willis, S.N.; Rodig, S.J.; Caron, T.; Almendinger, S.E.; Howell, O.W.; Reynolds, R.; O’Connor, K.C.; Hafler, D.A. Related B cell clones populate the meninges and parenchyma of patients with multiple sclerosis. Brain 2011, 134, 534–541. [Google Scholar] [CrossRef] [PubMed]
- Milo, R. Therapeutic strategies targeting B-cells in Multiple Sclerosis. Autoimmun. Rev. 2016, 15, 714–718. [Google Scholar] [CrossRef] [PubMed]
- Bruck, W.; Gold, R.; Lund, B.T.; Oreja-Guevara, C.; Prat, A.; Spencer, C.M.; Steinman, L.; Tintoré, M.; Vollmer, T.L.; Weber, M.S.; et al. Therapeutic decisions in multiple sclerosis: Moving beyond efficacy. JAMA Neurol. 2013, 70, 1315–1324. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Giovannoni, G.; Comi, G.; Cook, S.; Rammohan, K.; Rieckmann, P.; Sorensen, P.S.; Vermersch, P.; Chang, P.; Hamlett, A.; Musch, B.; et al. A placebo-controlled trial of oral cladribine for relapsing multiple sclerosis. N. Engl. J. Med. 2010, 362, 416–426. [Google Scholar] [CrossRef] [Green Version]
- Goodman, G.R.; Beutler, E.; Saven, A. Cladribine in the treatment of hairy-cell leukaemia. Best Pract. Res. Clin. Haematol. 2003, 16, 101–116. [Google Scholar] [CrossRef]
- Musella, A.; Mandolesi, G.; Gentile, A.; Rossi, S.; Studer, V.; Motta, C.; Sepman, H.; Fresegna, D.; Haji, N.; Paolillo, A.; et al. Cladribine interferes with IL-1β synaptic effects in experimental multiple sclerosis. J. Neuroimmunol. 2013, 264, 8–13. [Google Scholar] [CrossRef]
- Baker, D.; Pryce, G.; Herrod, S.S.; Schmierer, K. Potential mechanisms of action related to the efficacy and safety of cladribine. Mult. Scler. Relat. Disord. 2019, 30, 176–186. [Google Scholar] [CrossRef]
- Joseph, S.B.; Arrildt, K.T.; Sturdevant, C.B.; Swanstrom, R. HIV-1 target cells in the CNS. J. Neuro 2015, 21, 276–289. [Google Scholar] [CrossRef]
- Smolders, J.; Remmerswaal, E.B.; Schuurman, K.G.; Melief, J.; van Eden, C.G.; van Lier, R.A.; Huitinga, I.; Hamann, J. Characteristics of differentiated CD8(+) and CD4 (+) T cells present in the human brain. Acta Neuropathol. 2013, 126, 525–535. [Google Scholar] [CrossRef]
- Ho, E.L.; Ronquillo, R.; Altmeppen, H.; Spudich, S.S.; Price, R.W.; Sinclair, E. Cellular composition of cerebrospinal fluid in HIV-1 infected and uninfected subjects. PLoS ONE 2013, 8, e66188. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liliemark, J. The clinical pharmacokinetics of cladribine. Clin. Pharmacokinet. 1997, 32, 120–131. [Google Scholar] [CrossRef] [PubMed]
- Hermann, R.; Karlsson, M.O.; Novakovic, A.M.; Terranova, N.; Fluck, M.; Munafo, A. The clinical pharmacology of cladribine tablets for the treatment of relapsing multiple sclerosis. Clin. Pharmacokinet. 2019, 58, 283–297. [Google Scholar] [CrossRef] [Green Version]
- Leist, T.P.; Weissert, R. Cladribine: Mode of action and implications for treatment of multiple sclerosis. Clin. Neuropharmacol. 2011, 34, 28–35. [Google Scholar] [CrossRef] [PubMed]
- Rogalińska, M.; Błoński, J.Z.; Hanausek, M.; Walaszek, Z.; Robak, T.; Kiliańska, Z.M. 2-Chlorodeoxyadenosine alone and in combination with cyclophosphamide and mitoxantrone induce apoptosis in B chronic lymphocytic leukemia cells in vivo. Cancer Detect. Prev. 2004, 28, 433–442. [Google Scholar] [CrossRef] [PubMed]
- Inwards, D.J.; Fishkin, P.A.S.; Hillman, D.W.; Brown, D.W.; Ansell, S.M.; Kurtin, P.J.; Fonseca, R.; Morton, R.F.; Veeder, M.H.; Witzig, T.E. Long-term results of the treatment of patients with mantle cell lymphoma with cladribine (2-CDA) alone (95-80-53) or 2-CDA and rituximab (N0189) in the North Central Cancer Treatment Group. Cancer 2008, 113, 108–116. [Google Scholar] [CrossRef] [Green Version]
- Tobinai, K.; Watanabe, T.; Tanimoto, K.; Maruyama, D.; Nakata, M.; Itoh, K.; Morishima, Y.; Ogura, M.; Usui, N.; Kasai, M.; et al. Phase I/II and pharmacokinetic study of cladribine with 2-h infusion in Japanese patients with relapsed indolent B-cell lymphoma mostly pretreated with rituximab. Cancer Sci. 2009, 100, 1344–1350. [Google Scholar] [CrossRef]
- Inaba, H.; Stewart, C.F.; Crews, K.R.; Yang, S.; Pounds, S.; Pui, C.H.; Rubnitz, J.E.; Razzouk, B.I.; Ribeiro, R.C. Combination of cladribine plus topotecan for recurrent or refractory pediatric acute myeloid leukemia. Cancer 2010, 116, 98–105. [Google Scholar] [CrossRef]
- Oguri, T.; Ozasa, H.; Uemura, T.; Takakuwa, O.; Kunii, E.; Kasai, D.; Ohkubo, H.; Miyazaki, M.; Maeno, K.; Sato, S. Preclinical rationale for synergistic interaction of pemetrexed and cytotoxic nucleoside analogues. Oncol. Lett. 2012, 4, 571–575. [Google Scholar] [CrossRef] [Green Version]
- Robak, T.; Robak, P. Purine nucleoside analogs in the treatment of rarer chronic lymphoid leukemias. Curr. Pharm. Des. 2012, 18, 3373–3388. [Google Scholar] [CrossRef]
- Wang, B.; Lyu, H.; Pei, S.; Song, D.; Ni, J.; Liu, B. Cladribine in combination with entinostat synergistically elicits anti-proliferative/anti-survival effects on multiple myeloma cells. Cell Cycle 2018, 17, 985–996. [Google Scholar] [CrossRef] [PubMed]
- Kopadze, T.; Döbert, M.; Leussink, V.I.; Dehmel, T.; Kieseier, B.C. Caldribine impedes in vitro migration of mononuclear cells: A possible implication for treating multilple sclerosis. Eur. J. Neurol. 2009, 16, 409–412. [Google Scholar] [CrossRef] [PubMed]
- Jørgensen, L.Ø.; Hyrlov, K.H.; Elkjaer, M.L.; Weber, A.B.; Pedersen, A.E.; Fex Svenningsen, A.; Illes, Z. Cladribine modifies functional properties of microglia. Clin. Exp. Immunol. 2020. [Google Scholar] [CrossRef] [PubMed]
- European Medicines Agency. European Public Assessment Report (EPAR) of 2017 (EMA/405631/2017); European Medicines Agency: Amsterdam, The Nederland, 2017. [Google Scholar]
- Lincz, L.F. Deciphering the apoptotic pathway: All roads lead to death. Immunol. Cell Biol. 1998, 76, 1–19. [Google Scholar] [CrossRef]
- Koopman, G.; Reutelingsperger, C.P.; Kuijten, G.A.; Keehnen, R.M.; Pals, S.T.; van Oers, M.H. Annexin V for flow cytometric detection of phosphatidylserine expression on B cells undergoing apoptosis. Blood 1994, 84, 1415–1420. [Google Scholar] [CrossRef] [Green Version]
- Elmore, S. Apoptosis: A review of programmed cell death. Toxicol. Pathol. 2007, 35, 495–516. [Google Scholar] [CrossRef]
- Lawrimore, C.J.; Coleman, L.G.; Crews, F.T. Ethanol induces interferon expression in neurons via TRAIL: Role of astrocyte-to-neuron signalling. Psychopharmacology 2019, 236, 2881–2897. [Google Scholar] [CrossRef] [Green Version]
- Lawrimore, C.J.; Crews, F.T. Ethanol, TLR3, and TLR4 agonists have unique innate immune responses in neuron-like SH-SY5Y and microglia-like BV2. Alcohol. Clin. Exp. Res. 2017, 41, 939–954. [Google Scholar] [CrossRef] [Green Version]
- Lehmann, S.M.; Rosenberger, K.; Kruger, C.; Habbel, P.; Derkow, K.; Kaul, D.; Rybak, A.; Brandt, C.; Schott, E.; Wulczyn, F.G.; et al. Extracellularly delivered single-stranded viral RNA causes neurodegeneration dependent on TLR7. J. Immunol. 2012, 189, 1448–1458. [Google Scholar] [CrossRef] [Green Version]
- Tang, S.C.; Arumugam, T.V.; Xu, X.; Cheng, A.; Mughal, M.R.; Jo, D.G.; Lathia, J.D.; Siler, D.A.; Chigurupati, S.; Ouyang, X.; et al. Pivotal role for neuronal toll-like receptors in ischemic brain injury and functional deficits. Proc. Natl. Acad. Sci. USA 2007, 104, 13798–13803. [Google Scholar] [CrossRef] [Green Version]
- Junn, E.; Mouradian, M.M. Apoptotic signaling in dopamine-induced cell death: The role of oxidative stress, p38 mitogen-activated protein kinase, cytochrome c and caspases. J. Neurochem. 2001, 78, 374–383. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ouyang, M.; Shen, X. Critical role of ASK1 in the 6-hydroxydopamine-induced apoptosis in human neuroblastoma SH-SY5Y cells. J. Neurochem. 2006, 97, 234–244. [Google Scholar] [CrossRef] [PubMed]
- Blum, D.; Torch, S.; Lambeng, N.; Nissou, M.F.; Benabid, A.L.; Sadoul, R.; Verna, J.M. Molecular pathways involved in the neurotoxicity of 6-OHDA, dopamine and MPTP: Contribution to the apoptotic theory in Parkinson’s disease. Prog. Neurobiol. 2001, 65, 135–172. [Google Scholar] [CrossRef]
- Schober, A. Classic toxin-induced animal models of Parkinson’s disease: 6-OHDA and MPTP. Cell Tissue Res. 2004, 318, 215–224. [Google Scholar] [CrossRef]
- Tiffany-Castiglioni, E.; Perez-Polo, J.R. Stimulation of resistance to 6-hydroxydopamine in a human neuroblastoma cell line by nerve growth factor. Neurosci. Lett. 1981, 26, 157–161. [Google Scholar] [CrossRef]
- Ikeda, Y.; Tsuji, S.; Satoh, A.; Ishikura, M.; Shirasawa, T.; Shimizu, T. Protective effects of astaxanthin on 6-hydroxydopamine-induced apoptosis in human neuroblastoma SH-SY5Y cells. J. Neurochem. 2008, 107, 1730–1740. [Google Scholar] [CrossRef] [PubMed]
- Bradl, M.; Lassmann, H. Progressive multiple sclerosis. Semin. Immunopathol. 2009, 31, 455–465. [Google Scholar] [CrossRef]





© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ruggieri, M.; Gargano, C.D.; Ferretta, A.; Manni, A.; Capacchione, A.; Frigeri, A.; Iaffaldano, P.; Trojano, M.; Paolicelli, D. Effect of Cladribine on Neuronal Apoptosis: New Insight of In Vitro Study in Multiple Sclerosis Therapy. Brain Sci. 2020, 10, 548. https://doi.org/10.3390/brainsci10080548
Ruggieri M, Gargano CD, Ferretta A, Manni A, Capacchione A, Frigeri A, Iaffaldano P, Trojano M, Paolicelli D. Effect of Cladribine on Neuronal Apoptosis: New Insight of In Vitro Study in Multiple Sclerosis Therapy. Brain Sciences. 2020; 10(8):548. https://doi.org/10.3390/brainsci10080548
Chicago/Turabian StyleRuggieri, Maddalena, Concetta Domenica Gargano, Anna Ferretta, Alessia Manni, Antonio Capacchione, Antonio Frigeri, Pietro Iaffaldano, Maria Trojano, and Damiano Paolicelli. 2020. "Effect of Cladribine on Neuronal Apoptosis: New Insight of In Vitro Study in Multiple Sclerosis Therapy" Brain Sciences 10, no. 8: 548. https://doi.org/10.3390/brainsci10080548
APA StyleRuggieri, M., Gargano, C. D., Ferretta, A., Manni, A., Capacchione, A., Frigeri, A., Iaffaldano, P., Trojano, M., & Paolicelli, D. (2020). Effect of Cladribine on Neuronal Apoptosis: New Insight of In Vitro Study in Multiple Sclerosis Therapy. Brain Sciences, 10(8), 548. https://doi.org/10.3390/brainsci10080548