
Effects of Increasing Stimulated Area in Spatiotemporally Congruent Unisensory and Multisensory Conditions


Abstract
:1. Introduction
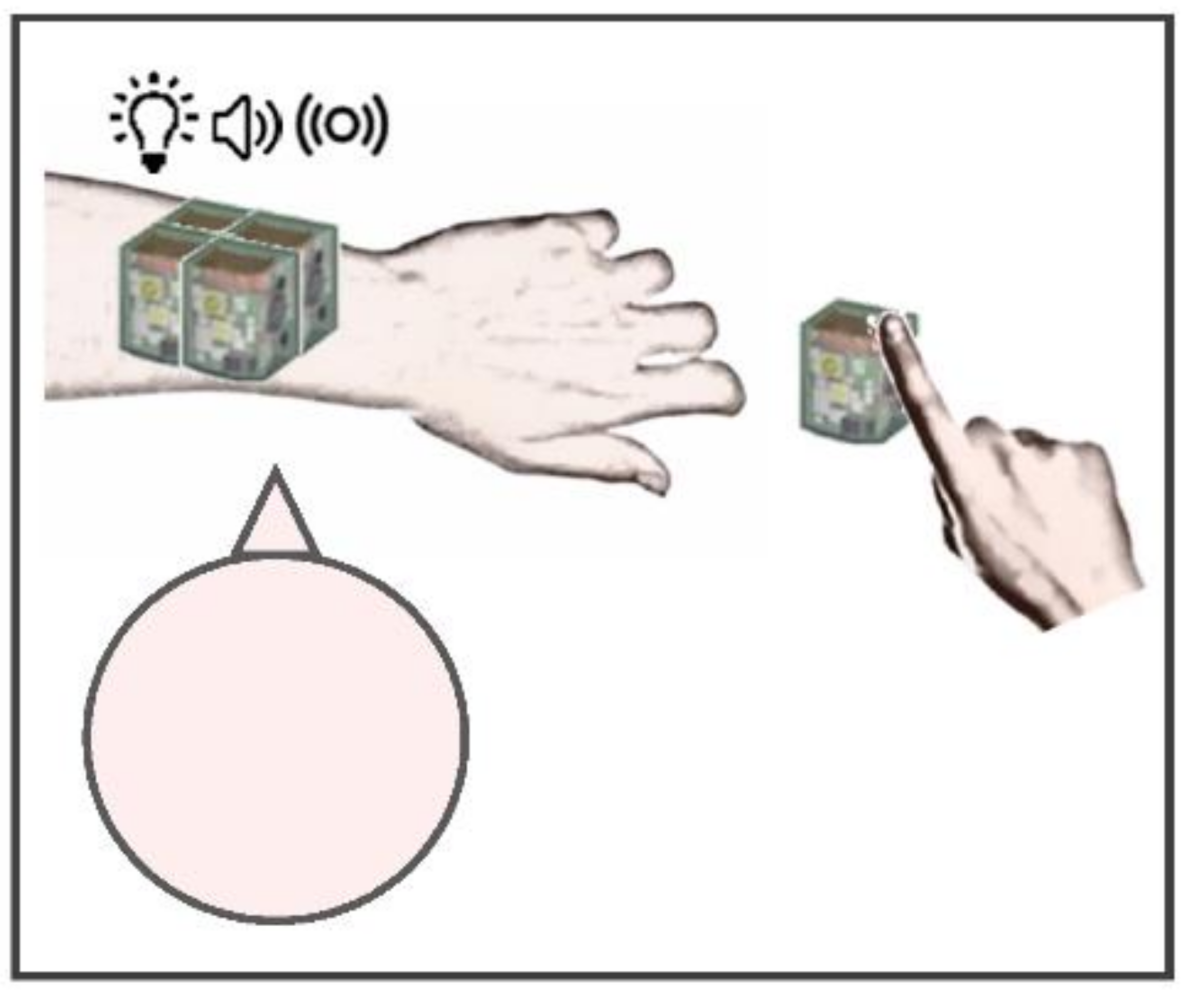
2. Materials and Methods
2.1. Participants
- -
- effect size dz: 1.18 (Cohen’s d = 1.09; see Schiatti et al., 2020 [48]);
- -
- α err. prob. = 0.05;
- -
- power (1 − β err. prob.) = 0.95.
2.2. Experimental Setup and Protocol
2.3. Data Analysis and Statistics
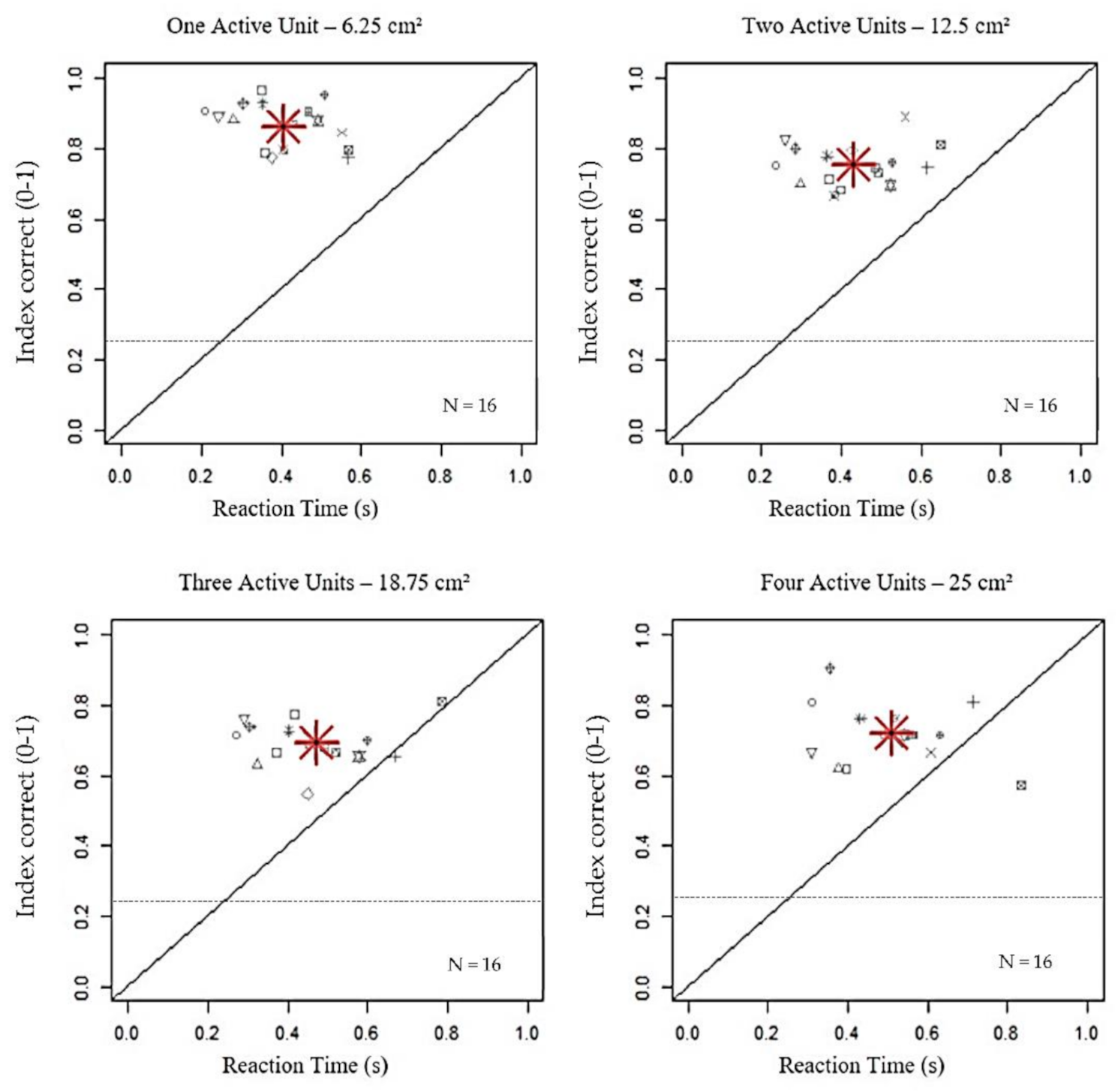
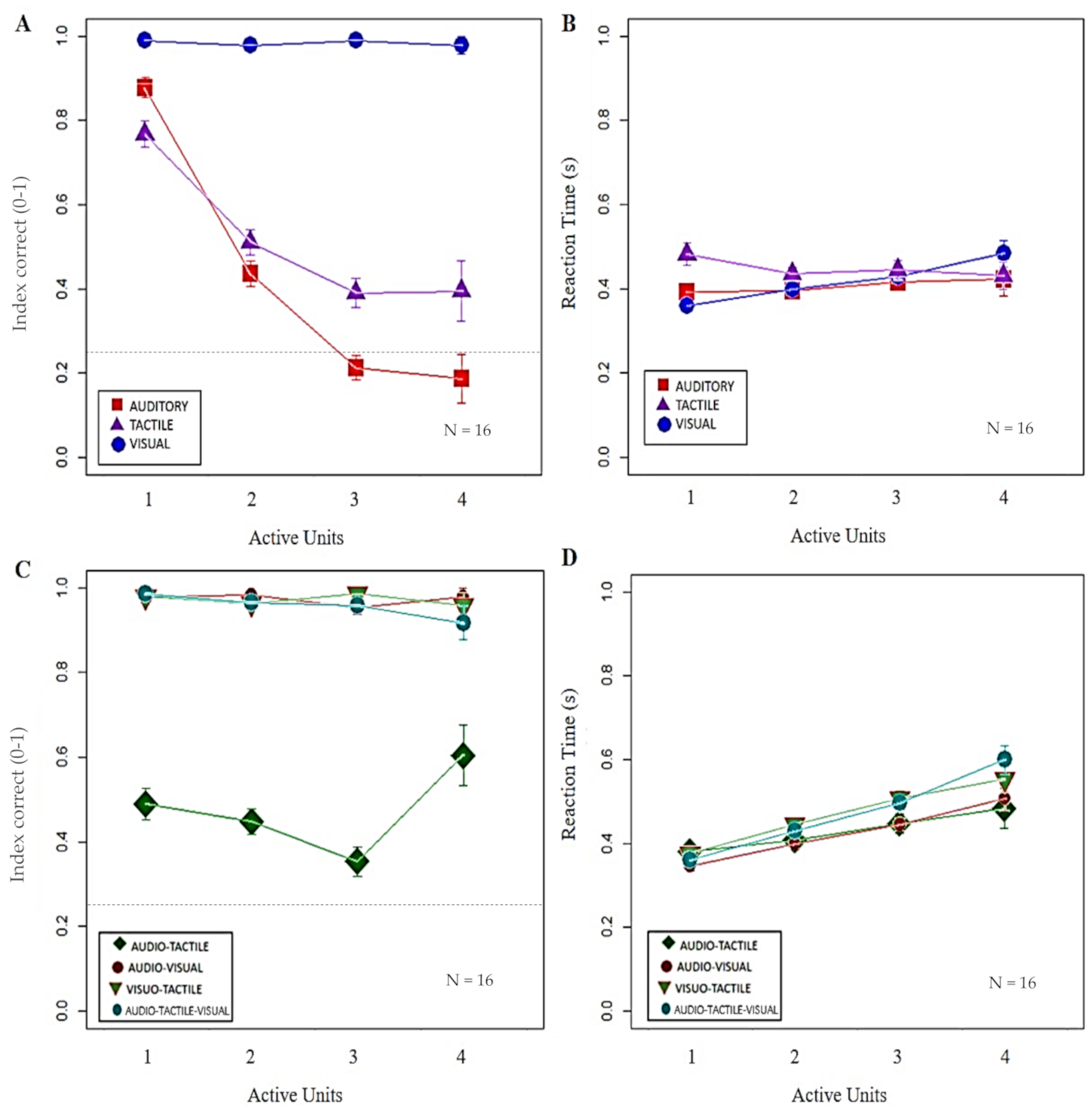
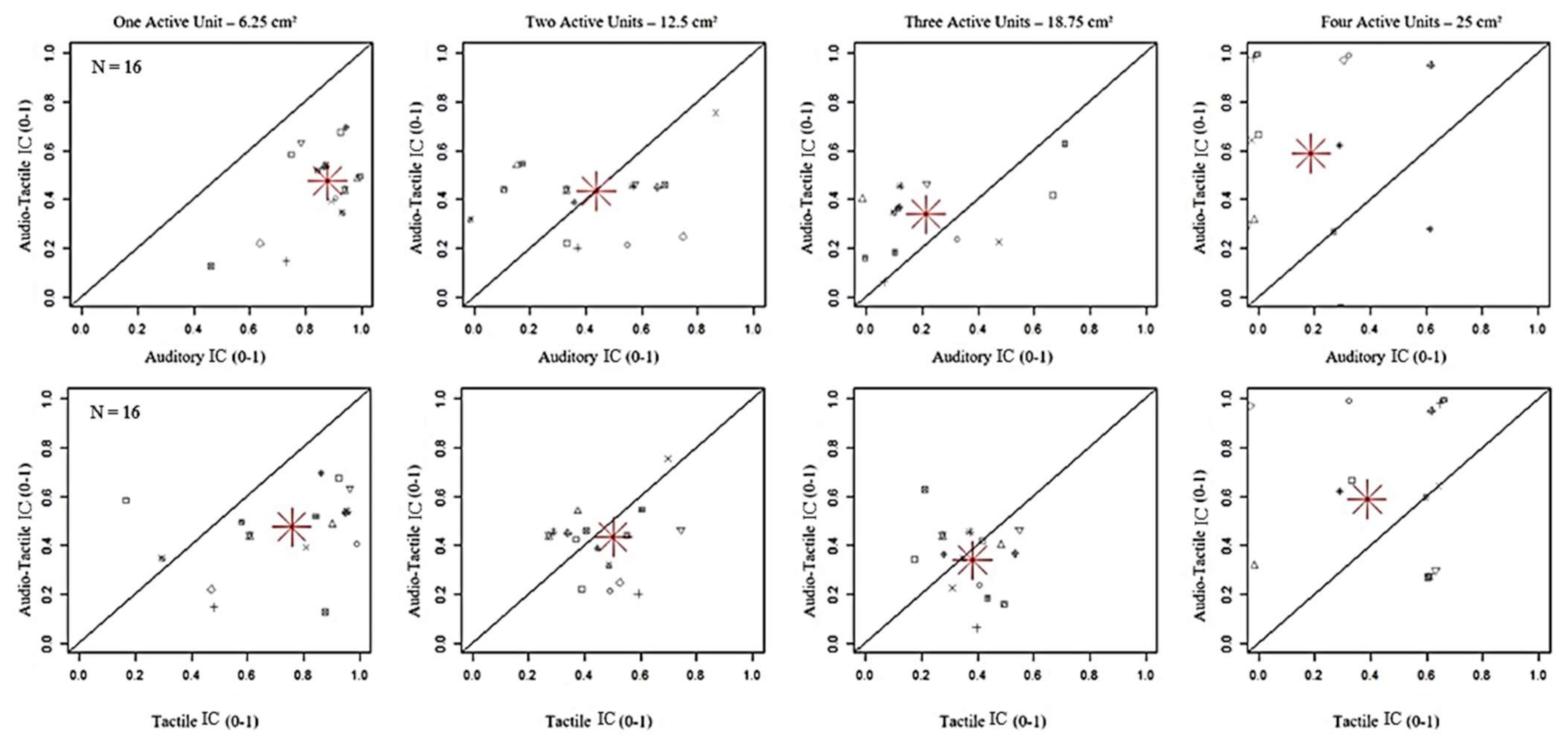
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Hart, R.A.; Moore, R.A. The development of spatial cognition: A review. In Image & Environment: Cognitive Mapping and Spatial Behavior; Aldine Transaction: London, UK, 1973. [Google Scholar]
- Vasilyeva, M.; Lourenco, S.F. Spatial development. In The Handbook of Life-Span Development; John Wiley & Sons, Inc.: Hoboken, NJ, USA, 2010; Volume 1. [Google Scholar]
- Thinus-Blanc, C.; Gaunet, F. Representation of space in blind persons: Vision as a spatial sense? Psychol. Bull. 1997, 121, 20–42. [Google Scholar] [CrossRef]
- Pasqualotto, A.; Proulx, M.J. The role of visual experience for the neural basis of spatial cognition. Neurosci. Biobehav. Rev. 2012, 36, 1179–1187. [Google Scholar] [CrossRef] [PubMed]
- Cappagli, G.; Cocchi, E.; Gori, M. Auditory and proprioceptive spatial impairments in blind children and adults. Dev. Sci. 2015, 20. [Google Scholar] [CrossRef] [PubMed]
- Newell, F.N.; Woods, A.T.; Mernagh, M.; Bülthoff, H.H. Visual, haptic and crossmodal recognition of scenes. Exp. Brain Res. 2005, 161, 233–242. [Google Scholar] [CrossRef]
- Newport, R.; Rabb, B.; Jackson, S.R. Noninformative vision improves haptic spatial perception. Curr. Biol. 2002, 12, 1661–1664. [Google Scholar] [CrossRef] [Green Version]
- Postma, A.; Zuidhoek, S.; Noordzij, M.L.; Kappers, A.M.L. Differences between early-blind, late-blind, and blindfolded-sighted people in haptic spatial-configuration learning and resulting memory traces. Perception 2007, 36, 1253–1265. [Google Scholar] [CrossRef] [Green Version]
- Ungar, S.; Blades, M.; Spencer, C. Mental rotation of a tactile layout by young visually impaired children. Perception 1995, 24, 891–900. [Google Scholar] [CrossRef]
- Bigelow, A.E. Blind and sighted children’s spatial knowledge of their home environments. Int. J. Behav. Dev. 1996, 19, 797–816. [Google Scholar] [CrossRef]
- Cattaneo, Z.; Vecchi, T.; Cornoldi, C.; Mammarella, I.; Bonino, D.; Ricciardi, E.; Pietrini, P. Imagery and spatial processes in blindness and visual impairment. Neurosci. Biobehav. Rev. 2008, 32, 1346–1360. [Google Scholar] [CrossRef]
- Koustriava, E.; Papadopoulos, K. Mental rotation ability of individuals with visual impairments. J. Vis. Impair. Blind. 2010, 104, 570–575. [Google Scholar] [CrossRef]
- Maurer, D.; Lewis, T.L.; Mondloch, C.J. Missing sights: Consequences for visual cognitive development. Trends Cogn. Sci. 2005, 9, 144–151. [Google Scholar] [CrossRef]
- Lepore, N.; Shi, Y.; Lepore, F.; Fortin, M.; Voss, P.; Chou, Y.; Lord, C.; Lassonde, M.; Dinov, I.D.; Toga, A.W. Pattern of hippocampal shape and volume differences in blind subjects. Neuroimage 2009, 46, 949–957. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ruotolo, F.; Ruggiero, G.; Vinciguerra, M.; Iachini, T. Sequential vs simultaneous encoding of spatial information: A comparison between the blind and the sighted. Acta Psychol. 2012, 139, 382–389. [Google Scholar] [CrossRef]
- Gori, M. Multisensory Integration and Calibration in Children and Adults with and without Sensory and Motor Disabilities. Multisens. Res. 2015, 28, 71–99. [Google Scholar] [CrossRef] [PubMed]
- Stein, B.E.; Meredith, M.A. The Merging of the Senses. MIT Press 1993. [Google Scholar]
- Miller, J. Divided attention: Evidence for coactivation with redundant signals. Cogn. Psychol. 1982, 14, 247–279. [Google Scholar] [CrossRef]
- Diederich, A.; Colonius, H.; Bockhorst, D.; Tabeling, S. Visual-tactile spatial interaction in saccade generation. Exp. Brain Res. 2003, 148, 328–337. [Google Scholar] [CrossRef]
- Murray, M.M.; Molholm, S.; Michel, C.M.; Heslenfeld, D.J.; Ritter, W.; Javitt, D.C.; Schroeder, C.E.; Foxe, J.J. Grabbing your ear: Rapid auditory–somatosensory multisensory interactions in low-level sensory cortices are not constrained by stimulus alignment. Cereb. Cortex 2004, 15, 963–974. [Google Scholar] [CrossRef]
- Todd, J.W. Reaction to Multiple Stimuli; Science Press: Beijing, China, 1912. [Google Scholar]
- Alais, D.; Burr, D. The ventriloquist effect results from Near-Optimal Bimodal Integration. Curr. Biol. 2004, 14, 257–262. [Google Scholar] [CrossRef]
- Anderson, P.W.; Zahorik, P. Auditory and visual distance estimation. In Proceedings of the Meetings on Acoustics, Seattle, WA, USA, 23–27 May 2011; Volume 12. [Google Scholar]
- Bertelson, P.; Aschersleben, G. Temporal ventriloquism: Crossmodal interaction on the time dimension: Evidence from auditory–visual temporal order judgment. Int. J. Psychophysiol. 2003, 50, 147–155. [Google Scholar] [CrossRef]
- Botvinick, M.; Cohen, J. Rubber hands ‘feel’ touch that eyes see. Nature 1998, 391, 756. [Google Scholar] [CrossRef] [PubMed]
- Flanagan, J.R.; Beltzner, M.A. Independence of perceptual and sensorimotor predictions in the size–weight illusion. Nat. Neurosci. 2000, 3, 737–741. [Google Scholar] [CrossRef]
- Zahorik, P. Estimating sound source distance with and without vision. Optom. Vis. Sci. 2001, 78, 270–275. [Google Scholar] [CrossRef] [PubMed]
- Stevenson, R.A.; Fister, J.K.; Barnett, Z.P.; Nidiffer, A.R.; Wallace, M.T. Interactions between the spatial and temporal stimulus factors that influence multisensory integration in human performance. Exp. Brain Res. 2012, 219, 121–137. [Google Scholar] [CrossRef] [Green Version]
- Brain, W.R. Visual orientation with special reference to lesions of the right cerebral hemisphere. Brain J. Neurol. 1941, 64, 244–272. [Google Scholar] [CrossRef]
- Hall, E.T. The Hidden Dimension; Doubleday: Garden City, NY, USA, 1966; Volume 609. [Google Scholar]
- Rizzolatti, G.; Scandolara, C.; Matelli, M.; Gentilucci, M. Afferent properties of periarcuate neurons in macaque monkeys. II. Visual responses. Behav. Brain Res. 1981, 2, 147–163. [Google Scholar] [CrossRef]
- Previc, F.H. The neuropsychology of 3-D space. Psychol. Bull. 1998, 124, 123. [Google Scholar] [CrossRef]
- Sambo, C.F.; Forster, B. An ERP investigation on visuotactile interactions in peripersonal and extrapersonal space: Evidence for the spatial rule. J. Cogn. Neurosci. 2009, 21, 1550–1559. [Google Scholar] [CrossRef]
- Soto-Faraco, S.; Lyons, J.; Gazzaniga, M.; Spence, C.; Kingstone, A. The ventriloquist in motion: Illusory capture of dynamic information across sensory modalities. Cogn. Brain Res. 2002, 14, 139–146. [Google Scholar] [CrossRef]
- Spence, C.; Squire, S. Multisensory integration: Maintaining the perception of synchrony. Curr. Biol. 2003, 13, R519–R521. [Google Scholar] [CrossRef] [Green Version]
- Zampini, M.; Shore, D.I.; Spence, C. Audiovisual temporal order judgments. Exp. Brain Res. 2003, 152, 198–210. [Google Scholar] [CrossRef] [PubMed]
- Ernst, M.O.; Banks, M.S. Humans integrate visual and haptic information in a statistically optimal fashion. Nature 2002, 415, 429. [Google Scholar] [CrossRef] [PubMed]
- Anderson, S.J.; Burr, D.C. Spatial summation properties of directionally selective mechanisms in human vision. JOSA A 1991, 8, 1330–1339. [Google Scholar] [CrossRef]
- Anderson, S.J.; Burr, D.C. Receptive field size of human motion detection units. Vision Res. 1987, 27, 621–635. [Google Scholar] [CrossRef]
- Burr, D.C.; Morrone, M.C.; Vaina, L.M. Large receptive fields for optic flow detection in humans. Vision Res. 1998, 38, 1731–1743. [Google Scholar] [CrossRef]
- Gescheider, G.A.; Bolanowski, S.J.; Pope, J.V.; Verrillo, R.T. A four-channel analysis of the tactile sensitivity of the fingertip: Frequency selectivity, spatial summation, and temporal summation. Somatosens. Mot. Res. 2002, 19, 114–124. [Google Scholar] [CrossRef] [PubMed]
- Verrillo, R.T.; Bolanowski, S.J.; McGlone, F.P. Subjective magnitude of tactile roughness. Somatosens. Mot. Res. 1999, 16, 352–360. [Google Scholar] [CrossRef]
- Verrillo, R.T.; Gescheider, G.A. Enhancement and summation in the perception of two successive vibrotactile stimuli. Percept. Psychophys. 1975, 18, 128–136. [Google Scholar] [CrossRef]
- Marchand, S.; Arsenault, P. Spatial summation for pain perception: Interaction of inhibitory and excitatory mechanisms. Pain 2002, 95, 201–206. [Google Scholar] [CrossRef]
- Shushruth, S.; Ichida, J.M.; Levitt, J.B.; Angelucci, A. Comparison of spatial summation properties of neurons in macaque V1 and V2. J. Neurophysiol. 2009, 102, 2069–2083. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Levitt, J.B.; Lund, J.S. Contrast dependence of contextual effects in primate visual cortex. Nature 1997, 387, 73–76. [Google Scholar] [CrossRef] [PubMed]
- Li, C.-Y.; Lei, J.-J.; Yao, H.-S. Shift in speed selectivity of visual cortical neurons: A neural basis of perceived motion contrast. Proc. Natl. Acad. Sci. USA 1999, 96, 4052–4056. [Google Scholar] [CrossRef] [Green Version]
- Schiatti, L.; Cappagli, G.; Martolini, C.; Maviglia, A.; Signorini, S.; Gori, M.; Crepaldi, M. A Novel Wearable and Wireless Device to Investigate Perception in Interactive Scenarios. In Proceedings of the 42nd Annual International Conference of the IEEE Engineering in Medicine & Biology Society (EMBC), EMBS Learning Academy, Montreal, QC, Canada, 20–24 July 2020; pp. 3252–3255. [Google Scholar]
- Foulke, E. Perception, Cognition, and Mobility of Blind Pedestrians. Spatial Abilities: Development and Physiological Foundations; Potegal, M., Ed.; Academic Press: New York, NY, USA, 1982; pp. 55–76. [Google Scholar]
- Merabet, L.B.; Pascual-Leone, A. Neural reorganization following sensory loss: The opportunity of change. Nat. Rev. Neurosci. 2010, 11, 44–52. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pasqualotto, A.; Spiller, M.J.; Jansari, A.S.; Proulx, M.J. Visual experience facilitates allocentric spatial representation. Behav. Brain Res. 2013, 236, 175–179. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nardini, M.; Thomas, R.L.; Knowland, V.C.P.; Braddick, O.J.; Atkinson, J. A viewpoint-independent process for spatial reorientation. Cognition 2009, 112, 241–248. [Google Scholar] [CrossRef]
- Vasilyeva, M.; Lourenco, S.F. Development of spatial cognition. Wiley Interdiscip. Rev. Cogn. Sci. 2012, 3, 349–362. [Google Scholar] [CrossRef]
- Macaluso, E.; Maravita, A. The representation of space near the body through touch and vision. Neuropsychologia 2010, 48, 782–795. [Google Scholar] [CrossRef]
- Wandell, B.A.; Dumoulin, S.O.; Brewer, A.A. Visual field maps in human cortex. Neuron 2007, 56, 366–383. [Google Scholar] [CrossRef] [Green Version]
- Golomb, J.D.; Chun, M.M.; Mazer, J.A. The native coordinate system of spatial attention is retinotopic. J. Neurosci. 2008, 28, 10654–10662. [Google Scholar] [CrossRef] [PubMed]
- Gardner, J.L.; Merriam, E.P.; Movshon, J.A.; Heeger, D.J. Maps of visual space in human occipital cortex are retinotopic, not spatiotopic. J. Neurosci. 2008, 28, 3988–3999. [Google Scholar] [CrossRef] [Green Version]
- Noory, B.; Herzog, M.H.; Ogmen, H. Spatial properties of non-retinotopic reference frames in human vision. Vision Res. 2015, 113, 44–54. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Foxe, J.J. Multisensory integration: Frequency tuning of audio-tactile integration. Curr. Biol. 2009, 19, R373–R375. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Foxe, J.J.; Morocz, I.A.; Murray, M.M.; Higgins, B.A.; Javitt, D.C.; Schroeder, C.E. Multisensory auditory-somatosensory interactions in early cortical processing revealed by high-density electrical mapping. Brain Res. Cogn. Brain Res. 2000, 10, 77–83. [Google Scholar] [CrossRef]
- Foxe, J.J.; Wylie, G.R.; Martinez, A.; Schroeder, C.E.; Javitt, D.C.; Guilfoyle, D.; Ritter, W.; Murray, M.M. Auditory-somatosensory multisensory processing in auditory association cortex: An fMRI study. J. Neurophysiol. 2002, 88, 540–543. [Google Scholar] [CrossRef] [PubMed]
- Meredith, M.A.; Allman, B.L.; Keniston, L.P.; Clemo, H.R. Auditory influences on non-auditory cortices. Hear. Res. 2009, 258, 64–71. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schroeder, C.E.; Lindsley, R.W.; Specht, C.; Marcovici, A.; Smiley, J.F.; Javitt, D.C. Somatosensory input to auditory association cortex in the macaque monkey. J. Neurophysiol. 2001, 85, 1322–1327. [Google Scholar] [CrossRef] [PubMed]
- Keniston, L.P.; Henderson, S.C.; Meredith, M.A. Neuroanatomical identification of crossmodal auditory inputs to interneurons in somatosensory cortex. Exp. Brain Res. 2010, 202, 725–731. [Google Scholar] [CrossRef] [Green Version]
- Finocchietti, S.; Cappagli, G.; Gori, M. Encoding audio motion: Spatial impairment in early blind individuals. Front. Psychol. 2015, 6, 1357. [Google Scholar] [CrossRef] [PubMed]
- Brandwein, A.B.; Foxe, J.J.; Russo, N.N.; Altschuler, T.S.; Gomes, H.; Molholm, S. The development of audiovisual multisensory integration across childhood and early adolescence: A high-density electrical mapping study. Cereb. Cortex 2011, 21, 1042–1055. [Google Scholar] [CrossRef] [PubMed]
- Soto-Faraco, S.; Spence, C.; Kingstone, A. Cross-modal dynamic capture: Congruency effects in the perception of motion across sensory modalities. J. Exp. Psychol. Hum. Percept. Perform. 2004, 30, 330. [Google Scholar] [CrossRef] [PubMed]
- Sanabria, D.; Soto-Faraco, S.; Spence, C. Spatiotemporal interactions between audition and touch depend on hand posture. Exp. Brain Res. 2005, 165, 505–514. [Google Scholar] [CrossRef]
- Sherrick, C.E. The antagonisms of hearing and touch. In Hearing and Davis: Essays Honoring Hallowell Davis; Wasington University: Washington, DC, USA, 1976; pp. 149–158. [Google Scholar]
- Hötting, K.; Röder, B. Hearing cheats touch, but less in congenitally blind than in sighted individuals. Psychol. Sci. 2004, 15, 60–64. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Main Effects | Bonferroni-IC | Bonferroni-RT |
| Active Units | One vs. Two: p < 0.001 | One vs. Two: p = 0.02 |
| One vs. Three: p < 0.001 | One vs. Three: p < 0.001 | |
| One vs. Four: p < 0.001 | One vs. Four: p < 0.001 | |
| Two vs. Three: p < 0.001 | Two vs. Three: p < 0.001 | |
| Two vs. Four: p = 0.89 | Two vs. Four: p < 0.001 | |
| Three vs. Four: p = 1.00 | Three vs. Four: p = 0.008 | |
| Stimulation | T vs. TV: p = 1.00 | |
| T vs. A: p = 0.017 | ||
| V vs. A/T/AT: p < 0.001 | T vs. V: p = 0.004 | |
| AV vs. A/T/AT: p < 0.001 | T vs. AT: p = 0.025 | |
| TV vs. A/T/AT: p < 0.001 | T vs. AV: p < 0.001 | |
| ATV vs. A/T/AT: p < 0.001 | T vs. ATV: p < 0.001 | |
| TV vs. A: p = 0.004 | ||
| TV vs. V/AT/AV: p < 0.001 | ||
| Interaction Effects | Bonferroni-IC | Bonferroni-RT |
| Stimulation × Active Units | V-One vs. V-One/Two/Three/Four: p = 1.00 | V-One vs. V-Three/Four: p < 0.001 |
| A-One vs. A-Two/Three/Four: p < 0.001 | V-Two vs. V-Four: p = 0.004 | |
| A-Two vs. A-Three: p = 0.049 | A-One vs. A-One/Two/Three/Four: p = 1.00 | |
| T-One vs. T-Two/Three/Four: p < 0.001 | T-One vs. T-One/Two/Three/Four: p = 1.00 | |
| T-Two vs. T-Three: p < 0.001 | AV-One vs. AV-Two: p = 0.005 | |
| T-Two vs. T-Four: p = 0.007 | AV-One vs. AV-Three/Four: p < 0.001 | |
| T-Three vs. A-Three: p = 0.038 | ||
| AV/TV/ATV-One vs. AV/TV/ATV-Two/Three/Four: p = 1.00 | AV-Three vs. AV-Four: p = 0.037 | |
| AT-One vs. AT-Two/Four: p = 1.00 | TV-One vs. TV-Two: p = 0.024 | |
| AT-One vs. AT-Three: p = 0.72 | ATV-One/Two vs. ATV-Three/Four: p < 0.001 | |
| AT-Two vs. AT-Three/Four: p = 1.00 | ATV-Three vs. ATV-Four: p = 0.026 | |
| AT-Three vs. AT- Four: p = 0.19 | AT-One vs. AT-Two: p = 1.00 | |
| AT-One vs. A-One: p < 0.001 | AT-One vs. AT-Three: p = 0.88 | |
| AT-One vs. T-One: p < 0.001 | AT-One vs. AT-Four: p = 0.96 | |
| AT-Four vs. A-Four: p < 0.001 | AT-Two vs. AT-Three/Four: p = 1 | |
| AT-Four vs. T-Four: p = 1.00 | AT-Three vs. AT-Four: p = 1.00 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Martolini, C.; Cappagli, G.; Signorini, S.; Gori, M. Effects of Increasing Stimulated Area in Spatiotemporally Congruent Unisensory and Multisensory Conditions. Brain Sci. 2021, 11, 343. https://doi.org/10.3390/brainsci11030343
Martolini C, Cappagli G, Signorini S, Gori M. Effects of Increasing Stimulated Area in Spatiotemporally Congruent Unisensory and Multisensory Conditions. Brain Sciences. 2021; 11(3):343. https://doi.org/10.3390/brainsci11030343
Chicago/Turabian StyleMartolini, Chiara, Giulia Cappagli, Sabrina Signorini, and Monica Gori. 2021. "Effects of Increasing Stimulated Area in Spatiotemporally Congruent Unisensory and Multisensory Conditions" Brain Sciences 11, no. 3: 343. https://doi.org/10.3390/brainsci11030343
APA StyleMartolini, C., Cappagli, G., Signorini, S., & Gori, M. (2021). Effects of Increasing Stimulated Area in Spatiotemporally Congruent Unisensory and Multisensory Conditions. Brain Sciences, 11(3), 343. https://doi.org/10.3390/brainsci11030343