
The Organ of Vision and the Stomatognathic System—Review of Association Studies and Evidence-Based Discussion

 ,
, 
 ,
, {kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. The Relationship between Organ of Vision and Stomatognathic System Dysfunctions
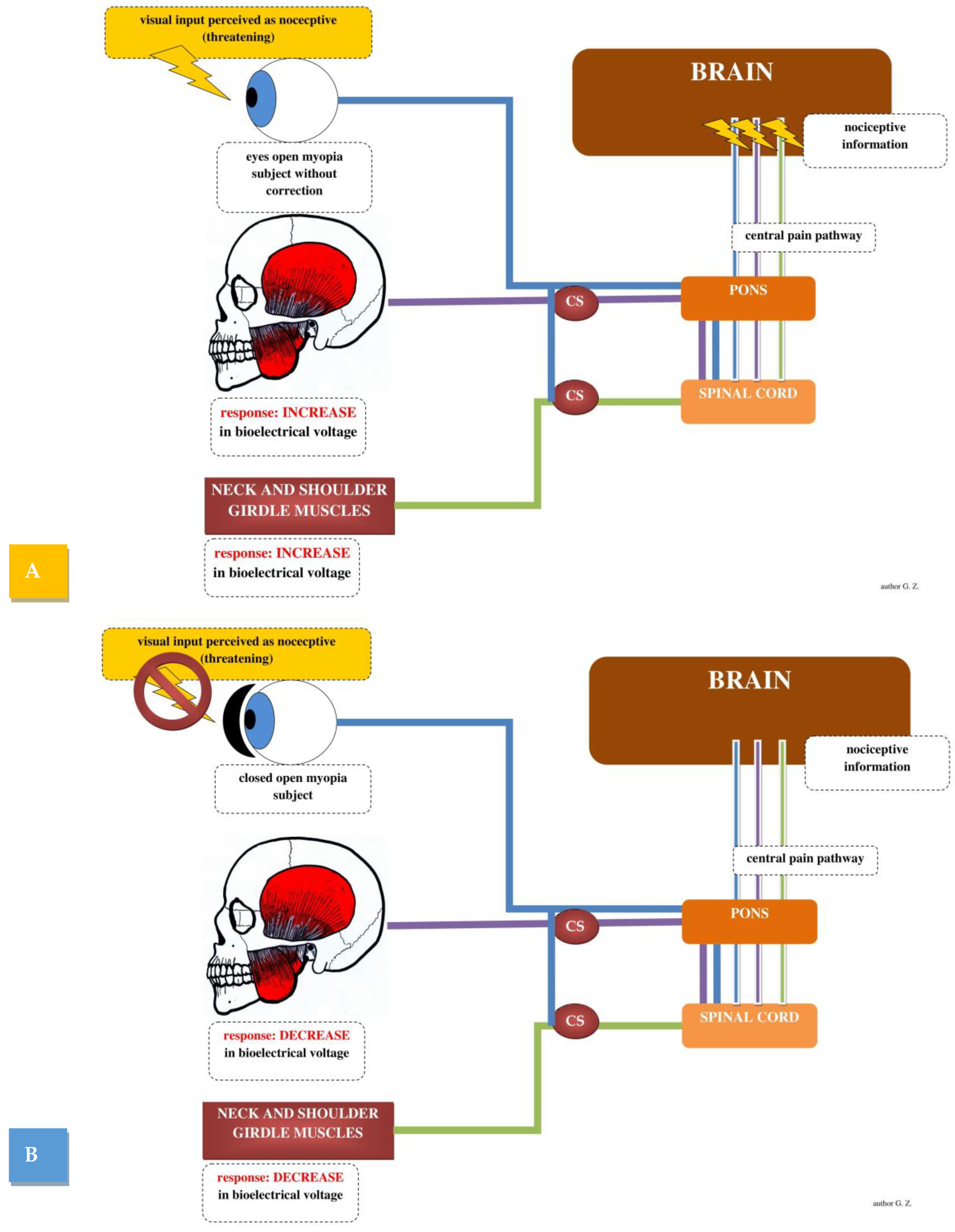
2.1. Visual Stimulation and Masticatory Muscles Activity
2.2. Structural Disorders in the Stomatognathic System and the Visual Impairment
3. Neurological Connection
3.1. Vestibulo–Ocular Reflex Connection
3.2. Central Sensitization Connection
4. Biochemical Connection
5. Muscular–Tendinous Connection
6. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Cuccia, A.; Caradonna, C. The Relationship between the Stomatognathic System and Body Posture. Clinics 2009, 64, 61–66. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kaltenborn, F.M. Manualne Mobilizacje Stawów Kończyn; Wydawnictwo Rolewski: Toruń, Poland, 1998. [Google Scholar]
- Okeson, J. Management of Temporomandibular Disorders and Occlusion, 7th ed.; Mosby: St. Louis, MO, USA, 2013; ISBN 978-0-323-08220-4. [Google Scholar]
- Maini, K.; Dua, A. Temporomandibular Joint Syndrome. In StatPearls; StatPearls Publishing: Treasure Island, FL, USA, 2021. [Google Scholar]
- Schuenke, M.; Schulte, E.; Schumacher, U. PROMETEUSZ. Atlas Anatomii Człowieka Tom III; MedPharm: Wrocław, Poland, 2015. [Google Scholar]
- Osiewicz, M.A.; Lobbezoo, F.; Loster, B.W.; Wilkosz, M.; Naeije, M.; Ohrbach, R. Research Diagnostic Criteria for Temporomandibular Disorders (RDC/TMD): The Polish Version of a Dual-Axis System for the Diagnosis of TMD.* RDC/TMD Form. J. Stomatol. 2013, 66, 576–649. [Google Scholar] [CrossRef]
- Al-Khotani, A.; Naimi-Akbar, A.; Albadawi, E.; Ernberg, M.; Hedenberg-Magnusson, B.; Christidis, N. Prevalence of Diagnosed Temporomandibular Disorders among Saudi Arabian Children and Adolescents. J. Headache Pain 2016, 17, 41. [Google Scholar] [CrossRef] [Green Version]
- Schiffman, E.; Ohrbach, R.; Truelove, E.; Look, J.; Anderson, G.; Goulet, J.-P.; List, T.; Svensson, P.; Gonzalez, Y.; Lobbezoo, F.; et al. Diagnostic Criteria for Temporomandibular Disorders (DC/TMD) for Clinical and Research Applications: Recommendations of the International RDC/TMD Consortium Network and Orofacial Pain Special Interest Group. J. Oral Facial Pain Headache 2014, 28, 6–27. [Google Scholar] [CrossRef]
- Martins-Júnior, R.L.; Palma, A.J.G.; Marquardt, E.J.; de Barros Gondin, T.M.; de Carvalho Kerber, F. Temporomandibular Disorders: A Report of 124 Patients. J. Contemp. Dent. Pract. 2010, 11, 71–78. [Google Scholar] [CrossRef]
- Jaeger, J.O.; Oakley, P.A.; Moore, R.R.; Ruggeroli, E.P.; Harrison, D.E. Resolution of Temporomandibular Joint Dysfunction (TMJD) by Correcting a Lateral Head Translation Posture Following Previous Failed Traditional Chiropractic Therapy: A CBP® Case Report. J. Phys. Ther. Sci. 2018, 30, 103–107. [Google Scholar] [CrossRef] [Green Version]
- Lluch, E.; Nijs, J.; De Kooning, M.; Van Dyck, D.; Vanderstraeten, R.; Struyf, F.; Roussel, N.A. Prevalence, Incidence, Localization, and Pathophysiology of Myofascial Trigger Points in Patients With Spinal Pain: A Systematic Literature Review. J. Manip. Physiol. Ther. 2015, 38, 587–600. [Google Scholar] [CrossRef]
- Chiarotto, A.; Clijsen, R.; Fernandez-de-Las-Penas, C.; Barbero, M. Prevalence of Myofascial Trigger Points in Spinal Disorders: A Systematic Review and Meta-Analysis. Arch. Phys. Med. Rehabil. 2016, 97, 316–337. [Google Scholar] [CrossRef] [PubMed]
- Marchili, N.; Ortu, E.; Pietropaoli, D.; Cattaneo, R.; Monaco, A. Dental Occlusion and Ophthalmology: A Literature Review. Open Dent. J. 2016, 10, 460–468. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lens, A.; Nemeth, S.C.; Ledford, J.K. Ocular Anatomy and Physiology, 2nd ed.; SLACK: Thorofare, NJ, USA, 2008; ISBN 978-1-55642-792-3. [Google Scholar]
- Carr, B.J.; Stell, W.K. The Science Behind Myopia. In Webvision: The Organization of the Retina and Visual System; Kolb, H., Fernandez, E., Nelson, R., Eds.; University of Utah Health Sciences Center: Salt Lake City, UT, USA, 1995. [Google Scholar]
- Baird, P.N.; Saw, S.-M.; Lanca, C.; Guggenheim, J.A.; Smith, E.L., III; Zhou, X.; Matsui, K.-O.; Wu, P.-C.; Sankaridurg, P.; Chia, A.; et al. Myopia. Nat. Rev. Dis. Primers 2020, 6, 99. [Google Scholar] [CrossRef]
- Resnikoff, S.; Jonas, J.B.; Friedman, D.; He, M.; Jong, M.; Nichols, J.J.; Ohno-Matsui, K.; Smith, E.L., III; Wildsoet, C.F.; Taylor, H.R.; et al. Myopia—A 21st Century Public Health Issue. Investig. Ophthalmol. Vis. Sci. 2019, 60, Mi–Mii. [Google Scholar] [CrossRef] [Green Version]
- Morgan, I.G.; French, A.N.; Ashby, R.S.; Guo, X.; Ding, X.; He, M.; Rose, K.A. The Epidemics of Myopia: Aetiology and Prevention. Prog. Retin. Eye Res. 2018, 62, 134–149. [Google Scholar] [CrossRef] [Green Version]
- Holden, B.A.; Fricke, T.R.; Wilson, D.A.; Jong, M.; Naidoo, K.S.; Sankaridurg, P.; Wong, T.Y.; Naduvilath, T.J.; Resnikoff, S. Global Prevalence of Myopia and High Myopia and Temporal Trends from 2000 through 2050. Ophthalmology 2016, 123, 1036–1042. [Google Scholar] [CrossRef] [Green Version]
- Naidoo, K.S.; Fricke, T.R.; Frick, K.D.; Jong, M.; Naduvilath, T.J.; Resnikoff, S.; Sankaridurg, P. Potential Lost Productivity Resulting from the Global Burden of Myopia: Systematic Review, Meta-Analysis, and Modeling. Ophthalmology 2019, 126, 338–346. [Google Scholar] [CrossRef] [Green Version]
- Fricke, T.R.; Jong, M.; Naidoo, K.S.; Sankaridurg, P.; Naduvilath, T.J.; Ho, S.M.; Wong, T.Y.; Resnikoff, S. Global Prevalence of Visual Impairment Associated with Myopic Macular Degeneration and Temporal Trends from 2000 through 2050: Systematic Review, Meta-Analysis and Modelling. Br. J. Ophthalmol. 2018, 102, 855–862. [Google Scholar] [CrossRef]
- Haarman, A.E.G.; Enthoven, C.A.; Tideman, J.W.L.; Tedja, M.S.; Verhoeven, V.J.M.; Klaver, C.C.W. The Complications of Myopia: A Review and Meta-Analysis. Investig. Ophthalmol. Vis. Sci. 2020, 61, 49. [Google Scholar] [CrossRef] [PubMed]
- Ludwig, P.E.; Lopez, M.J.; Czyz, C.N. Embryology, Eye Malformations. In StatPearls; StatPearls Publishing: Treasure Island, FL, USA, 2021. [Google Scholar]
- Öğütcen-Toller, M.; Keskin, M. Computerized 3-Dimensional Study of the Embryologic Development of the Human Masticatory Muscles and Temporomandibular Joint. J. Oral Maxillofac. Surg. 2000, 58, 1381–1386. [Google Scholar] [CrossRef] [PubMed]
- Ludwig, M.; Busch, L.C.; Winking, H. The Embryonic Development of Sensory Organs and the Skull in the Trisomy 16 Mouse, an Animal Model for Down’s Syndrome. Ann. Anat. Anat. Anz. 1997, 179, 525–533. [Google Scholar] [CrossRef]
- Krinsky-McHale, S.J.; Jenkins, E.C.; Zigman, W.B.; Silverman, W. Ophthalmic Disorders in Adults with down Syndrome. Curr. Gerontol. Geriatr. Res. 2012, 2012, 974253. [Google Scholar] [CrossRef]
- Kaczorowska, N.; Kaczorowski, K.; Laskowska, J.; Mikulewicz, M. Down Syndrome as a Cause of Abnormalities in the Craniofacial Region: A Systematic Literature Review. Adv. Clin. Exp. Med. 2019, 28, 1587–1592. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zieliński, G.; Matysik-Woźniak, A.; Rapa, M.; Baszczowski, M.; Ginszt, M.; Zawadka, M.; Szkutnik, J.; Rejdak, R.; Gawda, P. The Influence of Visual Input on Electromyographic Patterns of Masticatory and Cervical Spine Muscles in Subjects with Myopia. J. Clin. Med. 2021, 10, 5376. [Google Scholar] [CrossRef] [PubMed]
- Widmalm, S.-E.; Ericsson, S.G. The Influence of Eye Closure on Muscle Activity in the Anterior Temporal Region. J. Oral Rehabil. 1983, 10, 25–29. [Google Scholar] [CrossRef] [PubMed]
- Holmgren, K.; Sheikholeslam, A.; Riise, C. An Electromyographic Study of the Immediate Effect of an Occlusal Splint on the Postural Activity of the Anterior Temporal and Masseter Muscles in Different Body Positions with and without Visual Input. J. Oral Rehabil. 1985, 12, 483–490. [Google Scholar] [CrossRef] [PubMed]
- Miralles, R.; Valenzuela, S.; Ramirez, P.; Santander, H.; Palazzi, C.; Ormeño, G.; Zúñiga, C. Visual Input Effect on EMG Activity of Sternocleidomastoid and Masseter Muscles in Healthy Subjects and in Patients with Myogenic Cranio-Cervical-Mandibular Dysfunction. Cranio J. Craniomandib. Pract. 1998, 16, 168–184. [Google Scholar] [CrossRef]
- Monaco, A.; Cattaneo, R.; Spadaro, A.; Giannoni, M.; Di Martino, S.; Gatto, R. Visual Input Effect on EMG Activity of Masticatory and Postural Muscles in Healthy and in Myopic Children. Eur. J. Paediatr. Dent. 2006, 7, 18–22. [Google Scholar]
- Monaco, A.; Cattaneo, R.; Spadaro, A.; D’Andrea, P.; Marzo, G.; Gatto, R. Ocular Correction Effects on EMG Activity of Stomatognathic Muscles in Children with Functional Mandibular Lateral-Deviation: A Case Control Study. Eur. J. Paediatr. Dent. 2006, 7, 81–88. [Google Scholar]
- Spadaro, A.; Monaco, A.; Cattaneo, R.; Masci, C.; Gatto, R. Effect on Anterior Temporalis Surface EMG of Eyes Open-Closed Condition. Eur. J. Paediatr. Dent. 2010, 11, 210–212. [Google Scholar]
- Ciavarella, D.; Palazzo, A.; De Lillo, A.; Lo Russo, L.; Paduano, S.; Laino, L.; Chimenti, C.; Frezza, F.; Lo Muzio, L. Influence of Vision on Masticatory Muscles Function: Surface Electromyographic Evaluation. Ann. Stomatol. 2014, 5, 61–65. [Google Scholar] [CrossRef]
- Fiorucci, E.; Bucci, G.; Cattaneo, R.; Monaco, A. The Measurement of Surface Electromyographic Signal in Rest Position for the Correct Prescription of Eyeglasses. IEEE Trans. Instrum. Meas. 2012, 61, 419–428. [Google Scholar] [CrossRef]
- Monaco, A.; Ortu, E.; Giannoni, M.; D’Andrea, P.; Cattaneo, R.; Mummolo, A.; Pietropaoli, D. Standard Correction of Vision Worsens EMG Activity of Pericranial Muscles in Chronic TMD Subjects. Pain Res. Manag. 2020, 2020, e3932476. [Google Scholar] [CrossRef] [Green Version]
- Cuccia, A.M.; Caradonna, C. Binocular Motility System and Temporomandibular Joint Internal Derangement: A Study in Adults. Am. J. Orthod. Dentofac. Orthop. Off. Publ. Am. Assoc. Orthod. Its Const. Soc. Am. Board Orthod. 2008, 133, 640.e15–640.e20. [Google Scholar] [CrossRef] [PubMed]
- Monaco, A.; Streni, O.; Marci, M.C.; Sabetti, L.; Marzo, G.; Giannoni, M. Relationship between Mandibular Deviation and Ocular Convergence. J. Clin. Pediatr. Dent. 2004, 28, 135–138. [Google Scholar] [CrossRef]
- Monaco, A.; Spadaro, A.; Sgolastra, F.; Petrucci, A.; D’Andrea, P.D.; Gatto, R. Prevalence of Astigmatism in a Paediatric Population with Malocclusions. Eur. J. Paediatr. Dent. 2011, 12, 91–94. [Google Scholar] [PubMed]
- Monaco, A.; Sgolastra, F.; Cattaneo, R.; Petrucci, A.; Marci, M.C.; D’Andrea, P.D.; Gatto, R. Prevalence of Myopia in a Population with Malocclusions. Eur. J. Paediatr. Dent. 2012, 13, 256–258. [Google Scholar]
- Bollero, P.; Ricchiuti, M.R.; Laganà, G.; Di Fusco, G.; Lione, R.; Cozza, P. Correlations between Dental Malocclusions, Ocular Motility, and Convergence Disorders: A Cross-Sectional Study in Growing Subjects. Oral Implantol. 2017, 10, 289–294. [Google Scholar] [CrossRef]
- Vompi, C.; Serritella, E.; Galluccio, G.; Pistella, S.; Segnalini, A.; Giannelli, L.; Di Paolo, C. Evaluation of Vision in Gnathological and Orthodontic Patients with Temporomandibular Disorders: A Prospective Experimental Observational Cohort Study. J. Int. Soc. Prev. Community Dent. 2020, 10, 481–490. [Google Scholar] [CrossRef]
- Somisetty, S.; Das, J.M. Neuroanatomy, Vestibulo-Ocular Reflex. In StatPearls; StatPearls Publishing: Treasure Island, FL, USA, 2021. [Google Scholar]
- Stack, B.; Sims, A. The Relationship between Posture and Equilibrium and the Auriculotemporal Nerve in Patients with Disturbed Gait and Balance. Cranio J. Craniomandib. Pract. 2009, 27, 248–260. [Google Scholar] [CrossRef]
- Manivannan, M.; Suresh, P.K. On the Somatosensation of Vision. Ann. Neurosci. 2012, 19, 31–39. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Weir, C.R.; Knox, P.C.; Dutton, G.N. Does Extraocular Muscle Proprioception Influence Oculomotor Control? Br. J. Ophthalmol. 2000, 84, 1071–1074. [Google Scholar] [CrossRef] [Green Version]
- Donaldson, I.M. The Functions of the Proprioceptors of the Eye Muscles. Philos. Trans. R. Soc. Lond. Ser. B Biol. Sci. 2000, 355, 1685–1754. [Google Scholar] [CrossRef] [Green Version]
- Mangold, S.A.; Das, J.M. Neuroanatomy, Reticular Formation. In StatPearls; StatPearls Publishing: Treasure Island, FL, USA, 2021. [Google Scholar]
- Büttner-Ennever, J.A.; Büttner, U. Neuroanatomy of the Oculomotor System. The Reticular Formation. Rev. Oculomot. Res. 1988, 2, 119–176. [Google Scholar] [PubMed]
- Bradnam, L.; Barry, C. The Role of the Trigeminal Sensory Nuclear Complex in the Pathophysiology of Craniocervical Dystonia. J. Neurosci. 2013, 33, 18358–18367. [Google Scholar] [CrossRef] [Green Version]
- Torvik, A. Afferent Connections to the Sensory Trigeminal Nuclei, the Nucleus of the Solitary Tract and Adjacent Structures. An Experimental Study in the Rat. J. Comp. Neurol. 1956, 106, 51–141. [Google Scholar] [CrossRef]
- Alonso-Blanco, C.; Fernández-de-Las-Peñas, C.; de-la-Llave-Rincón, A.I.; Zarco-Moreno, P.; Galán-Del-Río, F.; Svensson, P. Characteristics of Referred Muscle Pain to the Head from Active Trigger Points in Women with Myofascial Temporomandibular Pain and Fibromyalgia Syndrome. J. Headache Pain 2012, 13, 625–637. [Google Scholar] [CrossRef] [Green Version]
- Zieliński, G.; Byś, A.; Szkutnik, J.; Majcher, P.; Ginszt, M. Electromyographic Patterns of Masticatory Muscles in Relation to Active Myofascial Trigger Points of the Upper Trapezius and Temporomandibular Disorders. Diagnostics 2021, 11, 580. [Google Scholar] [CrossRef]
- Ginszt, M.; Zieliński, G.; Berger, M.; Szkutnik, J.; Bakalczuk, M.; Majcher, P. Acute Effect of the Compression Technique on the Electromyographic Activity of the Masticatory Muscles and Mouth Opening in Subjects with Active Myofascial Trigger Points. Appl. Sci. 2020, 10, 7750. [Google Scholar] [CrossRef]
- Woolf, C.J. Central Sensitization: Implications for the Diagnosis and Treatment of Pain. Pain 2011, 152, S2–S15. [Google Scholar] [CrossRef]
- Filbrich, L.; Alamia, A.; Burns, S.; Legrain, V. Orienting Attention in Visual Space by Nociceptive Stimuli: Investigation with a Temporal Order Judgment Task Based on the Adaptive PSI Method. Exp. Brain Res. 2017, 235, 2069–2079. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van Ryckeghem, D.M.L.; Van Damme, S.; Crombez, G.; Eccleston, C.; Verhoeven, K.; Legrain, V. The Role of Spatial Attention in Attentional Control over Pain: An Experimental Investigation. Exp. Brain Res. 2011, 208, 269–275. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Williams, D.A. Phenotypic Features of Central Sensitization. J. Appl. Biobehav. Res. 2018, 23, e12135. [Google Scholar] [CrossRef]
- Mancini, F.; Beaumont, A.-L.; Hu, L.; Haggard, P.; Iannetti, G.D.D. Touch Inhibits Subcortical and Cortical Nociceptive Responses. Pain 2015, 156, 1936–1944. [Google Scholar] [CrossRef] [Green Version]
- Braz, J.; Solorzano, C.; Wang, X.; Basbaum, A.I. Transmitting Pain and Itch Messages: A Contemporary View of the Spinal Cord Circuits That Generate Gate Control. Neuron 2014, 82, 522–536. [Google Scholar] [CrossRef] [Green Version]
- Mendell, L.M. Constructing and Deconstructing the Gate Theory of Pain. Pain 2014, 155, 210–216. [Google Scholar] [CrossRef] [Green Version]
- Zorena, K.; Gładysiak, A.; Ślęzak, D. Early Intervention and Nonpharmacological Therapy of Myopia in Young Adults. J. Ophthalmol. 2018, 2018, 4680603. [Google Scholar] [CrossRef] [Green Version]
- Moore, M.K. Upper Crossed Syndrome and Its Relationship to Cervicogenic Headache. J. Manip. Physiol. Ther. 2004, 27, 414–420. [Google Scholar] [CrossRef]
- Fernández-de-Las-Peñas, C.; Gerwin, R.D.; Arendt-Nielsen, L.L. Tension-Type and Cervicogenic Headache: Pathophysiology, Diagnosis, and Management; Contemporary Issues in Physical Therapy and Rehabilitation Medicine; Jones and Bartlett Publishers: Burlington, MA, USA, 2010; ISBN 978-0-7637-5283-5. [Google Scholar]
- Saran, T.; Turska, M.; Kocki, T.; Zawadka, M.; Zieliński, G.; Turski, W.A.; Gawda, P. Effect of 4-Week Physical Exercises on Tryptophan, Kynurenine and Kynurenic Acid Content in Human Sweat. Sci. Rep. 2021, 11, 11092. [Google Scholar] [CrossRef]
- Walczak, K.; Wnorowski, A.; Turski, W.A.; Plech, T. Kynurenic Acid and Cancer: Facts and Controversies. Cell. Mol. Life Sci. CMLS 2020, 77, 1531–1550. [Google Scholar] [CrossRef] [Green Version]
- Sas, K.; Szabó, E.; Vécsei, L. Mitochondria, Oxidative Stress and the Kynurenine System, with a Focus on Ageing and Neuroprotection. Molecules 2018, 23, 191. [Google Scholar] [CrossRef] [Green Version]
- Ferreira, F.S.; Biasibetti-Brendler, H.; Pierozan, P.; Schmitz, F.; Bertó, C.G.; Prezzi, C.A.; Manfredini, V.; Wyse, A.T.S. Kynurenic Acid Restores Nrf2 Levels and Prevents Quinolinic Acid-Induced Toxicity in Rat Striatal Slices. Mol. Neurobiol. 2018, 55, 8538–8549. [Google Scholar] [CrossRef] [PubMed]
- Pierozan, P.; Biasibetti-Brendler, H.; Schmitz, F.; Ferreira, F.; Pessoa-Pureur, R.; Wyse, A.T.S. Kynurenic Acid Prevents Cytoskeletal Disorganization Induced by Quinolinic Acid in Mixed Cultures of Rat Striatum. Mol. Neurobiol. 2018, 55, 5111–5124. [Google Scholar] [CrossRef]
- Nguyen, K.H.; Patel, B.C.; Tadi, P. Anatomy, Head and Neck, Eye Retina. In StatPearls; StatPearls Publishing: Treasure Island, FL, USA, 2021. [Google Scholar]
- Rejdak, R.; Zielinska, E.; Shenk, Y.; Turski, W.A.; Okuno, E.; Zarnowski, T.; Zagorski, Z.; Zrenner, E.; Kohler, K. Ontogenic Changes of Kynurenine Aminotransferase I Activity and Its Expression in the Chicken Retina. Vis. Res. 2003, 43, 1513–1517. [Google Scholar] [CrossRef] [Green Version]
- Rejdak, R.; Zarnowski, T.; Turski, W.A.; Okuno, E.; Kocki, T.; Zagórski, Z.; Kohler, K.; Guenther, E.; Zrenner, E. Presence of Kynurenic Acid and Kynurenine Aminotransferases in the Inner Retina. Neuroreport 2001, 12, 3675–3678. [Google Scholar] [CrossRef]
- Rejdak, R.; Shenk, Y.; Schuettauf, F.; Turski, W.A.; Okuno, E.; Zagorski, Z.; Zrenner, E.; Kohler, K. Expression of Kynurenine Aminotransferases in the Rat Retina during Development. Vis. Res. 2004, 44, 1–7. [Google Scholar] [CrossRef] [Green Version]
- Rejdak, R.; Zarnowski, T.; Turski, W.A.; Kocki, T.; Zagorski, Z.; Zrenner, E.; Schuettauf, F. Alterations of Kynurenic Acid Content in the Retina in Response to Retinal Ganglion Cell Damage. Vis. Res. 2003, 43, 497–503. [Google Scholar] [CrossRef] [Green Version]
- Hu, P.; Hunt, N.H.; Arfuso, F.; Shaw, L.C.; Uddin, M.N.; Zhu, M.; Devasahayam, R.; Adamson, S.J.; Benson, V.L.; Chan-Ling, T.; et al. Increased Indoleamine 2,3-Dioxygenase and Quinolinic Acid Expression in Microglia and Müller Cells of Diabetic Human and Rodent Retina. Investig. Ophthalmol. Vis. Sci. 2017, 58, 5043. [Google Scholar] [CrossRef] [Green Version]
- Fiedorowicz, M.; Choragiewicz, T.; Thaler, S.; Schuettauf, F.; Nowakowska, D.; Wojtunik, K.; Reibaldi, M.; Avitabile, T.; Kocki, T.; Turski, W.A.; et al. Tryptophan and Kynurenine Pathway Metabolites in Animal Models of Retinal and Optic Nerve Damage: Different Dynamics of Changes. Front. Physiol. 2019, 10, 1254. [Google Scholar] [CrossRef]
- Atchison, D.A.; Pritchard, N.; Schmid, K.L.; Scott, D.H.; Jones, C.E.; Pope, J.M. Shape of the Retinal Surface in Emmetropia and Myopia. Investig. Ophthalmol. Vis. Sci. 2005, 46, 2698–2707. [Google Scholar] [CrossRef] [Green Version]
- Jonas, J.B.; Ohno-Matsui, K.; Panda-Jonas, S. Myopia: Anatomic Changes and Consequences for Its Etiology. Asia-Pac. J. Ophthalmol. 2019, 8, 355–359. [Google Scholar] [CrossRef]
- Wang, Y.X.; Panda-Jonas, S.; Jonas, J.B. Optic Nerve Head Anatomy in Myopia and Glaucoma, Including Parapapillary Zones Alpha, Beta, Gamma and Delta: Histology and Clinical Features. Prog. Retin. Eye Res. 2021, 83, 100933. [Google Scholar] [CrossRef] [PubMed]
- Agudelo, L.Z.; Femenía, T.; Orhan, F.; Porsmyr-Palmertz, M.; Goiny, M.; Martinez-Redondo, V.; Correia, J.C.; Izadi, M.; Bhat, M.; Schuppe-Koistinen, I.; et al. Skeletal Muscle PGC-1α1 Modulates Kynurenine Metabolism and Mediates Resilience to Stress-Induced Depression. Cell 2014, 159, 33–45. [Google Scholar] [CrossRef] [Green Version]
- Schlittler, M.; Goiny, M.; Agudelo, L.Z.; Venckunas, T.; Brazaitis, M.; Skurvydas, A.; Kamandulis, S.; Ruas, J.L.; Erhardt, S.; Westerblad, H.; et al. Endurance Exercise Increases Skeletal Muscle Kynurenine Aminotransferases and Plasma Kynurenic Acid in Humans. Am. J. Physiol. Cell Physiol. 2016, 310, C836–C840. [Google Scholar] [CrossRef] [Green Version]
- Mudry, J.M.; Alm, P.S.; Erhardt, S.; Goiny, M.; Fritz, T.; Caidahl, K.; Zierath, J.R.; Krook, A.; Wallberg-Henriksson, H. Direct Effects of Exercise on Kynurenine Metabolism in People with Normal Glucose Tolerance or Type 2 Diabetes. Diabetes Metab. Res. Rev. 2016, 32, 754–761. [Google Scholar] [CrossRef] [Green Version]
- Notarangelo, F.M.; Pocivavsek, A.; Schwarcz, R. Exercise Your Kynurenines to Fight Depression. Trends Neurosci. 2018, 41, 491–493. [Google Scholar] [CrossRef]
- Pedersen, B.K. Physical Activity and Muscle-Brain Crosstalk. Nat. Rev. Endocrinol. 2019, 15, 383–392. [Google Scholar] [CrossRef]
- Cervenka, I.; Agudelo, L.Z.; Ruas, J.L. Kynurenines: Tryptophan’s Metabolites in Exercise, Inflammation, and Mental Health. Science 2017, 357, eaaf9794. [Google Scholar] [CrossRef] [Green Version]
- Martin, K.S.; Azzolini, M.; Lira Ruas, J. The Kynurenine Connection: How Exercise Shifts Muscle Tryptophan Metabolism and Affects Energy Homeostasis, the Immune System, and the Brain. Am. J. Physiol.-Cell Physiol. 2020, 318, C818–C830. [Google Scholar] [CrossRef]
- Wyckelsma, V.L.; Lindkvist, W.; Venckunas, T.; Brazaitis, M.; Kamandulis, S.; Pääsuke, M.; Ereline, J.; Westerblad, H.; Andersson, D.C. Kynurenine Aminotransferase Isoforms Display Fiber-Type Specific Expression in Young and Old Human Skeletal Muscle. Exp. Gerontol. 2020, 134, 110880. [Google Scholar] [CrossRef]
- Suhr Thykjaer, A.; Lundberg, K.; Grauslund, J. Physical Activity in Relation to Development and Progression of Myopia—A Systematic Review. Acta Ophthalmol. 2017, 95, 651–659. [Google Scholar] [CrossRef] [Green Version]
- Barjandi, G.; Louca Jounger, S.; Löfgren, M.; Bileviciute-Ljungar, I.; Kosek, E.; Ernberg, M. Plasma Tryptophan and Kynurenine in Females with Temporomandibular Disorders and Fibromyalgia-An Exploratory Pilot Study. J. Oral Rehabil. 2020, 47, 150–157. [Google Scholar] [CrossRef]
- Tuka, B.; Nyári, A.; Cseh, E.K.; Körtési, T.; Veréb, D.; Tömösi, F.; Kecskeméti, G.; Janáky, T.; Tajti, J.; Vécsei, L. Clinical Relevance of Depressed Kynurenine Pathway in Episodic Migraine Patients: Potential Prognostic Markers in the Peripheral Plasma during the Interictal Period. J. Headache Pain 2021, 22, 60. [Google Scholar] [CrossRef]
- Fejes, A.; Párdutz, Á.; Toldi, J.; Vécsei, L. Kynurenine Metabolites and Migraine: Experimental Studies and Therapeutic Perspectives. Curr. Neuropharmacol. 2011, 9, 376–387. [Google Scholar] [CrossRef] [Green Version]
- Harle, D.E.; Evans, B.J.W. The Correlation between Migraine Headache and Refractive Errors. Optom. Vis. Sci. Off. Publ. Am. Acad. Optom. 2006, 83, 82–87. [Google Scholar] [CrossRef]
- Lajmi, H.; Choura, R.; Ben Achour, B.; Doukh, M.; Amin, Z.; Hmaied, W. Headache Associated with Refractive Errors: Characteristics and Risk Factors. Rev. Neurol. 2021, 177, 947–954. [Google Scholar] [CrossRef]
- Franco, A.L.; Gonçalves, D.A.G.; Castanharo, S.M.; Speciali, J.G.; Bigal, M.E.; Camparis, C.M. Migraine Is the Most Prevalent Primary Headache in Individuals with Temporomandibular Disorders. J. Orofac. Pain 2010, 24, 287–292. [Google Scholar]
- Goncalves, D.A.G.; Camparis, C.M.; Speciali, J.G.; Castanharo, S.M.; Ujikawa, L.T.; Lipton, R.B.; Bigal, M.E. Treatment of Comorbid Migraine and Temporomandibular Disorders: A Factorial, Double-Blind, Randomized, Placebo-Controlled Study. J. Orofac. Pain 2013, 27, 325–335. [Google Scholar] [CrossRef]
- Gatt, A.; Agarwal, S.; Zito, P.M. Anatomy, Fascia Layers. In StatPearls; StatPearls Publishing: Treasure Island, FL, USA, 2021. [Google Scholar]
- Schleip, R.; Jäger, H.; Klingler, W. What Is “Fascia”? A Review of Different Nomenclatures. J. Bodyw. Mov. Ther. 2012, 16, 496–502. [Google Scholar] [CrossRef]
- Stecco, C.; Macchi, V.; Porzionato, A.; Duparc, F.; De Caro, R. The Fascia: The Forgotten Structure. Ital. J. Anat. Embryol. Arch. Ital. Anat. Ed. Embriol. 2011, 116, 127–138. [Google Scholar]
- Klingler, W.; Velders, M.; Hoppe, K.; Pedro, M.; Schleip, R. Clinical Relevance of Fascial Tissue and Dysfunctions. Curr. Pain Headache Rep. 2014, 18, 439. [Google Scholar] [CrossRef]
- Bordoni, B.; Mahabadi, N.; Varacallo, M. Anatomy, Fascia. In StatPearls; StatPearls Publishing: Treasure Island, FL, USA, 2021. [Google Scholar]
- Findley, T.; Chaudhry, H.; Stecco, A.; Roman, M. Fascia Research—A Narrative Review. J. Bodyw. Mov. Ther. 2012, 16, 67–75. [Google Scholar] [CrossRef]
- Stecco, C.; Hammer, W.I. Functional Atlas of the Human Fascial System; Elsevier Ltd.: Edinburgh, UK, 2015; ISBN 978-0-7020-4430-4. [Google Scholar]
- Raja, G.P.; Bhat, N.S.; Fernández-de-las-Peñas, C.; Gangavelli, R.; Davis, F.; Shankar, R.; Prabhu, A. Effectiveness of Deep Cervical Fascial Manipulation and Yoga Postures on Pain, Function, and Oculomotor Control in Patients with Mechanical Neck Pain: Study Protocol of a Pragmatic, Parallel-Group, Randomized, Controlled Trial. Trials 2021, 22, 574. [Google Scholar] [CrossRef]
- Fredrick, D.R. Myopia. BMJ 2002, 324, 1195–1199. [Google Scholar] [CrossRef] [PubMed]
- Marumoto, T.; Sotoyama, M.; Villanueva, M.B.G.; Jonai, H.; Yamada, H.; Kanai, A.; Saito, S. Relationship Between Posture and Myopia Among Students. In Myopia Updates: Proceedings of the 6th International Conference on Myopia; Tokoro, T., Ed.; Springer: Tokyo, Japan, 1998; pp. 63–66. ISBN 978-4-431-66959-3. [Google Scholar]



Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zieliński, G.; Filipiak, Z.; Ginszt, M.; Matysik-Woźniak, A.; Rejdak, R.; Gawda, P. The Organ of Vision and the Stomatognathic System—Review of Association Studies and Evidence-Based Discussion. Brain Sci. 2022, 12, 14. https://doi.org/10.3390/brainsci12010014
Zieliński G, Filipiak Z, Ginszt M, Matysik-Woźniak A, Rejdak R, Gawda P. The Organ of Vision and the Stomatognathic System—Review of Association Studies and Evidence-Based Discussion. Brain Sciences. 2022; 12(1):14. https://doi.org/10.3390/brainsci12010014
Chicago/Turabian StyleZieliński, Grzegorz, Zuzanna Filipiak, Michał Ginszt, Anna Matysik-Woźniak, Robert Rejdak, and Piotr Gawda. 2022. "The Organ of Vision and the Stomatognathic System—Review of Association Studies and Evidence-Based Discussion" Brain Sciences 12, no. 1: 14. https://doi.org/10.3390/brainsci12010014
APA StyleZieliński, G., Filipiak, Z., Ginszt, M., Matysik-Woźniak, A., Rejdak, R., & Gawda, P. (2022). The Organ of Vision and the Stomatognathic System—Review of Association Studies and Evidence-Based Discussion. Brain Sciences, 12(1), 14. https://doi.org/10.3390/brainsci12010014