



Electrophysiological Evidence for A Number–Action Mapping in Infancy


{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Participants
2.2. Stimuli
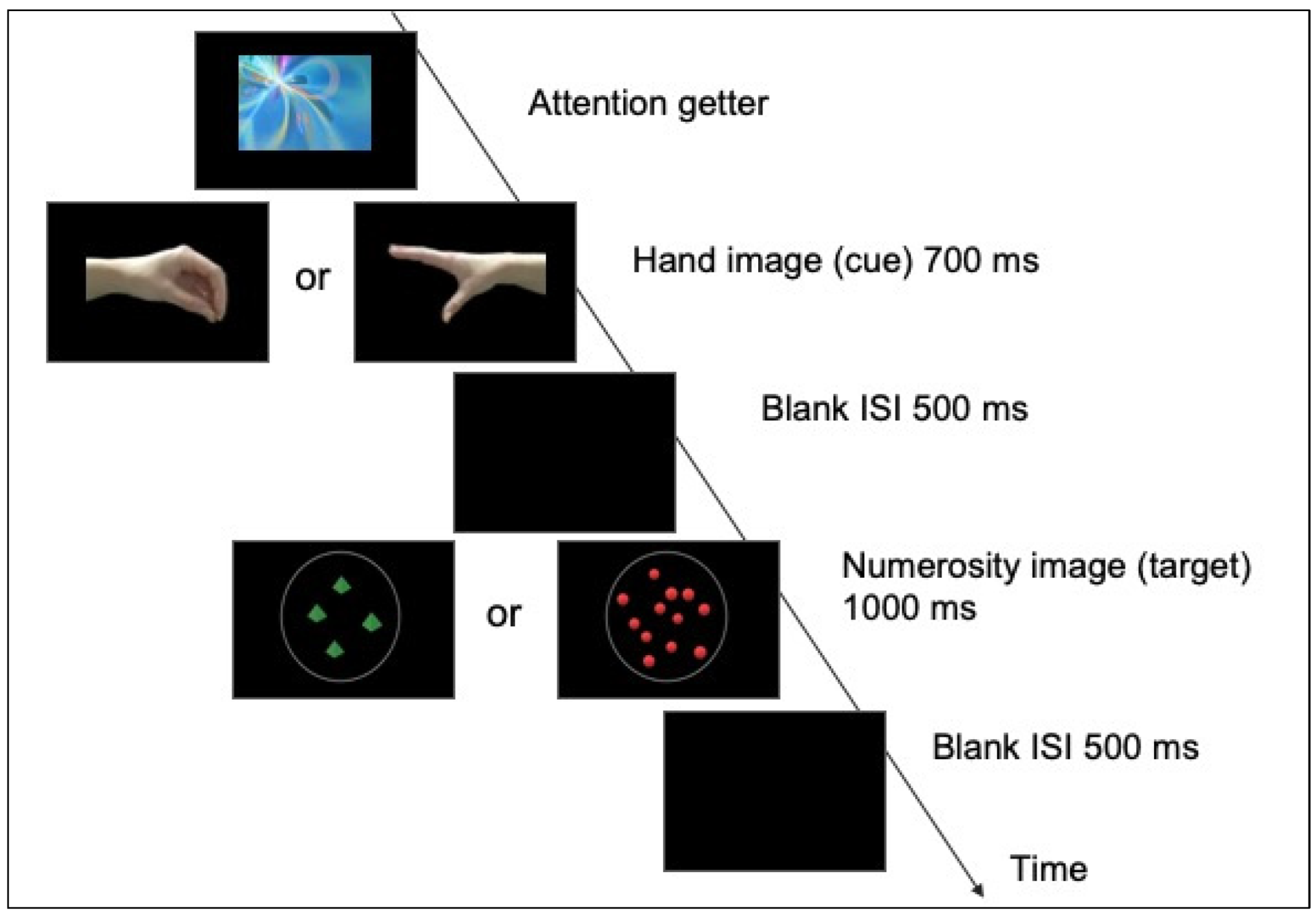
2.3. Procedure
2.4. EEG recording and Pre-Processing
2.5. Data and Statistical Analyses
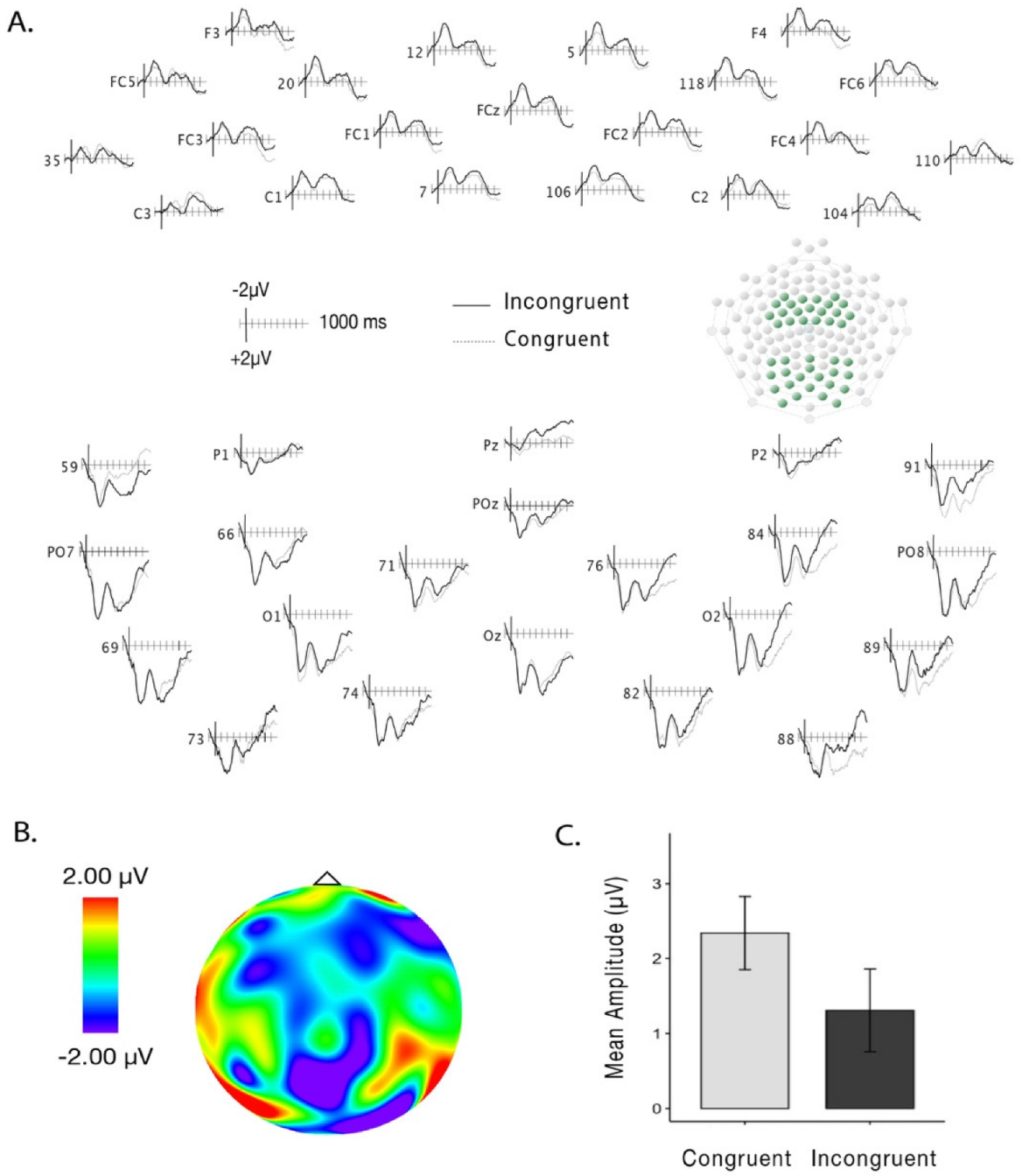
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Dehaene, S. The Number Sense: How the Mind Creates Mathematics, Revised and Updated Edition; Oxford University Press: New York, NY, USA, 2011; ISBN 978-0-19-975387-1. [Google Scholar]
- Izard, V.; Sann, C.; Spelke, E.S.; Streri, A. Newborn Infants Perceive Abstract Numbers. Proc. Natl. Acad. Sci. USA 2009, 106, 10382–10385. [Google Scholar] [CrossRef] [Green Version]
- Halberda, J.; Feigenson, L. Developmental Change in the Acuity of the ‘Number Sense’: The Approximate Number System in 3-, 4-, 5-, 6-Year-OLDS AND ADULTS 2008. Dev. Psychol. 2008, 44, 1457. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Libertus, M.E.; Brannon, E.M. Stable Individual Differences in Number Discrimination in Infancy. Dev. Sci. 2010, 13, 900–906. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, F.; Spelke, E.S. Large Number Discrimination in 6-Month-Old Infants. Cognition 2000, 74, B1–B11. [Google Scholar] [CrossRef]
- Schneider, M.; Thompson, C.A.; Rittle-Johnson, B. Associations of Magnitude Comparison and Number Line Estimation with Mathematical Competence: A Comparative Review. In Cognitive Development from a Strategy Perspective; Routledge: London, UK, 2017; ISBN 978-1-315-20044-6. [Google Scholar]
- Decarli, G.; Paris, E.; Tencati, C.; Nardelli, C.; Vescovi, M.; Surian, L.; Piazza, M. Impaired Large Numerosity Estimation and Intact Subitizing in Developmental Dyscalculia. PLoS ONE 2020, 15, e0244578. [Google Scholar] [CrossRef] [PubMed]
- Gallistel, C.R. Animal Cognition: The Representation of Space, Time and Number. Annu. Rev. Psychol. 1989, 40, 155–189. [Google Scholar] [CrossRef]
- De Corte, B.J.; Navarro, V.M.; Wasserman, E.A. Non-Cortical Magnitude Coding of Space and Time by Pigeons. Curr. Biol. 2017, 27, R1264–R1265. [Google Scholar] [CrossRef] [Green Version]
- Bortot, M.; Stancher, G.; Vallortigara, G. Transfer from Number to Size Reveals Abstract Coding of Magnitude in Honeybees. iScience 2020, 23, 101122. [Google Scholar] [CrossRef]
- Bueti, D.; Walsh, V. The Parietal Cortex and the Representation of Time, Space, Number and Other Magnitudes. Philos. Trans. R. Soc. B Biol. Sci. 2009, 364, 1831–1840. [Google Scholar] [CrossRef] [Green Version]
- Walsh, V. A Theory of Magnitude: Common Cortical Metrics of Time, Space and Quantity. Trends Cogn. Sci. 2003, 7, 483–488. [Google Scholar] [CrossRef]
- Walsh, V. A Theory of Magnitude: The Parts That Sum to Number. In The Oxford Handbook of Numerical Cognition; Oxford University Press: Oxford, UK, 2015; pp. 552–565. [Google Scholar]
- Anobile, G.; Arrighi, R.; Castaldi, E.; Burr, D.C. A Sensorimotor Numerosity System. Trends Cogn. Sci. 2021, 25, 24–36. [Google Scholar] [CrossRef] [PubMed]
- Ross, J. Visual Discrimination of Number without Counting. Perception 2003, 32, 867–870. [Google Scholar] [CrossRef]
- Whalen, J.; Gallistel, C.R.; Gelman, R. Nonverbal Counting in Humans: The Psychophysics of Number Representation. Psychol. Sci. 1999, 10, 130–137. [Google Scholar] [CrossRef]
- Hubbard, E.M.; Piazza, M.; Pinel, P.; Dehaene, S. Interactions between Number and Space in Parietal Cortex. Nat. Rev. Neurosci. 2005, 6, 435–448. [Google Scholar] [CrossRef] [PubMed]
- Simon, O.; Mangin, J.-F.; Cohen, L.; Le Bihan, D.; Dehaene, S. Topographical Layout of Hand, Eye, Calculation, and Language-Related Areas in the Human Parietal Lobe. Neuron 2002, 33, 475–487. [Google Scholar] [CrossRef] [Green Version]
- Arrighi, R.; Togoli, I.; Burr, D.C. A Generalized Sense of Number. Proc. R. Soc. B Biol. Sci. 2014, 281, 20141791. [Google Scholar] [CrossRef] [PubMed]
- Cicchini, G.M.; Anobile, G.; Burr, D.C. Spontaneous Perception of Numerosity in Humans. Nat. Commun. 2016, 7, 12536. [Google Scholar] [CrossRef] [Green Version]
- Wood, G.; Willmes, K.; Nuerk, H.-C.; Fischer, M.H. On the Cognitive Link between Space and Number: A Meta-Analysis of the SNARC Effect. Psychol. Sci. Q. 2008, 50, 489. [Google Scholar]
- Bulf, H.; de Hevia, M.D.; Macchi Cassia, V. Small on the Left, Large on the Right: Numbers Orient Visual Attention onto Space in Preverbal Infants. Dev. Sci. 2016, 19, 394–401. [Google Scholar] [CrossRef]
- de Hevia, M.D.; Veggiotti, L.; Streri, A.; Bonn, C.D. At Birth, Humans Associate “Few” with Left and “Many” with Right. Curr. Biol. 2017, 27, 3879–3884. [Google Scholar] [CrossRef]
- de Hevia, M.D.; Spelke, E.S. Number-Space Mapping in Human Infants. Psychol. Sci. 2010, 21, 653–660. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Di Giorgio, E.; Lunghi, M.; Rugani, R.; Regolin, L.; Dalla Barba, B.; Vallortigara, G.; Simion, F. A Mental Number Line in Human Newborns. Dev. Sci. 2019, 22, e12801. [Google Scholar] [CrossRef] [PubMed]
- de Hevia, M.D. How the Human Mind Grounds Numerical Quantities on Space. Child Dev. Perspect. 2021, 15, 44–50. [Google Scholar] [CrossRef]
- de Hevia, M.D.; Izard, V.; Coubart, A.; Spelke, E.S.; Streri, A. Representations of Space, Time, and Number in Neonates. Proc. Natl. Acad. Sci. USA 2014, 111, 4809–4813. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rugani, R.; Kelly, D.M.; Szelest, I.; Regolin, L.; Vallortigara, G. Is It Only Humans That Count from Left to Right? Biol. Lett. 2010, 6, 290–292. [Google Scholar] [CrossRef] [Green Version]
- Drucker, C.B.; Brannon, E.M. Rhesus Monkeys (Macaca Mulatta) Map Number onto Space. Cognition 2014, 132, 57–67. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Andres, M.; Davare, M.; Pesenti, M.; Olivier, E.; Seron, X. Number Magnitude and Grip Aperture Interaction. Neuroreport 2004, 15, 2773–2777. [Google Scholar]
- Andres, M.; Ostry, D.J.; Nicol, F.; Paus, T. Time Course of Number Magnitude Interference during Grasping. Cortex 2008, 44, 414–419. [Google Scholar] [CrossRef]
- Lindemann, O.; Abolafia, J.; Girardi, G.; Bekkering, H. Getting a Grip on Numbers: Numerical Magnitude Priming in Object Grasping. J. Exp. Psychol. Hum. Percept. Perform. 2008, 33, 1400–1409. [Google Scholar] [CrossRef] [Green Version]
- Badets, A.; Andres, M.; Di Luca, S.; Pesenti, M. Number Magnitude Potentiates Action Judgements. Exp. Brain Res. 2007, 180, 525–534. [Google Scholar] [CrossRef]
- Badets, A.; Bouquet, C.A.; Ric, F.; Pesenti, M. Number Generation Bias after Action Observation. Exp. Brain Res. 2012, 221, 43–49. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Badets, A.; Pesenti, M. Creating Number Semantics through Finger Movement Perception. Cognition 2010, 115, 46–53. [Google Scholar] [CrossRef] [PubMed]
- Ranzini, M.; Semenza, C.; Zorzi, M.; Cutini, S. Influences of Hand Action on the Processing of Symbolic Numbers: A Special Role of Pointing? PLoS ONE 2022, 17, e0269557. [Google Scholar] [CrossRef] [PubMed]
- Eger, E. Neuronal Foundations of Human Numerical Representations. Prog. Brain Res. 2016, 227, 1–27. [Google Scholar] [PubMed]
- Frey, S.H.; Vinton, D.; Norlund, R.; Grafton, S.T. Cortical Topography of Human Anterior Intraparietal Cortex Active during Visually Guided Grasping. Cogn. Brain Res. 2005, 23, 397–405. [Google Scholar] [CrossRef]
- Kadosh, R.C.; Henik, A.; Rubinsten, O.; Mohr, H.; Dori, H.; van de Ven, V.; Zorzi, M.; Hendler, T.; Goebel, R.; Linden, D.E.J. Are Numbers Special? Neuropsychologia 2005, 43, 1238–1248. [Google Scholar] [CrossRef]
- Sawamura, H.; Shima, K.; Tanji, J. Numerical Representation for Action in the Parietal Cortex of the Monkey. Nature 2002, 415, 918–922. [Google Scholar] [CrossRef]
- Andres, M.; Michaux, N.; Pesenti, M. Common Substrate for Mental Arithmetic and Finger Representation in the Parietal Cortex. Neuroimage 2012, 62, 1520–1528. [Google Scholar] [CrossRef] [PubMed]
- Sawamura, H.; Urgen, B.A.; Corbo, D.; Orban, G.A. A Parietal Region Processing Numerosity of Observed Actions: An FMRI Study. Eur. J. Neurosci. 2020, 52, 4732–4750. [Google Scholar] [CrossRef]
- Ranzini, M.; Scarpazza, C.; Radua, J.; Cutini, S.; Semenza, C.; Zorzi, M. A Common Neural Substrate for Number Comparison, Hand Reaching and Grasping: A SDM-PSI Meta-Analysis of Neuroimaging Studies. Cortex 2022, 148, 31–67. [Google Scholar] [CrossRef]
- Piazza, M.; Izard, V.; Pinel, P.; Le Bihan, D.; Dehaene, S. Tuning Curves for Approximate Numerosity in the Human Intraparietal Sulcus. Neuron 2004, 44, 547–555. [Google Scholar] [CrossRef] [PubMed]
- Cantlon, J.F.; Libertus, M.E.; Pinel, P.; Dehaene, S.; Brannon, E.M.; Pelphrey, K.A. The Neural Development of an Abstract Concept of Number. J. Cogn. Neurosci. 2009, 21, 2217–2229. [Google Scholar] [CrossRef]
- Hyde, D.C.; Boas, D.A.; Blair, C.; Carey, S. Near-Infrared Spectroscopy Shows Right Parietal Specialization for Number in Pre-Verbal Infants. Neuroimage 2010, 53, 647–652. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Izard, V.; Dehaene-Lambertz, G.; Dehaene, S. Distinct Cerebral Pathways for Object Identity and Number in Human Infants. PLoS Biol. 2008, 6, e11. [Google Scholar] [CrossRef] [PubMed]
- Wilcox, T.; Haslup, J.A.; Boas, D.A. Dissociation of Processing of Featural and Spatiotemporal Information in the Infant Cortex. Neuroimage 2010, 53, 1256–1263. [Google Scholar] [CrossRef] [Green Version]
- Rocha, N.A.C.F.; Silva, F.P.d.S.; Tudella, E. The Impact of Object Size and Rigidity on Infant Reaching. Infant Behav. Dev. 2006, 29, 251–261. [Google Scholar] [CrossRef]
- Libertus, K.; Gibson, J.; Hidayatallah, N.Z.; Hirtle, J.; Adcock, R.A.; Needham, A. Size Matters: How Age and Reaching Experiences Shape Infants’ Preferences for Different Sized Objects. Infant Behav. Dev. 2013, 36, 189–198. [Google Scholar] [CrossRef] [Green Version]
- Daum, M.M.; Vuori, M.T.; Prinz, W.; Aschersleben, G. Inferring the Size of a Goal Object from an Actor’s Grasping Movement in 6- and 9-Month-Old Infants. Dev. Sci. 2009, 12, 854–862. [Google Scholar] [CrossRef]
- Ambrosini, E.; Reddy, V.; de Looper, A.; Costantini, M.; Lopez, B.; Sinigaglia, C. Looking Ahead: Anticipatory Gaze and Motor Ability in Infancy. PLoS ONE 2013, 8, e67916. [Google Scholar] [CrossRef] [Green Version]
- Chinello, A.; Cattani, V.; Bonfiglioli, C.; Dehaene, S.; Piazza, M. Objects, Numbers, Fingers, Space: Clustering of Ventral and Dorsal Functions in Young Children and Adults. Dev. Sci. 2013, 16, 377–393. [Google Scholar] [CrossRef]
- Mareschal, D.; Johnson, M.H. The “What” and “Where” of Object Representations in Infancy. Cognition 2003, 88, 259–276. [Google Scholar] [CrossRef]
- Decarli, G.; Veggiotti, L.; de Hevia, M.D. The Link between Number and Action in Human Infants. Sci. Rep. 2022, 12, 3371. [Google Scholar] [CrossRef] [PubMed]
- Bettoni, R.; Addabbo, M.; Bulf, H.; Macchi Cassia, V. Electrophysiological Evidence of Space-Number Associations in 9-Month-Old Infants. Child Dev. 2021, 92, 2142–2152. [Google Scholar] [CrossRef] [PubMed]
- Lunghi, M.; Di Giorgio, E.; Benavides-Varela, S.; Simion, F. Covert Orienting of Attention in 3-Month-Old Infants: The Case of Biological Motion. Infant Behav. Dev. 2020, 58, 101422. [Google Scholar] [CrossRef]
- von Hofsten, C.; Rönnqvist, L. Preparation for Grasping an Object: A Developmental Study. J. Exp. Psychol. Hum. Percept. Perform. 1988, 14, 610–621. [Google Scholar] [CrossRef]
- Whyte, V.A.; McDonald, P.V.; Baillargeon, R.; Newell, K.M. Mouthing and Grasping of Objects by Young Infants. Ecol. Psychol. 1994, 6, 205–218. [Google Scholar] [CrossRef]
- Kutas, M.; Federmeier, K.D. Thirty Years and Counting: Finding Meaning in the N400 Component of the Event Related Brain Potential (ERP). Annu. Rev. Psychol. 2011, 62, 621–647. [Google Scholar] [CrossRef] [Green Version]
- Friedrich, M.; Friederici, A.D. Word Learning in 6-Month-Olds: Fast Encoding–Weak Retention. J. Cogn. Neurosci. 2011, 23, 3228–3240. [Google Scholar] [CrossRef]
- Parise, E.; Csibra, G. Electrophysiological Evidence for the Understanding of Maternal Speech by 9-Month-Old Infants. Psychol. Sci. 2012, 23, 728–733. [Google Scholar] [CrossRef] [Green Version]
- Rämä, P.; Sirri, L.; Serres, J. Development of Lexical–Semantic Language System: N400 Priming Effect for Spoken Words in 18- and 24-Month Old Children. Brain Lang. 2013, 125, 1–10. [Google Scholar] [CrossRef]
- Sirri, L.; Rämä, P. Cognitive and Neural Mechanisms Underlying Semantic Priming during Language Acquisition. J. Neurolinguistics 2015, 35, 1–12. [Google Scholar] [CrossRef]
- Torkildsen, J.v.K.; Syversen, G.; Simonsen, H.G.; Moen, I.; Lindgren, M. Electrophysiological Correlates of Auditory Semantic Priming in 24-Month-Olds. J. Neurolinguistics 2007, 20, 332–351. [Google Scholar] [CrossRef]
- Reid, V.M.; Hoehl, S.; Grigutsch, M.; Groendahl, A.; Parise, E.; Striano, T. The Neural Correlates of Infant and Adult Goal Prediction: Evidence for Semantic Processing Systems. Dev. Psychol. 2009, 45, 620–629. [Google Scholar] [CrossRef] [PubMed]
- Monroy, C.D.; Gerson, S.A.; Domínguez-Martínez, E.; Kaduk, K.; Hunnius, S.; Reid, V. Sensitivity to Structure in Action Sequences: An Infant Event-Related Potential Study. Neuropsychologia 2019, 126, 92–101. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reynolds, G.D.; Courage, M.L.; Richards, J.E. Infant Attention and Visual Preferences: Converging Evidence From Behavior, Event-Related Potentials, and Cortical Source Localization. Dev. Psychol. 2010, 46, 886–904. [Google Scholar] [CrossRef] [Green Version]
- Reynolds, G.D. Infant Visual Attention and Object Recognition. Behav. Brain Res. 2015, 285, 34–43. [Google Scholar] [CrossRef] [Green Version]
- Michel, C.; Kaduk, K.; Ní Choisdealbha, Á.; Reid, V.M. Event-Related Potentials Discriminate Familiar and Unusual Goal Outcomes in 5-Month-Olds and Adults. Dev. Psychol. 2017, 53, 1833–1843. [Google Scholar] [CrossRef]
- Craighero, L.; Leo, I.; Umiltà, C.; Simion, F. Newborns’ preference for goal-directed actions. Cognition. 2011, 120, 26–32. [Google Scholar] [CrossRef]
- Gratton, G.; Coles, M.G.H.; Donchin, E. A New Method for Off-Line Removal of Ocular Artifact. Electroencephalogr. Clin. Neurophysiol. 1983, 55, 468–484. [Google Scholar] [CrossRef]
- Hyde, D.C.; Porter, C.L.; Flom, R.; Stone, S.A. Relational Congruence Facilitates Neural Mapping of Spatial and Temporal Magnitudes in Preverbal Infants. Dev. Cogn. Neurosci. 2013, 6, 102–112. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2017. [Google Scholar]
- Rugani, R.; Betti, S.; Ceccarini, F.; Sartori, L. Act on Numbers: Numerical Magnitude Influences Selection and Kinematics of Finger Movement. Front. Psychol. 2017, 8, 1481. [Google Scholar] [CrossRef] [PubMed]
- Rugani, R.; Sartori, L. Numbers in Action. Front. Hum. Neurosci. 2016, 10, 388. [Google Scholar] [CrossRef] [PubMed]
- Karrer, R.; Monti, L.A. Event-Related Potentials of 4–7-Week-Old Infants in a Visual Recognition Memory Task. Electroencephalogr. Clin. Neurophysiol. 1995, 94, 414–424. [Google Scholar] [CrossRef]
- Mills, D.L.; Coffey-Corina, S.; Neville, H.J. Language Comprehension and Cerebral Specialization from 13 to 20 Months. Dev. Neuropsychol. 1997, 13, 397–445. [Google Scholar] [CrossRef]


Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Decarli, G.; Rämä, P.; Granjon, L.; Veggiotti, L.; de Hevia, M.D. Electrophysiological Evidence for A Number–Action Mapping in Infancy. Brain Sci. 2022, 12, 1480. https://doi.org/10.3390/brainsci12111480
Decarli G, Rämä P, Granjon L, Veggiotti L, de Hevia MD. Electrophysiological Evidence for A Number–Action Mapping in Infancy. Brain Sciences. 2022; 12(11):1480. https://doi.org/10.3390/brainsci12111480
Chicago/Turabian StyleDecarli, Gisella, Pia Rämä, Lionel Granjon, Ludovica Veggiotti, and Maria Dolores de Hevia. 2022. "Electrophysiological Evidence for A Number–Action Mapping in Infancy" Brain Sciences 12, no. 11: 1480. https://doi.org/10.3390/brainsci12111480
APA StyleDecarli, G., Rämä, P., Granjon, L., Veggiotti, L., & de Hevia, M. D. (2022). Electrophysiological Evidence for A Number–Action Mapping in Infancy. Brain Sciences, 12(11), 1480. https://doi.org/10.3390/brainsci12111480