

Surface-Based Cortical Measures in Multimodal Association Brain Regions Predict Chess Expertise

 , , , and
, , , and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Participants
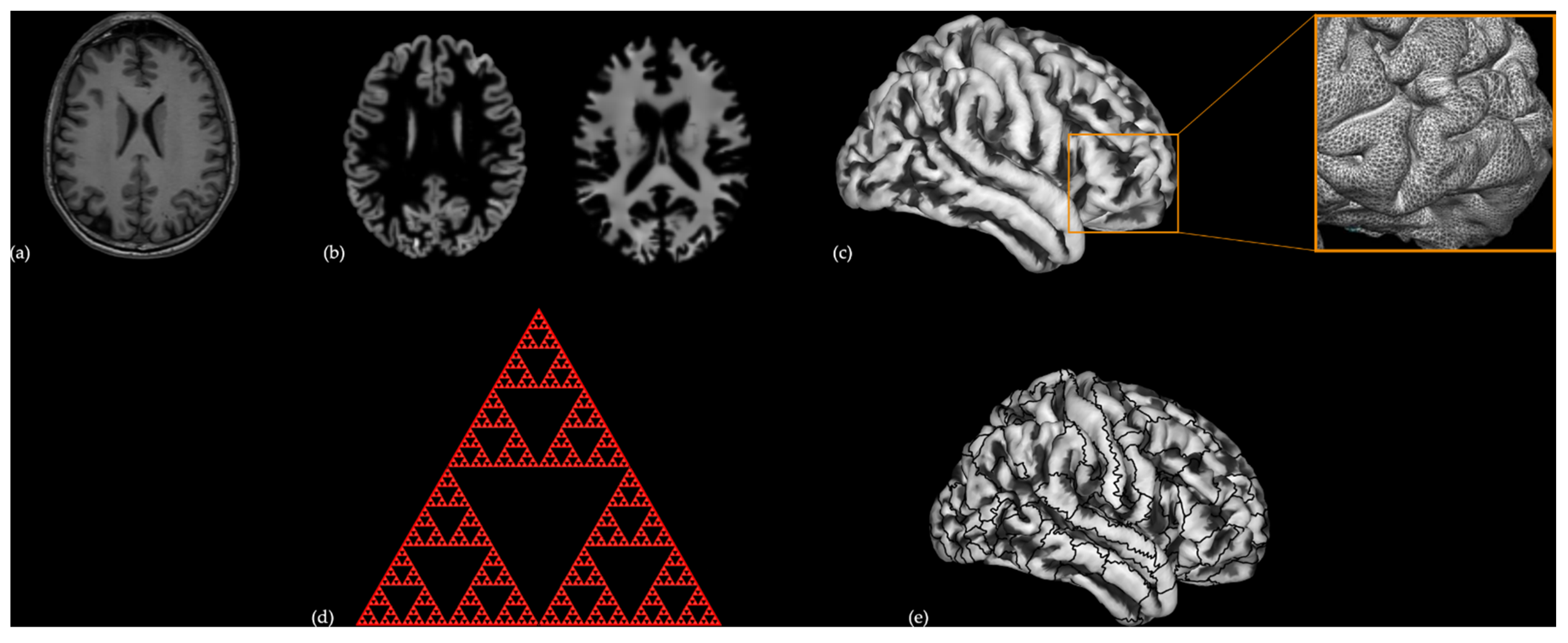
2.2. Brain Imaging
2.2.1. Data Acquisition
2.2.2. Pre-Processing
2.3. Statistical Analysis
2.3.1. Bivariate Analysis
2.3.2. Multivariate Analysis
3. Results
3.1. Cortical Complexity Assessed by FD
3.1.1. Bivariate Comparison
3.1.2. Correlations with Chess-Related Features in Chess Masters
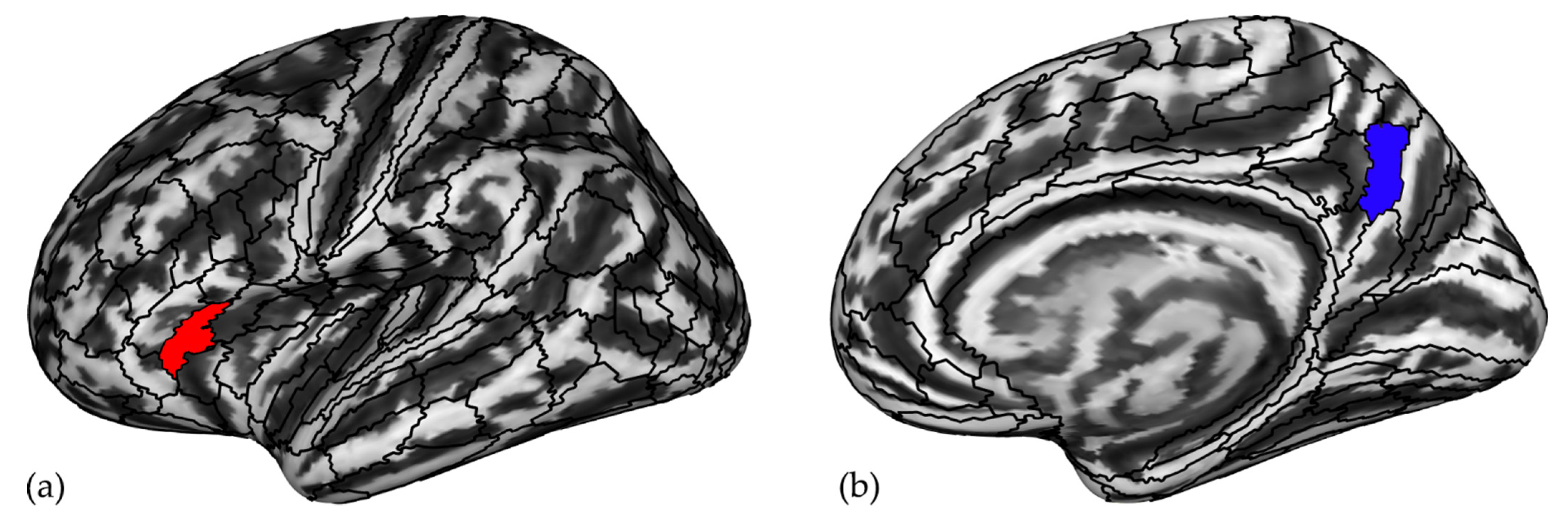
3.2. Regions Associated with Chess Expertise
3.3. Gyrification Index
3.4. Cortical Thickness
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Charness, N. The Impact of Chess Research on Cognitive Science. Psychol. Res. 1992, 54, 4–9. [Google Scholar] [CrossRef]
- Atherton, M.; Zhuang, J.; Bart, W.M.; Hu, X.; He, S. A Functional MRI Study of High-Level Cognition. I. The Game of Chess. Cogn. Brain Res. 2003, 16, 26–31. [Google Scholar] [CrossRef]
- Grabner, R.H.; Stern, E.; Neubauer, A.C. Individual Differences in Chess Expertise: A Psychometric Investigation. Acta Psychol. 2007, 124, 398–420. [Google Scholar] [CrossRef] [PubMed]
- Kelly, A.M.C.; Garavan, H. Human Functional Neuroimaging of Brain Changes Associated with Practice. Cereb. Cortex 2005, 15, 1089–1102. [Google Scholar] [CrossRef] [Green Version]
- Simon, H.; Chase, W. Skill in Chess. In Computer Chess Compendium; Levy, D., Ed.; Springer: New York, NY, USA, 1988; pp. 175–188. ISBN 9781475719703. [Google Scholar]
- Burgoyne, A.P.; Sala, G.; Gobet, F.; Macnamara, B.N.; Campitelli, G.; Hambrick, D.Z. The Relationship between Cognitive Ability and Chess Skill: A Comprehensive Meta-Analysis. Intelligence 2016, 59, 72–83. [Google Scholar] [CrossRef] [Green Version]
- Duan, X.; He, S.; Liao, W.; Liang, D.; Qiu, L.; Wei, L.; Li, Y.; Liu, C.; Gong, Q.; Chen, H. Reduced Caudate Volume and Enhanced Striatal-DMN Integration in Chess Experts. NeuroImage 2012, 60, 1280–1286. [Google Scholar] [CrossRef]
- Ouellette, D.J.; Hsu, D.-L.; Stefancin, P.; Duong, T.Q. Cortical Thickness and Functional Connectivity Changes in Chinese Chess Experts. PLoS ONE 2020, 15, e0239822. [Google Scholar] [CrossRef]
- Hänggi, J.; Brütsch, K.; Siegel, A.M.; Jäncke, L. The Architecture of the Chess Player’s Brain. Neuropsychologia 2014, 62, 152–162. [Google Scholar] [CrossRef]
- Mayeli, M.; Rahmani, F.; Aarabi, M.H. Comprehensive Investigation of White Matter Tracts in Professional Chess Players and Relation to Expertise: Region of Interest and DMRI Connectometry. Front. Neurosci. 2018, 12, 288. [Google Scholar] [CrossRef] [Green Version]
- Ghosh, S.S.; Kakunoori, S.; Augustinack, J.; Nieto-Castanon, A.; Kovelman, I.; Gaab, N.; Christodoulou, J.A.; Triantafyllou, C.; Gabrieli, J.D.E.; Fischl, B. Evaluating the Validity of Volume-Based and Surface-Based Brain Image Registration for Developmental Cognitive Neuroscience Studies in Children 4 to 11 Years of Age. Neuroimage 2010, 53, 85–93. [Google Scholar] [CrossRef]
- Scarpazza, C.; Tognin, S.; Frisciata, S.; Sartori, G.; Mechelli, A. False Positive Rates in Voxel-Based Morphometry Studies of the Human Brain: Should We Be Worried? Neurosci. Biobehav. Rev. 2015, 52, 49–55. [Google Scholar] [CrossRef] [PubMed]
- Goto, M.; Abe, O.; Hagiwara, A.; Fujita, S.; Kamagata, K.; Hori, M.; Aoki, S.; Osada, T.; Konishi, S.; Masutani, Y.; et al. Advantages of Using Both Voxel- and Surface-Based Morphometry in Cortical Morphology Analysis: A Review of Various Applications. Magn. Reson. Med. Sci. 2022, 21, 41–57. [Google Scholar] [CrossRef] [PubMed]
- Gregory, M.D.; Kippenhan, J.S.; Dickinson, D.; Carrasco, J.; Mattay, V.S.; Weinberger, D.R.; Berman, K.F. Regional Variations in Brain Gyrification Are Associated with General Cognitive Ability in Humans. Curr. Biol. 2016, 26, 1301–1305. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tadayon, E.; Pascual-Leone, A.; Santarnecchi, E. Differential Contribution of Cortical Thickness, Surface Area, and Gyrification to Fluid and Crystallized Intelligence. Cereb. Cortex 2020, 30, 215–225. [Google Scholar] [CrossRef]
- Jin, K.; Zhang, T.; Shaw, M.; Sachdev, P.; Cherbuin, N. Relationship Between Sulcal Characteristics and Brain Aging. Front Aging Neurosci. 2018, 10, 339. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, T.; Wen, W.; Zhu, W.; Kochan, N.A.; Trollor, J.N.; Reppermund, S.; Jin, J.S.; Luo, S.; Brodaty, H.; Sachdev, P.S. The Relationship between Cortical Sulcal Variability and Cognitive Performance in the Elderly. Neuroimage 2011, 56, 865–873. [Google Scholar] [CrossRef]
- Hofman, M. A The Fractal Geometry of Convoluted Brains. J. Hirnforsch. 1991, 32, 103–111. [Google Scholar]
- Jovanov-Milošević, N.; Petanjek, Z.; Petrović, D.; Judaš, M.; Kostović, I. Morphology, Molecular Phenotypes and Distribution of Neurons in Developing Human Corpus Callosum: Development of Human Intracallosal Neurons. Eur. J. Neurosci. 2010, 32, 1423–1432. [Google Scholar] [CrossRef]
- Losa, G.A.; Di Ieva, A.; Grizzi, F.; de Vico, G. On the Fractal Nature of Nervous Cell System. Front. Neuroanat. 2011, 5, 45. [Google Scholar] [CrossRef] [Green Version]
- Smith, T.G.; Lange, G.D.; Marks, W.B. Fractal Methods and Results in Cellular Morphology—Dimensions, Lacunarity and Multifractals. J. Neurosci. Methods 1996, 69, 123–136. [Google Scholar] [CrossRef] [Green Version]
- Zhang, L.; Liu, J.Z.; Dean, D.; Sahgal, V.; Yue, G.H. A Three-Dimensional Fractal Analysis Method for Quantifying White Matter Structure in Human Brain. J. Neurosci. Methods 2006, 150, 242–253. [Google Scholar] [CrossRef] [PubMed]
- Yotter, R.A.; Nenadic, I.; Ziegler, G.; Thompson, P.M.; Gaser, C. Local Cortical Surface Complexity Maps from Spherical Harmonic Reconstructions. Neuroimage 2011, 56, 961–973. [Google Scholar] [CrossRef] [PubMed]
- Caserta, F.; Eldred, W.D.; Fernandez, E.; Hausman, R.E.; Stanford, L.R.; Bulderev, S.V.; Schwarzer, S.; Stanley, H.E. Determination of Fractal Dimension of Physiologically Characterized Neurons in Two and Three Dimensions. J. Neurosci. Methods 1995, 56, 133–144. [Google Scholar] [CrossRef] [Green Version]
- Bernard, F.; Bossu, J.-L.; Gaillard, S. Identification of Living Oligodendrocyte Developmental Stages by Fractal Analysis of Cell Morphology. J. Neurosci. Res. 2001, 65, 439–445. [Google Scholar] [CrossRef] [PubMed]
- Franconeri, S.L.; Alvarez, G.A.; Cavanagh, P. Flexible Cognitive Resources: Competitive Content Maps for Attention and Memory. Trends Cogn. Sci. 2013, 17, 134–141. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- White, T.; Su, S.; Schmidt, M.; Kao, C.-Y.; Sapiro, G. The Development of Gyrification in Childhood and Adolescence. Brain Cogn. 2010, 72, 36–45. [Google Scholar] [CrossRef] [Green Version]
- Shyu, K.-K.; Wu, Y.-T.; Chen, T.-R.; Chen, H.-Y.; Hu, H.-H.; Guo, W.-Y. Analysis of Fetal Cortical Complexity from MR Images Using 3D Entropy Based Information Fractal Dimension. Nonlinear Dyn. 2010, 61, 363–372. [Google Scholar] [CrossRef]
- Takahashi, T.; Murata, T.; Omori, M.; Kosaka, H.; Takahashi, K.; Yonekura, Y.; Wada, Y. Quantitative Evaluation of Age-Related White Matter Microstructural Changes on MRI by Multifractal Analysis. J. Neurol. Sci. 2004, 225, 33–37. [Google Scholar] [CrossRef]
- Meregalli, V.; Alberti, F.; Madan, C.R.; Meneguzzo, P.; Miola, A.; Trevisan, N.; Sambataro, F.; Favaro, A.; Collantoni, E. Cortical Complexity Estimation Using Fractal Dimension: A Systematic Review of the Literature on Clinical and Nonclinical Samples. Eur. J. Neurosci. 2022, 55, 1547–1583. [Google Scholar] [CrossRef]
- Trevisan, N.; Miola, A.; Cattarinussi, G.; Kubera, K.M.; Hirjak, D.; Wolf, R.C.; Sambataro, F. Cortical Folding Complexity Is Distinctively Altered in Schizophrenia and Bipolar Disorder. Schizophr. Res. 2022, 241, 92–93. [Google Scholar] [CrossRef]
- Li, K.; Jiang, J.; Qiu, L.; Yang, X.; Huang, X.; Lui, S.; Gong, Q. A Multimodal MRI Dataset of Professional Chess Players. Sci. Data 2015, 2, 150044. [Google Scholar] [CrossRef]
- Ashburner, J. A Fast Diffeomorphic Image Registration Algorithm. Neuroimage 2007, 38, 95–113. [Google Scholar] [CrossRef] [PubMed]
- Dahnke, R.; Yotter, R.A.; Gaser, C. Cortical Thickness and Central Surface Estimation. Neuroimage 2013, 65, 336–348. [Google Scholar] [CrossRef] [PubMed]
- Glasser, M.F.; Coalson, T.S.; Robinson, E.C.; Hacker, C.D.; Harwell, J.; Yacoub, E.; Ugurbil, K.; Andersson, J.; Beckmann, C.F.; Jenkinson, M.; et al. A Multi-Modal Parcellation of Human Cerebral Cortex. Nature 2016, 536, 171–178. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lotze, M.; Scheler, G.; Tan, H.-R.M.; Braun, C.; Birbaumer, N. The Musician’s Brain: Functional Imaging of Amateurs and Professionals during Performance and Imagery. NeuroImage 2003, 20, 1817–1829. [Google Scholar] [CrossRef] [PubMed]
- Baumgaertner, A.; Buccino, G.; Lange, R.; McNamara, A.; Binkofski, F. Polymodal Conceptual Processing of Human Biological Actions in the Left Inferior Frontal Lobe: Conceptual Processing of Biological Hand Actions. Eur. J. Neurosci. 2007, 25, 881–889. [Google Scholar] [CrossRef]
- Bunge, S.A. How We Use Rules to Select Actions: A Review of Evidence from Cognitive Neuroscience. Cogn. Affect. Behav. Neurosci. 2004, 4, 564–579. [Google Scholar] [CrossRef] [Green Version]
- Kan, I.P.; Thompson-Schill, S.L. Effect of Name Agreement on Prefrontal Activity during Overt and Covert Picture Naming. Cogn. Affect. Behav. Neurosci. 2004, 4, 43–57. [Google Scholar] [CrossRef] [Green Version]
- León-Domínguez, U.; Martín-Rodríguez, J.F.; León-Carrión, J. Executive N-Back Tasks for the Neuropsychological Assessment of Working Memory. Behav. Brain Res. 2015, 292, 167–173. [Google Scholar] [CrossRef]
- Braver, T.S.; Barch, D.M. Extracting Core Components of Cognitive Control. Trends Cogn. Sci. 2006, 10, 529–532. [Google Scholar] [CrossRef]
- Assem, M.; Blank, I.A.; Mineroff, Z.; Ademoğlu, A.; Fedorenko, E. Activity in the Fronto-Parietal Multiple-Demand Network Is Robustly Associated with Individual Differences in Working Memory and Fluid Intelligence. Cortex 2020, 131, 1–16. [Google Scholar] [CrossRef] [PubMed]
- Zatorre, R.J.; Fields, R.D.; Johansen-Berg, H. Plasticity in Gray and White: Neuroimaging Changes in Brain Structure during Learning. Nat. Neurosci. 2012, 15, 528–536. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sydnor, V.J.; Larsen, B.; Bassett, D.S.; Alexander-Bloch, A.; Fair, D.A.; Liston, C.; Mackey, A.P.; Milham, M.P.; Pines, A.; Roalf, D.R.; et al. Neurodevelopment of the Association Cortices: Patterns, Mechanisms, and Implications for Psychopathology. Neuron 2021, 109, 2820–2846. [Google Scholar] [CrossRef] [PubMed]
- Caspari, N.; Arsenault, J.T.; Vandenberghe, R.; Vanduffel, W. Functional Similarity of Medial Superior Parietal Areas for Shift-Selective Attention Signals in Humans and Monkeys. Cereb. Cortex 2018, 28, 2085–2099. [Google Scholar] [CrossRef] [Green Version]
- Gogos, A.; Gavrilescu, M.; Davison, S.; Searle, K.; Adams, J.; Rossell, S.L.; Bell, R.; Davis, S.R.; Egan, G.F. Greater Superior than Inferior Parietal Lobule Activation with Increasing Rotation Angle during Mental Rotation: An FMRI Study. Neuropsychologia 2010, 48, 529–535. [Google Scholar] [CrossRef]
- Molenberghs, P.; Mesulam, M.M.; Peeters, R.; Vandenberghe, R.R.C. Remapping Attentional Priorities: Differential Contribution of Superior Parietal Lobule and Intraparietal Sulcus. Cereb. Cortex 2007, 17, 2703–2712. [Google Scholar] [CrossRef] [Green Version]
- Wolpert, D.M.; Goodbody, S.J.; Husain, M. Maintaining Internal Representations: The Role of the Human Superior Parietal Lobe. Nat. Neurosci. 1998, 1, 529–533. [Google Scholar] [CrossRef]
- Wang, J.; Yang, Y.; Fan, L.; Xu, J.; Li, C.; Liu, Y.; Fox, P.T.; Eickhoff, S.B.; Yu, C.; Jiang, T. Convergent Functional Architecture of the Superior Parietal Lobule Unraveled with Multimodal Neuroimaging Approaches: Parcellation of Superior Parietal Lobule. Hum. Brain Mapp. 2015, 36, 238–257. [Google Scholar] [CrossRef] [Green Version]
- Koenigs, M.; Barbey, A.K.; Postle, B.R.; Grafman, J. Superior Parietal Cortex Is Critical for the Manipulation of Information in Working Memory. J. Neurosci. 2009, 29, 14980–14986. [Google Scholar] [CrossRef] [Green Version]
- Cavanna, A.E.; Trimble, M.R. The Precuneus: A Review of Its Functional Anatomy and Behavioural Correlates. Brain 2006, 129, 564–583. [Google Scholar] [CrossRef] [Green Version]
- Zimmermann, M.; Kubik, V.; Persson, J.; Mäntylä, T. Monitoring Multiple Deadlines Relies on Spatial Processing in Posterior Parietal Cortex. J. Cogn. Neurosci. 2019, 31, 1468–1483. [Google Scholar] [CrossRef] [PubMed]
- Seitz, R.J.; Binkofski, F. Modular Organization of Parietal Lobe Functions as Revealed by Functional Activation Studies. Adv. Neurol. 2003, 93, 281–292. [Google Scholar]
- Grefkes, C.; Ritzl, A.; Zilles, K.; Fink, G.R. Human Medial Intraparietal Cortex Subserves Visuomotor Coordinate Transformation. NeuroImage 2004, 23, 1494–1506. [Google Scholar] [CrossRef] [PubMed]
- Borra, E.; Gerbella, M.; Rozzi, S.; Luppino, G. The Macaque Lateral Grasping Network: A Neural Substrate for Generating Purposeful Hand Actions. Neurosci. Biobehav. Rev. 2017, 75, 65–90. [Google Scholar] [CrossRef]
- Seeley, W.W.; Menon, V.; Schatzberg, A.F.; Keller, J.; Glover, G.H.; Kenna, H.; Reiss, A.L.; Greicius, M.D. Dissociable Intrinsic Connectivity Networks for Salience Processing and Executive Control. J. Neurosci. 2007, 27, 2349–2356. [Google Scholar] [CrossRef]
- Song, L.; Peng, Q.; Liu, S.; Wang, J. Changed Hub and Functional Connectivity Patterns of the Posterior Fusiform Gyrus in Chess Experts. Brain Imaging Behav. 2020, 14, 797–805. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Zuo, C.; Wang, D.; Tao, S.; Hao, L. Reduced Thalamus Volume and Enhanced Thalamus and Fronto-Parietal Network Integration in the Chess Experts. Cereb. Cortex 2020, 30, 5560–5569. [Google Scholar] [CrossRef]
- Aminoff, E.M.; Kveraga, K.; Bar, M. The Role of the Parahippocampal Cortex in Cognition. Trends Cogn. Sci. 2013, 17, 379–390. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bilalić, M.; Langner, R.; Erb, M.; Grodd, W. Mechanisms and Neural Basis of Object and Pattern Recognition: A Study with Chess Experts. J. Exp. Psychol. Gen. 2010, 139, 728–742. [Google Scholar] [CrossRef] [Green Version]
- Rilling, J.K.; Sanfey, A.G.; Aronson, J.A.; Nystrom, L.E.; Cohen, J.D. The Neural Correlates of Theory of Mind within Interpersonal Interactions. NeuroImage 2004, 22, 1694–1703. [Google Scholar] [CrossRef]
- Tettamanti, M.; Vaghi, M.M.; Bara, B.G.; Cappa, S.F.; Enrici, I.; Adenzato, M. Effective Connectivity Gateways to the Theory of Mind Network in Processing Communicative Intention. NeuroImage 2017, 155, 169–176. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lissek, S.; Peters, S.; Fuchs, N.; Witthaus, H.; Nicolas, V.; Tegenthoff, M.; Juckel, G.; Brüne, M. Cooperation and Deception Recruit Different Subsets of the Theory-of-Mind Network. PLoS ONE 2008, 3, e2023. [Google Scholar] [CrossRef]
- Baker, C.M.; Burks, J.D.; Briggs, R.G.; Stafford, J.; Conner, A.K.; Glenn, C.A.; Sughrue, M.E. A Connectomic Atlas of the Human Cerebrum—Chapter 4: The Medial Frontal Lobe, Anterior Cingulate Gyrus, and Orbitofrontal Cortex. Oper. Neurosurg. 2018, 15 (Suppl. 1), S122–S174. [Google Scholar] [CrossRef] [PubMed]
- Blanton, R.E.; Levitt, J.G.; Thompson, P.M.; Narr, K.L.; Capetillo-Cunliffe, L.; Nobel, A.; Singerman, J.D.; McCracken, J.T.; Toga, A.W. Mapping Cortical Asymmetry and Complexity Patterns in Normal Children. Psychiatry Res. Neuroimaging 2001, 107, 29–43. [Google Scholar] [CrossRef]
- Premi, E.; Gazzina, S.; Diano, M.; Girelli, A.; Calhoun, V.D.; Iraji, A.; Gong, Q.; Li, K.; Cauda, F.; Gasparotti, R.; et al. Enhanced Dynamic Functional Connectivity (Whole-Brain Chronnectome) in Chess Experts. Sci. Rep. 2020, 10, 7051. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Professional Chess Players (N = 29) | Novices (N = 29) | p-Values * | |
|---|---|---|---|
| Age: mean (SD) | 28.72 (10.84) | 25.76 (6.95) | 0.22 |
| Sex: females (%) | 9 (31.03%) | 15 (51.72%) | 0.11 |
| Education: years (SD) | 13.27 (2.79) | 13.92 (3.15) | 0.41 |
| Elo rank: mean (SD) | 2401.1 (134.6) | - | - |
| Age at which they started professional training: mean years (SD) | 17 (5.8) | - | - |
| Duration of daily training: mean hours (SD) | 4.12 (1.79) | - | - |
| Predictors with Bootstrap | ||||||
|---|---|---|---|---|---|---|
| B | Bias | SE | p | 95% CI | ||
| Lower | Upper | |||||
| Age (years) | −0.10 | −5.98 | 34.31 | 0.025 | −52.27 | 1.27 |
| Sex (male) | 1.01 | 61.15 | 561.57 | 0.317 | −135.33 | 595.34 |
| Education | −0.24 | −13.81 | 107.35 | 0.100 | −118.85 | 16.58 |
| ROIs | ||||||
| Left FOP5 | 11.07 | 485.79 | 2869.57 | 0.001 | 5.94 | 3844.66 |
| Left PFt | −7.25 | −412.05 | 2380.84 | 0.000 | −3538.41 | −4.18 |
| Left 8BM | −16.41 | −698.42 | 3863.81 | 0.001 | −5819.85 | −9.42 |
| Right TF | −8.92 | −404.94 | 2514.75 | 0.004 | −3164.28 | −3.45 |
| Right 7 m | −7.67 | −214.67 | 1686.45 | 0.002 | −1616.48 | 63.18 |
| Intercept | 74.24 | 3258.26 | 18,208.48 | 0.000 | 51.04 | 26,353.63 |
| Classification table | ||||||
| Predicted | ||||||
| Observed | Novices | Professional chess players | Correct % | |||
| Novices | 26 | 3 | 89.7% | |||
| Professional chess masters | 1 | 28 | 96.6% | |||
| Overall percentage | 93.1% | |||||
| Model fit | Hosmer and Lemeshow test | |||||
| Nagelkerke R2 | Chi-2 | p | ||||
| 0.793 | 5.724 | 0.678 | ||||
| Predictors with Bootstrap | ||||||
|---|---|---|---|---|---|---|
| Predictors | B | Bias | SE | p | 95% CI | |
| Lower | Upper | |||||
| Education | 1.04 | 0.19 | 0.95 | 0.106 | −0.27 | 3.00 |
| Age (years) | 0.04 | 0.01 | 0.07 | 0.367 | −0.05 | 0.18 |
| Male/female | −0.10 | −0.01 | 0.19 | 0.441 | −0.48 | 0.24 |
| ROIs | ||||||
| Right a24pr | −0.37 | −0.08 | 0.26 | 0.007 | −0.95 | −0.09 |
| Right STSdp | 0.55 | 0.14 | 0.44 | 0.007 | 0.20 | 1.46 |
| Intercept | −5.64 | −2.13 | 13.46 | 0.503 | −34.00 | 11.46 |
| Classification table | ||||||
| Predicted | ||||||
| Observed | Novices | Professional chess masters | Correct % | |||
| Novices | 21 | 8 | 72.4% | |||
| Professional chess masters | 10 | 19 | 65.5% | |||
| Overall percentage | 69.0% | |||||
| Model fit | Hosmer and Lemeshow test | |||||
| Nagelkerke R2 | Chi-2 | p | ||||
| 0.359 | 7.030 | 0.533 | ||||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Trevisan, N.; Jaillard, A.; Cattarinussi, G.; De Roni, P.; Sambataro, F. Surface-Based Cortical Measures in Multimodal Association Brain Regions Predict Chess Expertise. Brain Sci. 2022, 12, 1592. https://doi.org/10.3390/brainsci12111592
Trevisan N, Jaillard A, Cattarinussi G, De Roni P, Sambataro F. Surface-Based Cortical Measures in Multimodal Association Brain Regions Predict Chess Expertise. Brain Sciences. 2022; 12(11):1592. https://doi.org/10.3390/brainsci12111592
Chicago/Turabian StyleTrevisan, Nicolò, Assia Jaillard, Giulia Cattarinussi, Prisca De Roni, and Fabio Sambataro. 2022. "Surface-Based Cortical Measures in Multimodal Association Brain Regions Predict Chess Expertise" Brain Sciences 12, no. 11: 1592. https://doi.org/10.3390/brainsci12111592
APA StyleTrevisan, N., Jaillard, A., Cattarinussi, G., De Roni, P., & Sambataro, F. (2022). Surface-Based Cortical Measures in Multimodal Association Brain Regions Predict Chess Expertise. Brain Sciences, 12(11), 1592. https://doi.org/10.3390/brainsci12111592