
Assessing Anti-Social and Aggressive Behavior in a Zebrafish (Danio rerio) Model of Parkinson’s Disease Chronically Exposed to Rotenone

 , , ,
, , ,  , , ,
, , , 

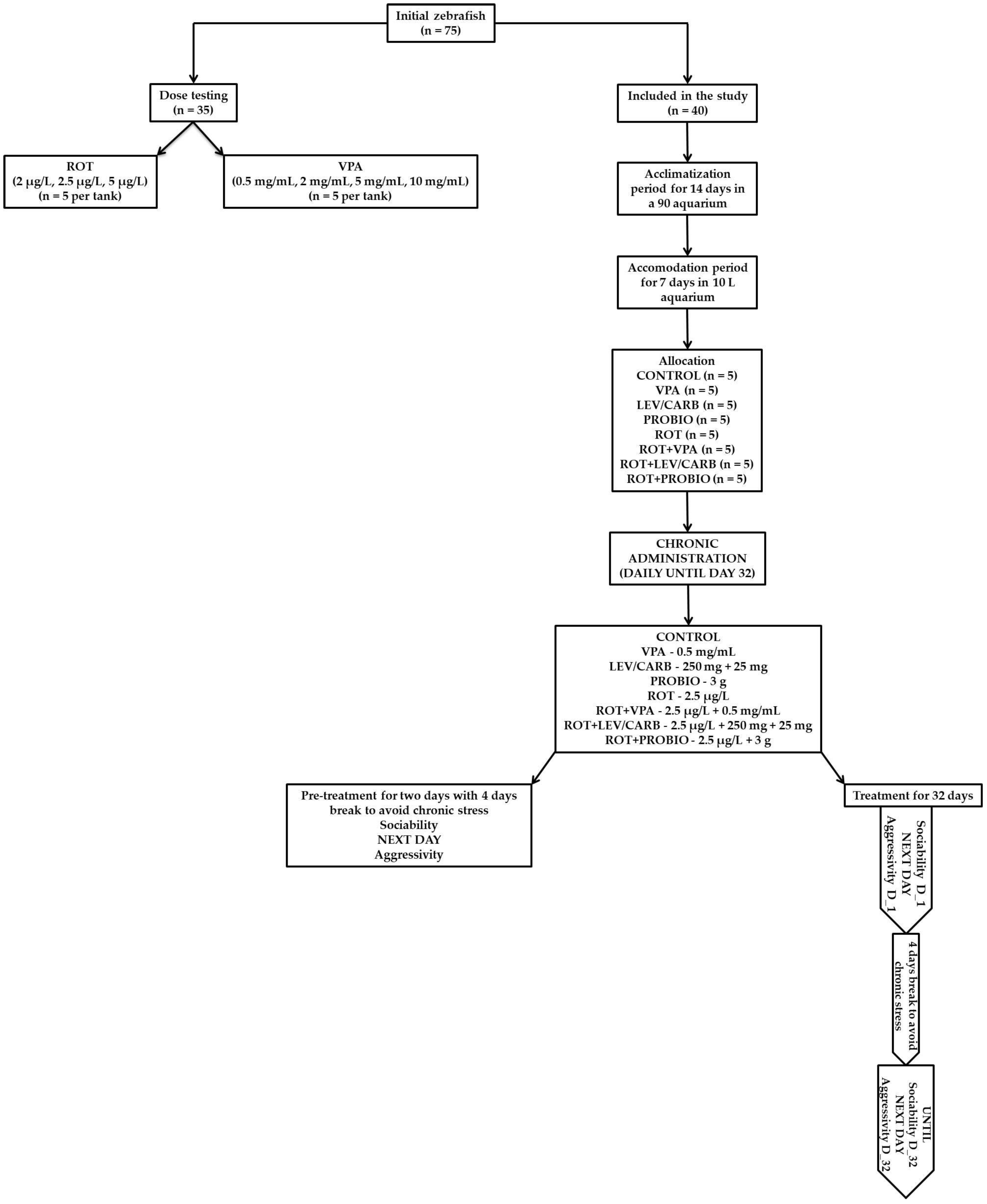{kind=link}
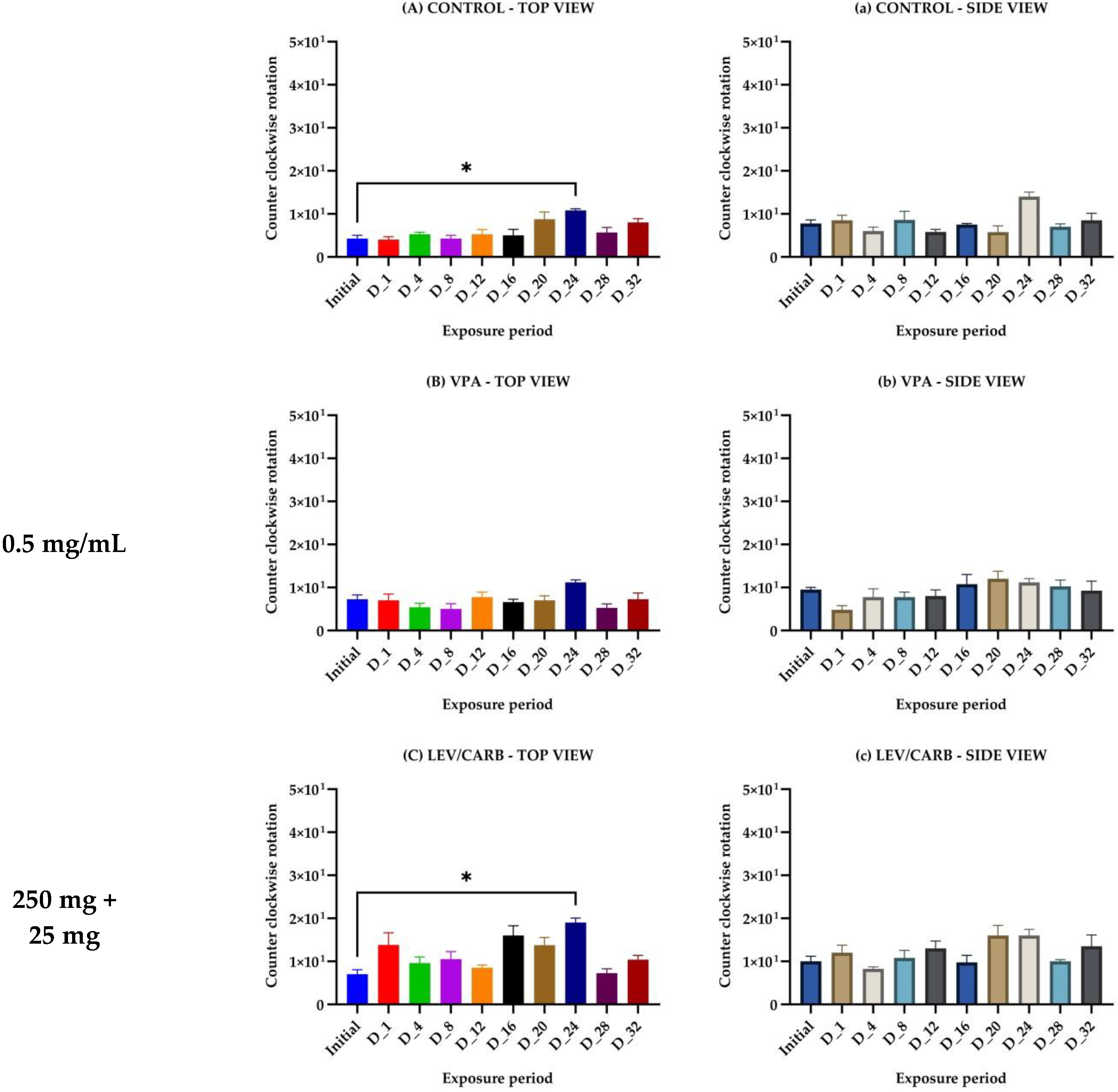{kind=link}
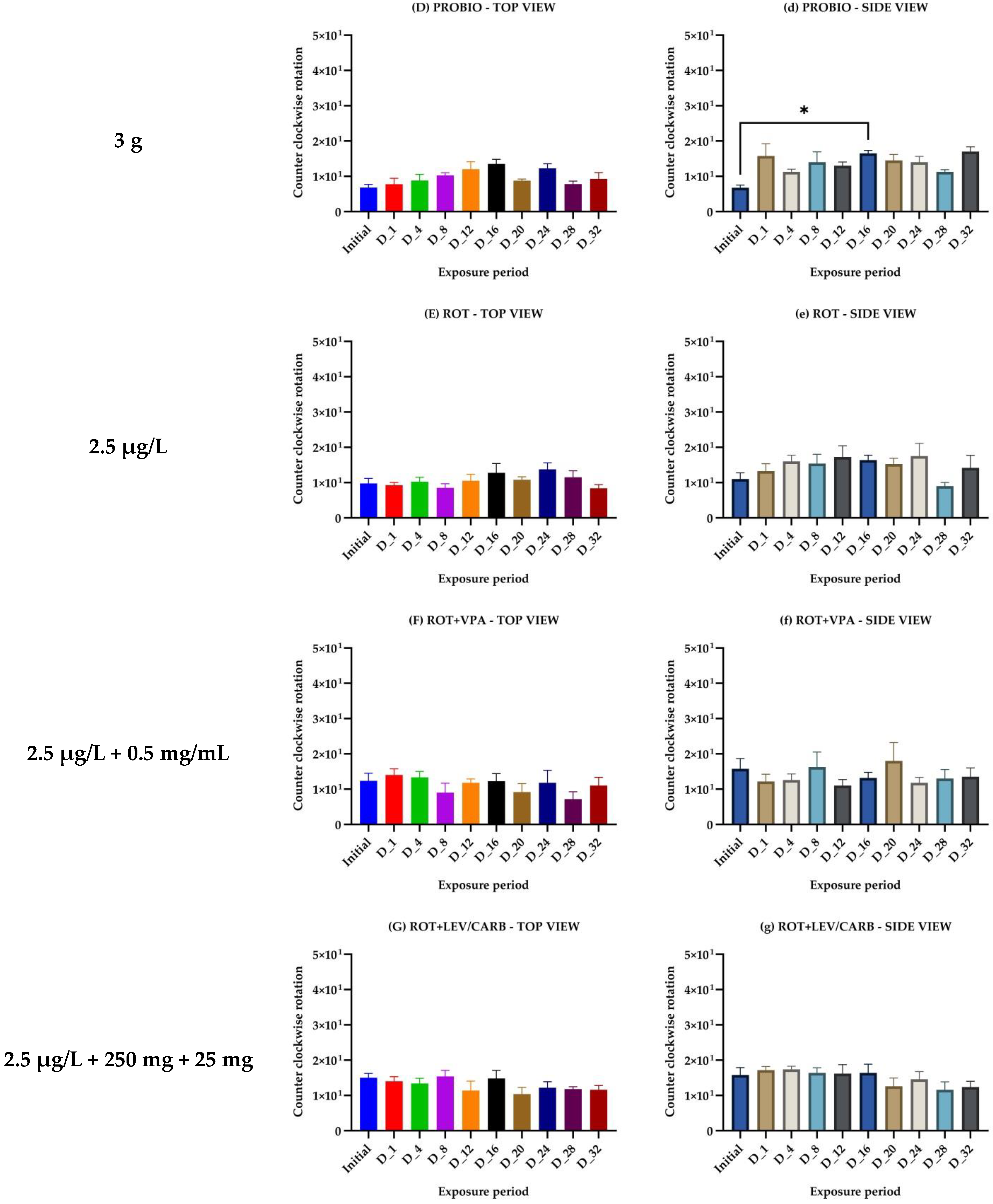{kind=link}
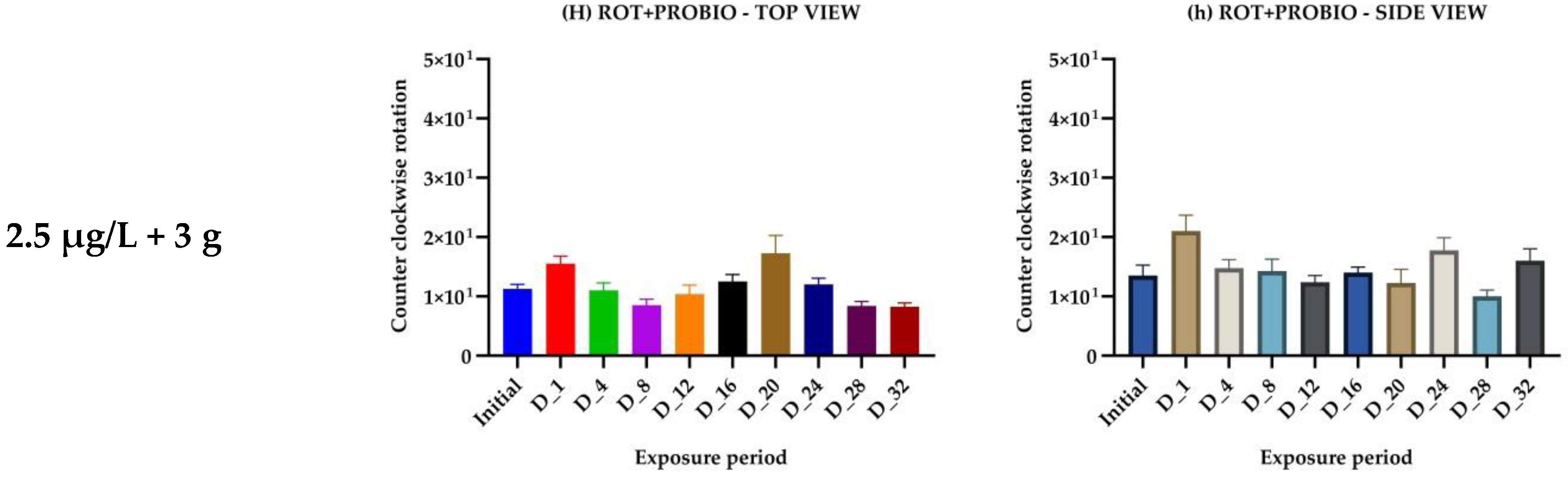{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Animal Maintenance
2.2. Ethical Note
2.3. Ant(Agonists) and Lactic Acid Lacteria Strains
2.4. Behavioral Testing
2.4.1. Anti-Social and Aggressivity Behaviors
2.4.2. Cycling Rotation
2.5. Statistical Analyses
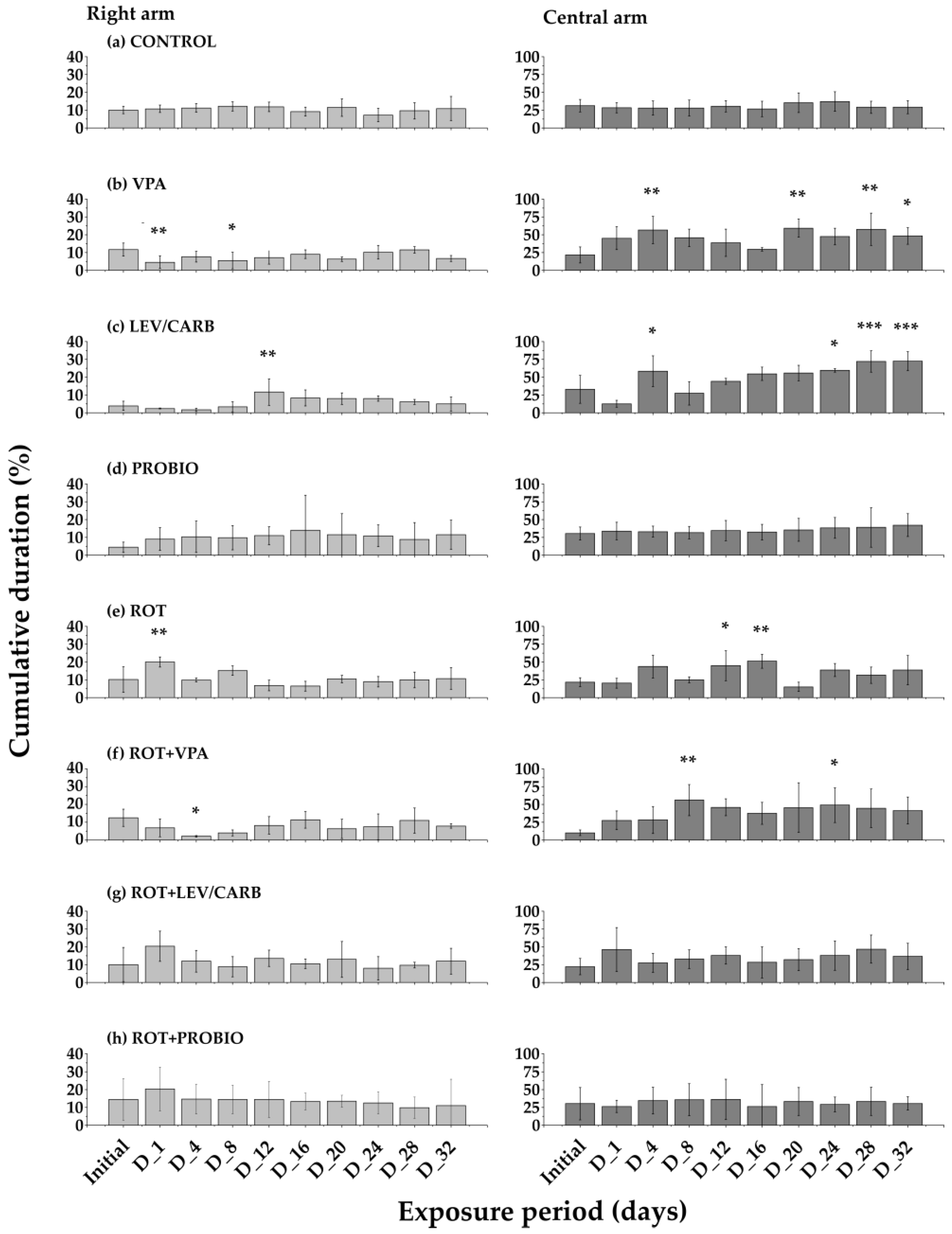
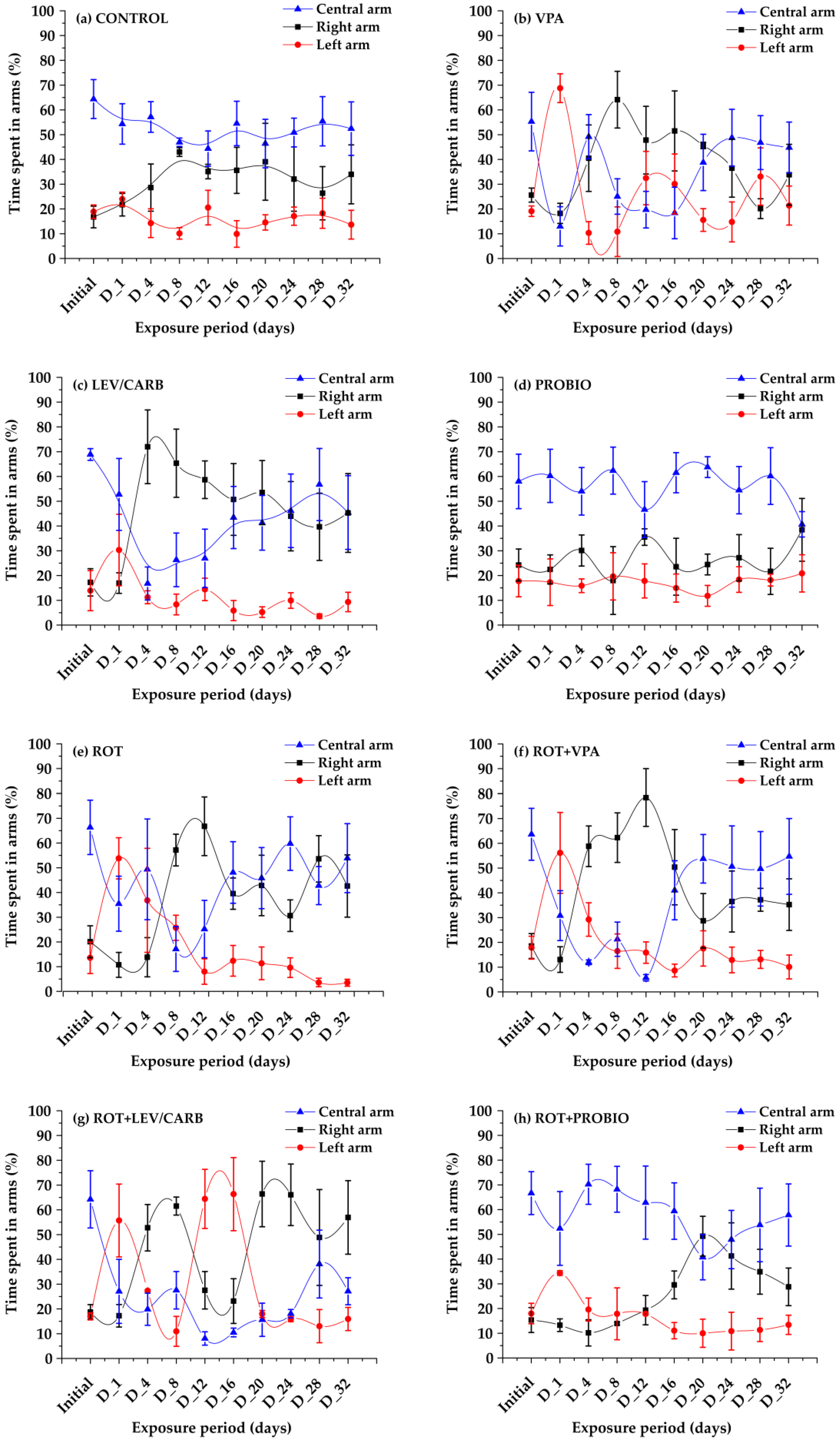
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Robea, M.-A.; Balmus, I.-M.; Ciobica, A.; Strungaru, S.; Plavan, G.; Gorgan, L.D.; Savuca, A.; Nicoara, M. Parkinson’s Disease-Induced Zebrafish Models: Focussing on Oxidative Stress Implications and Sleep Processes. Oxid. Med. Cell. Longev. 2020, 2020, 1370837. [Google Scholar] [CrossRef] [PubMed]
- Sherer, T.B.; Betarbet, R.; Testa, C.M.; Seo, B.B.; Richardson, J.R.; Kim, J.H.; Miller, G.W.; Yagi, T.; Matsuno-Yagi, A.; Greenamyre, J.T. Mechanism of toxicity in rotenone models of Parkinson’s disease. J. Neurosci. 2003, 23, 10756. [Google Scholar] [CrossRef] [PubMed]
- Bové, J.; Prou, D.; Perier, C.; Przedborski, S. Toxin-induced models of Parkinson’s disease. NeuroRX 2005, 2, 484–494. [Google Scholar] [CrossRef] [Green Version]
- Lawana, V.; Cannon, J.R. Chapter Five—Rotenone neurotoxicity: Relevance to Parkinson’s disease. In Neurotoxicity of Pesticides; Aschner, M., Costa, L.G., Eds.; Academic Press: Cambridge, MA, USA, 2020; Volume 4, pp. 209–254. ISBN 2468-7480. [Google Scholar]
- Gupta, R.C. (Ed.) Chapter 52—Rotenone; Academic Press: Boston, MA, USA, 2012; pp. 620–623. ISBN 978-0-12-385926-6. [Google Scholar]
- Gupta, R.C. Veterinary Toxicology: Basic and Clinical Principles; Elsevier: New York, NY, USA; Academic Press: London, UK, 2007; ISBN 9780123704672. [Google Scholar]
- Bastías-Candia, S.; Zolezzi, J.M.; Inestrosa, N.C. Revisiting the Paraquat-Induced Sporadic Parkinson’s Disease-Like Model. Mol. Neurobiol. 2019, 56, 1044–1055. [Google Scholar] [CrossRef]
- Yurtsever, İ.; Üstündağ, Ü.V.; Ünal, İ.; Ateş, P.S.; Emekli-Alturfan, E. Rifampicin decreases neuroinflammation to maintain mitochondrial function and calcium homeostasis in rotenone-treated zebrafish. Drug Chem. Toxicol. 2020, 45, 1–8. [Google Scholar] [CrossRef]
- Gao, F.; Chen, D.; Hu, Q.; Wang, G. Rotenone Directly Induces BV2 Cell Activation via the p38 MAPK Pathway. PLoS ONE 2013, 8, e72046. [Google Scholar]
- Hijaz, B.A.; Volpicelli-Daley, L.A. Initiation and propagation of α-synuclein aggregation in the nervous system. Mol. Neurodegener. 2020, 15, 19. [Google Scholar] [CrossRef] [PubMed]
- Reed, B.; Jennings, M. Guidance on the Housing and Care of Zebrafish Danio rerio; RSPCA: Horsham, UK, 2011. [Google Scholar]
- Ilie, O.-D.; Paduraru, E.; Robea, M.-A.; Balmus, I.-M.; Jijie, R.; Nicoara, M.; Ciobica, A.; Nita, I.-B.; Dobrin, R.; Doroftei, B. The Possible Role of Bifidobacterium longum BB536 and Lactobacillus rhamnosus HN001 on Locomotor Activity and Oxidative Stress in a Rotenone-Induced Zebrafish Model of Parkinson’s Disease. Oxid. Med. Cell. Longev. 2021, 2021, 9629102. [Google Scholar] [CrossRef]
- Wang, Y.; Liu, W.; Yang, J.; Wang, F.; Sima, Y.; Zhong, Z.; Wang, H.; Hu, L.-F.; Liu, C.-F. Parkinson’s disease-like motor and non-motor symptoms in rotenone-treated zebrafish. Neurotoxicology 2017, 58, 103–109. [Google Scholar] [CrossRef]
- Khotimah, H.; Ali, M.; Sumitro, S.B.; Widodo, M.A. Decreasing α-synuclein aggregation by methanolic extract of Centella asiatica in zebrafish Parkinson’s model. Asian Pac. J. Trop. Biomed. 2015, 5, 948–954. [Google Scholar] [CrossRef] [Green Version]
- Ünal, İ.; Çalışkan-Ak, E.; Üstündağ, Ü.V.; Ateş, P.S.; Alturfan, A.A.; Altinoz, M.A.; Elmaci, I.; Emekli-Alturfan, E. Neuroprotective effects of mitoquinone and oleandrin on Parkinson’s disease model in zebrafish. Int. J. Neurosci. 2020, 130, 574–582. [Google Scholar] [CrossRef] [PubMed]
- Khotimah, H.; Sumitro, S.; Widodo, M. Zebrafish Parkinson’s model: Rotenone decrease motility, dopamine, and increase α-synuclein aggregation and apoptosis of zebrafish brain. Int. J. Pharm. Tech. Res. 2015, 8, 614–621. [Google Scholar]
- Cansız, D.; Ünal, İ.; Üstündağ, Ü.V.; Alturfan, A.A.; Altinoz, M.A.; Elmacı, İ.; Emekli-Alturfan, E. Caprylic acid ameliorates rotenone induced inflammation and oxidative stress in the gut-brain axis in Zebrafish. Mol. Biol. Rep. 2021, 48, 5259–5273. [Google Scholar] [CrossRef] [PubMed]
- Strungaru, S.-A.; Robea, M.A.; Plavan, G.; Todirascu-Ciornea, E.; Ciobica, A.; Nicoara, M. Acute exposure to methylmercury chloride induces fast changes in swimming performance, cognitive processes and oxidative stress of zebrafish (Danio rerio) as reference model for fish community. J. Trace Elem. Med. Biol. 2018, 47, 115–123. [Google Scholar] [CrossRef]
- Strungaru, S.-A.; Plavan, G.; Ciobica, A.; Nicoara, M.; Robea, M.A.; Solcan, C.; Petrovici, A. Toxicity and chronic effects of deltamethrin exposure on zebrafish (Danio rerio) as a reference model for freshwater fish community. Ecotoxicol. Environ. Saf. 2019, 171, 854–862. [Google Scholar] [CrossRef]
- Kalueff, A.V. (Ed.) Illustrated Zebrafish Neurobehavioral Glossary BT—The Rights and Wrongs of Zebrafish: Behavioral Phenotyping of Zebrafish; Springer International Publishing: Cham, Switzerland, 2017; pp. 291–317. ISBN 978-3-319-33774-6. [Google Scholar]
- Jijie, R.; Solcan, G.; Nicoara, M.; Micu, D.; Strungaru, S.-A. Antagonistic effects in zebrafish (Danio rerio) behavior and oxidative stress induced by toxic metals and deltamethrin acute exposure. Sci. Total Environ. 2020, 698, 134299. [Google Scholar] [CrossRef]
- Paduraru, E.; Flocea, E.-I.; Lazado, C.C.; Simionov, I.-A.; Nicoara, M.; Ciobica, A.; Faggio, C.; Jijie, R. Vitamin C Mitigates Oxidative Stress and Behavioral Impairments Induced by Deltamethrin and Lead Toxicity in Zebrafish. Int. J. Mol. Sci. 2021, 22, 12714. [Google Scholar] [CrossRef]
- Jijie, R.; Mihalache, G.; Balmus, I.-M.; Strungaru, S.-A.; Baltag, E.S.; Ciobica, A.; Nicoara, M.; Faggio, C. Zebrafish as a Screening Model to Study the Single and Joint Effects of Antibiotics. Pharmaceuticals 2021, 14, 578. [Google Scholar] [CrossRef]
- Kalueff, A.V.; Gebhardt, M.; Stewart, A.M.; Cachat, J.M.; Brimmer, M.; Chawla, J.S.; Craddock, C.; Kyzar, E.J.; Roth, A.; Landsman, S.; et al. Towards a Comprehensive Catalog of Zebrafish Behavior 1.0 and Beyond. Zebrafish 2013, 10, 70–86. [Google Scholar] [CrossRef]
- Faria, M.; Prats, E.; Rosas Ramírez, J.R.; Bellot, M.; Bedrossiantz, J.; Pagano, M.; Valls, A.; Gomez-Canela, C.; Porta, J.M.; Mestres, J.; et al. Androgenic activation, impairment of the monoaminergic system and altered behavior in zebrafish larvae exposed to environmental concentrations of fenitrothion. Sci. Total Environ. 2021, 775, 145671. [Google Scholar] [CrossRef] [PubMed]
- Lieschke, G.J.; Currie, P.D. Animal models of human disease: Zebrafish swim into view. Nat. Rev. Genet. 2007, 8, 353–367. [Google Scholar] [CrossRef] [PubMed]
- Ünal, İ.; Üstündağ, Ü.V.; Ateş, P.S.; Eğilmezer, G.; Alturfan, A.A.; Yiğitbaşı, T.; Emekli-Alturfan, E. Rotenone impairs oxidant/antioxidant balance both in brain and intestines in zebrafish. Int. J. Neurosci. 2019, 129, 363–368. [Google Scholar] [CrossRef] [PubMed]
- Lv, D.; Li, L.; Chen, J.; Wei, S.-Z.; Wang, F.; Hu, H.; Xie, A.-M.; Liu, C.-F. Sleep deprivation caused a memory defects and emotional changes in a rotenone-based zebrafish model of Parkinson’s disease. Behav. Brain Res. 2019, 372, 112031. [Google Scholar] [CrossRef] [PubMed]
- Carriere, C.H.; Kang, N.H.; Niles, L.P. Neuroprotection by valproic acid in an intrastriatal rotenone model of Parkinson’s disease. Neuroscience 2014, 267, 114–121. [Google Scholar] [CrossRef]
- Monti, B.; Gatta, V.; Piretti, F.; Raffaelli, S.S.; Virgili, M.; Contestabile, A. Valproic Acid is Neuroprotective in the Rotenone Rat Model of Parkinson’s Disease: Involvement of α-Synuclein. Neurotox. Res. 2010, 17, 130–141. [Google Scholar] [CrossRef]
- Idalencio, R.; Lopes, T.M.; Soares, S.M.; Pompermaier, A.; de Alcantara Barcellos, H.H.; Kalichak, F.; Fagundes, M.; de Oliveira, C.M.; Barcellos, L.J.G. Effect of levodopa/carbidopa on stress response in zebrafish. J. Comp. Physiol. A 2021, 207, 393–399. [Google Scholar] [CrossRef]
- Tan, A.H.; Hor, J.W.; Chong, C.W.; Lim, S.-Y. Probiotics for Parkinson’s disease: Current evidence and future directions. JGH Open 2021, 5, 414–419. [Google Scholar] [CrossRef]
- Robea, M.A.; Ciobica, A.; Curpan, A.-S.; Plavan, G.; Strungaru, S.; Lefter, R.; Nicoara, M. Preliminary Results Regarding Sleep in a Zebrafish Model of Autism Spectrum Disorder. Brain Sci. 2021, 11, 556. [Google Scholar] [CrossRef]
- Zimmermann, F.F.; Gaspary, K.V.; Leite, C.E.; De Paula Cognato, G.; Bonan, C.D. Embryological exposure to valproic acid induces social interaction deficits in zebrafish (Danio rerio): A developmental behavior analysis. Neurotoxicol. Teratol. 2015, 52, 36–41. [Google Scholar] [CrossRef]
- Liu, X.; Zhang, Y.; Lin, J.; Xia, Q.; Guo, N.; Li, Q. Social Preference Deficits in Juvenile Zebrafish Induced by Early Chronic Exposure to Sodium Valproate. Front. Behav. Neurosci. 2016, 10, 201. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tan, W.-H.; Bird, L.M.; Sadhwani, A.; Barbieri-Welge, R.L.; Skinner, S.A.; Horowitz, L.T.; Bacino, C.A.; Noll, L.M.; Fu, C.; Hundley, R.J.; et al. A randomized controlled trial of levodopa in patients with Angelman syndrome. Am. J. Med. Genet. A 2018, 176, 1099–1107. [Google Scholar] [CrossRef] [PubMed]
- Riday, T.T.; Dankoski, E.C.; Krouse, M.C.; Fish, E.W.; Walsh, P.L.; Han, J.E.; Hodge, C.W.; Wightman, R.M.; Philpot, B.D.; Malanga, C.J. Pathway-specific dopaminergic deficits in a mouse model of Angelman syndrome. J. Clin. Investig. 2012, 122, 4544–4554. [Google Scholar] [CrossRef]
- Newman, E.J.; Grosset, D.G.; Kennedy, P.G.E. The Parkinsonism-Hyperpyrexia Syndrome. Neurocrit. Care 2009, 10, 136–140. [Google Scholar] [CrossRef]
- Kutcher, E.O.; Egorov, A.Y.; Chernikova, N.A.; Filatova, E.V. Long-Term Ethanol Alcoholization Decreases Levodopa- Induced Aggressive Behavior in Rats. J. Evol. Biochem. Physiol. 2018, 54, 216–220. [Google Scholar] [CrossRef]
- Hira, S.; Saleem, U.; Anwar, F.; Ahmad, B. Antioxidants Attenuate Isolation- and L-DOPA-Induced Aggression in Mice. Front. Pharmacol. 2018, 8, 945. [Google Scholar] [CrossRef] [Green Version]
- Valcarce, D.G.; Martínez-Vázquez, J.M.; Riesco, M.F.; Robles, V. Probiotics reduce anxiety-related behavior in zebrafish. Heliyon 2020, 6, e03973. [Google Scholar] [CrossRef] [PubMed]
- Borrelli, L.; Aceto, S.; Agnisola, C.; De Paolo, S.; Dipineto, L.; Stilling, R.M.; Dinan, T.G.; Cryan, J.F.; Menna, L.F.; Fioretti, A. Probiotic modulation of the microbiota-gut-brain axis and behaviour in zebrafish. Sci. Rep. 2016, 6, 30046. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Davis, D.J.; Bryda, E.C.; Gillespie, C.H.; Ericsson, A.C. Microbial modulation of behavior and stress responses in zebrafish larvae. Behav. Brain Res. 2016, 311, 219–227. [Google Scholar] [CrossRef]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ilie, O.-D.; Duta, R.; Jijie, R.; Nita, I.-B.; Nicoara, M.; Faggio, C.; Dobrin, R.; Mavroudis, I.; Ciobica, A.; Doroftei, B. Assessing Anti-Social and Aggressive Behavior in a Zebrafish (Danio rerio) Model of Parkinson’s Disease Chronically Exposed to Rotenone. Brain Sci. 2022, 12, 898. https://doi.org/10.3390/brainsci12070898
Ilie O-D, Duta R, Jijie R, Nita I-B, Nicoara M, Faggio C, Dobrin R, Mavroudis I, Ciobica A, Doroftei B. Assessing Anti-Social and Aggressive Behavior in a Zebrafish (Danio rerio) Model of Parkinson’s Disease Chronically Exposed to Rotenone. Brain Sciences. 2022; 12(7):898. https://doi.org/10.3390/brainsci12070898
Chicago/Turabian StyleIlie, Ovidiu-Dumitru, Raluca Duta, Roxana Jijie, Ilinca-Bianca Nita, Mircea Nicoara, Caterina Faggio, Romeo Dobrin, Ioannis Mavroudis, Alin Ciobica, and Bogdan Doroftei. 2022. "Assessing Anti-Social and Aggressive Behavior in a Zebrafish (Danio rerio) Model of Parkinson’s Disease Chronically Exposed to Rotenone" Brain Sciences 12, no. 7: 898. https://doi.org/10.3390/brainsci12070898
APA StyleIlie, O.-D., Duta, R., Jijie, R., Nita, I.-B., Nicoara, M., Faggio, C., Dobrin, R., Mavroudis, I., Ciobica, A., & Doroftei, B. (2022). Assessing Anti-Social and Aggressive Behavior in a Zebrafish (Danio rerio) Model of Parkinson’s Disease Chronically Exposed to Rotenone. Brain Sciences, 12(7), 898. https://doi.org/10.3390/brainsci12070898