
Hindbrain Stimulation Modulates Object Recognition Discrimination Efficiency and Hippocampal Synaptic Connections


{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
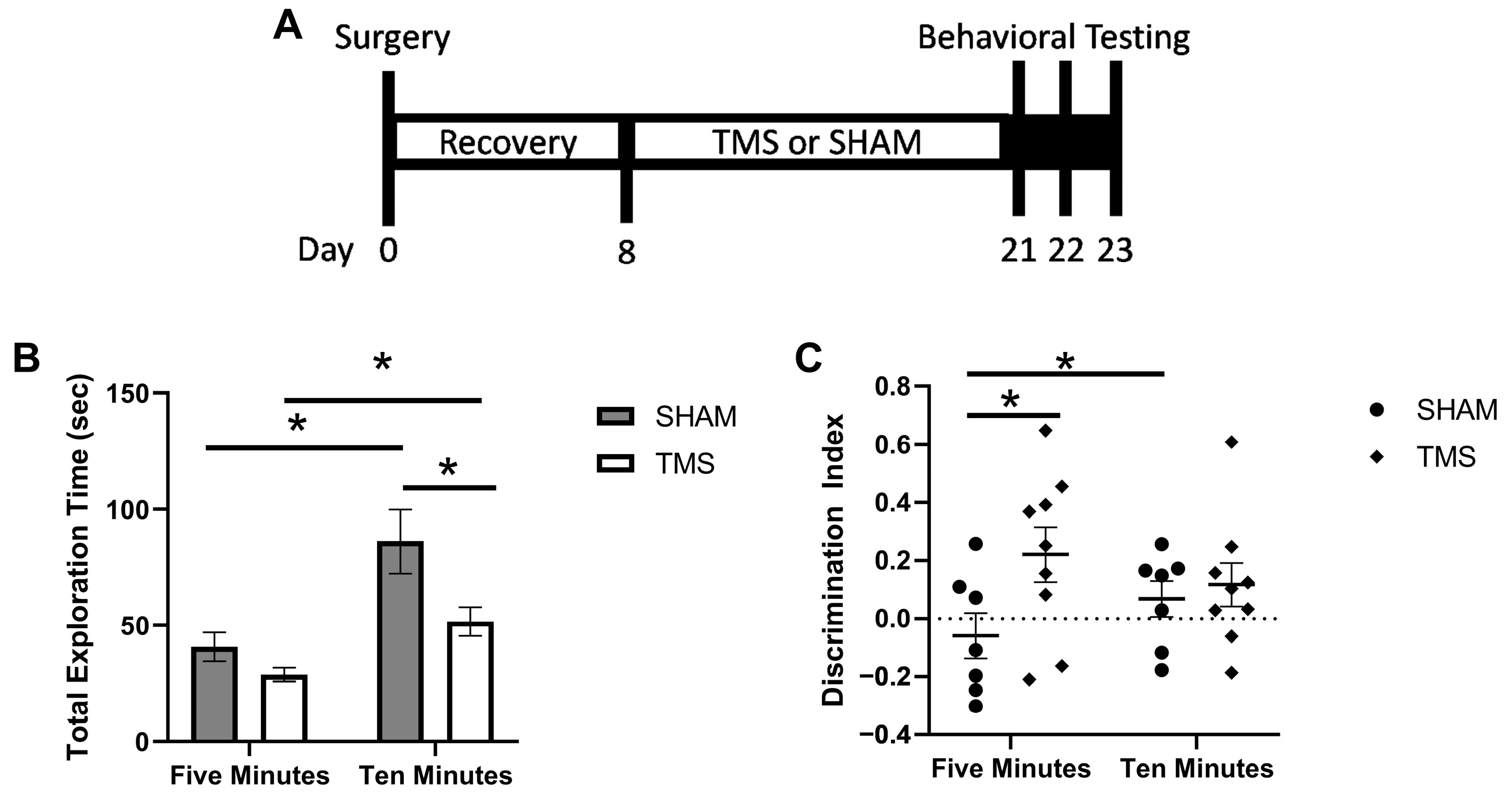
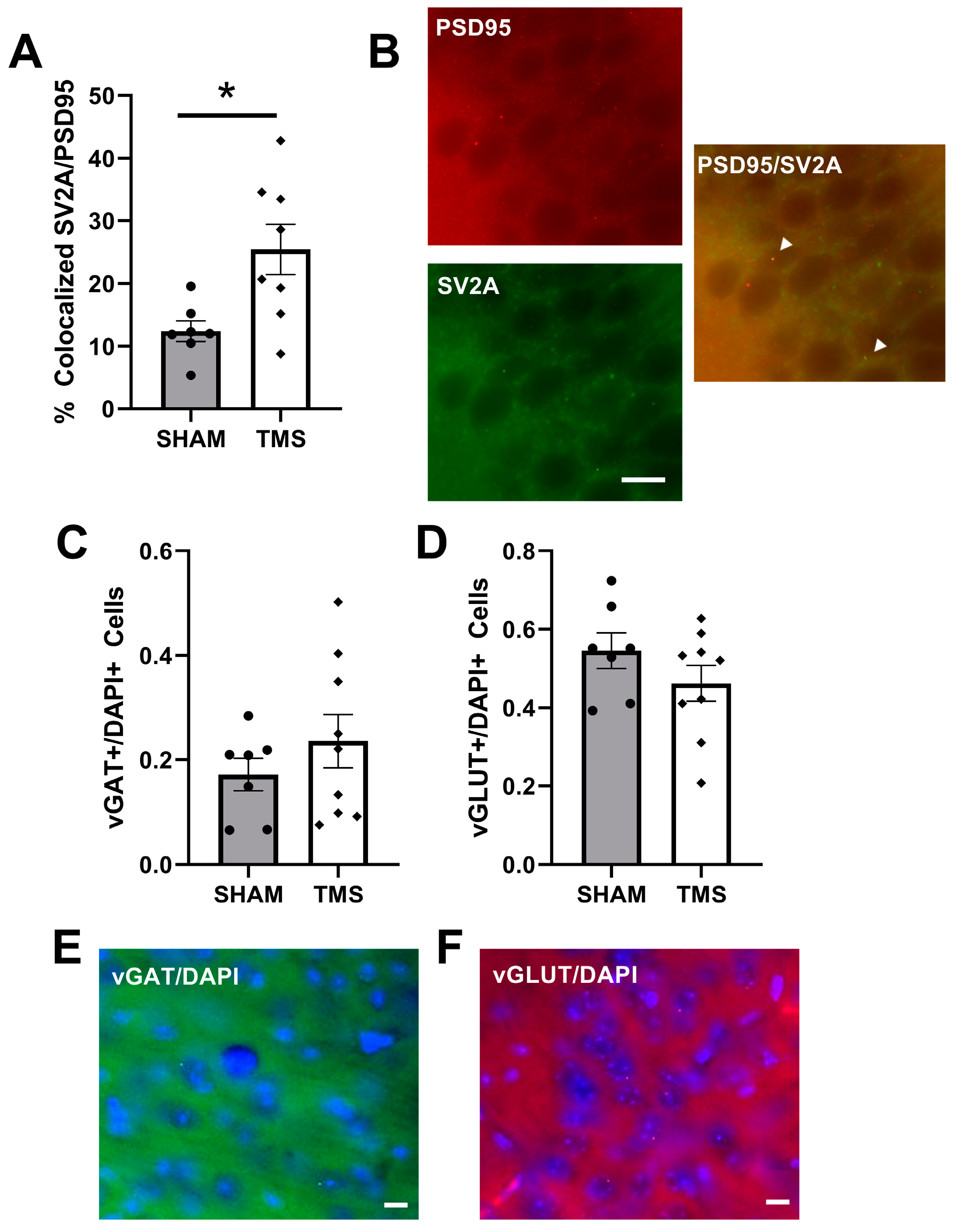
3. Results
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Villanueva, R. The Cerebellum and Neuropsychiatric Disorders. Psychiatry Res. 2012, 198, 527–532. [Google Scholar] [CrossRef] [PubMed]
- Schmahmann, J.D. The Cerebellum and Cognition. Neurosci. Lett. 2019, 688, 62–75. [Google Scholar] [CrossRef] [PubMed]
- Yu, W.; Krook-Magnuson, E. Cognitive Collaborations: Bidirectional Functional Connectivity between the Cerebellum and the Hippocampus. Front. Syst. Neurosci. 2015, 9, 177. [Google Scholar] [CrossRef] [PubMed]
- Rogers, T.D.; Dickson, P.E.; Heck, D.H.; Goldowitz, D.; Mittleman, G.; Blaha, C.D. Connecting the Dots of the Cerebro-Cerebellar Role in Cognitive Function: Neuronal Pathways for Cerebellar Modulation of Dopamine Release in the Prefrontal Cortex. Synapse 2011, 65, 1204–1212. [Google Scholar] [CrossRef] [PubMed]
- Barker, A.T.; Jalinous, R.; Freeston, I.L. Non-Invasive Magnetic Stimulation of Human Motor Cortex. Lancet 1985, 325, 1106–1107. [Google Scholar] [CrossRef] [PubMed]
- Baeken, C.; Raedt, R.D. Neurobiological Mechanisms of Repetitive Transcranial Magnetic Stimulation on the Underlying Neuro Circuitry in Unipolar Depression. Dialogues Clin. Neurosci. 2011, 13, 139. [Google Scholar] [CrossRef] [PubMed]
- Rossi, S.; Hallett, M.; Rossini, P.M.; Pascual-Leone, A.; Avanzini, G.; Bestmann, S.; Berardelli, A.; Brewer, C.; Canli, T.; Cantello, R.; et al. Safety, Ethical Considerations, and Application Guidelines for the Use of Transcranial Magnetic Stimulation in Clinical Practice and Research. Clin. Neurophysiol. 2009, 120, 2008–2039. [Google Scholar] [CrossRef]
- Perera, T.; George, M.S.; Grammer, G.; Janicak, P.G.; Pascual-Leone, A.; Wiercki, T.S. The Clinical TMS Society Consensus Review and Treatment Recommendations for TMS Therapy for Major Depressive Disorder. Brain Stimul. 2016, 9, 336–346. [Google Scholar] [CrossRef]
- Shivakumar, V.; Dinakaran, D.; Narayanaswamy, J.C.; Venkatasubramanian, G. Noninvasive Brain Stimulation in Obsessive–Compulsive Disorder. Indian J. Psychiatry 2019, 61, S66. [Google Scholar] [CrossRef]
- Zangen, A.; Moshe, H.; Martinez, D.; Barnea-Ygael, N.; Vapnik, T.; Bystritsky, A.; Duffy, W.; Toder, D.; Casuto, L.; Grosz, M.L.; et al. Repetitive Transcranial Magnetic Stimulation for Smoking Cessation: A Pivotal Multicenter Double-Blind Randomized Controlled Trial. World Psychiatry 2021, 20, 397–404. [Google Scholar] [CrossRef]
- Pell, G.S.; Harmelech, T.; Zibman, S.; Roth, Y.; Tendler, A.; Zangen, A. Efficacy of Deep TMS with the H1 Coil for Anxious Depression. J. Clin. Med. 2022, 11, 1015. [Google Scholar] [CrossRef]
- Machado, S.; Arias-Carrión, O.; Paes, F.; Vieira, R.T.; Caixeta, L.; Novaes, F.; Marinho, T.; Almada, L.F.; Silva, A.C.; Nardi, A.E. Repetitive Transcranial Magnetic Stimulation for Clinical Applications in Neurological and Psychiatric Disorders: An Overview. Eurasian J. Med. 2013, 45, 191–206. [Google Scholar] [CrossRef] [PubMed]
- Harrington, A.; Hammond-Tooke, G.D. Theta Burst Stimulation of the Cerebellum Modifies the TMS-Evoked N100 Potential, a Marker of GABA Inhibition. PLoS ONE 2015, 10, e0141284. [Google Scholar] [CrossRef] [PubMed]
- Koch, G.; Porcacchia, P.; Ponzo, V.; Carrillo, F.; Cáceres-Redondo, M.T.; Brusa, L.; Desiato, M.T.; Arciprete, F.; Lorenzo, F.D.; Pisani, A.; et al. Effects of Two Weeks of Cerebellar Theta Burst Stimulation in Cervical Dystonia Patients. Brain Stimul. 2014, 7, 564–572. [Google Scholar] [CrossRef] [PubMed]
- Heleven, E.; van Dun, K.; Witte, S.D.; Baeken, C.; Overwalle, F.V. The Role of the Cerebellum in Social and Non-Social Action Sequences: A Preliminary LF-rTMS Study. Front. Hum. Neurosci. 2021, 15, 85. [Google Scholar] [CrossRef]
- Grehl, S. In Vitro Magnetic Stimulation: A Simple Stimulation Device to Deliver Defined Low Intensity Electromagnetic Fields. Front. Neural Circuits 2016, 10, 85. [Google Scholar] [CrossRef]
- Dufor, T.; Grehl, S.; Tang, A.D.; Doulazmi, M.; Traoré, M.; Debray, N.; Dubacq, C.; Deng, Z.-D.; Mariani, J.; Lohof, A.M.; et al. Neural Circuit Repair by Low-Intensity Magnetic Stimulation Requires Cellular Magnetoreceptors and Specific Stimulation Patterns. Sci. Adv. 2019, 5, 9847. [Google Scholar] [CrossRef] [PubMed]
- Leuner, B.; Gould, E. Structural Plasticity and Hippocampal Function. Annu. Rev. Psychol. 2010, 61, 111–140. [Google Scholar] [CrossRef]
- Madore, M.R.; Poh, E.; Bollard, S.J.; Rivera, J.; Taylor, J.; Cheng, J.; Booth, E.; Nable, M.; Heath, A.; Yesavage, J.; et al. Moving Back in the Brain to Drive the Field Forward: Targeting Neurostimulation to Different Brain Regions in Animal Models of Depression and Neurodegeneration. J. Neurosci. Methods 2021, 360, 109261. [Google Scholar] [CrossRef]
- Zeidler, Z.; Hoffmann, K.; Krook-Magnuson, E. HippoBellum: Acute Cerebellar Modulation Alters Hippocampal Dynamics and Function. J. Neurosci. 2020, 40, 6910–6926. [Google Scholar] [CrossRef]
- Heath, A.M.; Brewer, M.; Yesavage, J.; McNerney, M.W. Improved Object Recognition Memory Using Post-Encoding Repetitive Transcranial Magnetic Stimulation. Brain Stimul. 2022, 15, 78–86. [Google Scholar] [CrossRef] [PubMed]
- Faul, F.; Erdfelder, E.; Buchner, A.; Lang, A.-G. Statistical Power Analyses Using G*Power 3.1: Tests for Correlation and Regression Analyses. Behav. Res. Methods 2009, 41, 1149–1160. [Google Scholar] [CrossRef] [PubMed]
- Timmann, D.; Drepper, J.; Maschke, M.; Kolb, F.P.; Böring, D.; Thilmann, A.F.; Diener, H.C. Motor Deficits Cannot Explain Impaired Cognitive Associative Learning in Cerebellar Patients. Neuropsychologia 2002, 40, 788–800. [Google Scholar] [CrossRef] [PubMed]
- Wong, C.H.Y.; Liu, J.; Lee, T.M.C.; Tao, J.; Wong, A.W.K.; Chau, B.K.H.; Chen, L.; Chan, C.C.H. Fronto-Cerebellar Connectivity Mediating Cognitive Processing Speed. NeuroImage 2021, 226, 117556. [Google Scholar] [CrossRef]
- Nixon, P.D.; Passingham, R.E. The Cerebellum and Cognition: Cerebellar Lesions Do Not Impair Spatial Working Memory or Visual Associative Learning in Monkeys. Eur. J. Neurosci. 1999, 11, 4070–4080. [Google Scholar] [CrossRef] [PubMed]
- Binder, S.; Dere, E.; Zlomuzica, A. A Critical Appraisal of the What-Where-When Episodic-like Memory Test in Rodents: Achievements, Caveats and Future Directions. Prog. Neurobiol. 2015, 130, 71–85. [Google Scholar] [CrossRef] [PubMed]
- Chao, O.Y.; Zhang, H.; Pathak, S.S.; Huston, J.P.; Yang, Y.M. Functional Convergence of Motor and Social Processes in Lobule IV/V of the Mouse Cerebellum. Cerebellum 2021, 20, 836–852. [Google Scholar] [CrossRef]


Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Heath, A.; Madore, M.; Diaz, K.; McNerney, M.W. Hindbrain Stimulation Modulates Object Recognition Discrimination Efficiency and Hippocampal Synaptic Connections. Brain Sci. 2023, 13, 1425. https://doi.org/10.3390/brainsci13101425
Heath A, Madore M, Diaz K, McNerney MW. Hindbrain Stimulation Modulates Object Recognition Discrimination Efficiency and Hippocampal Synaptic Connections. Brain Sciences. 2023; 13(10):1425. https://doi.org/10.3390/brainsci13101425
Chicago/Turabian StyleHeath, Alesha, Michelle Madore, Karina Diaz, and M. Windy McNerney. 2023. "Hindbrain Stimulation Modulates Object Recognition Discrimination Efficiency and Hippocampal Synaptic Connections" Brain Sciences 13, no. 10: 1425. https://doi.org/10.3390/brainsci13101425
APA StyleHeath, A., Madore, M., Diaz, K., & McNerney, M. W. (2023). Hindbrain Stimulation Modulates Object Recognition Discrimination Efficiency and Hippocampal Synaptic Connections. Brain Sciences, 13(10), 1425. https://doi.org/10.3390/brainsci13101425