

A Brief History of Stereotactic Atlases: Their Evolution and Importance in Stereotactic Neurosurgery

 ,
,  and
and {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. The Discovery of Brain Localization and Cranio-Cerebral Topography before Atlases
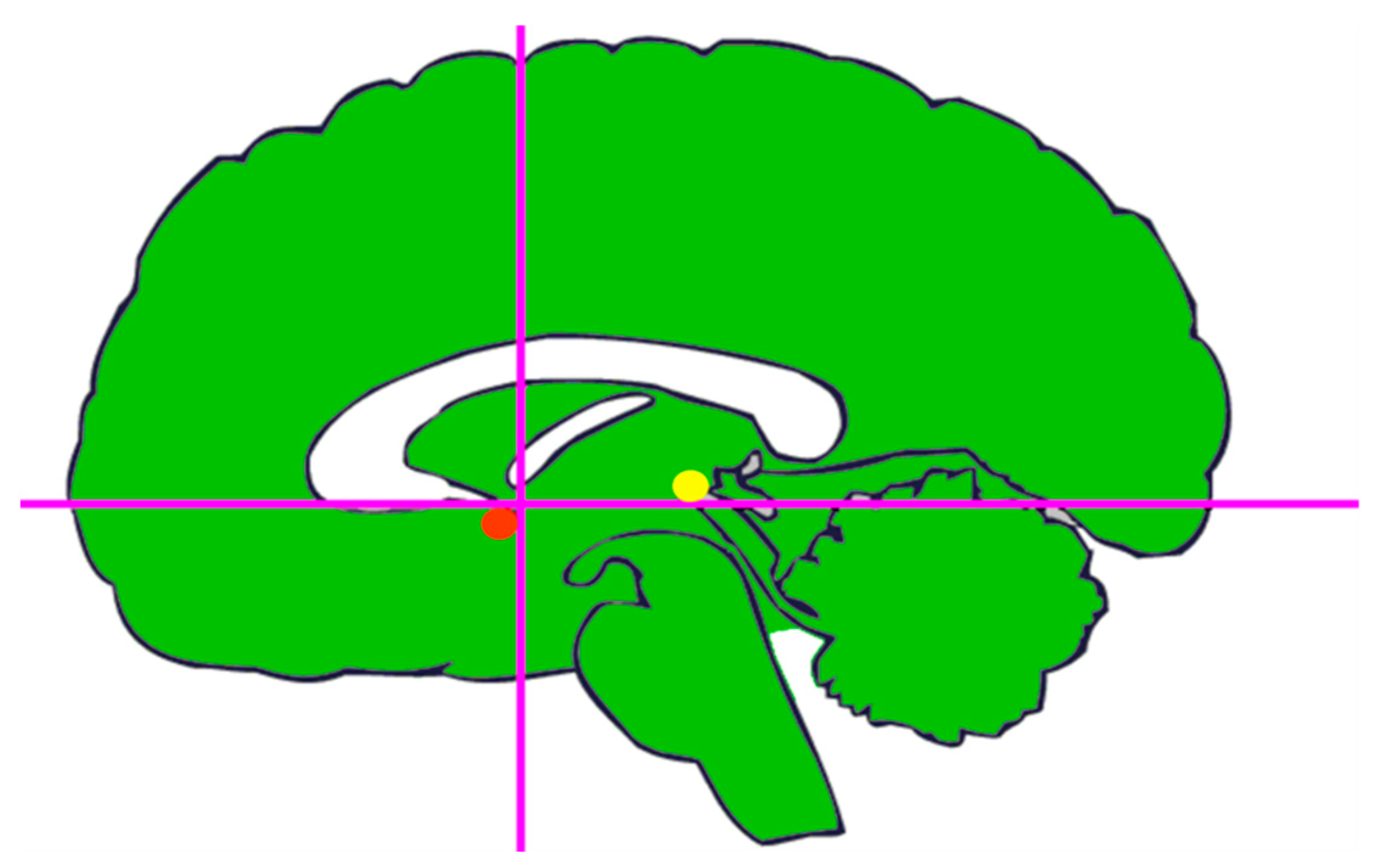
3. The Invention of the Stereotactic Atlas
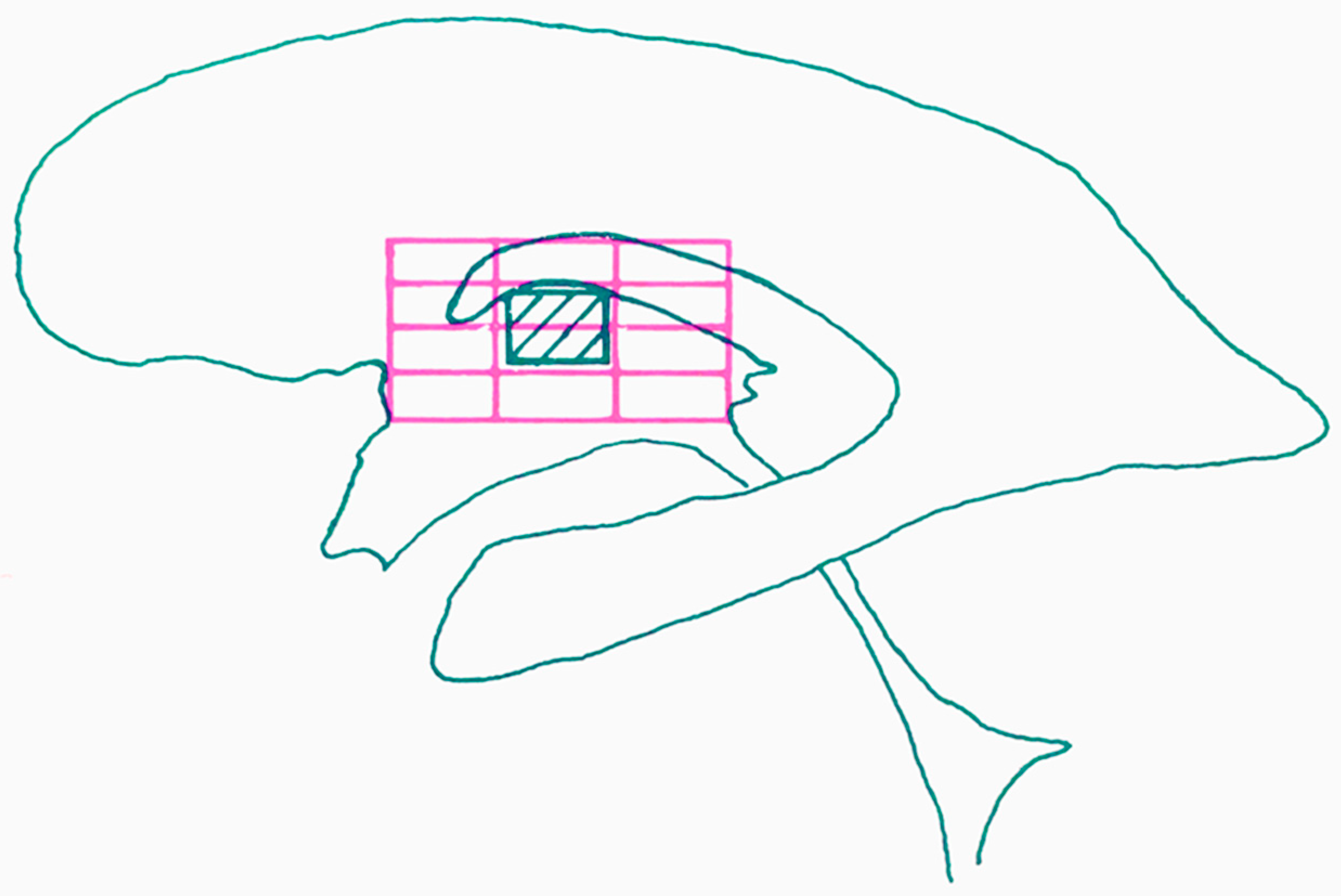
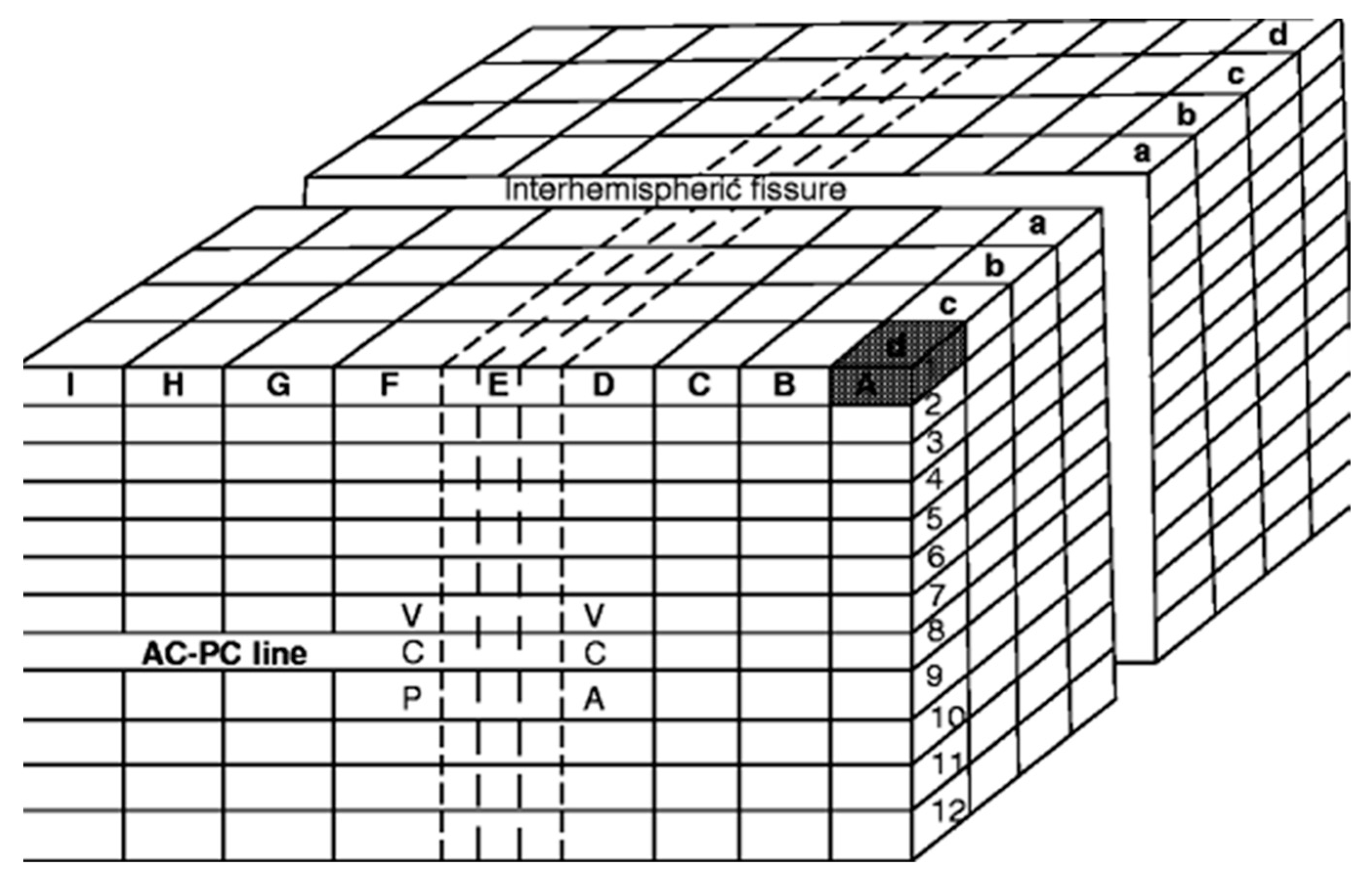
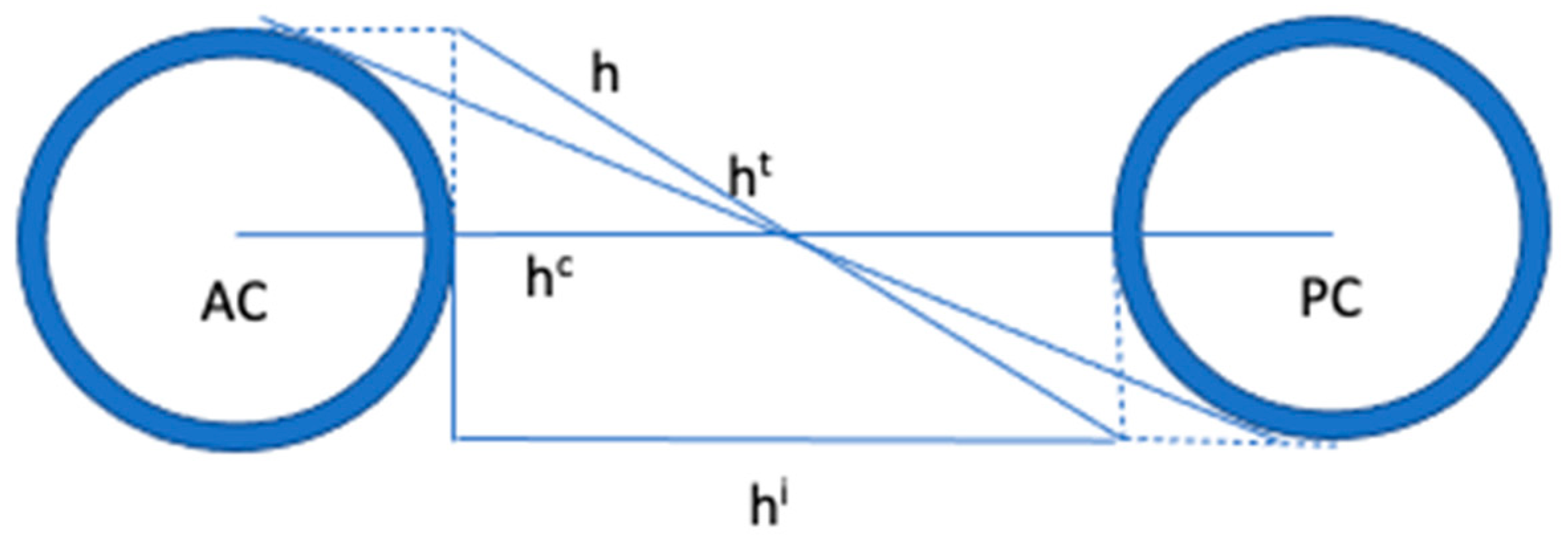
4. The Proportional System of Talairach
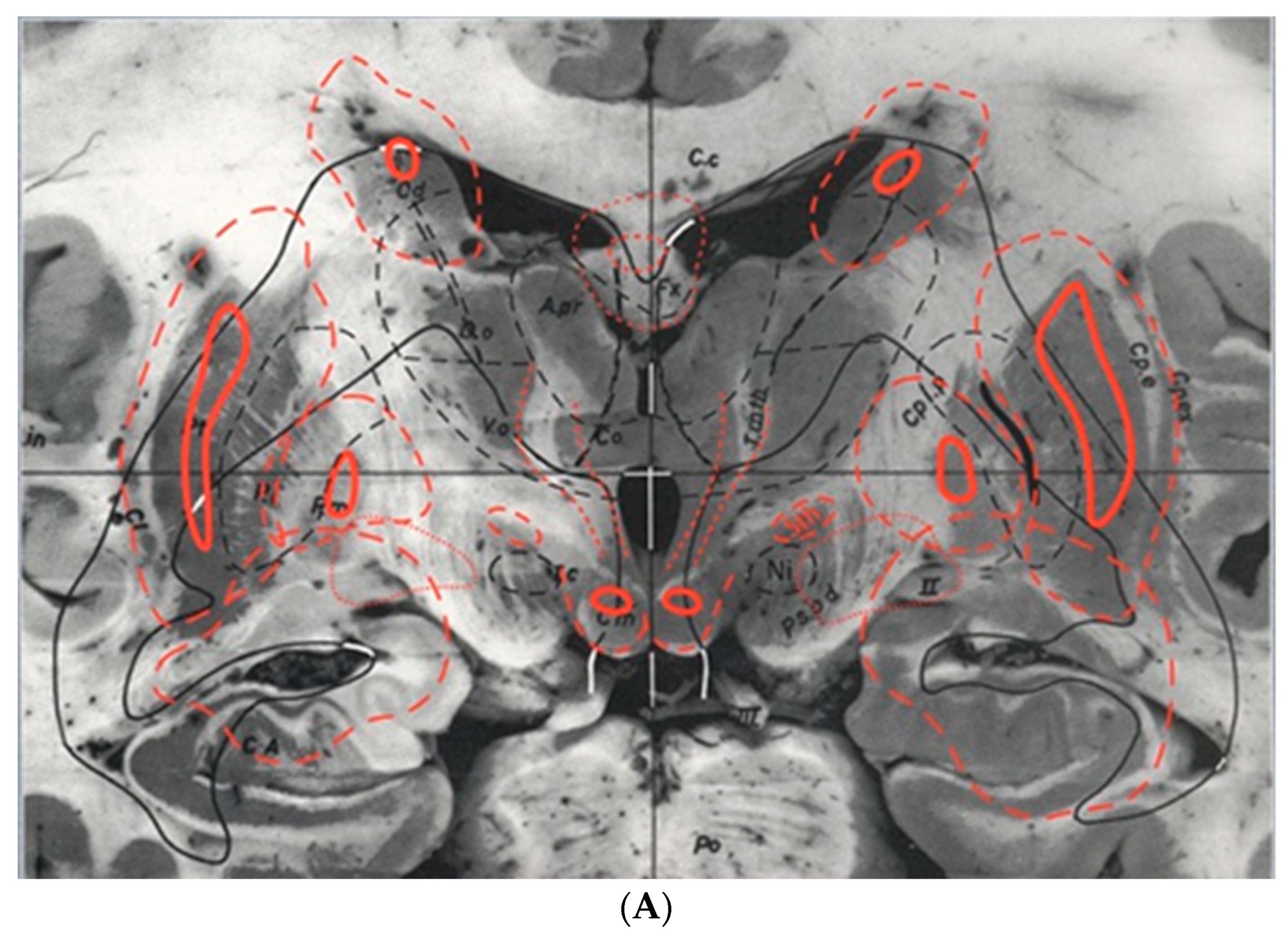
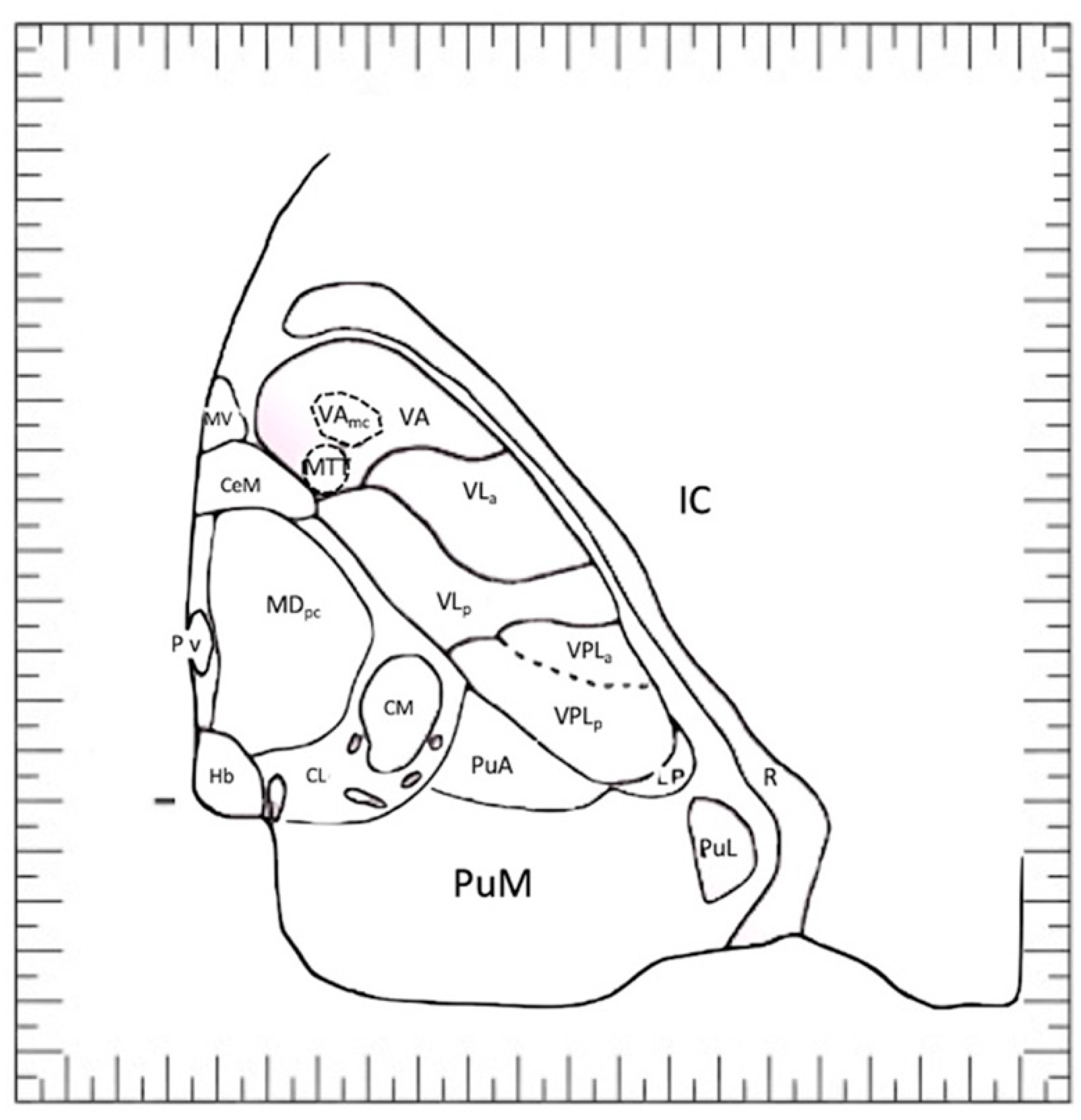
5. The ‘Schaltenbrand and Bailey’ Atlas
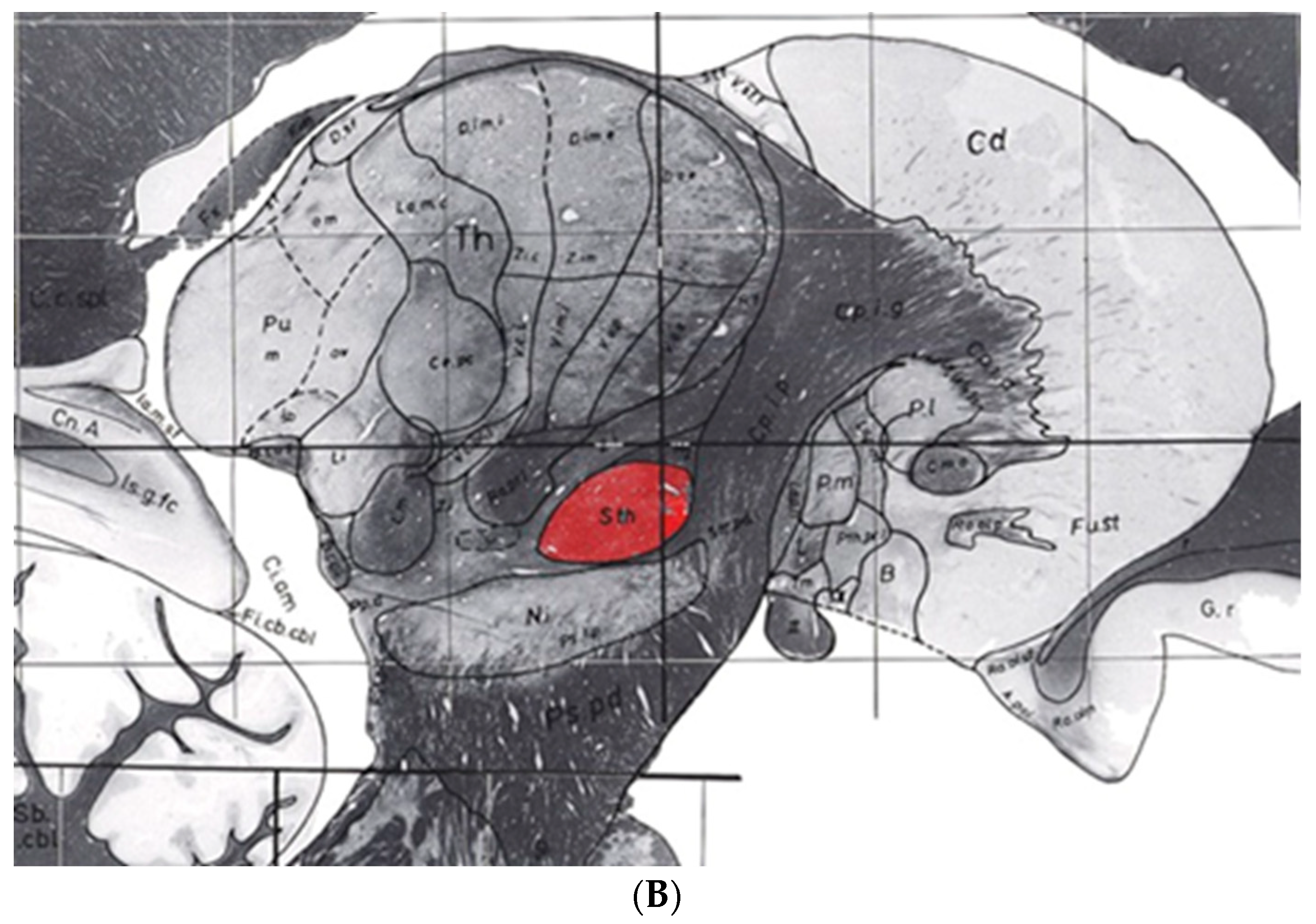
6. An Atlas Showing Variations in Human Diencephalon
7. The ‘Talairach’ and ‘Schaltenbrand’ Atlases of the CT Era
7.1. Consistencies and Inconsistencies of the Talairach and Tournoux 1988 and the Schaltenbrand and Wahren Atlases
7.2. The Talairach–Nowinski System
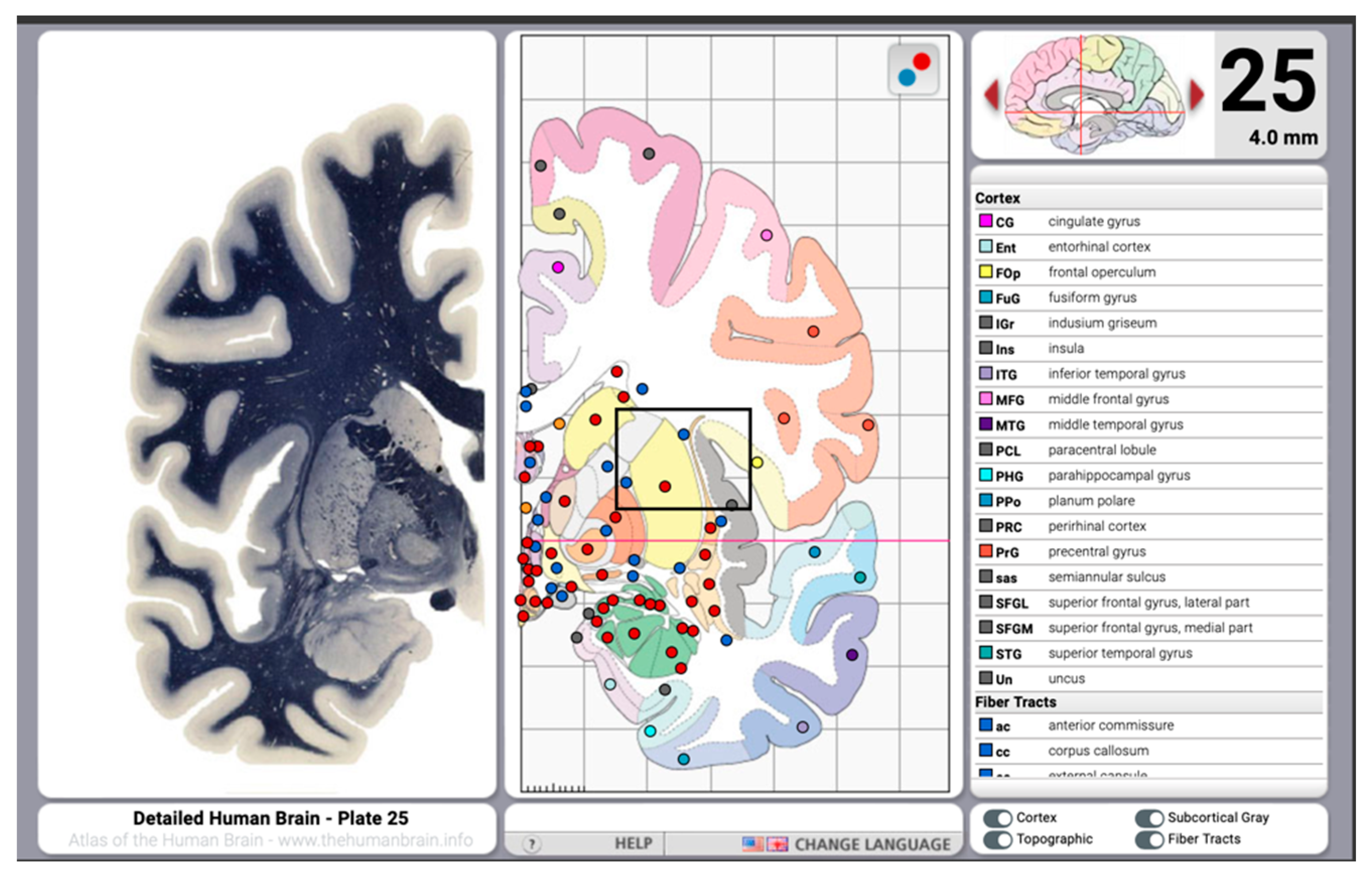
8. Digital Histological Atlases
9. Three-Dimensional Atlases
9.1. Early Computer-Based Atlases
9.2. Atlas-to-Scan Warping
9.3. Digital Atlases from MRI-Data
10. Three-Dimensional Rendered Atlases from Histological Sections and Multiatlas Collections
11. Probabilistic and Functional Atlases
12. Brain Atlas Applications
12.1. Education
12.2. Research
12.3. Clinical Applications
13. Ongoing Projects
14. Conclusions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Coffey, R. Printed Stereotactic Atlases, Review. In Textbook of Stereotactic and Functional Neurosurgery; Springer: Berlin/Heidelberg, Germany, 2009; pp. 347–371. [Google Scholar]
- Nowinski, W.L.; Thirunavuukarasuu, A.; Benabid, A. The Cerefy Clinical Brain Atlas: Enhanced Edition with Surgical Planning and Intraoperative Support; Thieme: New York, NY, USA, 2005. [Google Scholar]
- Nowinski, W.L.; Yang, G.L.; Yeo, T.T. Computer-Aided Stereotactic Functional Neurosurgery Enhanced by the Use of the Multiple Brain Atlas Database. IEEE Trans. Med. Imaging 2000, 19, 62–69. [Google Scholar] [CrossRef] [PubMed]
- Broca, P. About the articulation language site. Bull. Soc. Anth. 1861, 6, 330–357, 398–407. [Google Scholar]
- Jackson, J. Convulsive Spasms of the Right Hand and Arm Preceding Epileptic Seizures1. Med. Times Gaz. 1863, 1, 110–111. [Google Scholar]
- Reis, C.; Sankar, T.; Crusius, M.; Zabramski, J.; Deshmukh, P.; Rhoton, A.J.; Spetzler, R.; Preul, M. Comparative Study of Cranial Topographic Procedures: Broca’s Legacy toward Practical Brain Surgery. Neurosurgery 2008, 62, 294–310. [Google Scholar] [CrossRef]
- Ribas, G.C.; Yasuda, A.; Ribas, E.C.; Nishikuni, K.; Rodrigues, A.J., Jr. Surgical Anatomy of Microneurosurgical Sulcal Key Points. Neurosurgery 2006, 59 (Suppl. S4), ONS-177. [Google Scholar] [CrossRef] [PubMed]
- Broca, P. About the cranial-cerebral topography. Rev. Anthr. 1876, 5, 193–248. [Google Scholar]
- Turner, W. Relations of the Convolutions to the Skull. In The Functions of the Brain; Smith, Elder & Co.: London, UK, 1876. [Google Scholar]
- Reid, R. Observations on the Relation of the Principle Fissures and Convolutions of the Cerebrum to the Outer Surface of the Scalp. Lancet 1884, 2, 539–540. [Google Scholar] [CrossRef]
- Poirier, P. Cranial-Cerebral Topography; Lescrosnier et Babè: Paris, France, 1891. [Google Scholar]
- Chipault, A. Operatory Surgery of the Nervous System; Rueff et Cie: Paris, France, 1894. [Google Scholar]
- Taylor, E.; Haugton, W. Some Recent Researches on the Topography of the Convolutions and Fissures of the Brain. Trans. R. Acad. 1900, 18, 511–519. [Google Scholar] [CrossRef]
- Horsley, V.; Clarke, R. The Structure and Function of the Cerebellum Examined by a New Method. Brain 1908, 31, 124–154. [Google Scholar] [CrossRef]
- Paxinos, G.; Watson, C. The Rat Brain in Stereotaxic Coordinates, 4th ed.; Academic Press: London, UK, 1998. [Google Scholar]
- Spiegel, E.; Wycis, H. Stereoencephalotomy, Thalamotomy and Related Procedures. I. Methods and Stereotaxic Atlas of the Human Brain; Grune and Stratton: New York, NY, USA, 1952. [Google Scholar]
- Spiegel, E.; Wycis, H. Stereoencephalotomy. II. Clinical and Physiological Applications; Grune and Stratton: New York, NY, USA, 1962. [Google Scholar]
- Talairach, J.; David, M.; Tournoux, P.; Corredor, H.; Kvasina, T. Atlas d’Anatomie Stereotaxique. Repérage Radiologique Indirect Des Noyaux Gris Centraux Des Régions Mésencéphalo-Sous-Optique et Hypothalamique de l’homme; Masson: Paris, France, 1957. [Google Scholar]
- Schaltenbrand, G.; Bailey, P. Introduction to Stereotaxis with an Atlas of the Human Brain; Thieme: Stuttgart, Germany, 1959. [Google Scholar]
- Van Buren, J.; Borke, R. Variations and Connections of the Human Thalamus; Springer: Berlin/Heidelberg, Germany, 1972. [Google Scholar]
- Schaltenbrand, G.; Wahren, W. Atlas for Stereotaxy of the Human Brain; Thieme: Stuttgart, Germany, 1977. [Google Scholar]
- Talairach, J.; Tournoux, P. Co-Planar Stereotaxic Atlas of the Human Brain: 3-Dimensional Proportional System: An Approach to Cerebral Imaging; Thieme: Stuttgart, Germany, 1988. [Google Scholar]
- Nowinski, W.L. Computerized Brain Atlases for Surgery of Movement Disorders. Semin. Neurosurg. 2001, 12, 183–194. [Google Scholar] [CrossRef]
- Niemann, K.; van Nieuwenhofen, I. One Atlas-Three Anatomies: Relationships of the Schaltenbrand and Wahren Microscopic Data. Acta Neurochir. 1999, 141, 1025–1038. [Google Scholar] [CrossRef] [PubMed]
- Nowinski, W.L.; Liu, J.; Thirunavuukarasuu, A. Quantification and Visualization of Three-Dimensional Inconsistency of the Ventrointermediate Nucleus of the Thalamus in the Schaltenbrand-Wahren Brain Atlas. Acta Neurochir. 2008, 150, 647–653. [Google Scholar] [CrossRef] [PubMed]
- Nowinski, W.L.; Thirunavuukarasuu, A. Quantification of Spatial Consistency in the Talairach and Tournoux Stereotactic Atlas. Acta Neurochir. 2009, 151, 1207–1213. [Google Scholar] [CrossRef] [PubMed]
- Niemann, K.; Naujokat, C.; Pohl, G.; Wollner, C.; von Keyserlingk, D. Verification of the Schaltenbrand and Wahren Stereotactic Atlas. Acta Neurochir. 1994, 129, 72–81. [Google Scholar] [CrossRef]
- Nowinski, W.L. Modified Talairach Landmarks. Acta Neurochir. 2001, 143, 1045–1057. [Google Scholar] [CrossRef]
- Mai, J.; Paxinos, G.; Assheuer, J. Atlas of the Human Brain; Academic Press: New York, NY, USA, 2004. [Google Scholar]
- Morel, A. Stereotactic Atlas of the Human Thalamus and Basal Ganglia; Informa Healthcare: New York, NY, USA, 2007. [Google Scholar]
- Nowinski, W.L. Evolution of Human Brain Atlases in Terms of Content, Applications, Functionality, and Availability. Neuroinformatics 2021, 19, 1–22. [Google Scholar] [CrossRef]
- Kall, B.A.; Kelly, P.J.; Goerss, S.; Frieder, G. Methodology and Clinical Experience with Computed Tomography and a Computer-Resident Stereotactic Atlas. Neurosurgery 1985, 17, 400–407. [Google Scholar] [CrossRef]
- Yoshida, M. Creation of a Three-Dimensional Atlas by Interpolation from Schaltenbrand-Bailey’s Atlas. Appl. Neurophysiol. 1987, 50, 45–48. [Google Scholar] [CrossRef]
- St-Jean, P.; Sadikot, A.F.; Collins, L.; Clonda, D.; Kasrai, R.; Evans, A.C.; Peters, T.M. Automated Atlas Integration and Interactive Three-Dimensional Visualization Tools for Planning and Guidance in Functional Neurosurgery. IEEE Trans. Med. Imaging 1998, 17, 672–680. [Google Scholar] [CrossRef]
- Nowinski, W.L.; Fang, A.; Nguyen, B.T.; Raphel, J.K.; Jagannathan, L.; Raghavan, R.; Bryan, R.N.; Miller, G.A. Multiple Brain Atlas Database and Atlas-Based Neuroimaging System. Comput. Aided Surg. 1997, 2, 42–66. [Google Scholar] [CrossRef]
- Sudhyadhom, A.; Okun, M.S.; Foote, K.D.; Rahman, M.; Bova, F.J. A Three-Dimensional Deformable Brain Atlas for DBS Targeting. I. Methodology for Atlas Creation and Artifact Reduction. Open Neuroimag. J. 2012, 6, 92–98. [Google Scholar] [CrossRef]
- Bohm, C.; Greitz, T.; Kingsley, D.; Berggren, B.M.; Olsson, L. Adjustable Computerized Stereotaxic Brain Atlas for Transmission and Emission Tomography. AJNR Am. J. Neuroradiol. 1983, 4, 731–733. [Google Scholar]
- Greitz, T.; Bohm, C.; Holte, S.; Eriksson, L. A Computerized Brain Atlas: Construction, Anatomical Content, and Some Applications. J. Comput. Assist. Tomogr. 1991, 15, 26–38. [Google Scholar] [CrossRef]
- Bertrand, G.; Olivier, A.; Thompson, C.J. Computer Display of Stereotaxic Brain Maps and Probe Tracts. In Advances in Stereotactic and Functional Neurosurgery: Proceedings of the 1st Meeting of the European Society for Stereotactic and Functional Neurosurgery, Edinburgh 1972; Springer: Vienna, Austria, 1974; Supplement 21; pp. 235–243. [Google Scholar] [CrossRef]
- Lacadie, C.M.; Fulbright, R.K.; Rajeevan, N.; Constable, R.T.; Papademetris, X. More Accurate Talairach Coordinates for Neuroimaging Using Non-Linear Registration. Neuroimage 2008, 42, 717–725. [Google Scholar] [CrossRef]
- Cox, R.W. AFNI: Software for Analysis and Visualization of Functional Magnetic Resonance Neuroimages. Comput. Biomed. Res. 1996, 29, 162–173. [Google Scholar] [CrossRef]
- Evans, A.C.; Collins, D.L.; Mills, S.R.; Brown, E.D.; Kelly, R.L.; Peters, T.M. 3D Statistical Neuroanatomical Models from 305 MRI Volumes. In Proceedings of the 1993 IEEE Conference Record Nuclear Science Symposium and Medical Imaging Conference, San Francisco, CA, USA, 31 October–6 November 1993; IEEE: San Francisco, CA, USA, 1993; pp. 1813–1817. [Google Scholar]
- Ono, M.; Kubik, S.; Abernathey, C. Atlas of the Cerebral Sulci; Georg Thieme Verlag/Thieme Medical Publishers: Stuttgart, Germany, 1990. [Google Scholar]
- Nowinski, W.L.; Qian, G.; Bhanu Prakash, K.N.; Hu, Q.; Aziz, A. Fast Talairach Transformation for Magnetic Resonance Neuroimages. J. Comput. Assist. Tomogr. 2006, 30, 629–641. [Google Scholar] [CrossRef]
- Brett, M.; Christoff, K.; Cusack, R.; Lancaster, J. Using the Talairach Atlas with the MN1 Template. NeuroImage 2001, 13, 85. [Google Scholar] [CrossRef]
- Carmack, P.S.; Spence, J.; Gunst, R.F.; Schucany, W.R.; Woodward, W.A.; Haley, R.W. Improved Agreement between Talairach and MNI Coordinate Spaces in Deep Brain Regions. Neuroimage 2004, 22, 367–371. [Google Scholar] [CrossRef]
- Chau, W.; McIntosh, A.R. The Talairach Coordinate of a Point in the MNI Space: How to Interpret It. Neuroimage 2005, 25, 408–416. [Google Scholar] [CrossRef]
- Lancaster, J.; Summerlin, J.; Rainey, L.; Freitas, C.; Fox, P. The Talairach Daemon a Database Server for Talairach Atlas Labels. Neuroimage 1997, 5, S633. [Google Scholar]
- Lucerna, S.; Salpietro, F.; Alafaci, C.; Tomasello, F. An in Vivo Atlas of Deep Brain Structures; Springer: Vienna, Austria, 2002. [Google Scholar]
- Yoshida, M. Three-Dimensional Maps by Interpolation from the Schaltenbrand and Bailey Atlas. In Computers in Stereotactic Neurosurgery; Blackwell: Boston, MA, USA, 1992; pp. 143–152. [Google Scholar]
- Talairach, J.; Tournoux, P. Referentially Oriented Cerebral MRI Anatomy: Atlas of Stereotaxic Anatomical Correlations for Gray and White Matter; Thieme: Stuttgart, Germany, 1993. [Google Scholar]
- Nowinski, W.L.; Bryan, R.N.; Raghavan, R. The Electronic Clinical Brain Atlas: Multiplanar Navigation of the Human Brain; Thieme: New York, NY, USA, 1997. [Google Scholar]
- Nowinski, W.L.; Thirunavuukarasuu, A. The Cerefy Clinical Brain Atlas on CD-ROM; Thieme: New York, NY, USA, 2004. [Google Scholar]
- Nowinski, W.L.; Thirunavuukarasuu, A.; Kennedy, D. Brain Atlas for Functional Imaging: Clinical and Research Applications; Thieme: New York, NY, USA, 2000. [Google Scholar]
- Nowinski, W.L. Anatomical and Probabilistic Functional Atlases in Stereotactic and Functional Neurosurgery. In Textbook of Stereotactic and Functional Neurosurgery; Springer: Berlin/Heidelberg, Germany, 2009; pp. 395–442. [Google Scholar]
- Nowinski, W.L. Co-Registration of the Schaltenbrand-Wahren Microseries with the Probabilistic Functional Atlas. Ster. Funct. Neurosurg. 2004, 82, 142–146. [Google Scholar] [CrossRef]
- Nowinski, W.L.; Liu, J.; Thirunavuukarasuu, A. Quantification and Visualization of the Three-Dimensional Inconsistency of the Subthalamic Nucleus in the Schaltenbrand-Wahren Brain Atlas. Ster. Funct. Neurosurg. 2006, 84, 46–55. [Google Scholar] [CrossRef]
- Yelnik, J.; Bardinet, E.; Dormont, D.; Malandain, G.; Ourselin, S.; Tandé, D.; Karachi, C.; Ayache, N.; Cornu, P.; Agid, Y. A Three-Dimensional, Histological and Deformable Atlas of the Human Basal Ganglia. I. Atlas Construction Based on Immunohistochemical and MRI Data. Neuroimage 2007, 34, 618–638. [Google Scholar] [CrossRef]
- Thompson, C.J.; Hardy, T.; Bertrand, G. A System for Anatomical and Functional Mapping of the Human Thalamus. Comput. Biomed. Res. 1977, 10, 9–24. [Google Scholar] [CrossRef]
- Tasker, R.; Organ, L.; Hawrylyshyn, P. The Thalamus and Midbrain of Man: A Physiological Atlas Using Electrical Stimulation; Charles C. Thomas: Springfield, IL, USA, 1982. [Google Scholar]
- Giorgi, C.; Cerchiari, U.; Broggi, G.; Birk, P.; Struppeler, A. Digital Image Processing to Handle Neuroanatomical Information and Neurophysiological Data. Appl. Neurophysiol. 1985, 48, 30–33. [Google Scholar] [CrossRef]
- Yoshida, M.; Okada, K.; Nagase, A.; Kuga, S.; Shirahama, M.; Watanabe, M.; Kuramoto, S. Neurophysiological Atlas of the Human Thalamus and Adjacent Structures. Computer-Assisted Mapping. Appl. Neurophysiol. 1982, 45, 406–409. [Google Scholar] [CrossRef]
- Bertrand, G.; Blundell, J.; Musella, R. Electrical Stimulation of the Internal Capsule and Neighboring Structures During Stereotactic Procedures; Harvey Cushing Soc.: Philadelphia, PA, SUA, 1963. [Google Scholar]
- Finnis, K.W.; Starreveld, Y.P.; Parrent, A.G.; Sadikot, A.F.; Peters, T.M. Three-Dimensional Database of Subcortical Electrophysiology for Image-Guided Stereotactic Functional Neurosurgery. IEEE Trans. Med. Imaging 2003, 22, 93–104. [Google Scholar] [CrossRef]
- Nowinski, W.L.; Belov, D.; Benabid, A.L. An Algorithm for Rapid Calculation of a Probabilistic Functional Atlas of Subcortical Structures from Electrophysiological Data Collected during Functional Neurosurgery Procedures. Neuroimage 2003, 18, 143–155. [Google Scholar] [CrossRef]
- Nowinski, W.L.; Belov, D.; Pollak, P.; Benabid, A.L. A Probabilistic Functional Atlas of the Human Subthalamic Nucleus. Neuroinformatics 2004, 2, 381–398. [Google Scholar] [CrossRef]
- Nowinski, W.L.; Belov, D.; Thirunavuukarasuu, A.; Benabid, A.L. A Probabilistic Functional Atlas of the VIM Nucleus Constructed from Pre-, Intra- and Postoperative Electrophysiological and Neuroimaging Data Acquired during the Surgical Treatment of Parkinson’s Disease Patients. Ster. Funct. Neurosurg. 2005, 83, 190–196. [Google Scholar] [CrossRef]
- Nowinski, W.L.; Belov, D.; Benabid, A.-L. A Community-Centric Internet Portal for Stereotactic and Functional Neurosurgery with a Probabilistic Functional Atlas. Ster. Funct. Neurosurg. 2002, 79, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Haegelen, C.; Baumgarten, C.; Houvenaghel, J.-F.; Zhao, Y.; Péron, J.; Drapier, S.; Jannin, P.; Morandi, X. Functional Atlases for Analysis of Motor and Neuropsychological Outcomes after Medial Globus Pallidus and Subthalamic Stimulation. PLoS ONE 2018, 13, e0200262. [Google Scholar] [CrossRef]
- Nowinski, W.L. Advances in Neuroanatomy through Brain Atlasing. Anatomia 2023, 2, 28–42. [Google Scholar] [CrossRef]
- Nowinski, W.L. NOWinBRAIN: A Large, Systematic, and Extendable Repository of 3D Reconstructed Images of a Living Human Brain Cum Head and Neck. J. Digit. Imaging 2022, 35, 98–114. [Google Scholar] [CrossRef]
- Nowinski, W.L. Bridging Neuroradiology and Neuroanatomy: NOWinBRAIN-a Repository with Sequences of Correlated and Labeled Planar-Surface Neuroimages. Neuroradiol. J. 2023, 36, 94–103. [Google Scholar] [CrossRef]
- Aljabar, P.; Heckemann, R.A.; Hammers, A.; Hajnal, J.V.; Rueckert, D. Multi-Atlas Based Segmentation of Brain Images: Atlas Selection and Its Effect on Accuracy. Neuroimage 2009, 46, 726–738. [Google Scholar] [CrossRef]
- Artaechevarria, X.; Munoz-Barrutia, A.; Ortiz-de-Solorzano, C. Combination Strategies in Multi-Atlas Image Segmentation: Application to Brain MR Data. IEEE Trans. Med. Imaging 2009, 28, 1266–1277. [Google Scholar] [CrossRef]
- Labra, N.; Guevara, P.; Duclap, D.; Houenou, J.; Poupon, C.; Mangin, J.-F.; Figueroa, M. Fast Automatic Segmentation of White Matter Streamlines Based on a Multi-Subject Bundle Atlas. Neuroinformatics 2017, 15, 71–86. [Google Scholar] [CrossRef]
- Li, X.; Chen, L.; Kutten, K.; Ceritoglu, C.; Li, Y.; Kang, N.; Hsu, J.T.; Qiao, Y.; Wei, H.; Liu, C.; et al. Multi-Atlas Tool for Automated Segmentation of Brain Gray Matter Nuclei and Quantification of Their Magnetic Susceptibility. Neuroimage 2019, 191, 337–349. [Google Scholar] [CrossRef]
- Lötjönen, J.M.; Wolz, R.; Koikkalainen, J.R.; Thurfjell, L.; Waldemar, G.; Soininen, H.; Rueckert, D. Alzheimer’s Disease Neuroimaging Initiative Fast and Robust Multi-Atlas Segmentation of Brain Magnetic Resonance Images. Neuroimage 2010, 49, 2352–2365. [Google Scholar] [CrossRef]
- Zaffino, P.; Ciardo, D.; Raudaschl, P.; Fritscher, K.; Ricotti, R.; Alterio, D.; Marvaso, G.; Fodor, C.; Baroni, G.; Amato, F.; et al. Multi Atlas Based Segmentation: Should We Prefer the Best Atlas Group over the Group of Best Atlases? Phys. Med. Biol. 2018, 63, 12NT01. [Google Scholar] [CrossRef] [PubMed]
- Makowski, C.; van der Meer, D.; Dong, W.; Wang, H.; Wu, Y.; Zou, J.; Liu, C.; Rosenthal, S.B.; Hagler, D.J.; Fan, C.C.; et al. Discovery of Genomic Loci of the Human Cerebral Cortex Using Genetically Informed Brain Atlases. Science 2022, 375, 522–528. [Google Scholar] [CrossRef] [PubMed]
- Nowinski, W.L.; Chua, B.C.; Volkau, I.; Puspitasari, F.; Marchenko, Y.; Runge, V.M.; Knopp, M.V. Simulation and Assessment of Cerebrovascular Damage in Deep Brain Stimulation Using a Stereotactic Atlas of Vasculature and Structure Derived from Multiple 3- and 7-Tesla Scans. J. Neurosurg. 2010, 113, 1234–1241. [Google Scholar] [CrossRef]
- Nowinski, W.L.; Qian, G.; Kirgaval Nagaraja, B.P.; Thirunavuukarasuu, A.; Hu, Q.; Ivanov, N.; Parimal, A.S.; Runge, V.M.; Beauchamp, N.J. Analysis of Ischemic Stroke MR Images by Means of Brain Atlases of Anatomy and Blood Supply Territories. Acad. Radiol. 2006, 13, 1025–1034. [Google Scholar] [CrossRef]
- Nowinski, W.L. Human Brain Atlases in Stroke Management. Neuroinformatics 2020, 18, 549–567. [Google Scholar] [CrossRef] [PubMed]
- Nowinski, W.L.; Chua, B.C. Bridging Neuroanatomy, Neuroradiology and Neurology: Three-Dimensional Interactive Atlas of Neurological Disorders. Neuroradiol. J. 2013, 26, 252–262. [Google Scholar] [CrossRef]
- Sim, K.; Yang, G.L.; Loh, D.; Poon, L.Y.; Sitoh, Y.Y.; Verma, S.; Keefe, R.; Collinson, S.; Chong, S.A.; Heckers, S.; et al. White Matter Abnormalities and Neurocognitive Deficits Associated with the Passivity Phenomenon in Schizophrenia: A Diffusion Tensor Imaging Study. Psychiatry Res. 2009, 172, 121–127. [Google Scholar] [CrossRef]
- Toga, A.W.; Thompson, P.M.; Mori, S.; Amunts, K.; Zilles, K. Towards Multimodal Atlases of the Human Brain. Nat. Rev. Neurosci. 2006, 7, 952–966. [Google Scholar] [CrossRef] [PubMed]
- Van Essen, D.C.; Smith, S.M.; Barch, D.M.; Behrens, T.E.J.; Yacoub, E.; Ugurbil, K. WU-Minn HCP Consortium The WU-Minn Human Connectome Project: An Overview. Neuroimage 2013, 80, 62–79. [Google Scholar] [CrossRef]
- Sunkin, S.M.; Ng, L.; Lau, C.; Dolbeare, T.; Gilbert, T.L.; Thompson, C.L.; Hawrylycz, M.; Dang, C. Allen Brain Atlas: An Integrated Spatio-Temporal Portal for Exploring the Central Nervous System. Nucleic Acids Res. 2013, 41, D996–D1008. [Google Scholar] [CrossRef]
- Amunts, K.; Lepage, C.; Borgeat, L.; Mohlberg, H.; Dickscheid, T.; Rousseau, M.-É.; Bludau, S.; Bazin, P.-L.; Lewis, L.B.; Oros-Peusquens, A.-M.; et al. BigBrain: An Ultrahigh-Resolution 3D Human Brain Model. Science 2013, 340, 1472–1475. [Google Scholar] [CrossRef]
- Assaf, Y.; Alexander, D.C.; Jones, D.K.; Bizzi, A.; Behrens, T.E.J.; Clark, C.A.; Cohen, Y.; Dyrby, T.B.; Huppi, P.S.; Knoesche, T.R.; et al. The CONNECT Project: Combining Macro- and Micro-Structure. Neuroimage 2013, 80, 273–282. [Google Scholar] [CrossRef] [PubMed]
- Jiang, T. Brainnetome: A New -Ome to Understand the Brain and Its Disorders. Neuroimage 2013, 80, 263–272. [Google Scholar] [CrossRef] [PubMed]
- Jorgenson, L.A.; Newsome, W.T.; Anderson, D.J.; Bargmann, C.I.; Brown, E.N.; Deisseroth, K.; Donoghue, J.P.; Hudson, K.L.; Ling, G.S.F.; MacLeish, P.R.; et al. The BRAIN Initiative: Developing Technology to Catalyse Neuroscience Discovery. Philos. Trans. R. Soc. Lond. B Biol. Sci. 2015, 370, 20140164. [Google Scholar] [CrossRef] [PubMed]
- Markram, H.; Muller, E.; Ramaswamy, S.; Reimann, M.W.; Abdellah, M.; Sanchez, C.A.; Ailamaki, A.; Alonso-Nanclares, L.; Antille, N.; Arsever, S.; et al. Reconstruction and Simulation of Neocortical Microcircuitry. Cell 2015, 163, 456–492. [Google Scholar] [CrossRef]
- Amunts, K.; Ebell, C.; Muller, J.; Telefont, M.; Knoll, A.; Lippert, T. The Human Brain Project: Creating a European Research Infrastructure to Decode the Human Brain. Neuron 2016, 92, 574–581. [Google Scholar] [CrossRef]
- Zuo, X.-N.; He, Y.; Betzel, R.F.; Colcombe, S.; Sporns, O.; Milham, M.P. Human Connectomics across the Life Span. Trends Cogn. Sci. 2017, 21, 32–45. [Google Scholar] [CrossRef]
- Sadato, N.; Morita, K.; Kasai, K.; Fukushi, T.; Nakamura, K.; Nakazawa, E.; Okano, H.; Okabe, S. Neuroethical Issues of the Brain/MINDS Project of Japan. Neuron 2019, 101, 385–389. [Google Scholar] [CrossRef]
- Chin, A.-L.; Yang, S.-M.; Chen, H.-H.; Li, M.-T.; Lee, T.-T.; Chen, Y.-J.; Lee, T.-K.; Petibois, C.; Cai, X.; Low, C.-M.; et al. A Synchrotron X-Ray Imaging Strategy to Map Large Animal Brains. Chin. J. Phys. 2020, 65, 24–32. [Google Scholar] [CrossRef]
- Chen, S.; He, Z.; Han, X.; He, X.; Li, R.; Zhu, H.; Zhao, D.; Dai, C.; Zhang, Y.; Lu, Z.; et al. How Big Data and High-Performance Computing Drive Brain Science. Genom. Proteom. Bioinform. 2019, 17, 381–392. [Google Scholar] [CrossRef]














Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Conti, A.; Gambadauro, N.M.; Mantovani, P.; Picciano, C.P.; Rosetti, V.; Magnani, M.; Lucerna, S.; Tuleasca, C.; Cortelli, P.; Giannini, G. A Brief History of Stereotactic Atlases: Their Evolution and Importance in Stereotactic Neurosurgery. Brain Sci. 2023, 13, 830. https://doi.org/10.3390/brainsci13050830
Conti A, Gambadauro NM, Mantovani P, Picciano CP, Rosetti V, Magnani M, Lucerna S, Tuleasca C, Cortelli P, Giannini G. A Brief History of Stereotactic Atlases: Their Evolution and Importance in Stereotactic Neurosurgery. Brain Sciences. 2023; 13(5):830. https://doi.org/10.3390/brainsci13050830
Chicago/Turabian StyleConti, Alfredo, Nicola Maria Gambadauro, Paolo Mantovani, Canio Pietro Picciano, Vittoria Rosetti, Marcello Magnani, Sebastiano Lucerna, Constantin Tuleasca, Pietro Cortelli, and Giulia Giannini. 2023. "A Brief History of Stereotactic Atlases: Their Evolution and Importance in Stereotactic Neurosurgery" Brain Sciences 13, no. 5: 830. https://doi.org/10.3390/brainsci13050830
APA StyleConti, A., Gambadauro, N. M., Mantovani, P., Picciano, C. P., Rosetti, V., Magnani, M., Lucerna, S., Tuleasca, C., Cortelli, P., & Giannini, G. (2023). A Brief History of Stereotactic Atlases: Their Evolution and Importance in Stereotactic Neurosurgery. Brain Sciences, 13(5), 830. https://doi.org/10.3390/brainsci13050830