


Transcranial Direct Current Stimulation (tDCS) Ameliorates Stress-Induced Sleep Disruption via Activating Infralimbic-Ventrolateral Preoptic Projections


Abstract
:1. Introduction
- (a)
- Current Intensity and Duration: Many studies suggest that the impact of current duration and intensity on outcomes is not necessarily linear [32]. An example study indicated that excessively long stimulation times may affect the expected effects of polarity, such as anodal stimulation causing inhibition of cortical excitability or vice versa [33];
- (b)
- Electrode Placement: For tDCS to form a complete circuit, two electrodes are required. The target electrode (anode or cathode) is placed on the specific brain region to be modulated, while the reference or counter electrode (the one with the opposite polarity to the target electrode) is placed in the corresponding area based on the hypothesis or target configuration [34,35,36]. When placing the reference electrode, it is necessary to consider the distance between the two electrodes so that the current can pass through the target brain region. If the distance between the two electrodes is too close, it may lead to a shunting effect, causing the current to dissipate through the scalp [35]. In clinical applications, especially in the field of human psychiatric disorders, selected brain stimulation areas often include the motor cortex and dorsolateral prefrontal cortex. Some research results have shown that after stimulating these regions, patients experience significant improvements in sleep quality [37] and related indicators such as sleep efficiency or number of awakenings [29,38];
- (c)
- Current Density and Charge Density: Current density (A/m2) is the amount of electric charge passing through a unit area during a given period, while charge density (C/m2) is the multiplication of current density and the duration. In a study investigating brain tissue damage in rats following tDCS stimulation, the findings suggested that brain tissue can be harmed when both current density and charge density surpass a certain threshold [39]. This implies that even with high current density if the duration is very short, the impact on the tissue may be weak. Conversely, when applying extremely low current density and prolonging the duration, even with a high final charge density, it does not result in tissue damage. Factors influencing current density also include the electrode’s contact area with the skull, essentially the size of the electrode. Therefore, when setting the current intensity, adjustments should be made based on the electrode’s contact area used rather than using the same current intensity directly. Furthermore, reducing the impedance between the electrode and the contact surface is also of significant importance [35]. In most experiments, the primary method for ensuring conductivity involves the use of saline, while some studies employ conductive gel [34]. However, in our experiment, the extremely low current we applied led to significant resistance when using saline as the conducting solution. This resistance prevented the formation of a complete circuit. As a result, we opted for chloride-free conductive gel. This not only ensured the flow of current but also prevented any potential chemical reactions between the electrodes and the conducting solution.
2. Materials and Methods
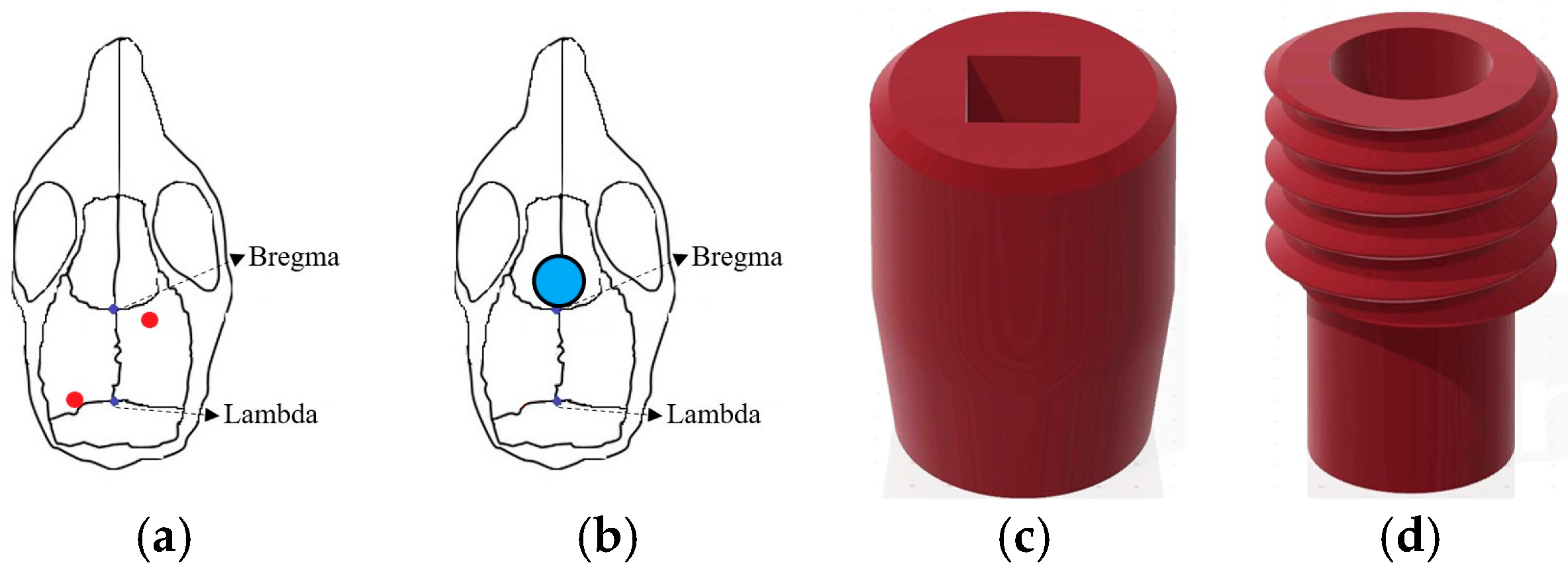
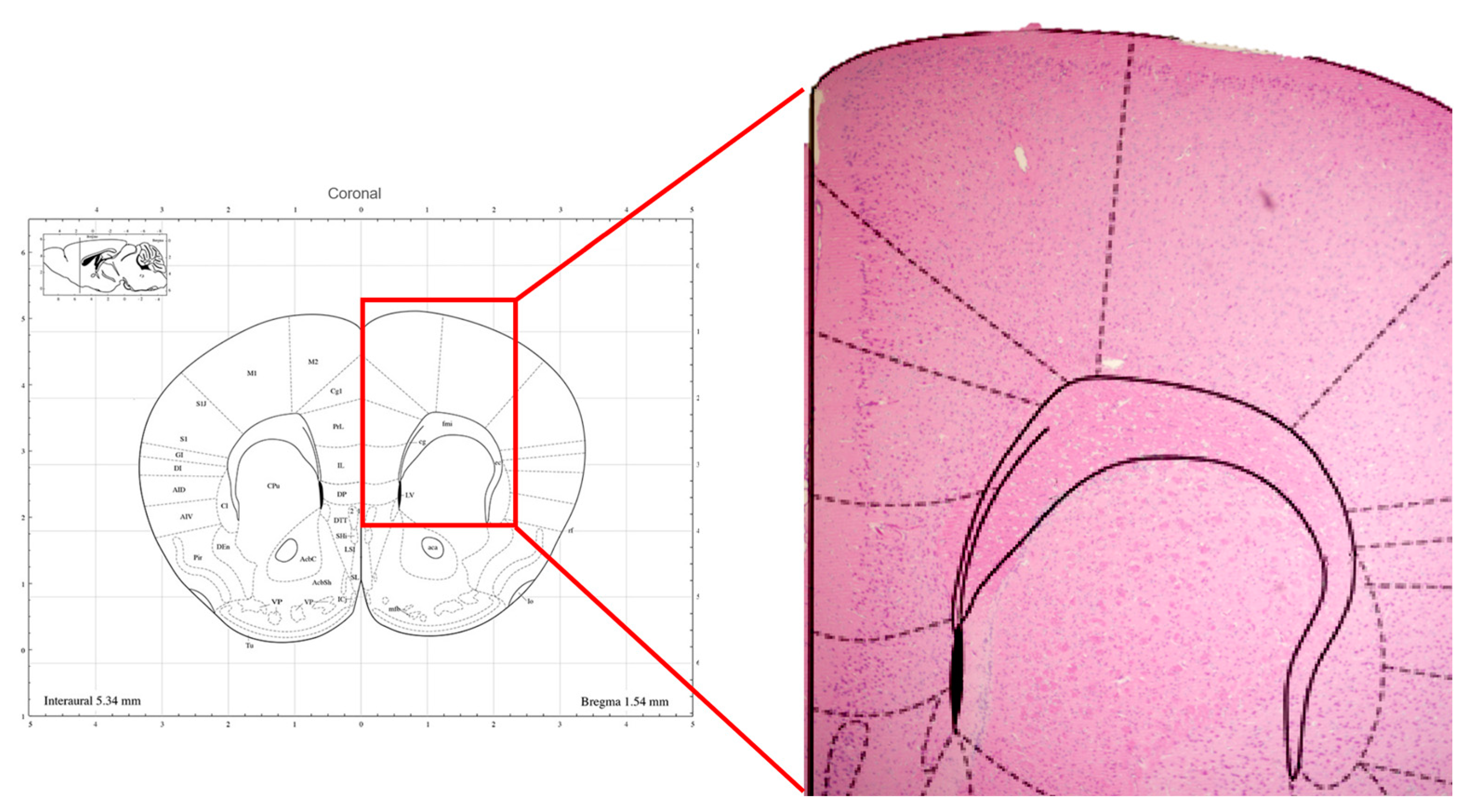
2.1. Electroencephalography (EEG) and tDCS Electrodes Implantation
2.2. Microinjection of Fluorogold
2.3. Designer Receptors Exclusively Activated by Designer Drugs (DREADDs) and Cre/DIO System
2.4. EEG Recording and Analysis
2.5. Brain Slice Preparation
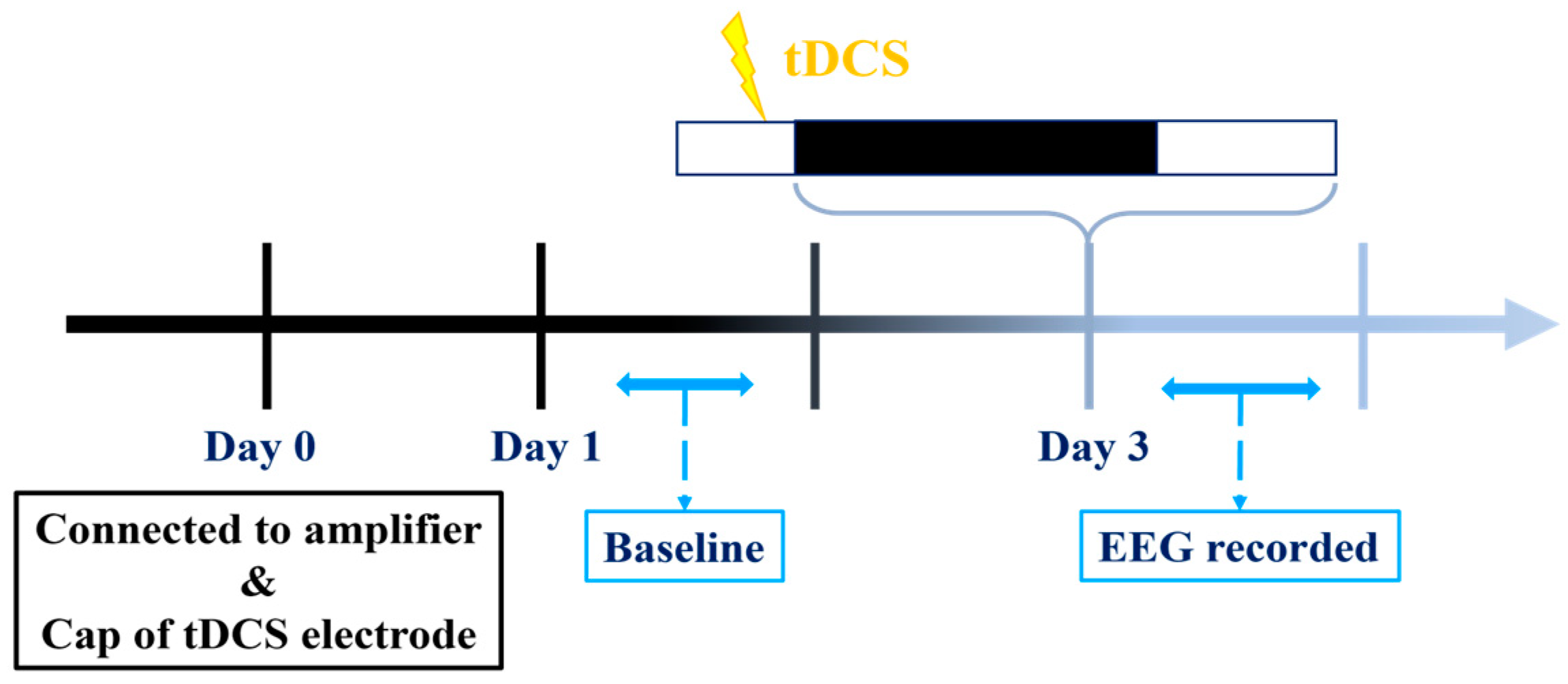
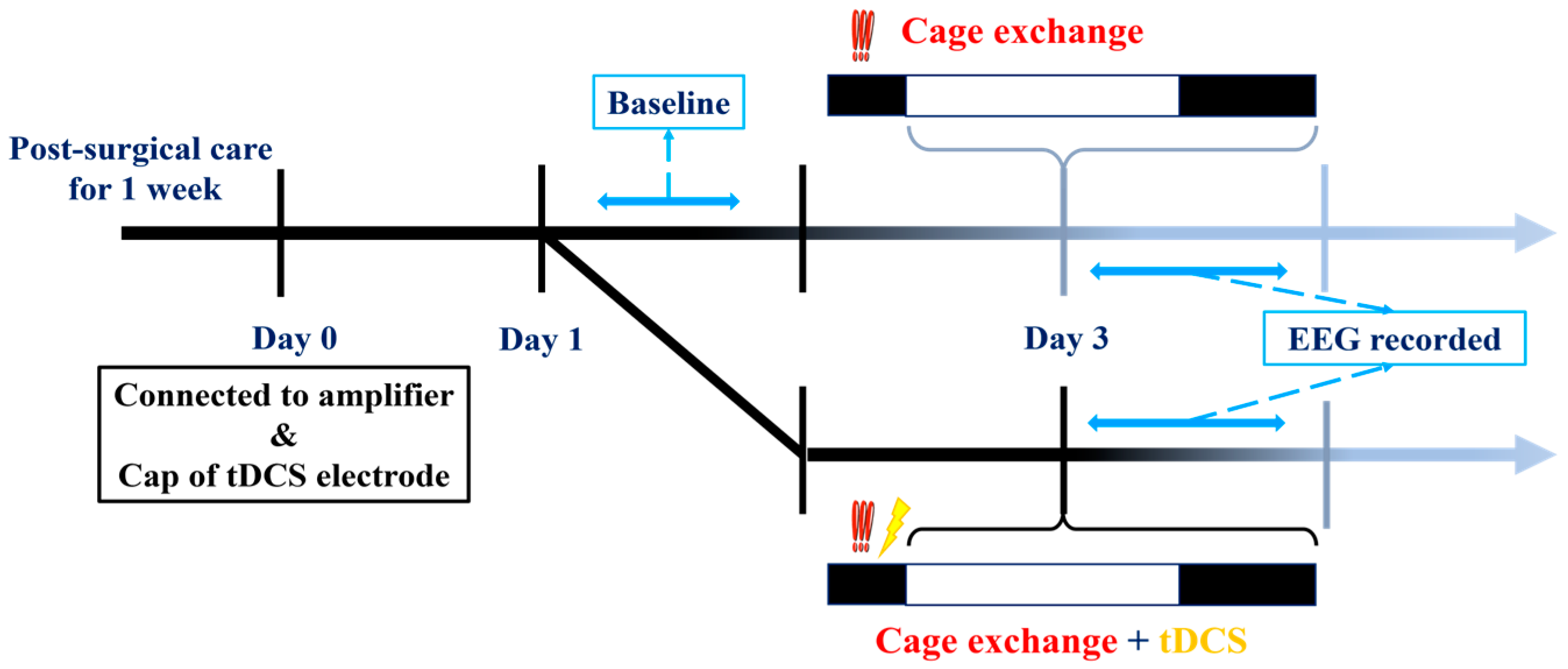
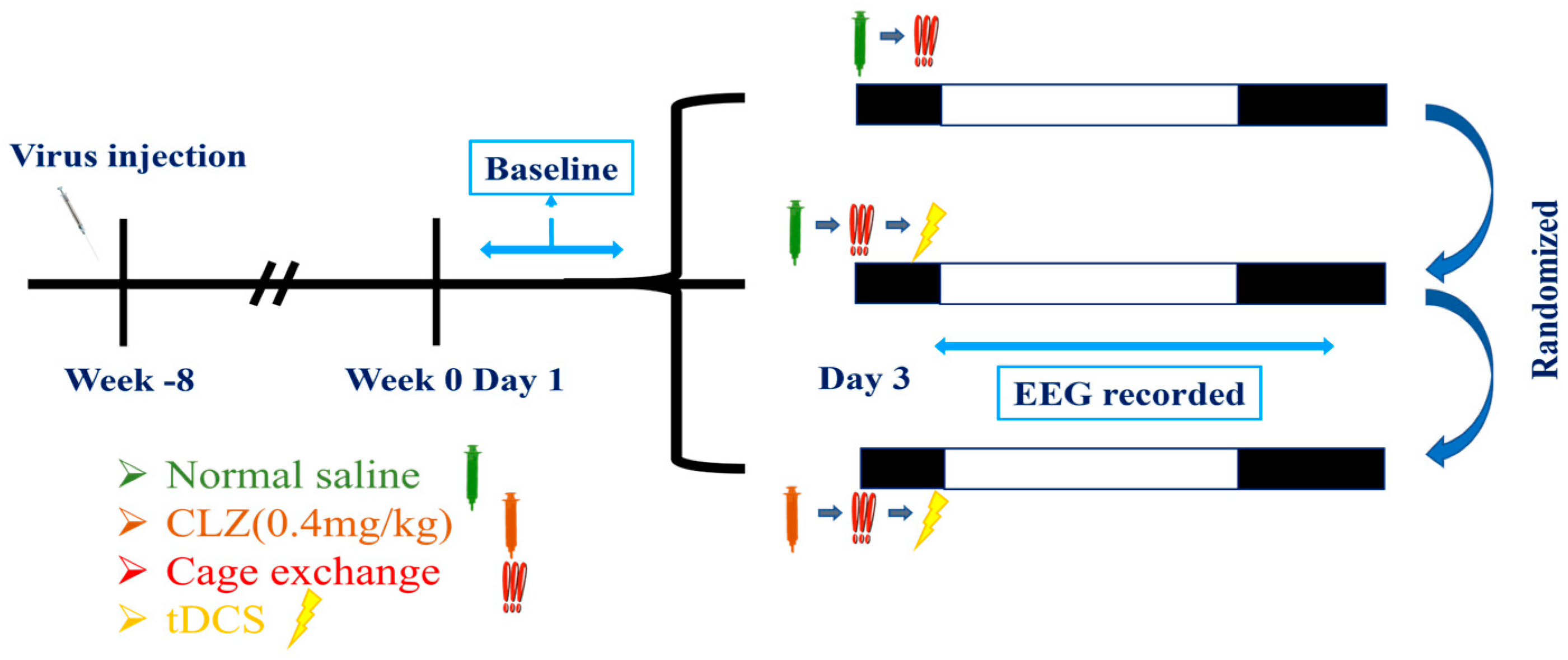
2.6. Experimental Protocols
2.6.1. Establishing a Suitable tDCS Parameters
2.6.2. tDCS Effects on Stress-Induced Insomnia Model
2.6.3. Validate the Projection from IL to VLPO
2.6.4. Determine the Effect of tDCS on Stress-Induced Insomnia Is Mediated by the IL-VLPO Pathway
- The first group received an intraperitoneal injection of saline. After a 15 min interval, the mouse was relocated to a cage already occupied by another mouse to induce insomnia. EEG recording commenced after tDCS stimulation;
- The second group received an intraperitoneal injection of clozapine (clz, 0.4 mg/kg) to inhibit the activation of the IL to VLPO pathway [55]. Fifteen minutes after the intraperitoneal injection, we conducted cage exchanging and initiated tDCS stimulation with parameters consistent with the previous settings. EEG recording began after completing the stimulation;
- The third group received an intraperitoneal injection of saline. After a 15 min interval, they underwent cage exchanging, followed by EEG recording.
2.7. Statistical Analysis
3. Results
3.1. Optimal Configuration of tDCS for Mice
3.1.1. Parameters of tDCS
3.1.2. The Influence of tDCS on Sleep in Naïve Mice
3.1.3. Effects of tDCS on Sleep Architecture and Quality
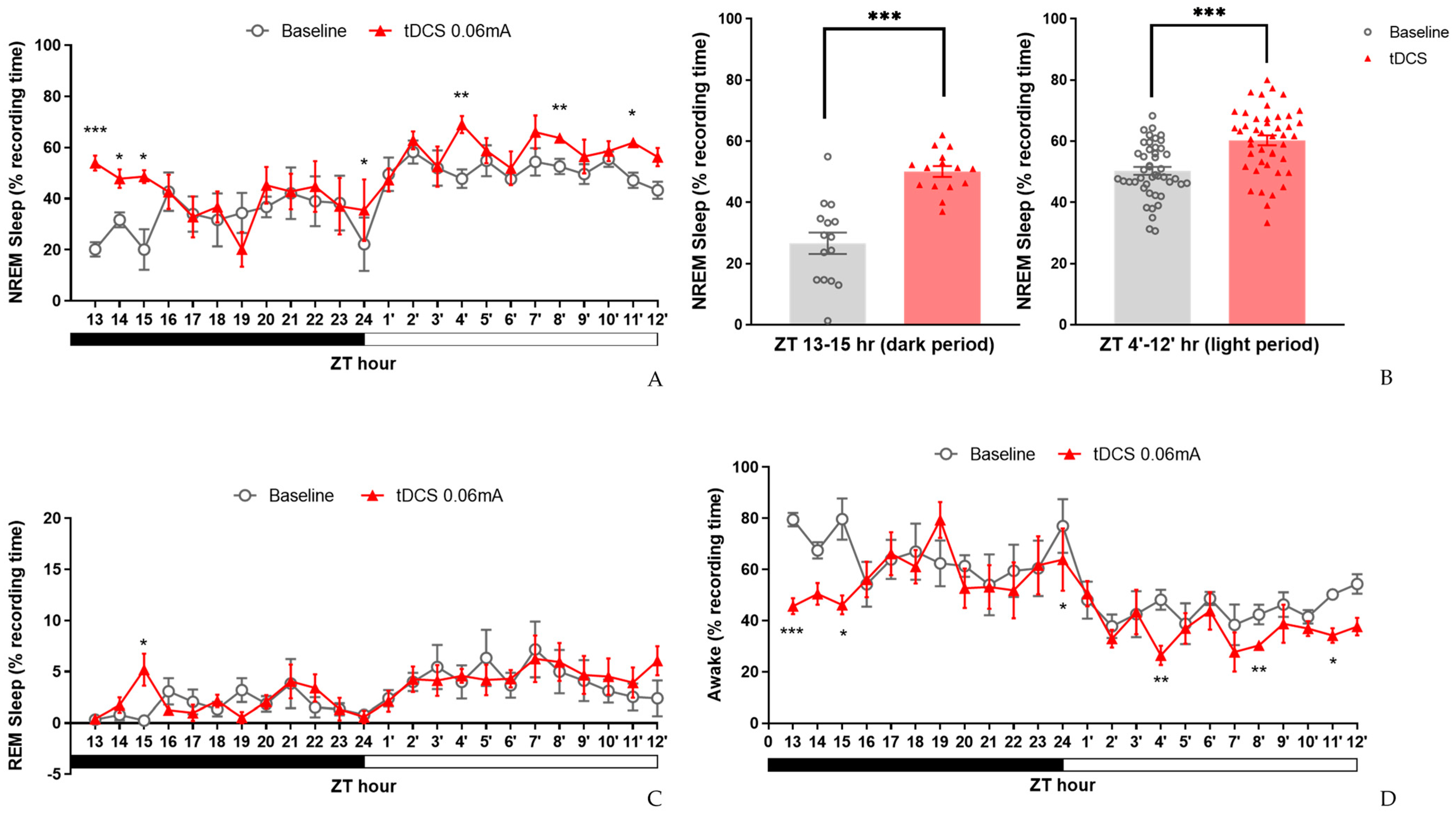
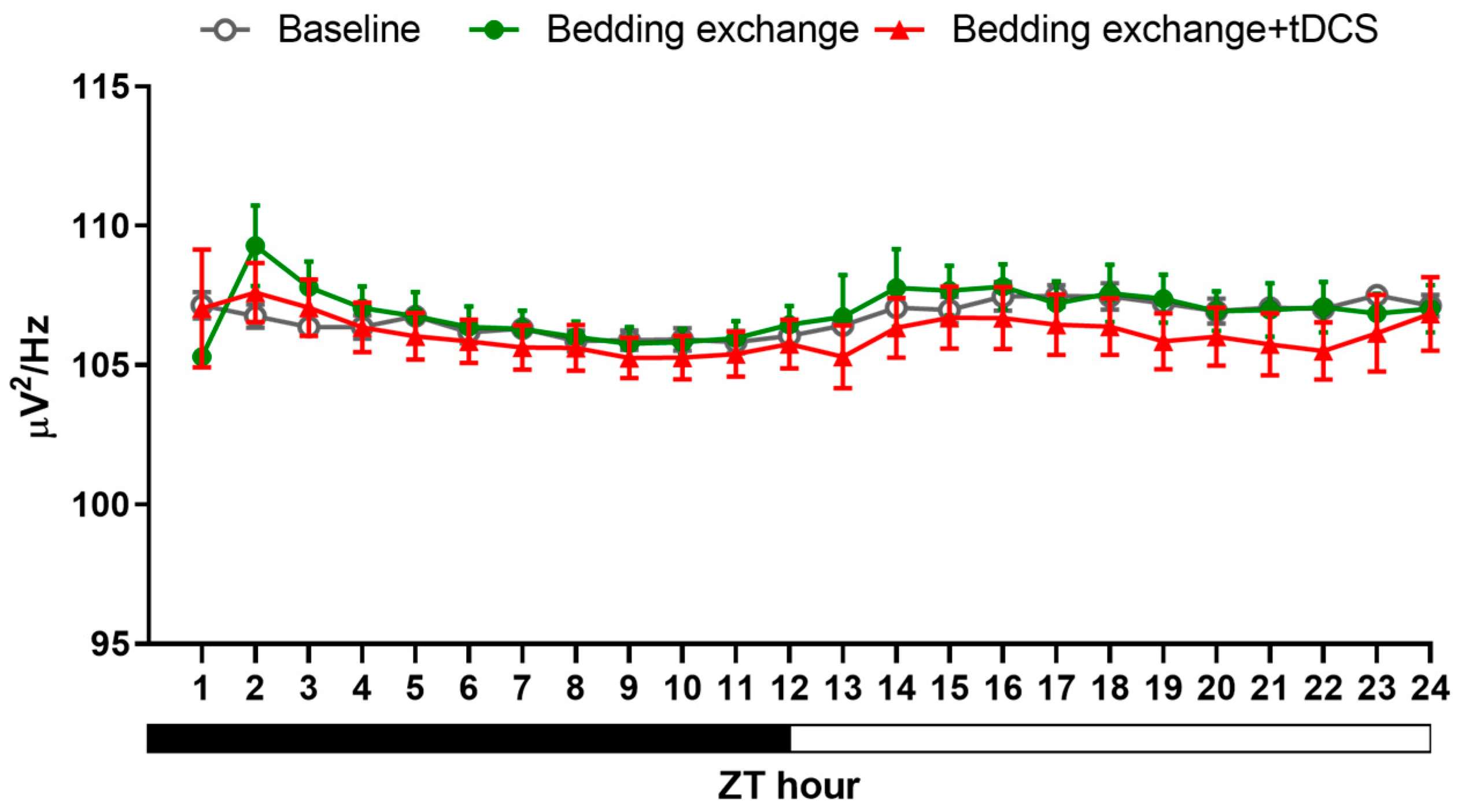
3.2. The Therapeutic Effect of tDCS in Stress-Induced Insomnia Mice
3.2.1. Sleep Modifications in Stress-Induced Insomnia Mice following tDCS Intervention
3.2.2. Alterations of Sleep Architectures between Groups during Different Phases
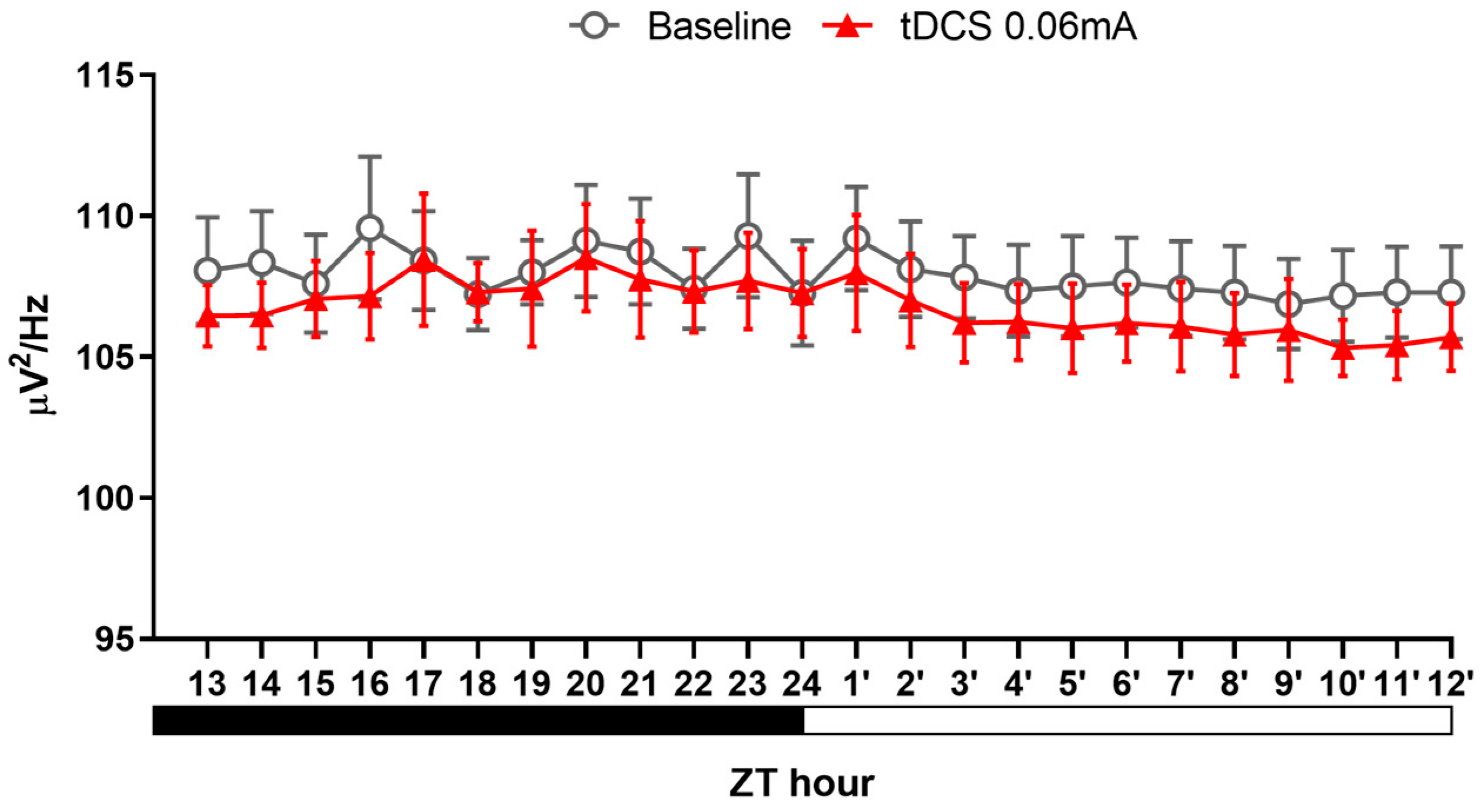
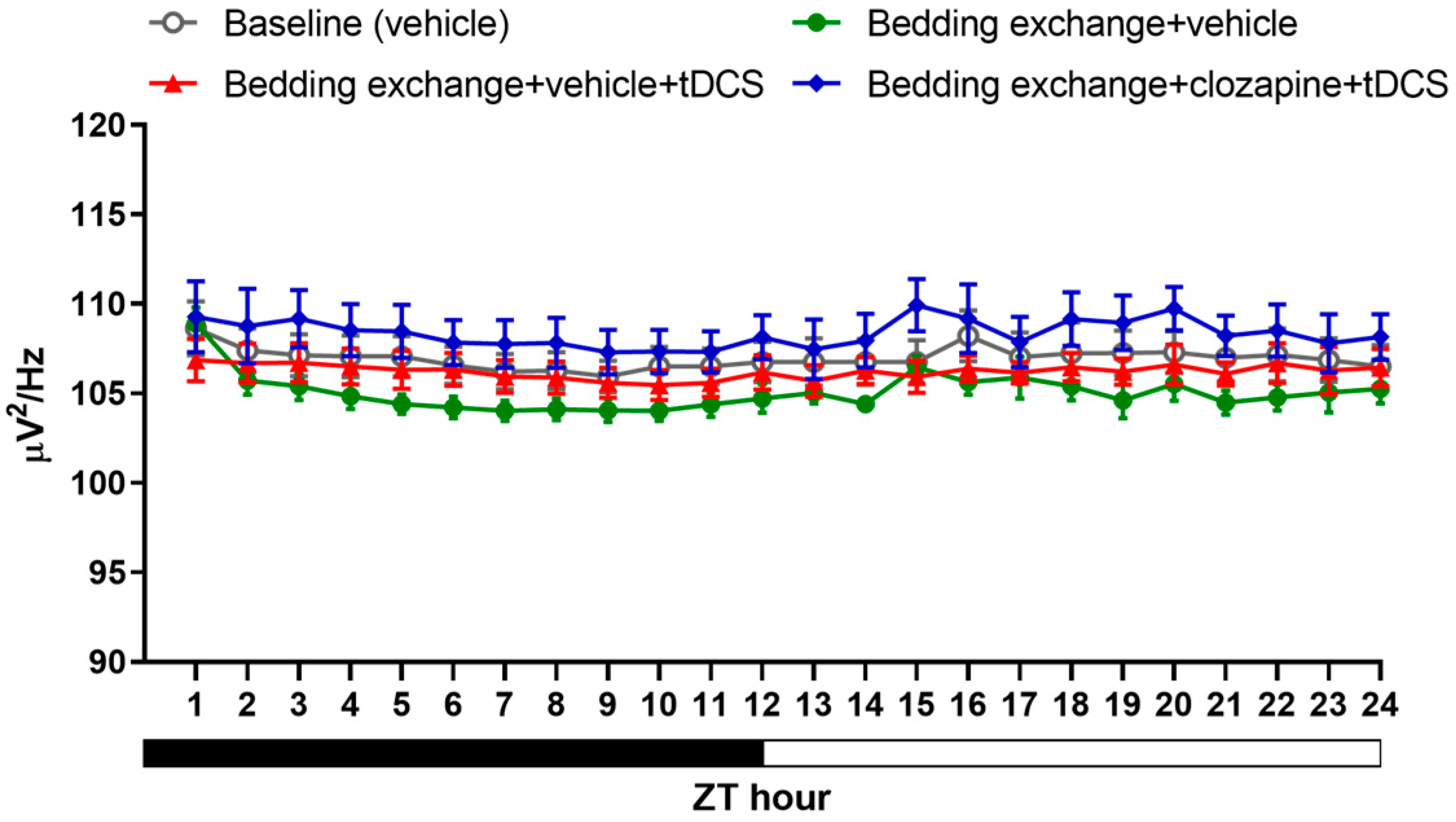
3.2.3. Alteration in NREM Delta Power
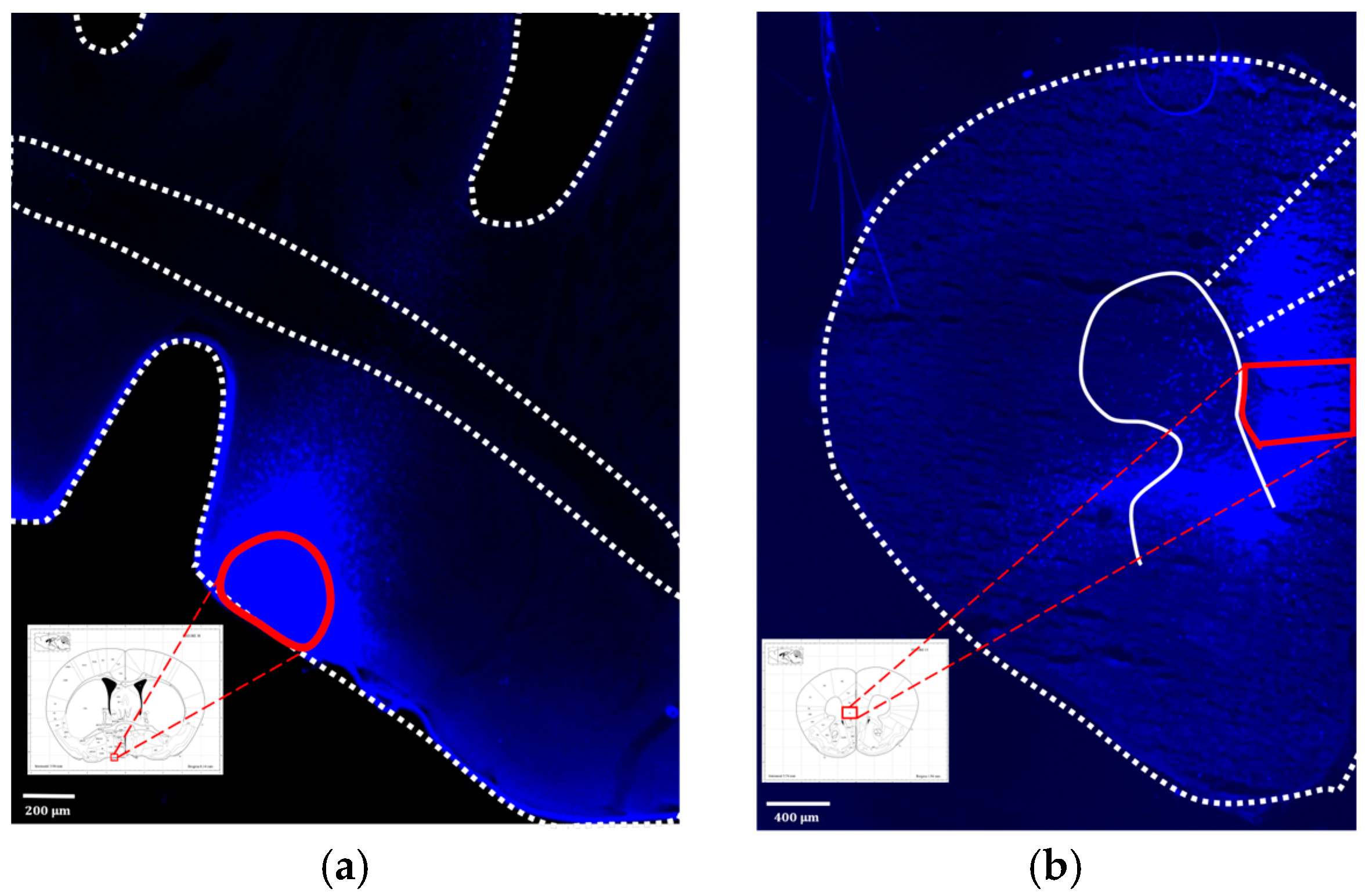
3.3. Confirming the IL to VLPO Pathway in Mice
3.4. The Manipulation of the IL to VLPO Pathway
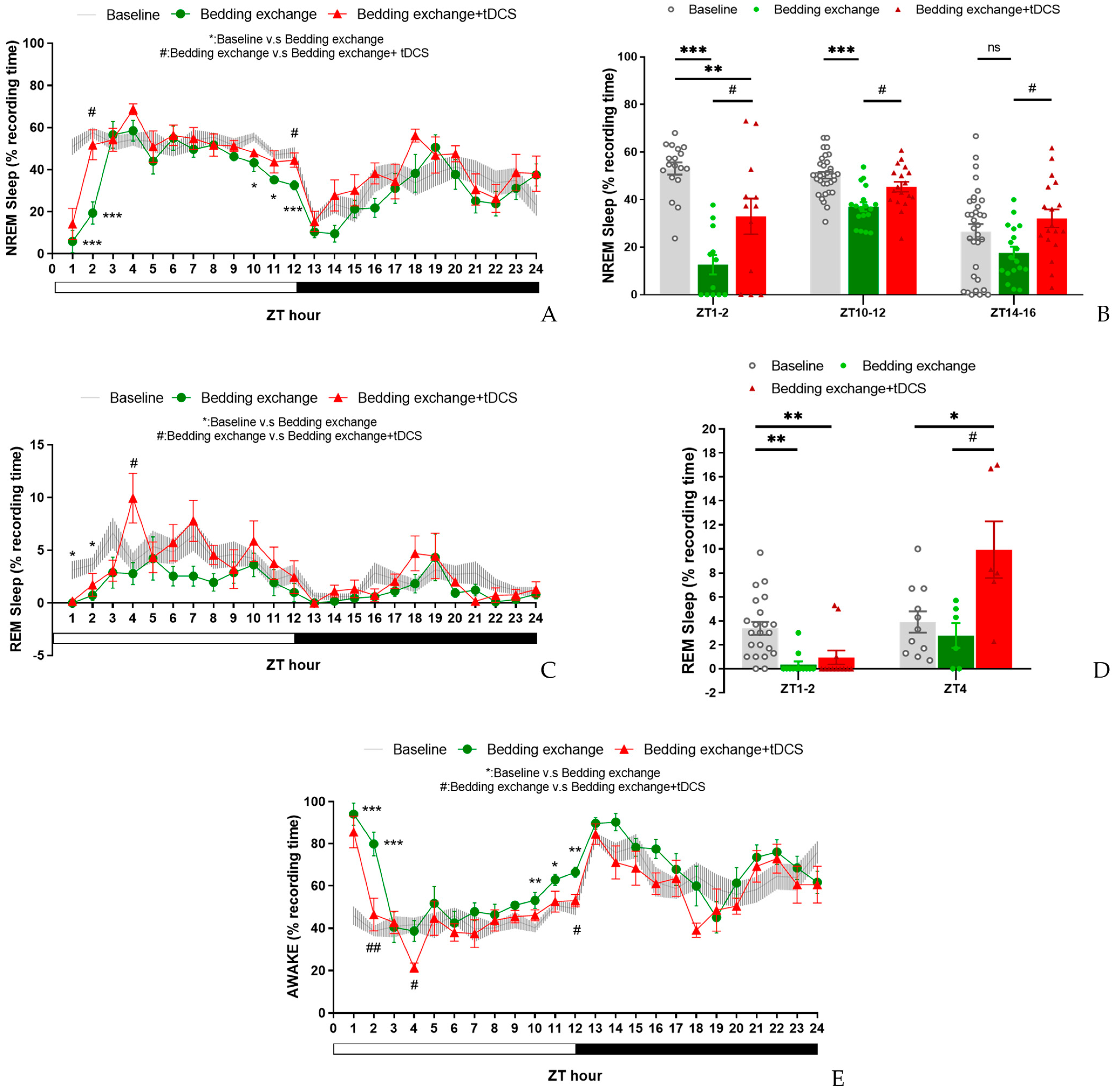
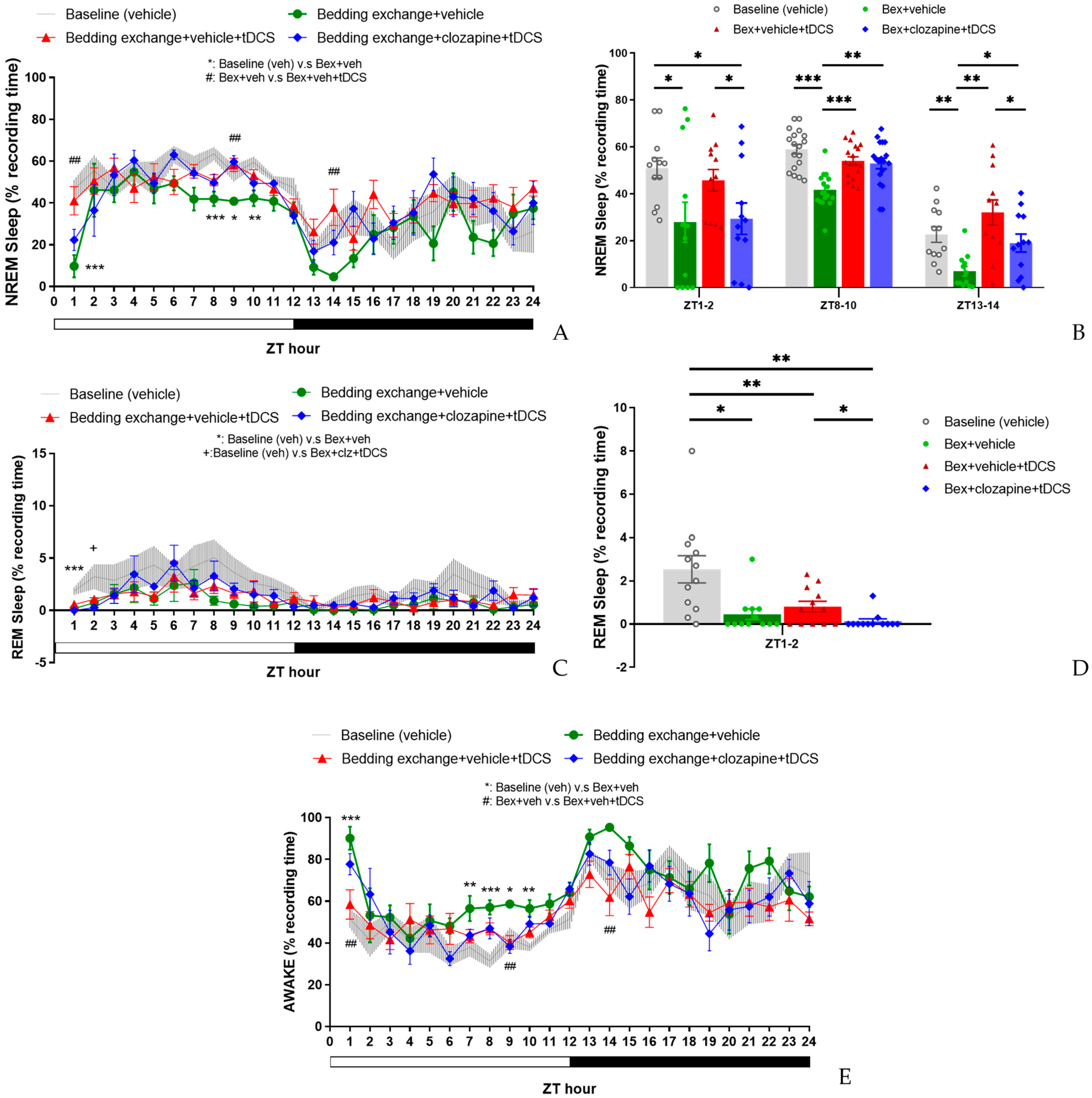
3.4.1. Effect of tDCS after Specifically Blocking the IL-VLPO Pathway
3.4.2. Alterations in Sleep Architecture and Quality
3.4.3. Fluorescence Image of Brain Slices after Microinjection of Virus
4. Discussion
5. Conclusions
6. Limitation
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Levenson, J.C.; Kay, D.B.; Buysse, D.J. The pathophysiology of insomnia. Chest 2015, 147, 1179–1192. [Google Scholar] [CrossRef] [PubMed]
- Bollu, P.C.; Kaur, H. Sleep Medicine: Insomnia and Sleep. Mo. Med. 2019, 116, 68–75. [Google Scholar] [PubMed]
- Roth, T. Insomnia: Definition, prevalence, etiology, and consequences. J. Clin. Sleep Med. 2007, 3 (Suppl. S5), S7–S10. [Google Scholar] [CrossRef] [PubMed]
- Saper, C.B.; Fuller, P.M.; Pedersen, N.P.; Lu, J.; Scammell, T.E. Sleep state switching. Neuron 2010, 68, 1023–1042. [Google Scholar] [CrossRef] [PubMed]
- Sherin, J.; Shiromani, P.; McCarley, R.; Saper, C. Activation of ventrolateral preoptic neurons during sleep. Science 1996, 271, 216–219. [Google Scholar] [CrossRef] [PubMed]
- Gallopin, T.; Luppi, P.H.; Cauli, B.; Urade, Y.; Rossier, J.; Hayaishi, O.; Lambolez, B.; Fort, P. The endogenous somnogen adenosine excites a subset of sleep-promoting neurons via A2A receptors in the ventrolateral preoptic nucleus. Neuroscience 2005, 134, 1377–1390. [Google Scholar] [CrossRef] [PubMed]
- Arrigoni, E.; Fuller, P.M. The Sleep-Promoting Ventrolateral Preoptic Nucleus: What Have We Learned over the Past 25 Years? Int. J. Mol. Sci. 2022, 23, 2905. [Google Scholar] [CrossRef] [PubMed]
- Scammell, T.; Gerashchenko, D.; Urade, Y.; Onoe, H.; Saper, C.; Hayaishi, O. Activation of ventrolateral preoptic neurons by the somnogen prostaglandin D2. Proc. Natl. Acad. Sci. USA 1998, 95, 7754–7759. [Google Scholar] [CrossRef]
- Lu, J.; Greco, M.A.; Shiromani, P.; Saper, C.B. Effect of lesions of the ventrolateral preoptic nucleus on NREM and REM sleep. J. Neurosci. 2000, 20, 3830–3842. [Google Scholar] [CrossRef]
- Sateia, M.J.; Buysse, D.J.; Krystal, A.D.; Neubauer, D.N.; Heald, J.L. Clinical Practice Guideline for the Pharmacologic Treatment of Chronic Insomnia in Adults: An American Academy of Sleep Medicine Clinical Practice Guideline. J. Clin. Sleep Med. 2017, 13, 307–349. [Google Scholar] [CrossRef]
- Doyno, C.R.; White, C.M. Sedative-Hypnotic Agents That Impact Gamma-Aminobutyric Acid Receptors: Focus on Flunitrazepam, Gamma-Hydroxybutyric Acid, Phenibut, and Selank. J. Clin. Pharmacol. 2021, 61 (Suppl. S2), S114–S128. [Google Scholar] [CrossRef] [PubMed]
- Gottesmann, C. GABA mechanisms and sleep. Neuroscience 2002, 111, 231–239. [Google Scholar] [CrossRef] [PubMed]
- Plante, D.T.; Jensen, J.E.; Winkelman, J.W. The role of GABA in primary insomnia. Sleep 2012, 35, 741–742. [Google Scholar] [CrossRef] [PubMed]
- Deoras, K.S.; Moul, D.E. Hypnotics. In Encyclopedia of the Neurological Sciences, 2nd ed.; Aminoff, M.J., Daroff, R.B., Eds.; Academic Press: Oxford, UK, 2014; pp. 646–649. [Google Scholar]
- Vozoris, N.T. Do benzodiazepines contribute to respiratory problems? Expert. Rev. Respir. Med. 2014, 8, 661–663. [Google Scholar] [CrossRef] [PubMed]
- Sarmiento, C.I.; San-Juan, D.; Prasath, V.B.S. Letter to the Editor: Brief history of transcranial direct current stimulation (tDCS): From electric fishes to microcontrollers. Psychol. Med. 2016, 46, 3259–3261. [Google Scholar] [CrossRef] [PubMed]
- Yamada, Y.; Sumiyoshi, T. Neurobiological Mechanisms of Transcranial Direct Current Stimulation for Psychiatric Disorders; Neurophysiological, Chemical, and Anatomical Considerations. Front. Human Neurosci. 2021, 15, 631838. [Google Scholar] [CrossRef] [PubMed]
- Nitsche, M.A.; Cohen, L.G.; Wassermann, E.M.; Priori, A.; Lang, N.; Antal, A.; Paulus, W.; Hummel, F.; Boggio, P.S.; Fregni, F.; et al. Transcranial direct current stimulation: State of the art 2008. Brain Stimul. 2008, 1, 206–223. [Google Scholar] [CrossRef] [PubMed]
- Podda, M.V.; Cocco, S.; Mastrodonato, A.; Fusco, S.; Leone, L.; Barbati, S.A.; Colussi, C.; Ripoli, C.; Grassi, C. Anodal transcranial direct current stimulation boosts synaptic plasticity and memory in mice via epigenetic regulation of Bdnf expression. Sci. Rep. 2016, 6, 22180. [Google Scholar] [CrossRef]
- Kronberg, G.; Bridi, M.; Abel, T.; Bikson, M.; Parra, L.C. Direct Current Stimulation Modulates LTP and LTD: Activity Dependence and Dendritic Effects. Brain Stimul. 2017, 10, 51–58. [Google Scholar] [CrossRef]
- Liu, H.-H.; He, X.-K.; Chen, H.-Y.; Peng, C.-W.; Rotenberg, A.; Juan, C.-H.; Pei, Y.-C.; Liu, H.-L.; Chiang, Y.-H.; Wang, J.-Y.; et al. Neuromodulatory Effects of Transcranial Direct Current Stimulation on Motor Excitability in Rats. Neural Plast. 2019, 2019, 4252943. [Google Scholar] [CrossRef]
- Vergallito, A.; Varoli, E.; Pisoni, A.; Mattavelli, G.; Del Mauro, L.; Feroldi, S.; Vallar, G.; Romero Lauro, L.J. State-dependent effectiveness of cathodal transcranial direct current stimulation on cortical excitability. NeuroImage 2023, 277, 120242. [Google Scholar] [CrossRef] [PubMed]
- Zhao, H.; Qiao, L.; Fan, D.; Zhang, S.; Turel, O.; Li, Y.; Li, J.; Xue, G.; Chen, A.; He, Q. Modulation of Brain Activity with Noninvasive Transcranial Direct Current Stimulation (tDCS): Clinical Applications and Safety Concerns. Front. Psychol. 2017, 8, 685. [Google Scholar] [CrossRef] [PubMed]
- Pochon, J.B.; Levy, R.; Poline, J.B.; Crozier, S.; Lehéricy, S.; Pillon, B.; Deweer, B.; Le Bihan, D.; Dubois, B. The role of dorsolateral prefrontal cortex in the preparation of forthcoming actions: An fMRI study. Cereb. Cortex 2001, 11, 260–266. [Google Scholar] [CrossRef] [PubMed]
- Barbey, A.K.; Koenigs, M.; Grafman, J. Dorsolateral prefrontal contributions to human working memory. Cortex 2013, 49, 1195–1205. [Google Scholar] [CrossRef] [PubMed]
- Xia, X.; Li, Y.; Wang, Y.; Xia, J.; Lin, Y.; Zhang, X.; Liu, Y.; Zhang, J. Functional role of dorsolateral prefrontal cortex in the modulation of cognitive bias. Psychophysiology 2021, 58, e13894. [Google Scholar] [CrossRef] [PubMed]
- Minichino, A.; Bersani, F.S.; Spagnoli, F.; Corrado, A.; De Michele, F.; Calò, W.K.; Primavera, M.; Yang, B.; Bernabei, L.; Macrì, F.; et al. Prefronto-Cerebellar Transcranial Direct Current Stimulation Improves Sleep Quality in Euthymic Bipolar Patients: A Brief Report. Behav. Neurol. 2014, 2014, 876521. [Google Scholar] [CrossRef] [PubMed]
- Kim, Y.J.; Ku, J.; Kim, H.J.; Im, D.J.; Lee, H.S.; Han, K.A.; Kang, Y.J. Randomized, sham controlled trial of transcranial direct current stimulation for painful diabetic polyneuropathy. Ann. Rehabil. Med. 2013, 37, 766–776. [Google Scholar] [CrossRef]
- Herrero Babiloni, A.; Bellemare, A.; Beetz, G.; Vinet, S.A.; Martel, M.O.; Lavigne, G.J.; De Beaumont, L. The effects of non-invasive brain stimulation on sleep disturbances among different neurological and neuropsychiatric conditions: A systematic review. Sleep. Med. Rev. 2021, 55, 101381. [Google Scholar] [CrossRef]
- Frase, L.; Selhausen, P.; Krone, L.; Tsodor, S.; Jahn, F.; Feige, B.; Maier, J.G.; Mainberger, F.; Piosczyk, H.; Kuhn, M.; et al. Differential effects of bifrontal tDCS on arousal and sleep duration in insomnia patients and healthy controls. Brain Stimul. 2019, 12, 674–683. [Google Scholar] [CrossRef]
- Jung, K.; Jun, J. Efficacy of transcranial direct-current stimulation on chronic insomnia. Brain Stimul. Basic Transl. Clin. Res. Neuromodulation 2019, 12, 557. [Google Scholar] [CrossRef]
- Esmaeilpour, Z.; Marangolo, P.; Hampstead, B.M.; Bestmann, S.; Galletta, E.; Knotkova, H.; Bikson, M. Incomplete evidence that increasing current intensity of tDCS boosts outcomes. Brain Stimul. 2018, 11, 310–321. [Google Scholar] [CrossRef] [PubMed]
- Monte-Silva, K.; Kuo, M.-F.; Hessenthaler, S.; Fresnoza, S.; Liebetanz, D.; Paulus, W.; Nitsche, M.A. Induction of Late LTP-Like Plasticity in the Human Motor Cortex by Repeated Non-Invasive Brain Stimulation. Brain Stimul. 2013, 6, 424–432. [Google Scholar] [CrossRef]
- Jackson, M.P.; Rahman, A.; Lafon, B.; Kronberg, G.; Ling, D.; Parra, L.C.; Bikson, M. Animal models of transcranial direct current stimulation: Methods and mechanisms. Clin. Neurophysiol. 2016, 127, 3425–3454. [Google Scholar] [CrossRef] [PubMed]
- Thair, H.; Holloway, A.L.; Newport, R.; Smith, A.D. Transcranial Direct Current Stimulation (tDCS): A Beginner’s Guide for Design and Implementation. Front. Neurosci. 2017, 11, 641. [Google Scholar] [CrossRef] [PubMed]
- De Souza Nicolau, E.; de Alvarenga, K.A.F.; Tenza-Ferrer, H.; Nogueira, M.C.A.; Rezende, F.D.; Nicolau, N.F.; Collodetti, M.; de Miranda, D.M.; Magno, L.A.V.; Romano-Silva, M.A. Transcranial direct current stimulation (tDCS) in mice. JoVE J. Vis. Exp. 2018, 139, e58517. [Google Scholar]
- Zhou, Q.; Yu, C.; Yu, H.; Zhang, Y.; Liu, Z.; Hu, Z.; Yuan, T.-F.; Zhou, D. The effects of repeated transcranial direct current stimulation on sleep quality and depression symptoms in patients with major depression and insomnia. Sleep Med. 2020, 70, 17–26. [Google Scholar] [CrossRef]
- Mohebbian, B.; Najafi, M.; Sabahi, P. The effect of transcranial direct current stimulation on sleep quality, resilience, and optimism. Curr. Psychol. 2023, 42, 5785–5792. [Google Scholar] [CrossRef]
- Liebetanz, D.; Koch, R.; Mayenfels, S.; König, F.; Paulus, W.; Nitsche, M.A. Safety limits of cathodal transcranial direct current stimulation in rats. Clin. Neurophysiol. 2009, 120, 1161–1167. [Google Scholar] [CrossRef]
- Schwierin, B.; Borbély, A.A.; Tobler, I. Effects of N6-cyclopentyladenosine and caffeine on sleep regulation in the rat. Eur. J. Pharmacol. 1996, 300, 163–171. [Google Scholar] [CrossRef]
- Yanik, G.; Glaum, S.; Radulovacki, M. The dose-response effects of caffeine on sleep in rats. Brain Res. 1987, 403, 177–180. [Google Scholar] [CrossRef]
- Revel, F.G.; Gottowik, J.; Gatti, S.; Wettstein, J.G.; Moreau, J.L. Rodent models of insomnia: A review of experimental procedures that induce sleep disturbances. Neurosci. Biobehav. Rev. 2009, 33, 874–899. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.; Lee, H.J.; Cho, C.H. Mediation Effect of Insomnia Symptoms on Relation Between Stress and Quality of Life. Psychiatry Investig. 2022, 19, 229–238. [Google Scholar] [CrossRef] [PubMed]
- Herbst, E.; Metzler, T.J.; Lenoci, M.; McCaslin, S.E.; Inslicht, S.; Marmar, C.R.; Neylan, T.C. Adaptation effects to sleep studies in participants with and without chronic posttraumatic stress disorder. Psychophysiology 2010, 47, 1127–1133. [Google Scholar] [CrossRef] [PubMed]
- Toth, L.A.; Bhargava, P. Animal models of sleep disorders. Comp. Med. 2013, 63, 91–104. [Google Scholar] [PubMed]
- Jou, S.B.; Tsai, C.J.; Fang, C.Y.; Yi, P.L.; Chang, F.C. Effects of N(6) -(4-hydroxybenzyl) adenine riboside in stress-induced insomnia in rodents. J. Sleep Res. 2021, 30, e13156. [Google Scholar] [CrossRef] [PubMed]
- Brown, V.J.; Bowman, E.M. Rodent models of prefrontal cortical function. Trends Neurosci. 2002, 25, 340–343. [Google Scholar] [CrossRef]
- Carlén, M. What constitutes the prefrontal cortex? Science 2017, 358, 478–482. [Google Scholar] [CrossRef] [PubMed]
- Laubach, M.; Amarante, L.M.; Swanson, K.; White, S.R. What, If Anything, Is Rodent Prefrontal Cortex? eNeuro 2018, 5, ENEURO.0315-18.2018. [Google Scholar] [CrossRef]
- Chou, T.C.; Bjorkum, A.A.; Gaus, S.E.; Lu, J.; Scammell, T.E.; Saper, C.B. Afferents to the ventrolateral preoptic nucleus. J. Neurosci. 2002, 22, 977–990. [Google Scholar] [CrossRef]
- Naumann, T.; Härtig, W.; Frotscher, M. Retrograde tracing with Fluoro-Gold: Different methods of tracer detection at the ultrastructural level and neurodegenerative changes of back-filled neurons in long-term studies. J. Neurosci. Methods 2000, 103, 11–21. [Google Scholar] [CrossRef]
- Yao, F.; Zhang, E.; Gao, Z.; Ji, H.; Marmouri, M.; Xia, X. Did you choose appropriate tracer for retrograde tracing of retinal ganglion cells? The differences between cholera toxin subunit B and Fluorogold. PLoS ONE 2018, 13, e0205133. [Google Scholar] [CrossRef] [PubMed]
- Roth, B.L. DREADDs for neuroscientists. Neuron 2016, 89, 683–694. [Google Scholar] [CrossRef] [PubMed]
- Thompson, K.J.; Khajehali, E.; Bradley, S.J.; Navarrete, J.S.; Huang, X.P.; Slocum, S.; Jin, J.; Liu, J.; Xiong, Y.; Olsen, R.H.J.; et al. DREADD Agonist 21 Is an Effective Agonist for Muscarinic-Based DREADDs in Vitro and in Vivo. ACS Pharmacol. Transl. Sci. 2018, 1, 61–72. [Google Scholar] [CrossRef] [PubMed]
- Jendryka, M.; Palchaudhuri, M.; Ursu, D.; van der Veen, B.; Liss, B.; Kätzel, D.; Nissen, W.; Pekcec, A. Pharmacokinetic and pharmacodynamic actions of clozapine-N-oxide, clozapine, and compound 21 in DREADD-based chemogenetics in mice. Sci. Rep. 2019, 9, 4522. [Google Scholar] [CrossRef]
- Ruiz-Velasco, V.; Ikeda, S.R. Multiple G-protein betagamma combinations produce voltage-dependent inhibition of N-type calcium channels in rat superior cervical ganglion neurons. J. Neurosci. 2000, 20, 2183–2191. [Google Scholar] [CrossRef] [PubMed]
- Sadana, R.; Dessauer, C.W. Physiological roles for G protein-regulated adenylyl cyclase isoforms: Insights from knockout and overexpression studies. Neurosignals 2009, 17, 5–22. [Google Scholar] [CrossRef] [PubMed]
- Nagy, A. Cre recombinase: The universal reagent for genome tailoring. Genesis 2000, 26, 99–109. [Google Scholar] [CrossRef]
- Yamabe, M.; Horie, K.; Shiokawa, H.; Funato, H.; Yanagisawa, M.; Kitagawa, H. MC-SleepNet: Large-scale Sleep Stage Scoring in Mice by Deep Neural Networks. Sci. Rep. 2019, 9, 15793. [Google Scholar] [CrossRef]
- Long, S.; Ding, R.; Wang, J.; Yu, Y.; Lu, J.; Yao, D. Sleep Quality and Electroencephalogram Delta Power. Front. Neurosci. 2021, 15, 803507. [Google Scholar] [CrossRef]
- Pikhovych, A.; Walter, H.L.; Mahabir, E.; Fink, G.R.; Graf, R.; Schroeter, M.; Rueger, M.A. Transcranial direct current stimulation in the male mouse to promote recovery after stroke. Lab. Anim. 2016, 50, 212–216. [Google Scholar] [CrossRef]
- Sánchez-León, C.A.; Cordones, I.; Ammann, C.; Ausín, J.M.; Gómez-Climent, M.A.; Carretero-Guillén, A.; Sánchez-Garrido Campos, G.; Gruart, A.; Delgado-García, J.M.; Cheron, G.; et al. Immediate and after effects of transcranial direct-current stimulation in the mouse primary somatosensory cortex. Sci. Rep. 2021, 11, 3123. [Google Scholar] [CrossRef] [PubMed]
- Huang, Y.-Z.; Lu, M.-K.; Antal, A.; Classen, J.; Nitsche, M.; Ziemann, U.; Ridding, M.; Hamada, M.; Ugawa, Y.; Jaberzadeh, S.; et al. Plasticity induced by non-invasive transcranial brain stimulation: A position paper. Clin. Neurophysiol. 2017, 128, 2318–2329. [Google Scholar] [CrossRef] [PubMed]
- Quirk, G.J.; Garcia, R.; González-Lima, F. Prefrontal mechanisms in extinction of conditioned fear. Biol. Psychiatry 2006, 60, 337–343. [Google Scholar] [CrossRef]
- Peters, J.; LaLumiere, R.T.; Kalivas, P.W. Infralimbic prefrontal cortex is responsible for inhibiting cocaine seeking in extinguished rats. J. Neurosci. 2008, 28, 6046–6053. [Google Scholar] [CrossRef] [PubMed]
- Smith, K.S.; Graybiel, A.M. Habit formation. Dialogues Clin. Neurosci. 2016, 18, 33–43. [Google Scholar] [CrossRef]














{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
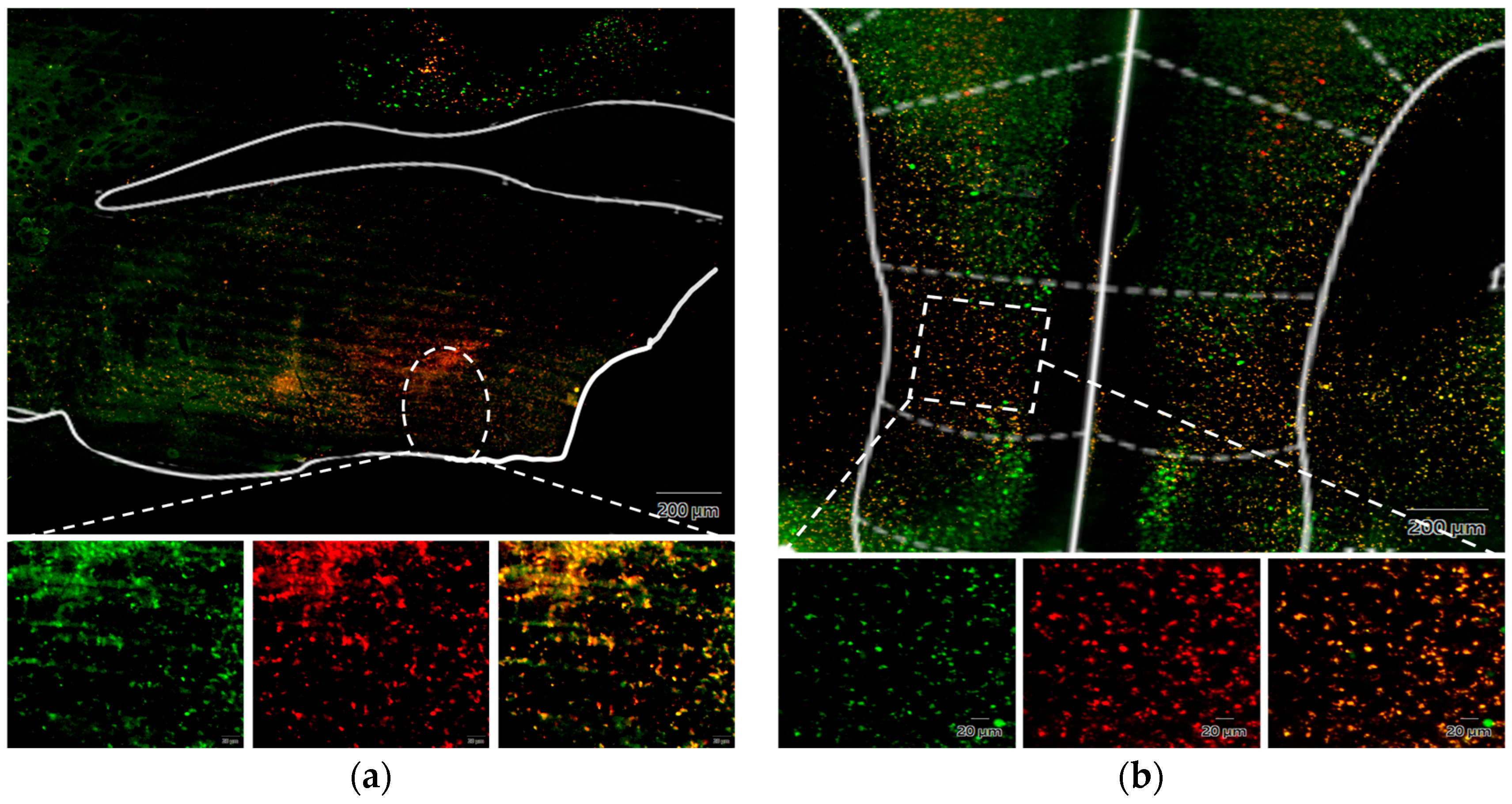{kind=link}
| Group | ZT Hour | L:D Cycle | Bout Number | Bout Duration | ||||
|---|---|---|---|---|---|---|---|---|
| NREM | WAKE | REM | NREM | WAKE | REM | |||
| Control | 13–24 | D | 9.5 ± 2.0 | 6.7 ± 1.5 | 1.1 ± 0.2 | 2.1 ± 0.4 | 10.2 ± 3.7 | 0.5 ± 0.1 |
| tDCS | 13–24 | D | 8.4 ± 1.4 | 5.9 ± 0.7 | 1.1 ± 0.1 | 3.0 ± 0.2 * | 8.0 ± 2.2 | 0.6 ± 0.1 |
| Control | 1′–12′ | L | 14.3 ± 2.0 | 9.7 ± 1.3 | 1.9 ± 0.5 | 2.4 ± 0.4 | 2.9 ± 0.5 | 1.0 ± 0.2 |
| tDCS | 1′–12′ | L | 12.3 ± 1.3 | 8.2 ± 0.9 | 2.1 ± 0.2 | 3.2 ± 0.4 | 2.7 ± 0.3 | 1.2 ± 0.2 |
| Control | 13–15 | D | 8.3 ± 1.7 | 5.9 ± 1.3 | 0.3 ± 0.1 | 1.6 ± 0.1 | 12.0 ± 3.8 | 0.3 ± 0.1 |
| tDCS | 13–15 | D | 9.6 ± 1.6 | 6.1 ± 0.8 | 1.3 ± 0.2 * | 3.8 ± 0.7 * | 4.5 ± 0.6 | 0.7 ± 0.1 |
| Control | 4′–12′ | L | 15.3 ± 2.0 | 10.4 ± 1.4 | 1.8 ± 0.6 | 2.1 ± 0.3 | 2.6 ± 0.4 | 1.0 ± 0.2 |
| tDCS | 4′–12′ | L | 13.2 ± 1.2 | 8.5 ± 0.9 | 2.2 ± 0.1 | 3.0 ± 0.3 * | 2.3 ± 0.2 | 1.3 ± 0.2 |
| Group | ZT Hour | L:D Cycle | Bout Number | Bout Duration | ||||
|---|---|---|---|---|---|---|---|---|
| NREM | WAKE | REM | NREM | WAKE | REM | |||
| Control | 1–12 | L | 13.8 ± 1.4 | 9.5 ± 0.9 | 1.9 ± 0.2 | 2.6 ± 0.2 | 3.1 ± 0.4 | 1.0 ± 0.1 |
| Bex | 1–12 | L | 11.2 ± 0.9 | 7.9 ± 0.5 | 1.2 ± 0.3 | 2.2 ± 0.2 | 8.7 ± 1.3 ** | 0.6 ± 0.2 |
| Bex + tDCS | 1–12 | L | 13.4 ± 1.3 | 8.5 ± 0.9 | 1.9 ± 0.4 | 2.3 ± 0.2 | 5.6 ± 1.4 | 1.0 ± 0.2 |
| Control | 13–24 | D | 8.6 ± 1.2 | 6.3 ± 0.9 | 1.0 ± 0.1 | 2.2 ± 0.2 | 12.2 ± 3.1 | 0.6 ± 0.1 |
| Bex | 13–24 | D | 7.8 ± 1.0 | 5.5 ± 0.8 | 0.6 ± 0.1 * | 2.2 ± 0.3 | 12.3 ± 2.1 | 0.4 ± 0.1 |
| Bex + tDCS | 13–24 | D | 9.4 ± 1.3 | 6.6 ± 0.9 | 0.9 ± 0.1 | 2.3 ± 0.2 | 10.1 ± 3.1 | 0.6 ± 0.1 |
| Control | 1–2 | L | 12.9 ± 1.9 | 7.9 ± 0.9 | 2.0 ± 0.2 | 2.9 ± 0.3 | 3.3 ± 0.5 | 1.0 ± 0.1 |
| Bex | 1–2 | L | 2.2 ± 0.7 * | 1.8 ± 0.5 ** | 0.3 ± 0.2 *** | 1.9 ± 0.6 | 34.5 ± 7.0 *** | 0.1 ± 0.1 ** |
| Bex + tDCS | 1–2 | L | 7.5 ± 1.8 | 5.6 ± 1.6 | 0.4 ± 0.3 *** | 2.0 ± 0.2 | 19.2 ± 7.0 | 0.3 ± 0.2 ** |
| Control | 4 | L | 14.7 ± 1.4 | 11.2 ± 1.0 | 2.2 ± 0.4 | 2.3 ± 0.3 | 2.2 ± 0.3 | 0.9 ± 0.1 |
| Bex | 4 | L | 11.3 ± 1.5 | 7.8 ± 0.9 | 1.8 ± 0.7 | 3.2 ± 0.4 | 2.7 ± 0.6 | 0.6 ± 0.2 |
| Bex + tDCS | 4 | L | 14.8 ± 1.9 | 7.7 ± 0.7 | 4.5 ± 0.9 + | 2.9 ± 0.4 | 1.1 ± 0.2 + | 1.4 ± 0.2 + |
| Control | 10–12 | L | 15.0 ± 1.3 | 10.8 ± 0.8 | 1.3 ± 0.3 | 2.1 ± 0.2 | 2.6 ± 0.3 | 0.7 ± 0.2 |
| Bex | 10–12 | L | 13.2 ± 0.9 | 9.7 ± 0.6 | 1.1 ± 0.4 | 1.6 ± 0.1 | 3.9 ± 0.3 * | 0.6 ± 0.2 |
| Bex + tDCS | 10–12 | L | 15.9 ± 1.7 | 10.5 ± 1.0 | 1.6 ± 0.5 | 1.8 ± 0.1 | 2.9 ± 0.4 | 1.0 ± 0.3 |
| Control | 14–16 | D | 7.6 ± 1.4 | 5.3 ± 0.9 | 0.8 ± 0.2 | 1.8 ± 0.2 | 15.3 ± 4.7 | 0.5 ± 0.1 |
| Bex | 14–16 | D | 4.7 ± 1.2 | 3.2 ± 0.8 | 0.4 ± 0.1 | 2.4 ± 0.4 | 18.2 ± 2.9 | 0.2 ± 0.1 |
| Bex + tDCS | 14–16 | D | 8.5 ± 1.8 | 5.6 ± 1.0 | 0.6 ± 0.3 | 2.5 ± 0.2 | 9.3 ± 2.5 | 0.5 ± 0.3 |
| Group | ZT Hour | L:D Cycle | Bout Number | Bout Duration | ||||
|---|---|---|---|---|---|---|---|---|
| NREM | WAKE | REM | NREM | WAKE | REM | |||
| Control | 1–12 | L | 11.8 ± 0.6 | 7.6 ± 0.3 | 1.9 ± 0.6 | 2.9 ± 0.2 | 3.4 ± 0.3 ** | 0.7 ± 0.1 |
| Bex + veh | 1–12 | L | 11.5 ± 0.4 | 8.5 ± 0.3 | 0.9 ± 0.4 | 2.3 ± 0.2 | 8.3 ± 1.2 | 0.4 ± 0.2 |
| Bex + veh + tDCS | 1–12 | L | 14.3 ± 1.3 | 10.3 ± 1.1 | 1.3 ± 0.3 | 2.2 ± 0.2 | 3.2 ± 0.4 ** | 0.4 ± 0.1 |
| Bex + clz + tDCS | 1–12 | L | 12.0 ± 0.8 | 8.4 ± 0.7 | 1.1 ± 0.2 | 2.6 ± 0.3 | 4.8 ± 1.0 * | 0.6 ± 0.1 |
| Control | 13–24 | D | 6.8 ± 0.6 | 5.2 ± 0.4 | 1.0 ± 0.2 | 2.6 ± 0.2 | 11.2 ± 1.7 | 0.4 ± 0.1 |
| Bex + veh | 13–24 | D | 5.7 ± 1.1 | 4.4 ± 0.7 | 0.3 ± 0.1 | 2.4 ± 0.3 | 16.5 ± 2.9 | 0.1 ± 0.1 |
| Bex + veh + tDCS | 13–24 | D | 10.4 ± 0.6 * | 7.6 ± 0.6 * | 0.7 ± 0.2 | 2.1 ± 0.2 | 5.6 ± 0.6 * | 0.3 ± 0.1 |
| Bex + clz + tDCS | 13–24 | D | 7.2 ± 1.4 | 5.5 ± 0.7 | 0.7 ± 0.2 | 2.7 ± 0.2 | 10.9 ± 2.2 | 0.3 ± 0.1 |
| Control | 1–2 | L | 9.3 ± 1.1 * | 5.9 ± 0.7 | 1.9 ± 0.5 ** | 3.4 ± 0.3 | 4.6 ± 0.7 ** | 0.7 ± 0.1 |
| Bex + veh | 1–2 | L | 3.5 ± 1.0 | 2.7 ± 0.7 | 0.3 ± 0.2 | 3.0 ± 0.9 | 30.5 ± 8.1 | 0.2 ± 0.1 |
| Bex + veh + tDCS | 1–2 | L | 12.0 ± 1.7 ** | 9.2 ± 1.1 *** | 0.8 ± 0.3 | 2.2 ± 0.2 | 4.2 ± 0.8 ** | 0.4 ± 0.1 |
| Bex + clz + tDCS | 1–2 | L | 5.8 ± 1.0 # | 5.4 ± 0.9 # | 0.2 ± 0.2 ++ | 2.9 ± 0.8 | 13.1 ± 5.7 | 0.0 ± 0.0 ++ |
| Control | 8–10 | L | 13.6 ± 1.1 | 8.5 ± 0.5 ## | 2.3 ± 0.7 | 2.8 ± 0.3 ** | 2.4 ± 0.2 | 0.7 ± 0.2 |
| Bex + veh | 8–10 | L | 15.2 ± 1.0 | 11.5 ± 0.3 | 0.7 ± 0.2 | 1.6 ± 0.2 | 3.1 ± 0.2 | 0.4 ± 0.1 |
| Bex + veh + tDCS | 8–10 | L | 17.7 ± 1.4 | 13.2 ± 1.3 | 1.4 ± 0.2 | 1.9 ± 0.2 + | 2.0 ± 0.2 ** | 0.6 ± 0.2 |
| Bex + clz + tDCS | 8–10 | L | 14.9 ± 0.7 | 9.6 ± 0.3 ## | 1.2 ± 0.4 | 2.2 ± 0.1 | 2.6 ± 0.2 | 0.8 ± 0.3 |
| Control | 13–14 | D | 6.6 ± 0.9 | 4.4 ± 0.9 | 0.3 ± 0.2 | 2.0 ± 0.3 | 12.4 ± 2.3 | 0.2 ± 0.1 |
| Bex + veh | 13–14 | D | 1.8 ± 0.4 | 1.8 ± 0.3 | 0.0 ± 0.0 | 1.8 ± 0.5 | 28.6 ± 5.1 | 0.0 ± 0.0 |
| Bex + veh + tDCS | 13–14 | D | 8.9 ± 1.6 ** | 6.3 ± 0.8 ** | 0.5 ± 0.3 | 1.9 ± 0.3 | 7.7 ± 2.6 * | 0.2 ± 0.1 |
| Bex + clz + tDCS | 13–14 | D | 6.0 ± 1.7 | 4.4 ± 1.0 | 0.4 ± 0.2 | 1.7 ± 0.4 | 16.2 ± 5.3 | 0.3 ± 0.1 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Su, Y.-J.; Yi, P.-L.; Chang, F.-C. Transcranial Direct Current Stimulation (tDCS) Ameliorates Stress-Induced Sleep Disruption via Activating Infralimbic-Ventrolateral Preoptic Projections. Brain Sci. 2024, 14, 105. https://doi.org/10.3390/brainsci14010105
Su Y-J, Yi P-L, Chang F-C. Transcranial Direct Current Stimulation (tDCS) Ameliorates Stress-Induced Sleep Disruption via Activating Infralimbic-Ventrolateral Preoptic Projections. Brain Sciences. 2024; 14(1):105. https://doi.org/10.3390/brainsci14010105
Chicago/Turabian StyleSu, Yu-Jie, Pei-Lu Yi, and Fang-Chia Chang. 2024. "Transcranial Direct Current Stimulation (tDCS) Ameliorates Stress-Induced Sleep Disruption via Activating Infralimbic-Ventrolateral Preoptic Projections" Brain Sciences 14, no. 1: 105. https://doi.org/10.3390/brainsci14010105
APA StyleSu, Y.-J., Yi, P.-L., & Chang, F.-C. (2024). Transcranial Direct Current Stimulation (tDCS) Ameliorates Stress-Induced Sleep Disruption via Activating Infralimbic-Ventrolateral Preoptic Projections. Brain Sciences, 14(1), 105. https://doi.org/10.3390/brainsci14010105