
A Four-Week High-Fat Diet Induces Anxiolytic-like Behaviors through Mature BDNF in the mPFC of Mice

Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals
2.2. Behavioral Tests
2.2.1. Open Field Test (OFT)
2.2.2. Light-Dark Box Test (LDT)
2.2.3. Elevated Plus Maze Test (EPM)
2.3. Immunofluorescence
2.4. Western Blotting
2.5. Real-Time Quantitative PCR
2.6. Stereotaxic Injection of Virus
2.7. Cannula Implantation and BDNF-Neutralizing Antibody Administration
2.8. Statistical Analysis
3. Results
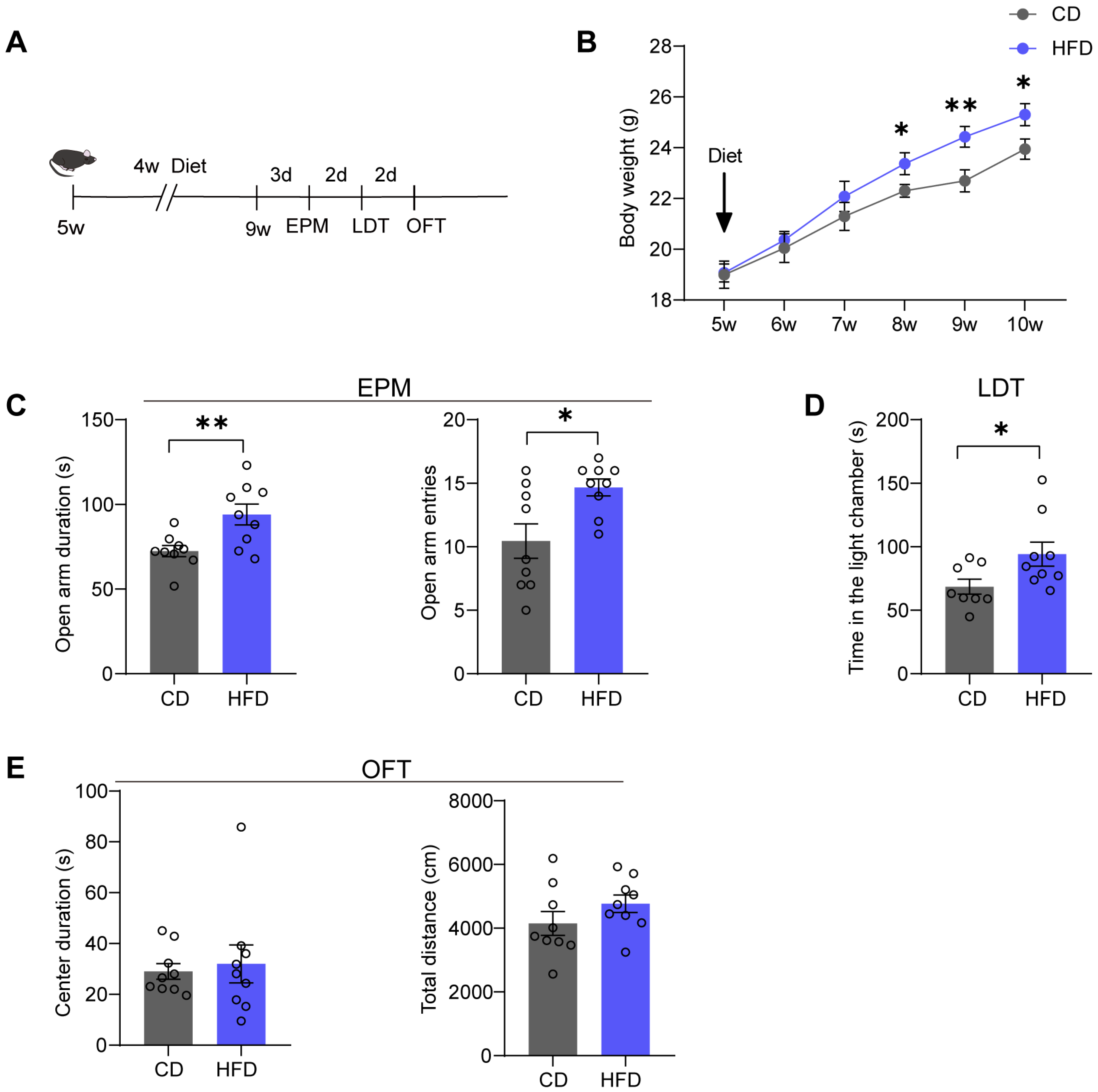
3.1. Mice Fed with an HFD for Four Weeks Exhibit Anxiolytic-like Behaviors
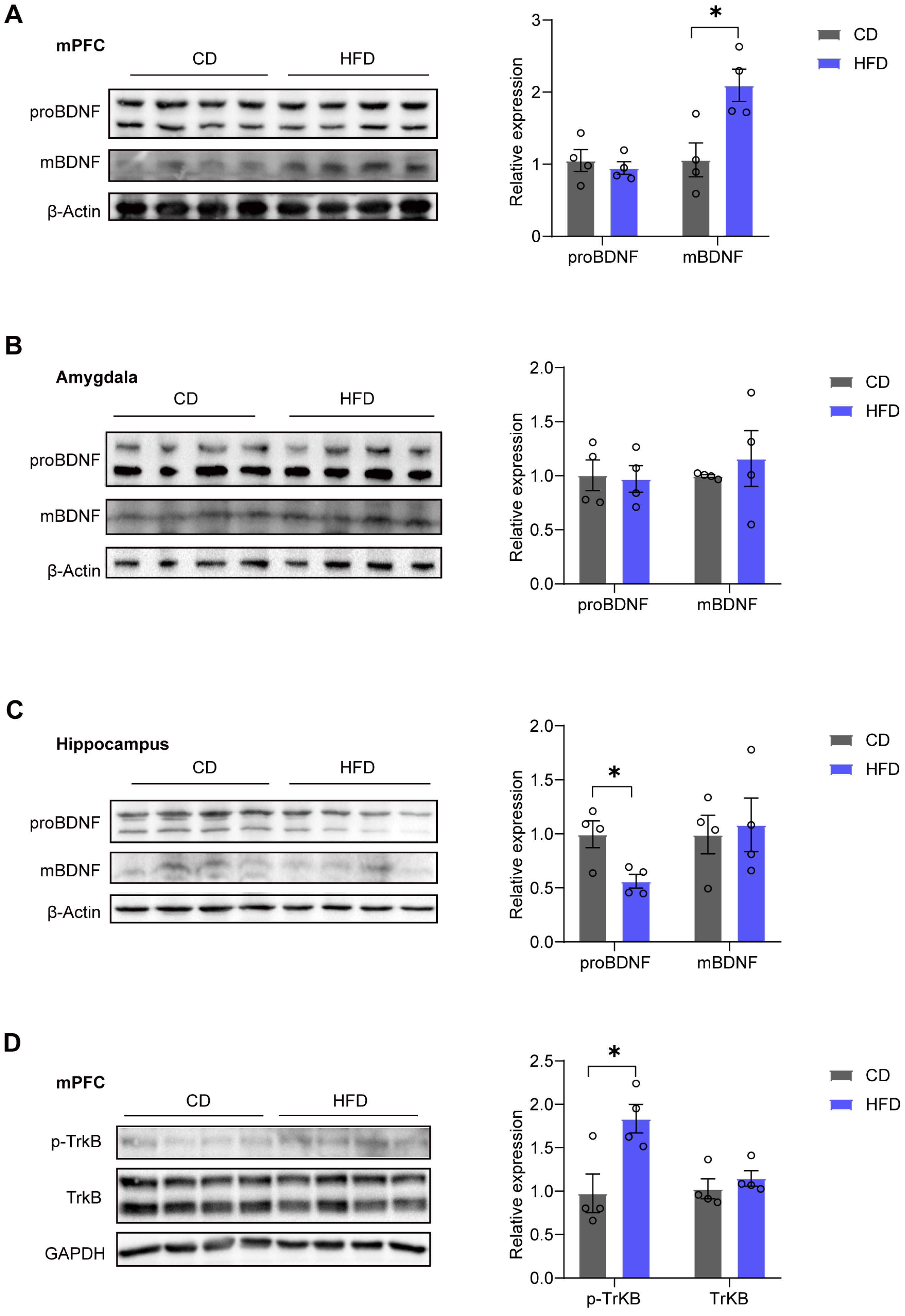
3.2. mBDNF and p-TrkB Levels in the mPFC Are Increased in HFD Mice
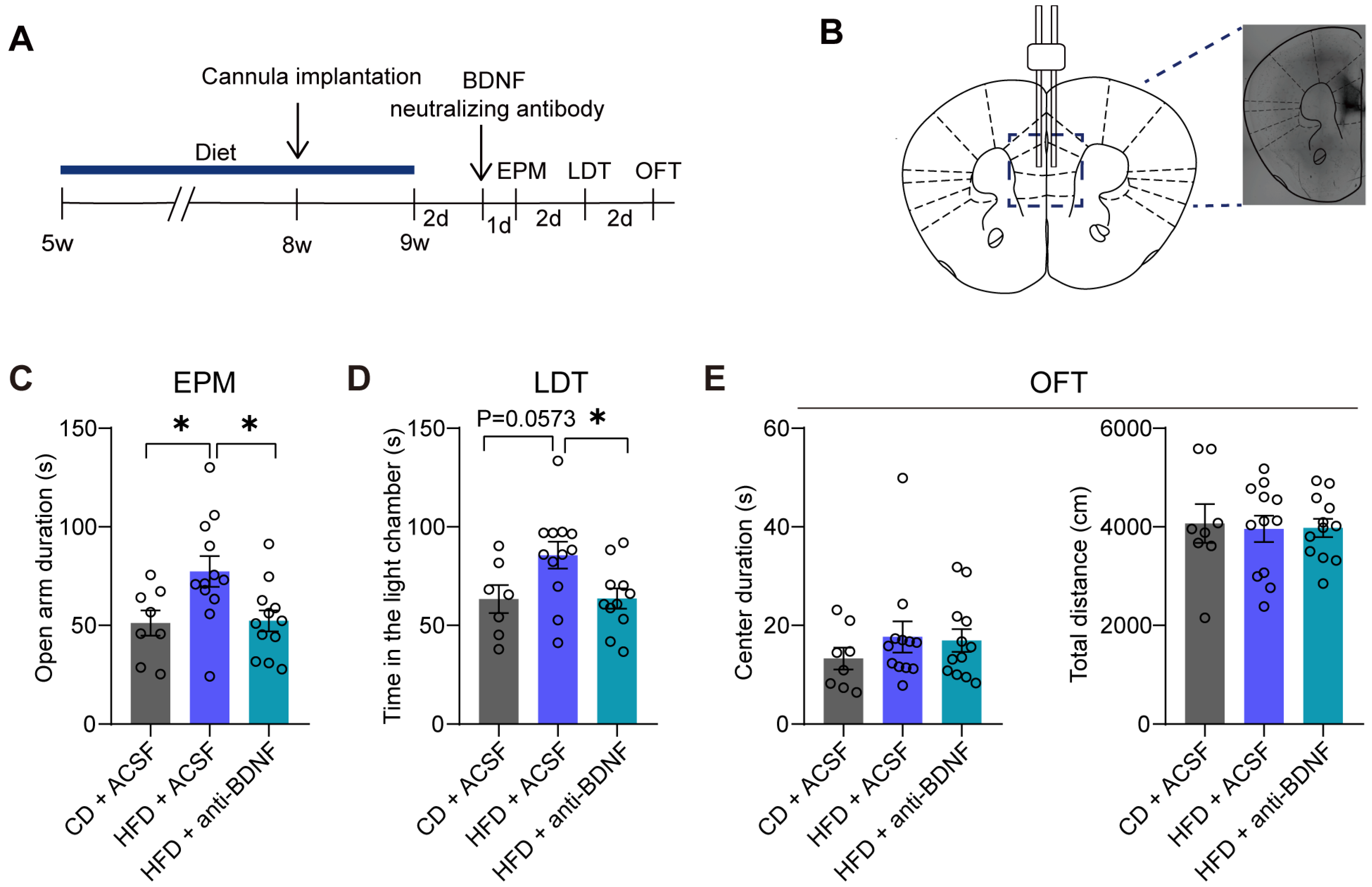
3.3. The Administration of a BDNF-Neutralizing Antibody into the mPFC Reverses Anxiolytic-like Behaviors Induced by the HFD
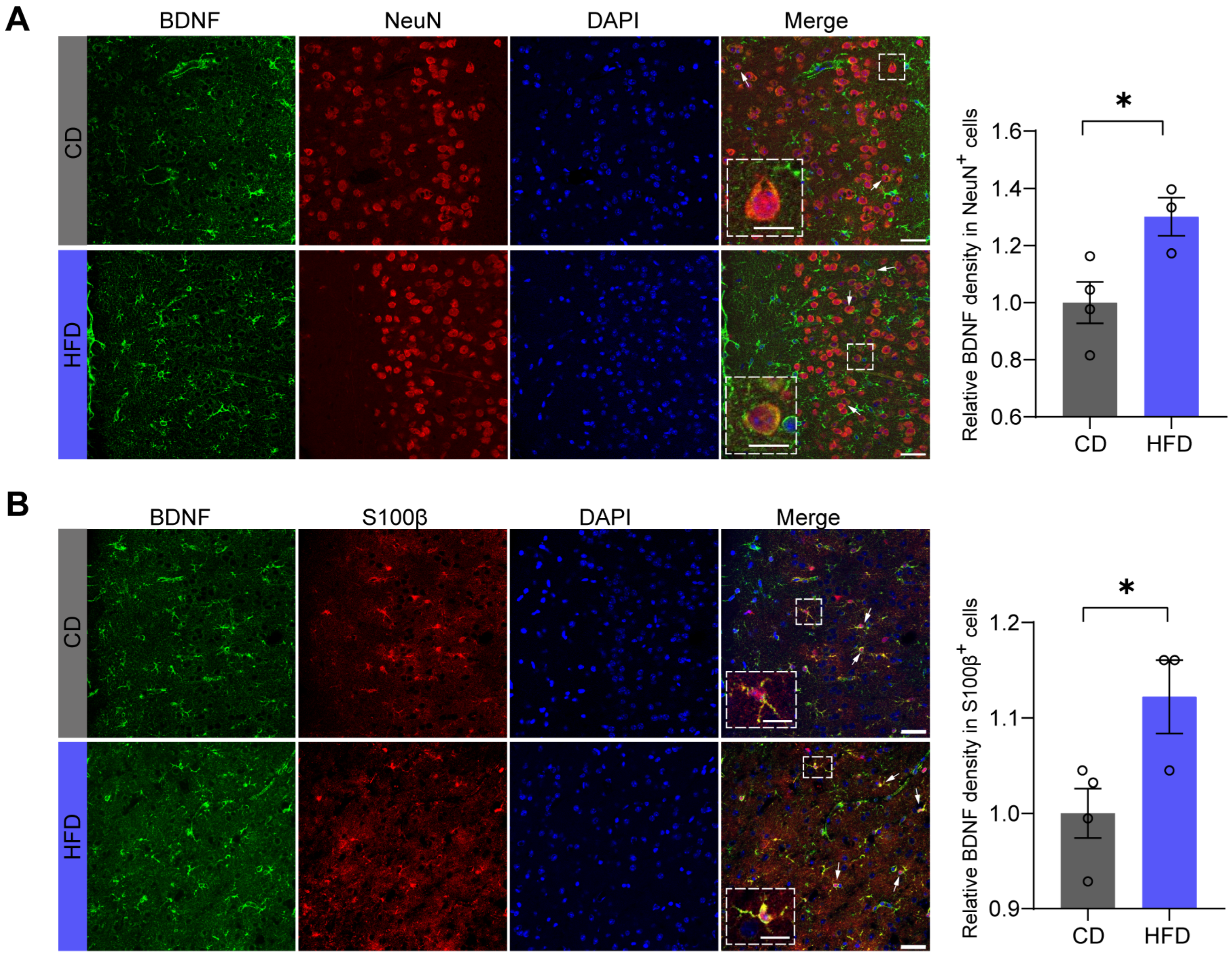
3.4. BDNF Levels Are Increased in Both Neurons and Astrocytes in the mPFC of Mice Fed an HFD
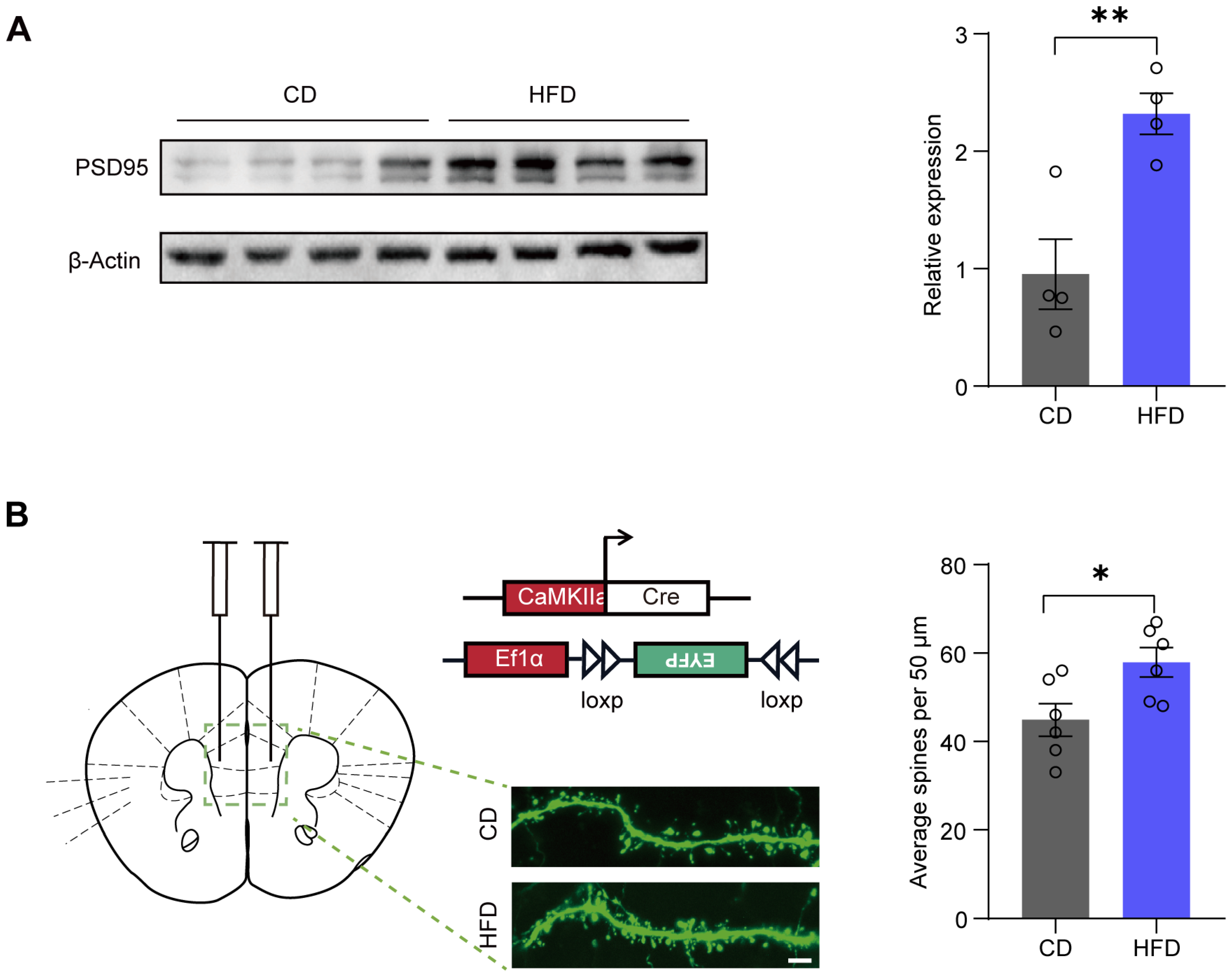
3.5. A Four-Week HFD Promotes the Expression of PSD95 and Increases the Density of Dendritic Spines
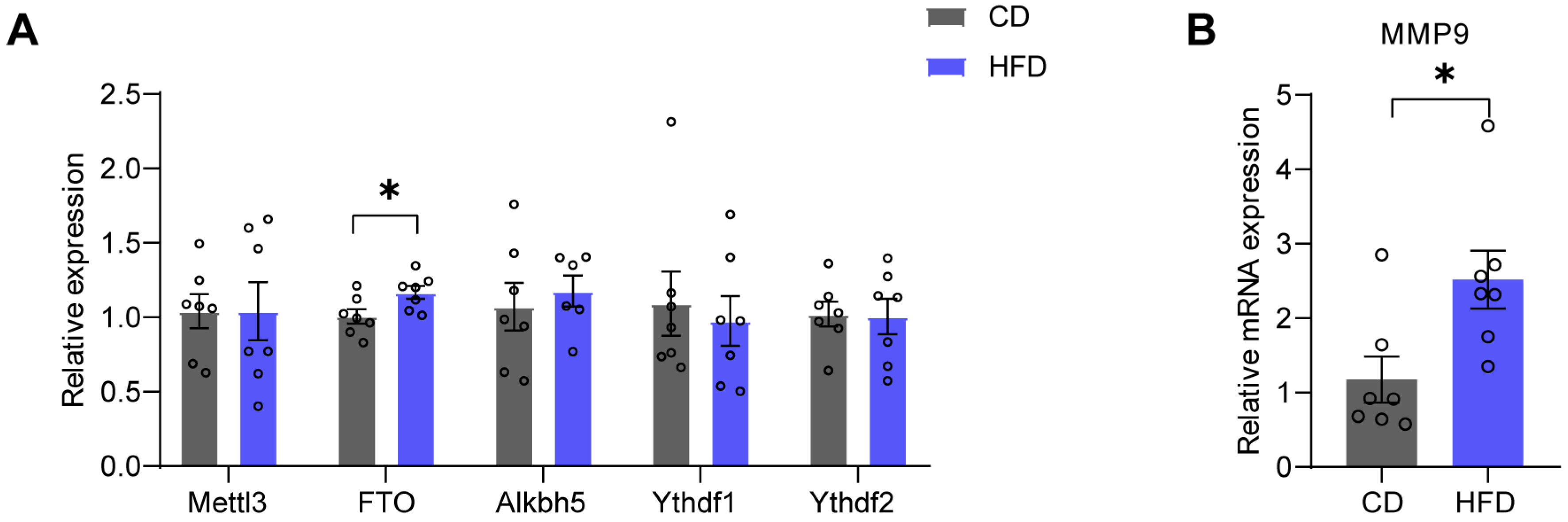
3.6. FTO and MMP9 Levels in the mPFC Are Increased in HFD Mice
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Pan, W.; Zhao, J.; Wu, J.; Xu, D.; Meng, X.; Jiang, P.; Shi, H.; Ge, X.; Yang, X.; Hu, M.; et al. Dimethyl itaconate ameliorates cognitive impairment induced by a high-fat diet via the gut-brain axis in mice. Microbiome 2023, 11, 30. [Google Scholar] [CrossRef]
- Chrobok, L.; Klich, J.D.; Sanetra, A.M.; Jeczmien-Lazur, J.S.; Pradel, K.; Palus-Chramiec, K.; Kepczynski, M.; Piggins, H.D.; Lewandowski, M.H. Rhythmic neuronal activities of the rat nucleus of the solitary tract are impaired by high-fat diet-implications for daily control of satiety. J. Physiol. Lond. 2022, 600, 751–767. [Google Scholar] [CrossRef] [PubMed]
- Vagena, E.; Ryu, J.K.; Baeza-Raja, B.; Walsh, N.M.; Syme, C.; Day, J.P.; Houslay, M.D.; Baillie, G.S. A high-fat diet promotes depression-like behavior in mice by suppressing hypothalamic PKA signaling. Transl. Psychiatry 2019, 9, 141. [Google Scholar] [CrossRef] [PubMed]
- de Noronha, S.R.; Campos, G.V.; Abreu, A.R.; de Souza, A.A.; Chianca, D.J.; de Menezes, R.C. High fat diet induced-obesity facilitates anxiety-like behaviors due to GABAergic impairment within the dorsomedial hypothalamus in rats. Behav. Brain Res. 2017, 316, 38–46. [Google Scholar] [CrossRef] [PubMed]
- Dutheil, S.; Ota, K.T.; Wohleb, E.S.; Rasmussen, K.; Duman, R.S. High-Fat Diet Induced Anxiety and Anhedonia: Impact on Brain Homeostasis and Inflammation. Neuropsychopharmacology 2016, 41, 1874–1887. [Google Scholar] [CrossRef] [PubMed]
- Finger, B.C.; Dinan, T.G.; Cryan, J.F. High-fat diet selectively protects against the effects of chronic social stress in the mouse. Neuroscience 2011, 192, 351–360. [Google Scholar] [CrossRef]
- Xu, L.; Xu, S.; Lin, L.; Gu, X.; Fu, C.; Fang, Y.; Li, X.; Wang, X. High-fat Diet Mediates Anxiolytic-like Behaviors in a Time-dependent Manner Through the Regulation of SIRT1 in the Brain. Neuroscience 2018, 372, 237–245. [Google Scholar] [CrossRef] [PubMed]
- Canetti, L.; Bachar, E.; Berry, E.M. Food and emotion. Behav. Process 2002, 60, 157–164. [Google Scholar] [CrossRef]
- Tomiyama, A.J.; Dallman, M.F.; Epel, E.S. Comfort food is comforting to those most stressed: Evidence of the chronic stress response network in high stress women. Psychoneuroendocrino 2011, 36, 1513–1519. [Google Scholar] [CrossRef]
- Xu, S.; Ju, Y.; Wei, X.; Ou, W.; Ma, M.; Lv, G.; Zhao, X.; Qin, Y.; Li, Y.; Li, L.; et al. Network analysis of suicide ideation and depression-anxiety symptoms among Chinese adolescents. Gen. Psychiatry 2024, 37, e101225. [Google Scholar] [CrossRef]
- Wang, C.S.; Kavalali, E.T.; Monteggia, L.M. BDNF signaling in context: From synaptic regulation to psychiatric disorders. Cell 2022, 185, 62–76. [Google Scholar] [CrossRef] [PubMed]
- Berton, O.; McClung, C.A.; Dileone, R.J.; Krishnan, V.; Renthal, W.; Russo, S.J.; Graham, D.; Tsankova, N.M.; Bolanos, C.A.; Rios, M.; et al. Essential role of BDNF in the mesolimbic dopamine pathway in social defeat stress. Science 2006, 311, 864–868. [Google Scholar] [CrossRef] [PubMed]
- Xu, H.; Steven, R.J.; Li, X.M. Dose-related effects of chronic antidepressants on neuroprotective proteins BDNF, Bcl-2 and Cu/Zn-SOD in rat hippocampus. Neuropsychopharmacology 2003, 28, 53–62. [Google Scholar] [CrossRef] [PubMed]
- Meng, F.; Liu, J.; Dai, J.; Wu, M.; Wang, W.; Liu, C.; Zhao, D.; Wang, H.; Zhang, J.; Li, M.; et al. Brain-derived neurotrophic factor in 5-HT neurons regulates susceptibility to depression-related behaviors induced by subchronic unpredictable stress. J. Psychiatry Res. 2020, 126, 55–66. [Google Scholar] [CrossRef] [PubMed]
- Yuan, Y.; Zhen, L.; Li, Z.; Xu, W.; Leng, H.; Xu, W.; Zheng, V.; Luria, V.; Pan, J.; Tao, Y.; et al. trans-Resveratrol ameliorates anxiety-like behaviors and neuropathic pain in mouse model of post-traumatic stress disorder. J. Psychopharmacol. 2020, 34, 726–736. [Google Scholar] [CrossRef] [PubMed]
- Shen, Z.; Zhu, J.; Yuan, Y.; Ren, L.; Qian, M.; Lin, M.; Cai, M.; Zhang, Z.; Shen, X. The roles of brain-derived neurotrophic factor (BDNF) and glial cell line-derived neurotrophic factor (GDNF) in predicting treatment remission in a Chinese Han population with generalized anxiety disorder. Psychiatry Res. 2019, 271, 319–324. [Google Scholar] [CrossRef]
- Carlino, D.; Francavilla, R.; Baj, G.; Kulak, K.; D’Adamo, P.; Ulivi, S.; Cappellani, S.; Gasparini, P.; Tongiorgi, E. Brain-derived neurotrophic factor serum levels in genetically isolated populations: Gender-specific association with anxiety disorder subtypes but not with anxiety levels or Val66Met polymorphism. PeerJ 2015, 3, e1252. [Google Scholar] [CrossRef] [PubMed]
- Lhullier, A.C.; Moreira, F.P.; Da, S.R.; Marques, M.B.; Bittencourt, G.; Pinheiro, R.T.; Souza, L.D.; Portela, L.V.; Lara, D.R.; Jansen, K.; et al. Increased serum neurotrophin levels related to alcohol use disorder in a young population sample. Alcohol. Clin. Exp. Res. 2015, 39, 30–33. [Google Scholar] [CrossRef]
- Pallanti, S.; Tofani, T.; Zanardelli, M.; Di Cesare, M.L.; Ghelardini, C. BDNF and Artemin are increased in drug-naive non-depressed GAD patients: Preliminary data. Int. J. Psychiatry Clin. 2014, 18, 255–260. [Google Scholar] [CrossRef]
- Chen, Z.Y.; Jing, D.; Bath, K.G.; Ieraci, A.; Khan, T.; Siao, C.J.; Herrera, D.G.; Toth, M.; Yang, C.; McEwen, B.S.; et al. Genetic variant BDNF (Val66Met) polymorphism alters anxiety-related behavior. Science 2006, 314, 140–143. [Google Scholar] [CrossRef]
- Jaehne, E.J.; Kent, J.N.; Antolasic, E.J.; Wright, B.J.; Spiers, J.G.; Creutzberg, K.C.; De Rosa, F.; Riva, M.A.; Sortwell, C.E.; Collier, T.J.; et al. Behavioral phenotyping of a rat model of the BDNF Val66Met polymorphism reveals selective impairment of fear memory. Transl. Psychiatry 2022, 12, 93. [Google Scholar] [CrossRef]
- Seibenhener, M.L.; Wooten, M.C. Use of the Open Field Maze to measure locomotor and anxiety-like behavior in mice. J. Vis. Exp. 2015, 96, e52434. [Google Scholar] [CrossRef]
- Ren, J.; Lu, C.L.; Huang, J.; Fan, J.; Guo, F.; Mo, J.W.; Huang, W.Y.; Kong, P.L.; Li, X.W.; Sun, L.R.; et al. A Distinct Metabolically Defined Central Nucleus Circuit Bidirectionally Controls Anxiety-Related Behaviors. J. Neurosci. 2022, 42, 2356–2370. [Google Scholar] [CrossRef]
- Garofalo, S.; Cocozza, G.; Mormino, A.; Bernardini, G.; Russo, E.; Ielpo, D.; Andolina, D.; Ventura, R.; Martinello, K.; Renzi, M.; et al. Natural killer cells and innate lymphoid cells 1 tune anxiety-like behavior and memory in mice via interferon-gamma and acetylcholine. Nat. Commun. 2023, 14, 3103. [Google Scholar] [CrossRef]
- Zhang, S.R.; Wu, J.L.; Chen, H.; Luo, R.; Chen, W.J.; Tang, L.J.; Li, X.W.; Yang, J.M.; Gao, T.M. ErbB4 knockdown in serotonergic neurons in the dorsal raphe induces anxiety-like behaviors. Neuropsychopharmacology 2020, 45, 1698–1706. [Google Scholar] [CrossRef]
- Bourin, M.; Hascoet, M. The mouse light/dark box test. Eur. J. Pharmacol. 2003, 463, 55–65. [Google Scholar] [CrossRef]
- Medina-Saldivar, C.; Pardo, G.; Pacheco-Otalora, L.F. Effect of MCH1, a fatty-acid amide hydrolase inhibitor, on the depressive-like behavior and gene expression of endocannabinoid and dopaminergic-signaling system in the mouse nucleus accumbens. Braz. J. Med. Biol. Res. 2024, 57, e12857. [Google Scholar] [CrossRef]
- Kong, Y.; Wang, Q.; Wu, D.Y.; Hu, J.; Zang, W.S.; Li, X.W.; Yang, J.M.; Gao, T.M. Involvement of P2X2 receptor in the medial prefrontal cortex in ATP modulation of the passive coping response to behavioral challenge. Genes Brain Behav. 2020, 19, e12691. [Google Scholar] [CrossRef]
- Fan, J.; Guo, F.; Mo, R.; Chen, L.Y.; Mo, J.W.; Lu, C.L.; Ren, J.; Zhong, Q.L.; Kuang, X.J.; Wen, Y.L.; et al. O-GlcNAc transferase in astrocytes modulates depression-related stress susceptibility through glutamatergic synaptic transmission. J. Clin. Investig. 2023, 133, 486–496. [Google Scholar] [CrossRef]
- Lin, R.; Wang, R.; Yuan, J.; Feng, Q.; Zhou, Y.; Zeng, S.; Ren, M.; Jiang, S.; Ni, H.; Zhou, C.; et al. Cell-type-specific and projection-specific brain-wide reconstruction of single neurons. Nat. Methods 2018, 15, 1033–1036. [Google Scholar] [CrossRef]
- Young, M.B.; Andero, R.; Ressler, K.J.; Howell, L.L. 3,4-Methylenedioxymethamphetamine facilitates fear extinction learning. Transl. Psychiatry 2015, 5, e634. [Google Scholar] [CrossRef] [PubMed]
- Nagahara, A.H.; Tuszynski, M.H. Potential therapeutic uses of BDNF in neurological and psychiatric disorders. Nat. Rev. Drug Discov. 2011, 10, 209–219. [Google Scholar] [CrossRef] [PubMed]
- Spychala, A.; Ruther, U. FTO affects hippocampal function by regulation of BDNF processing. PLoS ONE 2019, 14, e211937. [Google Scholar] [CrossRef]
- Maniam, J.; Morris, M.J. Long-term postpartum anxiety and depression-like behavior in mother rats subjected to maternal separation are ameliorated by palatable high fat diet. Behav. Brain Res. 2010, 208, 72–79. [Google Scholar] [CrossRef] [PubMed]
- Lam, Y.Y.; Tsai, S.F.; Chen, P.C.; Kuo, Y.M.; Chen, Y.W. Pioglitazone rescues high-fat diet-induced depression-like phenotypes and hippocampal astrocytic deficits in mice. Biomed. Pharmacother. 2021, 140, 111734. [Google Scholar] [CrossRef]
- Wu, H.; Zhang, W.; Huang, M.; Lin, X.; Chiou, J. Prolonged High-Fat Diet Consumption throughout Adulthood in Mice Induced Neurobehavioral Deterioration via Gut-Brain Axis. Nutrients 2023, 15, 392. [Google Scholar] [CrossRef]
- Andre, C.; Dinel, A.L.; Ferreira, G.; Laye, S.; Castanon, N. Diet-induced obesity progressively alters cognition, anxiety-like behavior and lipopolysaccharide-induced depressive-like behavior: Focus on brain indoleamine 2,3-dioxygenase activation. Brain Behav. Immun. 2014, 41, 10–21. [Google Scholar] [CrossRef]
- Virtuoso, A.; Tveden-Nyborg, P.; Schou-Pedersen, A.; Lykkesfeldt, J.; Muller, H.K.; Elfving, B.; Sorensen, D.B. A Long-Term Energy-Rich Diet Increases Prefrontal BDNF in Sprague-Dawley Rats. Nutrients 2021, 14, 126. [Google Scholar] [CrossRef]
- Miao, Z.; Mao, F.; Liang, J.; Szyf, M.; Wang, Y.; Sun, Z.S. Anxiety-Related Behaviours Associated with microRNA-206-3p and BDNF Expression in Pregnant Female Mice Following Psychological Social Stress. Mol. Neurobiol. 2018, 55, 1097–1111. [Google Scholar] [CrossRef]
- Wang, M.; Xie, Y.; Qin, D. Proteolytic cleavage of proBDNF to mBDNF in neuropsychiatric and neurodegenerative diseases. Brain Res. Bull. 2021, 166, 172–184. [Google Scholar] [CrossRef]
- Lee, R.; Kermani, P.; Teng, K.K.; Hempstead, B.L. Regulation of cell survival by secreted proneurotrophins. Science 2001, 294, 1945–1948. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Genes | Primers (5′–3′) | |
|---|---|---|
| Mettl3 | F | GGACACGTGGAGCTCTATCC |
| R | GCACGGGACTATCACTACGG | |
| FTO | F | TCAGCAGTGGCAGCTGAAAT |
| R | TCACCACGTCCCGAAACAAG | |
| Alkbh5 | F | CGGGAGAAGCTCAAGTCCAT |
| R | TATTTCCGCTTGGTGGTCCC | |
| Ythdf1 | F | GTGGACCCCCAGAGAACAAA |
| R | CCTCGCTGAGGGAGTAAGGA | |
| Ythdf2 | F | CCACAGGCAAGGCCGAATAA |
| R | GACCAAGCAGCTTCACCCAA | |
| MMP9 | F | GCAGAGGCATACTTGTACCG |
| R | TGATGTTATGATGGTCCCACTTG | |
| GAPDH | F | TCAAGAAGGTGGTGAAGCAG |
| R | TGGGAGTTGCTGTTGAAGTC |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Huang, H.; Huang, J.; Lu, W.; Huang, Y.; Luo, R.; Bathalian, L.; Chen, M.; Wang, X. A Four-Week High-Fat Diet Induces Anxiolytic-like Behaviors through Mature BDNF in the mPFC of Mice. Brain Sci. 2024, 14, 389. https://doi.org/10.3390/brainsci14040389
Huang H, Huang J, Lu W, Huang Y, Luo R, Bathalian L, Chen M, Wang X. A Four-Week High-Fat Diet Induces Anxiolytic-like Behaviors through Mature BDNF in the mPFC of Mice. Brain Sciences. 2024; 14(4):389. https://doi.org/10.3390/brainsci14040389
Chicago/Turabian StyleHuang, Huixian, Jia Huang, Wensi Lu, Yanjun Huang, Ran Luo, Luqman Bathalian, Ming Chen, and Xuemin Wang. 2024. "A Four-Week High-Fat Diet Induces Anxiolytic-like Behaviors through Mature BDNF in the mPFC of Mice" Brain Sciences 14, no. 4: 389. https://doi.org/10.3390/brainsci14040389
APA StyleHuang, H., Huang, J., Lu, W., Huang, Y., Luo, R., Bathalian, L., Chen, M., & Wang, X. (2024). A Four-Week High-Fat Diet Induces Anxiolytic-like Behaviors through Mature BDNF in the mPFC of Mice. Brain Sciences, 14(4), 389. https://doi.org/10.3390/brainsci14040389