
Effect of Experimental Hand Pain on Training-Induced Changes in Motor Performance and Corticospinal Excitability



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Subjects
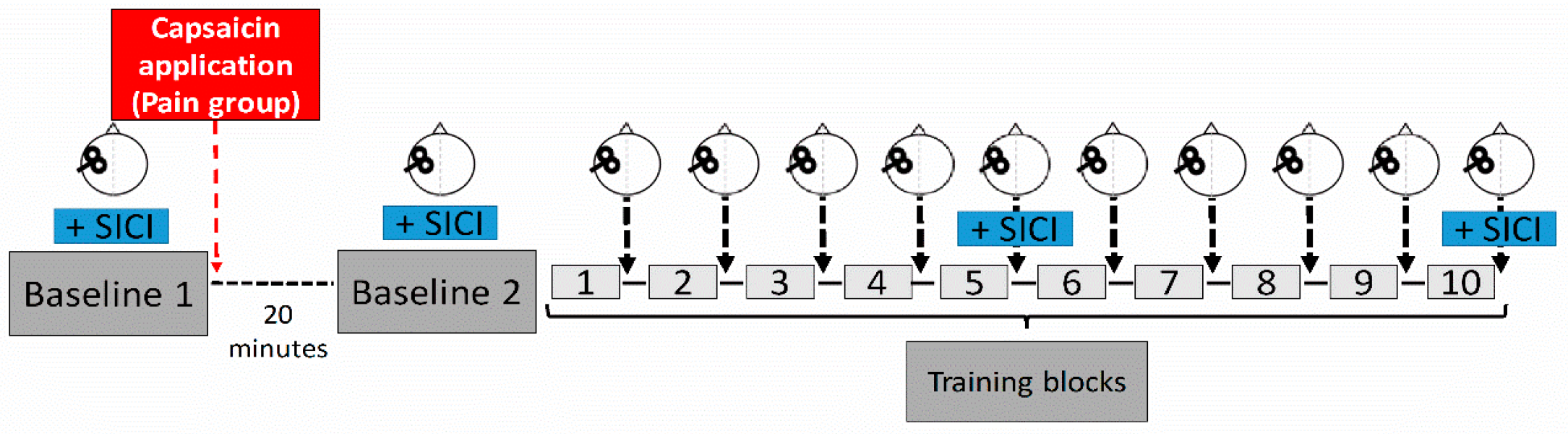
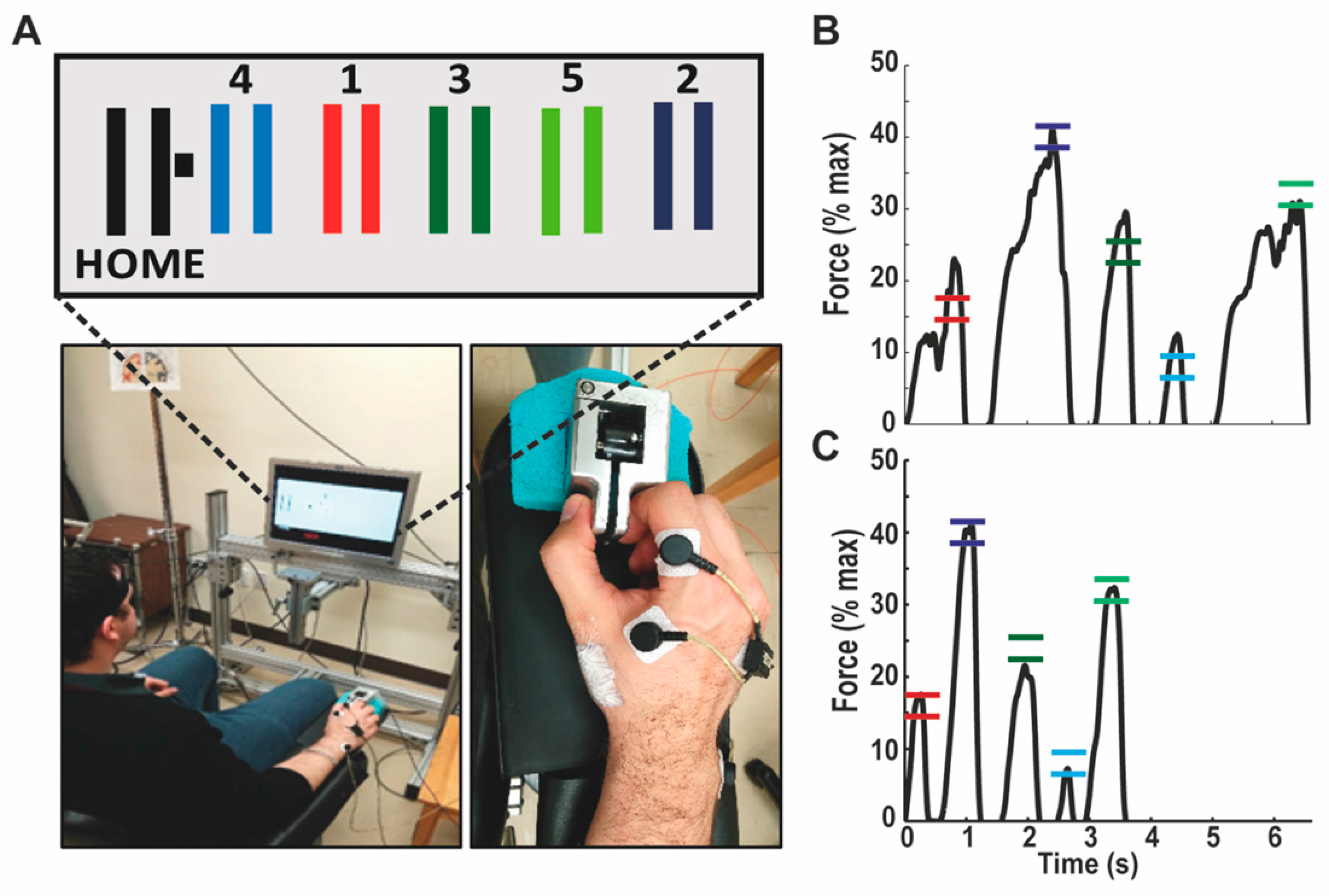
2.2. Experimental Design & Motor Task
2.3. Experimental Pain Model
2.4. EMG Recording and Neurophysiological Measures
2.5. Data Analysis
2.5.1. Behavioral Variables
2.5.2. Neurophysiological Variables
2.5.3. Statistical Analysis
3. Results
3.1. Group Characteristics
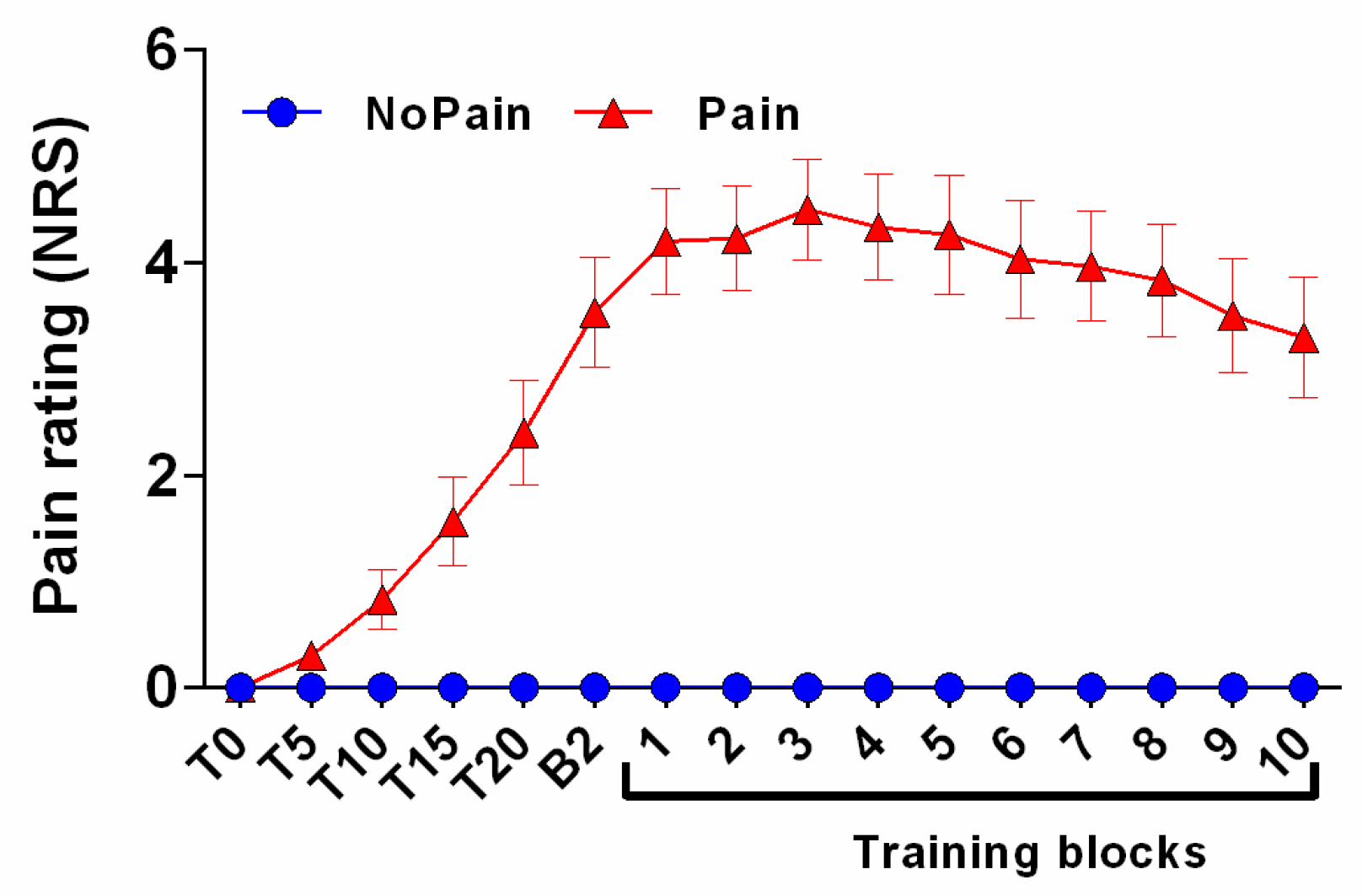
3.2. Pain Rating
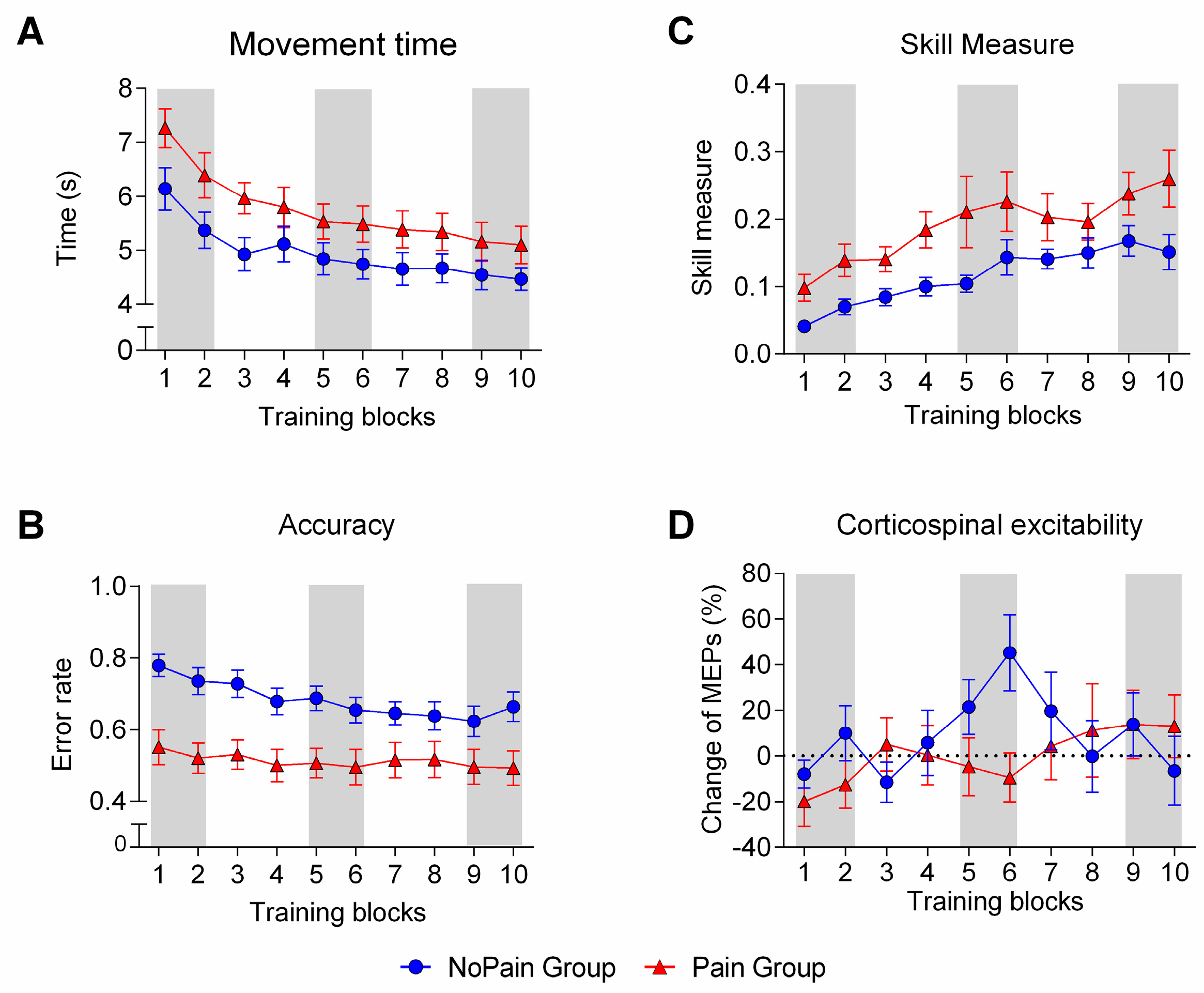
3.3. Behavioral Outcomes
3.4. Neurophysiological Outcomes
3.4.1. Pain-Related Changes
3.4.2. Training-Related Changes
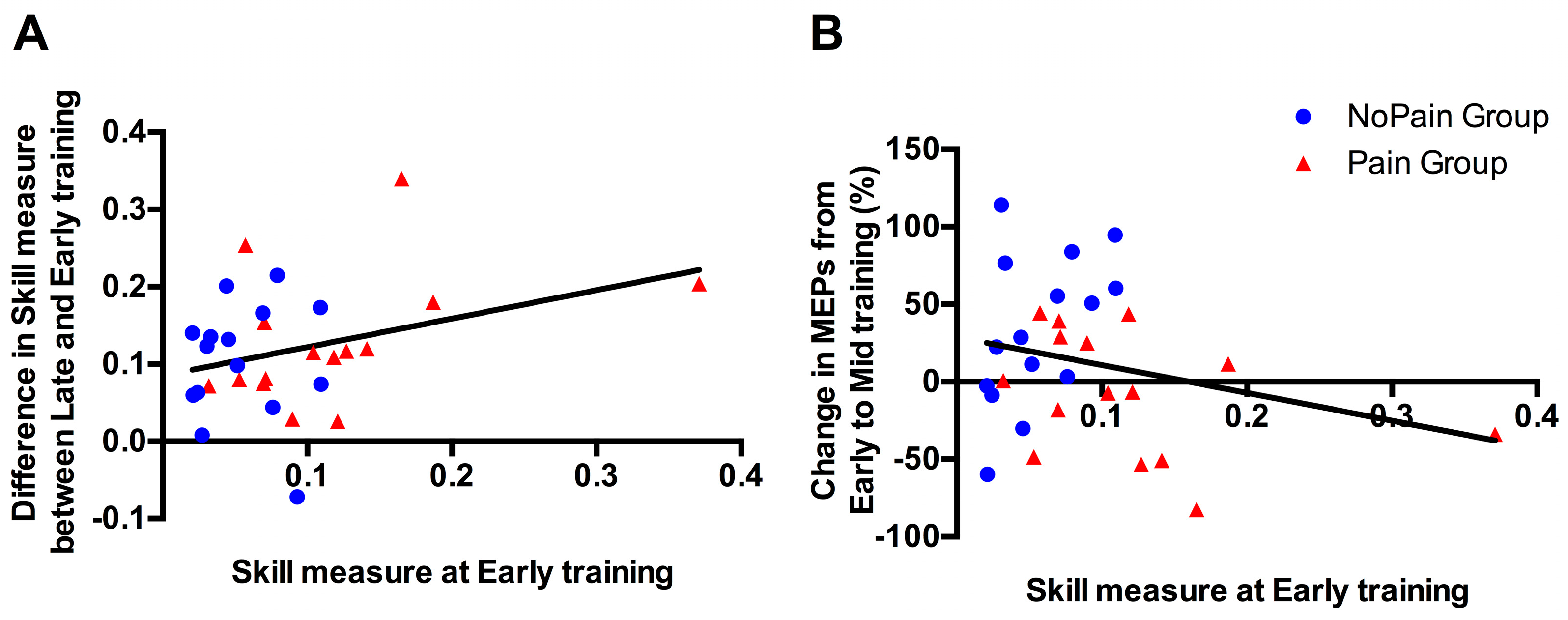
3.5. Correlational Analyses between Initial Performance and Training-Related Changes
3.6. Background EMG Levels throughout the Experiment
4. Discussion
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Dayan, E.; Cohen, L.G. Neuroplasticity subserving motor skill learning. Neuron 2011, 72, 443–454. [Google Scholar] [CrossRef] [PubMed]
- Della-Maggiore, V.; Landi, S.M.; Villalta, J.I. Sensorimotor adaptation: Multiple forms of plasticity in motor circuits. Neuroscientist 2015, 21, 109–125. [Google Scholar] [CrossRef] [PubMed]
- Doyon, J.; Benali, H. Reorganization and plasticity in the adult brain during learning of motor skills. Curr. Opin. Neurobiol. 2005, 15, 161–167. [Google Scholar] [CrossRef] [PubMed]
- Abraham, W.C. Metaplasticity: Tuning synapses and networks for plasticity. Nat. Rev. Neurosci. 2008, 9, 387. [Google Scholar] [CrossRef] [PubMed]
- Todd, G.; Ridding, M.C. The response to repetitive stimulation of human motor cortex is influenced by the history of synaptic activity. Restor. Neurol. Neurosci. 2010, 28, 459–467. [Google Scholar] [PubMed]
- Duffy, S.N.; Craddock, K.J.; Abel, T.; Nguyen, P.V. Environmental enrichment modifies the pka-dependence of hippocampal ltp and improves hippocampus-dependent memory. Learn Mem. 2001, 8, 26–34. [Google Scholar] [CrossRef] [PubMed]
- Van Praag, H.; Christie, B.R.; Sejnowski, T.J.; Gage, F.H. Running enhances neurogenesis, learning, and long-term potentiation in mice. Proc. Natl. Acad. Sci. USA 1999, 96, 13427–13431. [Google Scholar] [CrossRef] [PubMed]
- Foster, T.C.; Fugger, H.N.; Cunningham, S.G. Receptor blockade reveals a correspondence between hippocampal-dependent behavior and experience-dependent synaptic enhancement. Brain Res. 2000, 871, 39–43. [Google Scholar] [CrossRef]
- Garcia, R.; Musleh, W.; Tocco, G.; Thompson, R.F.; Baudry, M. Time-dependent blockade of STP and LTP in hippocampal slices following acute stress in mice. Neurosci. Lett. 1997, 233, 41–44. [Google Scholar] [CrossRef]
- Kim, J.J.; Foy, M.R.; Thompson, R.F. Behavioral stress modifies hippocampal plasticity through N-methyl-d-aspartate receptor activation. Proc. Natl. Acad. Sci. USA 1996, 93, 4750–4753. [Google Scholar] [CrossRef] [PubMed]
- Karabanov, A.; Ziemann, U.; Hamada, M.; George, M.S.; Quartarone, A.; Classen, J.; Massimini, M.; Rothwell, J.; Siebner, H.R. Consensus paper: Probing homeostatic plasticity of human cortex with non-invasive transcranial brain stimulation. Brain Stimul. 2015, 8, 993–1006. [Google Scholar] [CrossRef] [PubMed]
- Muller-Dahlhaus, F.; Ziemann, U. Metaplasticity in human cortex. Neuroscientist 2015, 21, 185–202. [Google Scholar] [CrossRef] [PubMed]
- Gentner, R.; Wankerl, K.; Reinsberger, C.; Zeller, D.; Classen, J. Depression of human corticospinal excitability induced by magnetic theta-burst stimulation: Evidence of rapid polarity-reversing metaplasticity. Cereb. Cortex 2008, 18, 2046–2053. [Google Scholar] [CrossRef] [PubMed]
- Iezzi, E.; Conte, A.; Suppa, A.; Agostino, R.; Dinapoli, L.; Scontrini, A.; Berardelli, A. Phasic voluntary movements reverse the aftereffects of subsequent theta-burst stimulation in humans. J. Neurophysiol. 2008, 100, 2070–2076. [Google Scholar] [CrossRef] [PubMed]
- Stefan, K.; Wycislo, M.; Gentner, R.; Schramm, A.; Naumann, M.; Reiners, K.; Classen, J. Temporary occlusion of associative motor cortical plasticity by prior dynamic motor training. Cereb. Cortex 2006, 16, 376–385. [Google Scholar] [CrossRef] [PubMed]
- Ziemann, U.; Ilic, T.V.; Pauli, C.; Meintzschel, F.; Ruge, D. Learning modifies subsequent induction of long-term potentiation-like and long-term depression-like plasticity in human motor cortex. J. Neurosci. 2004, 24, 1666–1672. [Google Scholar] [CrossRef] [PubMed]
- Ziemann, U.; Muellbacher, W.; Hallett, M.; Cohen, L.G. Modulation of practice-dependent plasticity in human motor cortex. Brain J. Neurol. 2001, 124, 1171–1181. [Google Scholar] [CrossRef]
- Boudreau, S.; Romaniello, A.; Wang, K.; Svensson, P.; Sessle, B.J.; Arendt-Nielsen, L. The effects of intra-oral pain on motor cortex neuroplasticity associated with short-term novel tongue-protrusion training in humans. Pain 2007, 132, 169–178. [Google Scholar] [CrossRef] [PubMed]
- Mavromatis, N.; Gagne, M.; Voisin, J.I.; Reilly, K.T.; Mercier, C. Experimental tonic hand pain modulates the corticospinal plasticity induced by a subsequent hand deafferentation. Neuroscience 2016, 330, 403–409. [Google Scholar] [CrossRef] [PubMed]
- Bank, P.J.; Peper, C.E.; Marinus, J.; Beek, P.J.; van Hilten, J.J. Motor consequences of experimentally induced limb pain: A systematic review. Eur. J. Pain 2013, 17, 145–157. [Google Scholar] [CrossRef] [PubMed]
- Hodges, P.W.; Tucker, K. Moving differently in pain: A new theory to explain the adaptation to pain. Pain 2011, 152, S90–S98. [Google Scholar] [CrossRef] [PubMed]
- Burns, E.; Chipchase, L.S.; Schabrun, S.M. Primary sensory and motor cortex function in response to acute muscle pain: A systematic review and meta-analysis. Eur. J. Pain 2016, 20, 1203–1213. [Google Scholar] [CrossRef] [PubMed]
- Bouffard, J.; Bouyer, L.J.; Roy, J.S.; Mercier, C. Tonic pain experienced during locomotor training impairs retention despite normal performance during acquisition. J. Neurosci. 2014, 34, 9190–9195. [Google Scholar] [CrossRef] [PubMed]
- Lamothe, M.; Roy, J.S.; Bouffard, J.; Gagne, M.; Bouyer, L.J.; Mercier, C. Effect of tonic pain on motor acquisition and retention while learning to reach in a force field. PLoS ONE 2014, 9, e99159. [Google Scholar] [CrossRef] [PubMed]
- Bilodeau, M.C.; Roosink, M.; Mercier, C. Effect of local versus remote tonic heat pain during training on acquisition and retention of a finger-tapping sequence task. Exp. Brain Res. 2016, 234, 475–482. [Google Scholar] [CrossRef] [PubMed]
- Ingham, D.; Tucker, K.J.; Tsao, H.; Hodges, P.W. The effect of pain on training-induced plasticity of the corticomotor system. Eur. J. Pain 2011, 15, 1028–1034. [Google Scholar] [CrossRef] [PubMed]
- Rittig-Rasmussen, B.; Kasch, H.; Fuglsang-Frederiksen, A.; Svensson, P.; Jensen, T.S. The role of neuroplasticity in experimental neck pain: A study of potential mechanisms impeding clinical outcomes of training. Man. Ther. 2014, 19, 288–293. [Google Scholar] [CrossRef] [PubMed]
- Dancey, E.; Murphy, B.; Srbely, J.; Yielder, P. The effect of experimental pain on motor training performance and sensorimotor integration. Exp. Brain Res. 2014, 232, 2879–2889. [Google Scholar] [CrossRef] [PubMed]
- Dancey, E.; Murphy, B.A.; Andrew, D.; Yielder, P. The effect of local vs remote experimental pain on motor learning and sensorimotor integration using a complex typing task. Pain 2016, 157, 1682–1695. [Google Scholar] [CrossRef] [PubMed]
- Dancey, E.; Murphy, B.; Andrew, D.; Yielder, P. The interactive effect of acute pain and motor learning acquisition on sensorimotor integration and motor learning outcomes. J. Neurophysiol. 2016, 116, 2210–2220. [Google Scholar] [CrossRef] [PubMed]
- Bonnard, M.; Gallea, C.; de Graaf, J.B.; Pailhous, J. Corticospinal control of the thumb-index grip depends on precision of force control: A transcranial magnetic stimulation and functional magnetic resonance imagery study in humans. Eur. J. Neurosci. 2007, 25, 872–880. [Google Scholar] [CrossRef] [PubMed]
- Reis, J.; Schambra, H.M.; Cohen, L.G.; Buch, E.R.; Fritsch, B.; Zarahn, E.; Celnik, P.A.; Krakauer, J.W. Noninvasive cortical stimulation enhances motor skill acquisition over multiple days through an effect on consolidation. Proc. Natl. Acad. Sci. USA 2009, 106, 1590–1595. [Google Scholar] [CrossRef] [PubMed]
- Farina, S.; Valeriani, M.; Rosso, T.; Aglioti, S.; Tamburin, S.; Fiaschi, A.; Tinazzi, M. Transient inhibition of the human motor cortex by capsaicin-induced pain. A study with transcranial magnetic stimulation. Neurosci. Lett. 2001, 314, 97–101. [Google Scholar] [CrossRef]
- Fierro, B.; de Tommaso, M.; Giglia, F.; Giglia, G.; Palermo, A.; Brighina, F. Repetitive transcranial magnetic stimulation (RTMS) of the dorsolateral prefrontal cortex (DLPFC) during capsaicin-induced pain: Modulatory effects on motor cortex excitability. Exp. Brain Res. 2010, 203, 31–38. [Google Scholar] [CrossRef] [PubMed]
- Garry, M.I.; Thomson, R.H.S. The effect of test TMS intensity on short-interval intracortical inhibition in different excitability states. Exp. Brain Res. 2009, 193, 267–274. [Google Scholar] [CrossRef] [PubMed]
- Rice, D.A.; Graven-Nielsen, T.; Lewis, G.N.; McNair, P.J.; Dalbeth, N. The effects of experimental knee pain on lower limb corticospinal and motor cortex excitability. Arthritis Res. Ther. 2015, 17, 204. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rice, D.A.; McNair, P.J.; Lewis, G.N.; Dalbeth, N. Quadriceps arthrogenic muscle inhibition: The effects of experimental knee joint effusion on motor cortex excitability. Arthritis Res. Ther. 2014, 16, 502. [Google Scholar] [CrossRef] [PubMed]
- Schabrun, S.M.; Hodges, P.W. Muscle pain differentially modulates short interval intracortical inhibition and intracortical facilitation in primary motor cortex. J. Pain. 2012, 13, 187–194. [Google Scholar] [CrossRef] [PubMed]
- Pascual-Leone, A.; Grafman, J.; Hallett, M. Modulation of cortical motor output maps during development of implicit and explicit knowledge. Science 1994, 263, 1287–1289. [Google Scholar] [CrossRef] [PubMed]
- Muellbacher, W.; Ziemann, U.; Boroojerdi, B.; Cohen, L.; Hallett, M. Role of the human motor cortex in rapid motor learning. Exp. Brain Res. 2001, 136, 431–438. [Google Scholar] [CrossRef] [PubMed]
- Orban de Xivry, J.J.; Ahmadi-Pajouh, M.A.; Harran, M.D.; Salimpour, Y.; Shadmehr, R. Changes in corticospinal excitability during reach adaptation in force fields. J. Neurophysiol. 2013, 109, 124–136. [Google Scholar] [CrossRef] [PubMed]
- Bouffard, J.; Bouyer, L.J.; Roy, J.S.; Mercier, C. Pain induced during both the acquisition and retention phases of locomotor adaptation does not interfere with improvements in motor performance. Neural Plast. 2016, 2016, 8539096. [Google Scholar] [CrossRef] [PubMed]
- Conte, A.; Gilio, F.; Iezzi, E.; Frasca, V.; Inghilleri, M.; Berardelli, A. Attention influences the excitability of cortical motor areas in healthy humans. Exp. Brain Res. 2007, 182, 109–117. [Google Scholar] [CrossRef] [PubMed]
- Stefan, K.; Wycislo, M.; Classen, J. Modulation of associative human motor cortical plasticity by attention. J. Neurophysiol. 2004, 92, 66–72. [Google Scholar] [CrossRef] [PubMed]





© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license ( http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mavromatis, N.; Neige, C.; Gagné, M.; Reilly, K.T.; Mercier, C. Effect of Experimental Hand Pain on Training-Induced Changes in Motor Performance and Corticospinal Excitability. Brain Sci. 2017, 7, 15. https://doi.org/10.3390/brainsci7020015
Mavromatis N, Neige C, Gagné M, Reilly KT, Mercier C. Effect of Experimental Hand Pain on Training-Induced Changes in Motor Performance and Corticospinal Excitability. Brain Sciences. 2017; 7(2):15. https://doi.org/10.3390/brainsci7020015
Chicago/Turabian StyleMavromatis, Nicolas, Cécilia Neige, Martin Gagné, Karen T. Reilly, and Catherine Mercier. 2017. "Effect of Experimental Hand Pain on Training-Induced Changes in Motor Performance and Corticospinal Excitability" Brain Sciences 7, no. 2: 15. https://doi.org/10.3390/brainsci7020015
APA StyleMavromatis, N., Neige, C., Gagné, M., Reilly, K. T., & Mercier, C. (2017). Effect of Experimental Hand Pain on Training-Induced Changes in Motor Performance and Corticospinal Excitability. Brain Sciences, 7(2), 15. https://doi.org/10.3390/brainsci7020015