Do EEG and Startle Reflex Modulation Vary with Self-Reported Aggression in Response to Violent Images?



Abstract
:1. Introduction
2. Materials and Methods
2.1. Participants
2.2. Participant Demographics
2.3. Measures
2.3.1. Online Questionnaire
2.3.2. Stimuli
2.3.3. EEG Recordings
2.3.4. Startle Reflex Modulation Recordings
2.3.5. Explicit Responses
2.4. Experimental Task and Procedure
2.5. Statistical Analysis
3. Results
3.1. Self-Reported Aggression, Impulsivity and Self-Monitoring
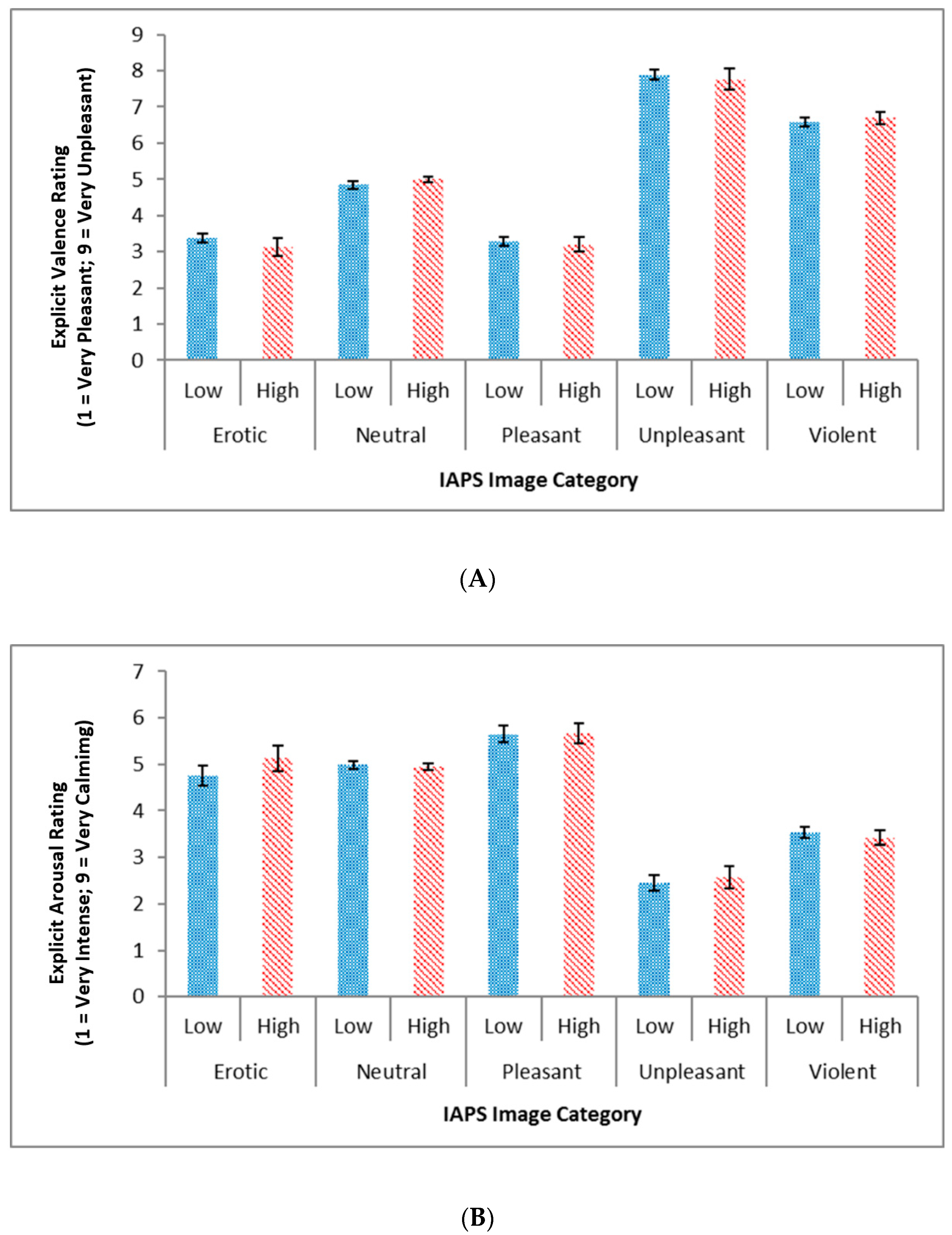
3.2. Explicit Responses
3.3. Physiological Measures
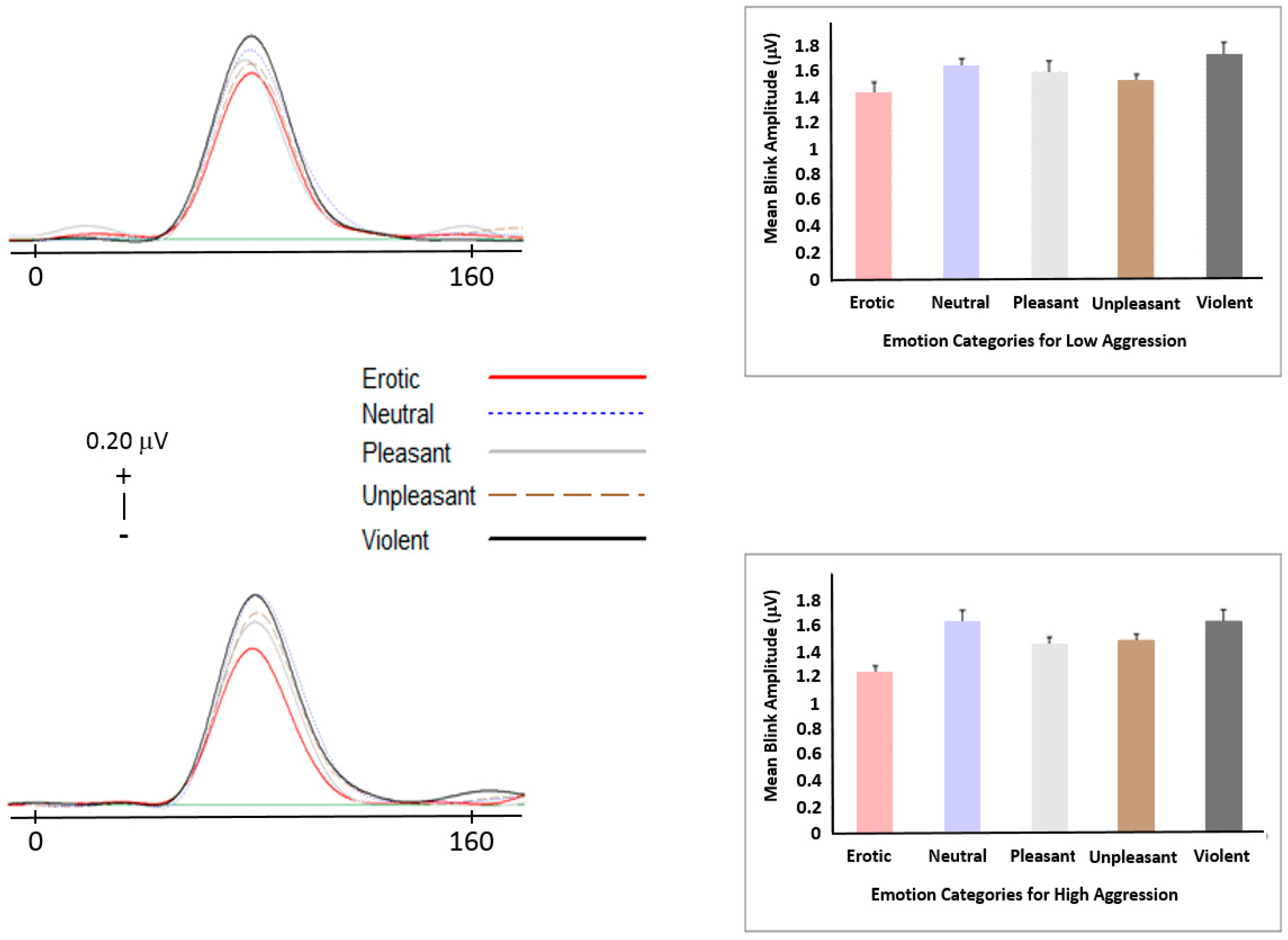
3.3.1. Startle Reflex Modulation (SRM)
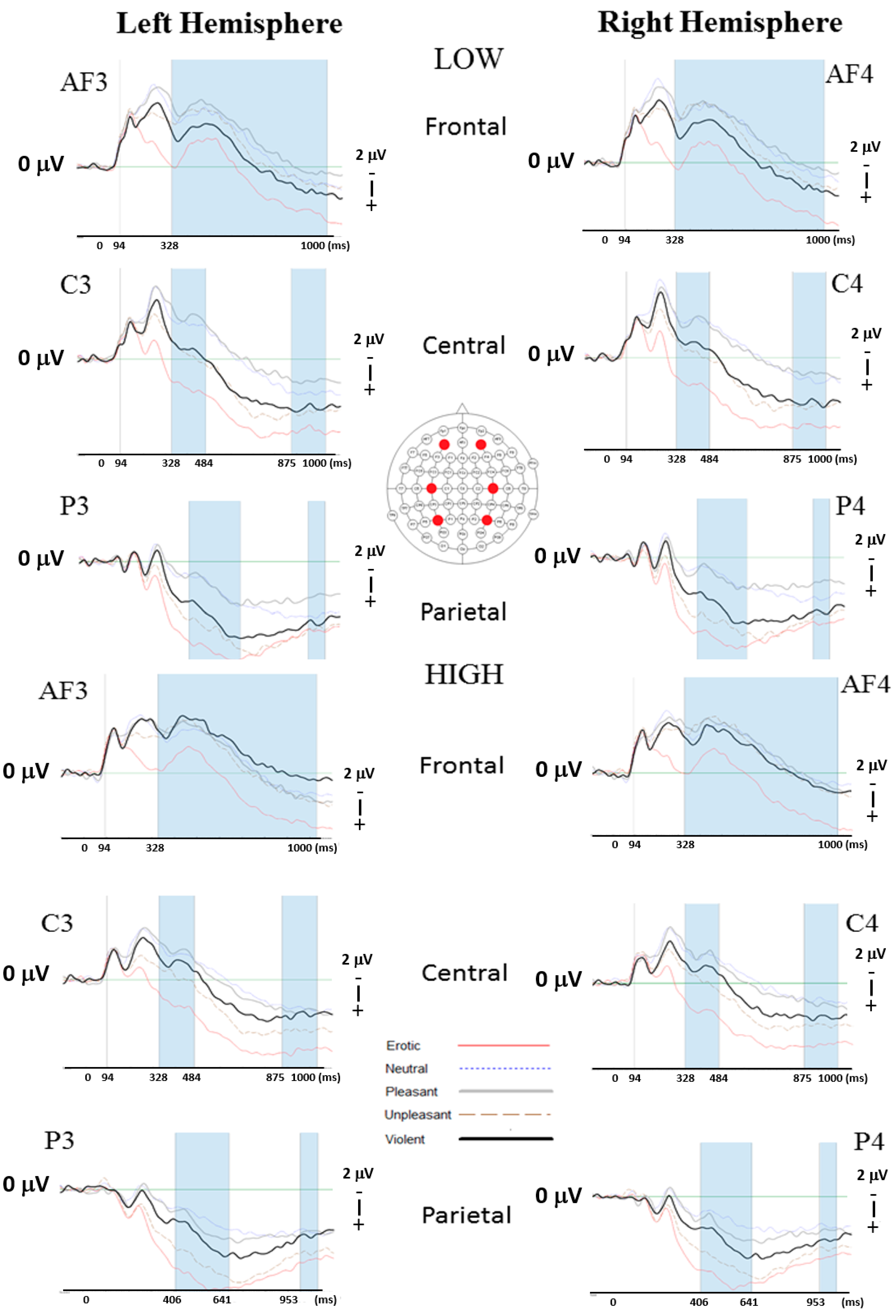
3.3.2. Event-Related Potentials (ERPs)
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Siegel, A.; Victoroff, J. Understanding human aggression: New insights from neuroscience. Int. J. Law Psychiatry 2009, 32, 209–215. [Google Scholar] [CrossRef] [PubMed]
- Barratt, E.S.; Monahan, J.; Steadman, H. Impulsiveness and aggression. Violence Ment. Disord. Dev. Risk Assess. 1994, 10, 61–79. [Google Scholar]
- Eron, L.D. The development of aggressive behavior from the perspective of a developing behaviorism. Am. Psychol. 1987, 42, 435–442. [Google Scholar] [CrossRef] [PubMed]
- American Psychiatric Association. Diagnostic and Statistical Manual of Mental Disorders (DSM-5); American Psychiatric Pub: Washington, DC, USA, 2013. [Google Scholar]
- The ICD-10 Classification of Mental and Behavioural Disorders: Diagnostic Criteria for Research; World Health Organization: Geneva, Switzerland, 1993.
- Fulmer, S.M.; Frijters, J.C. A Review of Self-Report and Alternative Approaches in the Measurement of Student Motivation. Educ. Psychol. Rev. 2009, 21, 219–246. [Google Scholar] [CrossRef]
- Van de Mortel, T.F. Faking it: Social desirability response bias in self-report research. Aust. J. Adv. Nurs. 2008, 25, 40–48. [Google Scholar]
- Hebert, J.R.; Clemow, L.; Pbert, L.; Ockene, I.S. Social Desirability Bias in Dietary Self-Report May Compromise the Validity of Dietary Intake Measures. Int. J. Epidemiol. 1995, 24, 389–398. [Google Scholar] [CrossRef]
- Kelly, C.A.; Soler-Hampejsek, E.; Mensch, B.S.; Hewett, P.C. Social desirability bias in sexual behavior reporting: Evidence from an interview mode experiment in rural Malawi. Int. Perspect. Sex. Reprod. Health 2013, 39, 14–21. [Google Scholar] [CrossRef]
- Walla, P. Non-Conscious Brain Processes Revealed by Magnetoencephalography (MEG). In Magnetoencephalography; IntechOpen: London, UK, 2011. [Google Scholar] [Green Version]
- Walla, P.; Brenner, G.; Koller, M. Objective Measures of Emotion Related to Brand Attitude: A New Way to Quantify Emotion-Related Aspects Relevant to Marketing. PLoS ONE 2011, 6, e26782. [Google Scholar] [CrossRef]
- Berridge, K.; Winkielman, P. What is an unconscious emotion? (The case for unconscious “liking”). Cogn. Emot. 2003, 17, 181–211. [Google Scholar] [CrossRef]
- Winkielman, P.; Berridge, K.C. Unconscious Emotion. Curr. Dir. Psychol. Sci. 2004, 13, 120–123. [Google Scholar] [CrossRef]
- Tamietto, M.; De Gelder, B. Neural bases of the non-conscious perception of emotional signals. Nat. Rev. Neurosci. 2010, 11, 697–709. [Google Scholar] [CrossRef] [PubMed]
- Moyer, K.E. Kinds of aggression and their physiological basis. Commun. Behav. Biol. 1968, 2, 65–87. [Google Scholar]
- Patton, J.H.; Stanford, M.S.; Barratt, E.S. Factor structure of the barratt impulsiveness scale. J. Clin. Psychol. 1995, 51, 768–774. [Google Scholar] [CrossRef]
- Walla, P.; Koller, M. Emotion is not What You Think It Is: Startle Reflex Modulation (SRM) as a Measure of Affective Processing in NeuroIs. In Information Systems and Neuroscience; Springer: Cham, Switzerland, 2015; pp. 181–186. [Google Scholar]
- Lang, P.J.; Bradley, M.M.; Cuthbert, B.N. Emotion, attention, and the startle reflex. Psychol. Rev. 1990, 97, 377–395. [Google Scholar] [CrossRef] [PubMed]
- Mavratzakis, A.; Herbert, C.; Walla, P. Emotional facial expressions evoke faster orienting responses, but weaker emotional responses at neural and behavioural levels compared to scenes: A simultaneous EEG and facial EMG study. NeuroImage 2016, 124, 931–946. [Google Scholar] [CrossRef]
- Geiser, M.; Walla, P. Objective Measures of Emotion During Virtual Walks through Urban Environments. Appl. Sci. 2011, 1, 1–11. [Google Scholar] [CrossRef]
- Koller, M.; Walla, P. Towards Alternative Ways to Measure Attitudes Related to Consumption: Introducing Startle Reflex Modulation. J. Agric. Food Ind. Organ. 2015, 13, 83–88. [Google Scholar] [CrossRef]
- Nesbitt, K.; Blackmore, K.; Hookham, G.; Kay-Lambkin, F.; Walla, P. Using the Startle Eye-Blink to Measure Affect in Players. In Serious Games Analytics; Springer Science and Business Media LLC: Cham, Switzerland, 2015; pp. 401–434. [Google Scholar]
- Walla, P.; Koller, M.; Meier, J.L. Consumer neuroscience to inform consumers-physiological methods to identify attitude formation related to over-consumption and environmental damage. Front. Hum. Neurosci. 2014, 8, 304. [Google Scholar] [CrossRef]
- Grahl, A.; Greiner, U.; Walla, P. Bottle Shape Elicits Gender-Specific Emotion: A Startle Reflex Modulation Study. Psychology 2012, 3, 548–554. [Google Scholar] [CrossRef] [Green Version]
- Allen, N.B.; Trinder, J.; Brennan, C. Affective startle modulation in clinical depression: Preliminary findings. Biol. Psychiatry 1999, 46, 542–550. [Google Scholar] [CrossRef]
- Walla, P.; Koller, M.; Brenner, G.; Bosshard, S. Evaluative conditioning of established brands: Implicit measures reveal other effects than explicit measures. J. Neurosci. Psychol. Econ. 2017, 10, 24–41. [Google Scholar] [CrossRef]
- Patrick, C.J. Psychophysiological correlates of aggression and violence: An integrative review. Philos. Trans. R. Soc. B: Biol. Sci. 2008, 363, 2543–2555. [Google Scholar] [CrossRef] [PubMed]
- Donchin, E.; Coles, M.G.H. Is the P300 component a manifestation of context updating? Behav. Brain Sci. 1988, 11, 357–374. [Google Scholar] [CrossRef]
- Hillyard, S.A.; Kutas, M. Electrophysiology of Cognitive Processing. Annu. Rev. Psychol. 1983, 34, 33–61. [Google Scholar] [CrossRef] [Green Version]
- Harmon-Jones, E.; Barratt, E.S.; Wigg, C. Impulsiveness, aggression, reading, and the P300 of the event-related potential. Pers. Individ. Differ. 1997, 22, 439–445. [Google Scholar] [CrossRef]
- Bond, A.J.; Surguy, S.M. Relationship between attitudinal hostility and P300 latencies. Prog. Neuro-Psychopharmacol. Biol. Psychiatry 2000, 24, 1277–1288. [Google Scholar] [CrossRef]
- Gerstle, J.E.; Mathias, C.W.; Stanford, M.S. Auditory P300 and self-reported impulsive aggression. Prog. Neuro-Psychopharmacol. Biol. Psychiatry 1998, 22, 575–583. [Google Scholar] [CrossRef]
- Stanford, M.S.; Houston, R.J.; Villemarette-Pittman, N.R.; Greve, K.W. Premeditated aggression: Clinical assessment and cognitive psychophysiology. Pers. Individ. Differ. 2003, 34, 773–781. [Google Scholar] [CrossRef]
- Barratt, E.S.; Stanford, M.S.; Kent, T.A.; Alan, F. Neuropsychological and cognitive psychophysiological substrates of impulsive aggression. Biol. Psychiatry 1997, 41, 1045–1061. [Google Scholar] [CrossRef]
- Koller, M.; Walla, P. Measuring Affective Information Processing in Information Systems and Consumer Research—Introducing Startle Reflex Modulation. In Proceedings of the 23th International Conference on Information Systems (ICIS), Orlando, FL, USA, 14 December 2012. [Google Scholar]
- Gagnon, J.; Aubin, M.; Emond, F.C.; Derguy, S.; Brochu, A.F.; Bessette, M.; Jolicoeur, P. An ERP study on hostile attribution bias in aggressive and nonaggressive individuals. Aggress. Behav. 2016, 43, 217–229. [Google Scholar] [CrossRef]
- Schupp, H.T.; Flaisch, T.; Stockburger, J.; Junghöfer, M. Emotion and attention: Event-related brain potential studies. In Motivation—Theory, Neurobiology and Applications; Elsevier BV: Amsterdam, The Netherlands, 2006; Volume 156, pp. 31–51. [Google Scholar]
- Patrick, C.J.; Bradley, M.M.; Lang, P.J. Emotion in the criminal psychopath: Startle reflex modulation. J. Abnorm. Psychol. 1993, 102, 82–92. [Google Scholar] [CrossRef] [PubMed]
- Vaidyanathan, U.; Hall, J.R.; Patrick, C.J.; Bernat, E.M. Clarifying the role of defensive reactivity deficits in psychopathy and antisocial personality using startle reflex methodology. J. Abnorm. Psychol. 2011, 120, 253–258. [Google Scholar] [CrossRef] [PubMed]
- Dawson, M.E.; Hazlett, E.A.; Filion, D.L.; Nuechterlein, K.H.; Al, E. Attention and schizophrenia: Impaired modulation of the startle reflex. J. Abnorm. Psychol. 1993, 102, 633–641. [Google Scholar] [CrossRef] [PubMed]
- Kumari, V.; Das, M.; Hodgins, S.; Zachariah, E.; Barkataki, I.; Howlett, M.; Sharma, T. Association between violent behaviour and impaired prepulse inhibition of the startle response in antisocial personality disorder and schizophrenia. Behav. Brain Res. 2005, 158, 159–166. [Google Scholar] [CrossRef] [PubMed]
- Hazlett, E.A.; Speiser, L.J.; Goodman, M.; Roy, M.; Carrizal, M.; Wynn, J.K.; Williams, W.C.; Romero, M.; Minzenberg, M.J.; Siever, L.J.; et al. Exaggerated Affect-Modulated Startle During Unpleasant Stimuli in Borderline Personality Disorder. Biol. Psychiatry 2007, 62, 250–255. [Google Scholar] [CrossRef] [PubMed]
- Walla, P.; Rosser, L.; Scharfenberger, J.; Duregger, C.; Bosshard, S. Emotion Ownership: Different Effects on Explicit Ratings and Implicit Responses. Psychology 2013, 4, 213–216. [Google Scholar] [CrossRef] [Green Version]
- Buss, A.H.; Durkee, A. An inventory for assessing different kinds of hostility. J. Consult. Psychol. 1957, 21, 343–349. [Google Scholar] [CrossRef]
- LimeSurvey Partner Services. Available online: www.limesurvey.com (accessed on 1 June 2019).
- Snyder, M.; Gangestad, S. On the nature of self-monitoring: Matters of assessment, matters of validity. J. Pers. Soc. Psychol. 1986, 51, 125–139. [Google Scholar] [CrossRef]
- Lang, P.; Bradley, M.; Cuthbert, B. International Affective Picture System (IAPS): Affective Ratings of Pictures and Instruction Manual; Technical Report A-8; University of Florida: Gainesville, FL, USA, 2008. [Google Scholar]
- Bartholow, B.D.; Bushman, B.J.; Sestir, M.A. Chronic violent video game exposure and desensitization to violence: Behavioral and event-related brain potential data. J. Exp. Soc. Psychol. 2006, 42, 532–539. [Google Scholar] [CrossRef]
- Kunaharan, S.; Halpin, S.; Sitharthan, T.; Bosshard, S.; Walla, P. Conscious and Non-Conscious Measures of Emotion: Do They Vary with Frequency of Pornography Use? Appl. Sci. 2017, 7, 493. [Google Scholar] [CrossRef]
- Zheng, Y.; Li, J.H. Attentional Bias in Individuals with Different Level of Implicit/Explicit Aggression: Behavioral and ERP Evidence. J. Psychol. Sci. 2014, 37, 40–47. [Google Scholar]
- Wang, Y.; Zhao, Y.; Qiu, J.; Ybarra, O.; Liu, L.; Huang, Y. Neural Correlates of Aggression among Individuals from Low and High Socioeconomic Status: An ERP Study. Int. J. Psychol. Stud. 2012, 4, 37. [Google Scholar] [CrossRef]
- Surguy, S.M.; Bond, A.J. P300 to emotionally relevant stimuli as an indicator of aggression levels. Aggress. Behav. 2006, 32, 253–260. [Google Scholar] [CrossRef]
- Meffert, H.; Gazzola, V.; Boer, J.A.D.; Bartels, A.A.J.; Keysers, C. Reduced spontaneous but relatively normal deliberate vicarious representations in psychopathy. Brain 2013, 136, 2550–2562. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Krumpal, I. Determinants of social desirability bias in sensitive surveys: A literature review. Qual. Quant. 2013, 47, 2025–2047. [Google Scholar] [CrossRef]
- Bailey, K.; West, R.; Anderson, C.A. The association between chronic exposure to video game violence and affective picture processing: An ERP study. Cogn. Affect. Behav. Neurosci. 2011, 11, 259–276. [Google Scholar] [CrossRef] [PubMed]
- Lu, H.; Wang, Y.; Xu, S.; Wang, Y.; Zhang, R.; Li, T. Aggression differentially modulates brain responses to fearful and angry faces: An exploratory study. Neuroreport 2015, 26, 663–668. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Variable | N | (%) |
|---|---|---|
| Marital Status | ||
| Married | 1 | 1.9 |
| Never married | 39 | 75 |
| DeFacto/living with a partner | 12 | 23.1 |
| Highest Level of Completed Education | ||
| Secondary school completed | 22 | 42.3 |
| Trade qualification | 1 | 1.9 |
| University or another tertiary study | 29 | 55.8 |
| Employment | ||
| Part time | 7 | 13.5 |
| Student | 45 | 86.5 |
| Country of Birth | ||
| Australia | 47 | 90.4 |
| Other | 5 | 9.6 |
| Ethnicity * | ||
| African | 1 | 1.9 |
| Asian | 2 | 3.8 |
| Caucasian | 47 | 90.4 |
| Other | 1 | 1.9 |
| Low BDHI Scorers (N = 26) | High BDHI Scorers (N = 26) | Total Sample (N = 52) | ||||
|---|---|---|---|---|---|---|
| Mean | SD | Mean | SD | Mean | SD | |
| Age | 21.19 | 3.48 | 21.38 | 2.68 | 21.00 | 3.1 |
| BDHI total | 20.77 | 5.98 | 38.81 | 8.94 | 29.79 | 11.81 |
| Snyder total | 10.08 | 3.57 | 11.58 | 3.38 | 10.82 | 3.52 |
| BIS-11 total | 62.35 * | 8.12 | 70.12 * | 11.81 | 66.23 | 10.77 |
| Electrode Sites | Time (ms) | p-Value |
|---|---|---|
| 94 | 0.05 | |
| 484 | 0.045 | |
| 641 | 0.011 | |
| AF3/AF4 | 719 | 0.008 |
| 797 | 0.007 | |
| 875 | 0.014 | |
| 953 | 0.017 | |
| 1031 | 0.012 | |
| 484 | 0.026 | |
| C3/C4 | 875 | 0.036 |
| 953 | 0.04 | |
| 1031 | 0.017 | |
| 406 | 0.044 | |
| 484 | 0.015 | |
| P3/P4 | 563 | 0.017 |
| 641 | 0.034 | |
| 953 | 0.049 | |
| 1031 | 0.015 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kunaharan, S.; Halpin, S.; Sitharthan, T.; Walla, P. Do EEG and Startle Reflex Modulation Vary with Self-Reported Aggression in Response to Violent Images? Brain Sci. 2019, 9, 298. https://doi.org/10.3390/brainsci9110298
Kunaharan S, Halpin S, Sitharthan T, Walla P. Do EEG and Startle Reflex Modulation Vary with Self-Reported Aggression in Response to Violent Images? Brain Sciences. 2019; 9(11):298. https://doi.org/10.3390/brainsci9110298
Chicago/Turabian StyleKunaharan, Sajeev, Sean Halpin, Thiagarajan Sitharthan, and Peter Walla. 2019. "Do EEG and Startle Reflex Modulation Vary with Self-Reported Aggression in Response to Violent Images?" Brain Sciences 9, no. 11: 298. https://doi.org/10.3390/brainsci9110298
APA StyleKunaharan, S., Halpin, S., Sitharthan, T., & Walla, P. (2019). Do EEG and Startle Reflex Modulation Vary with Self-Reported Aggression in Response to Violent Images? Brain Sciences, 9(11), 298. https://doi.org/10.3390/brainsci9110298