Mechanisms for Auditory Perception: A Neurocognitive Study of Second Language Learning of Mandarin Chinese

{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Methods
2.1. Participants
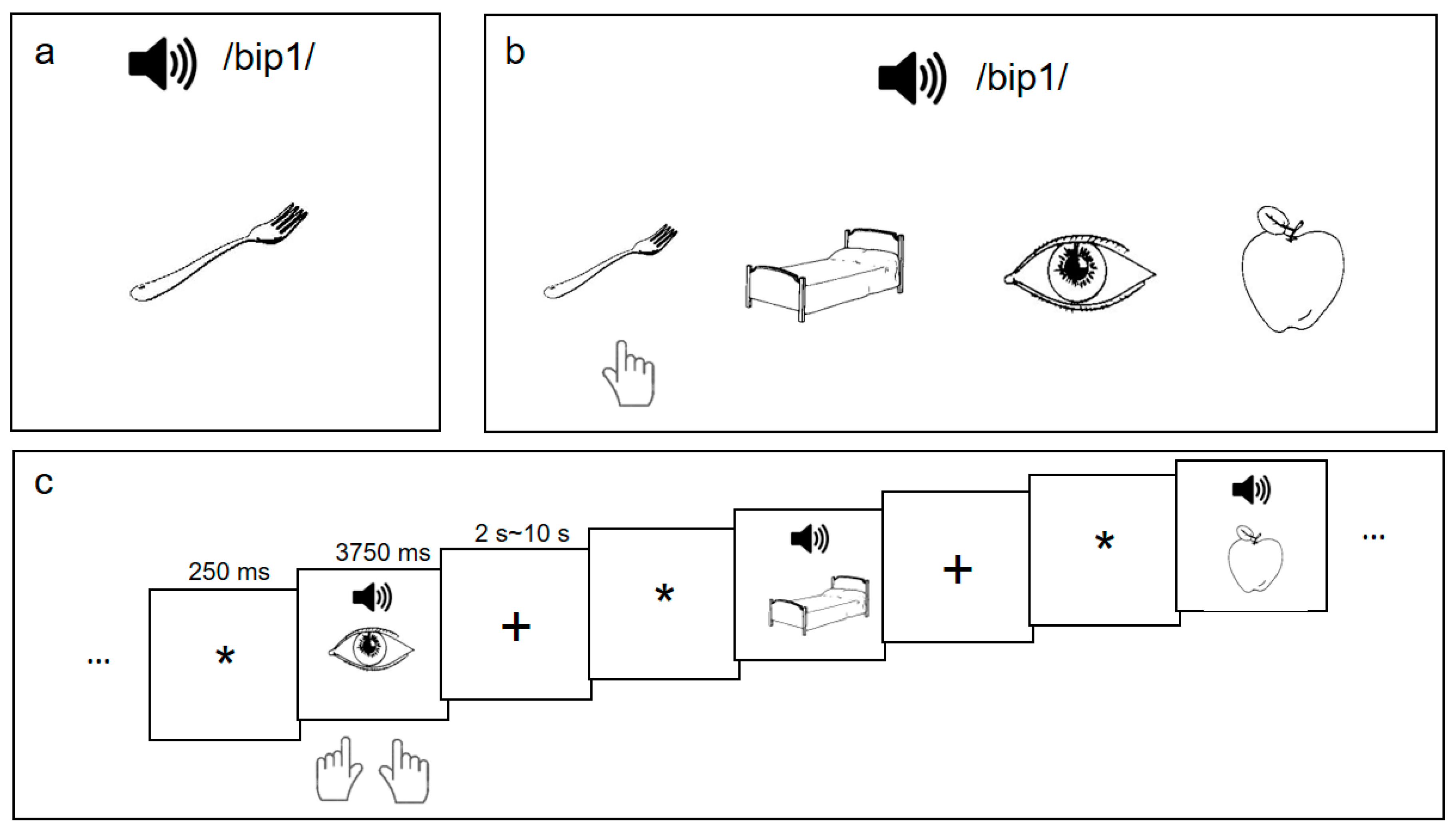
2.2. Materials and Procedure
2.3. MRI Acquisition
2.4. fMRI Data Analysis
3. Results
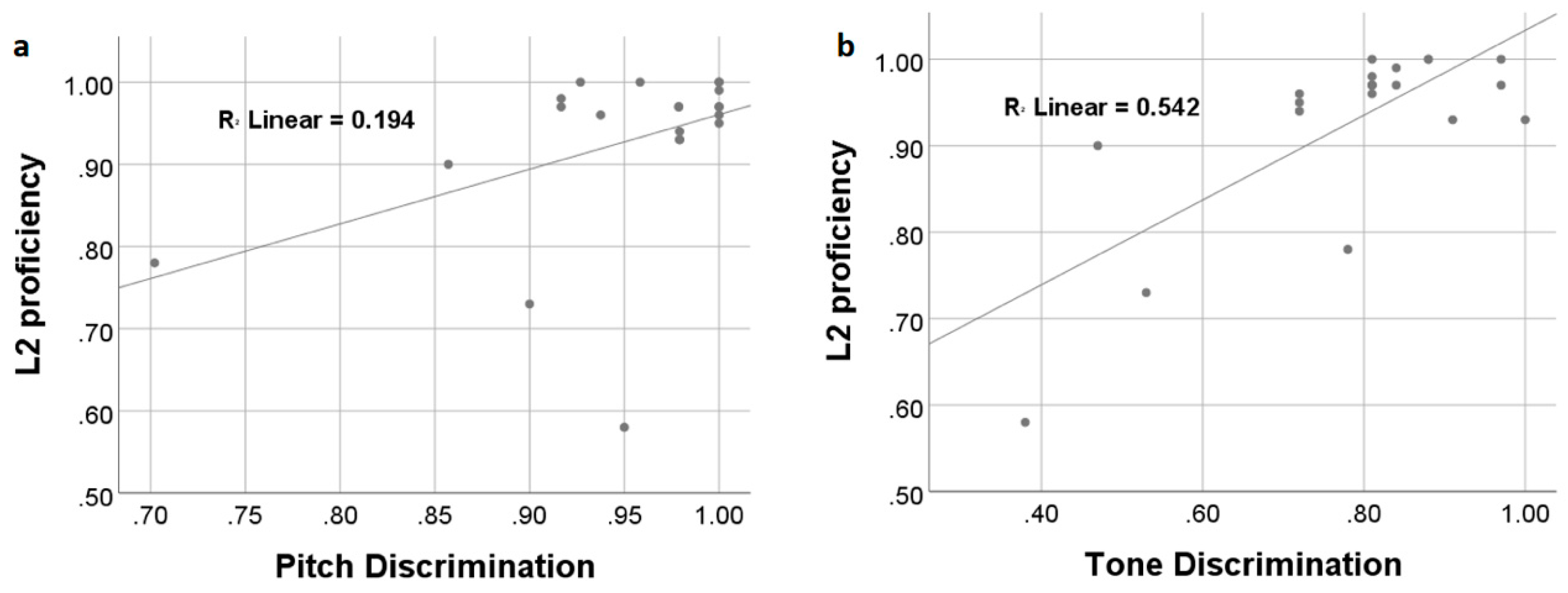
3.1. Behavioral Results
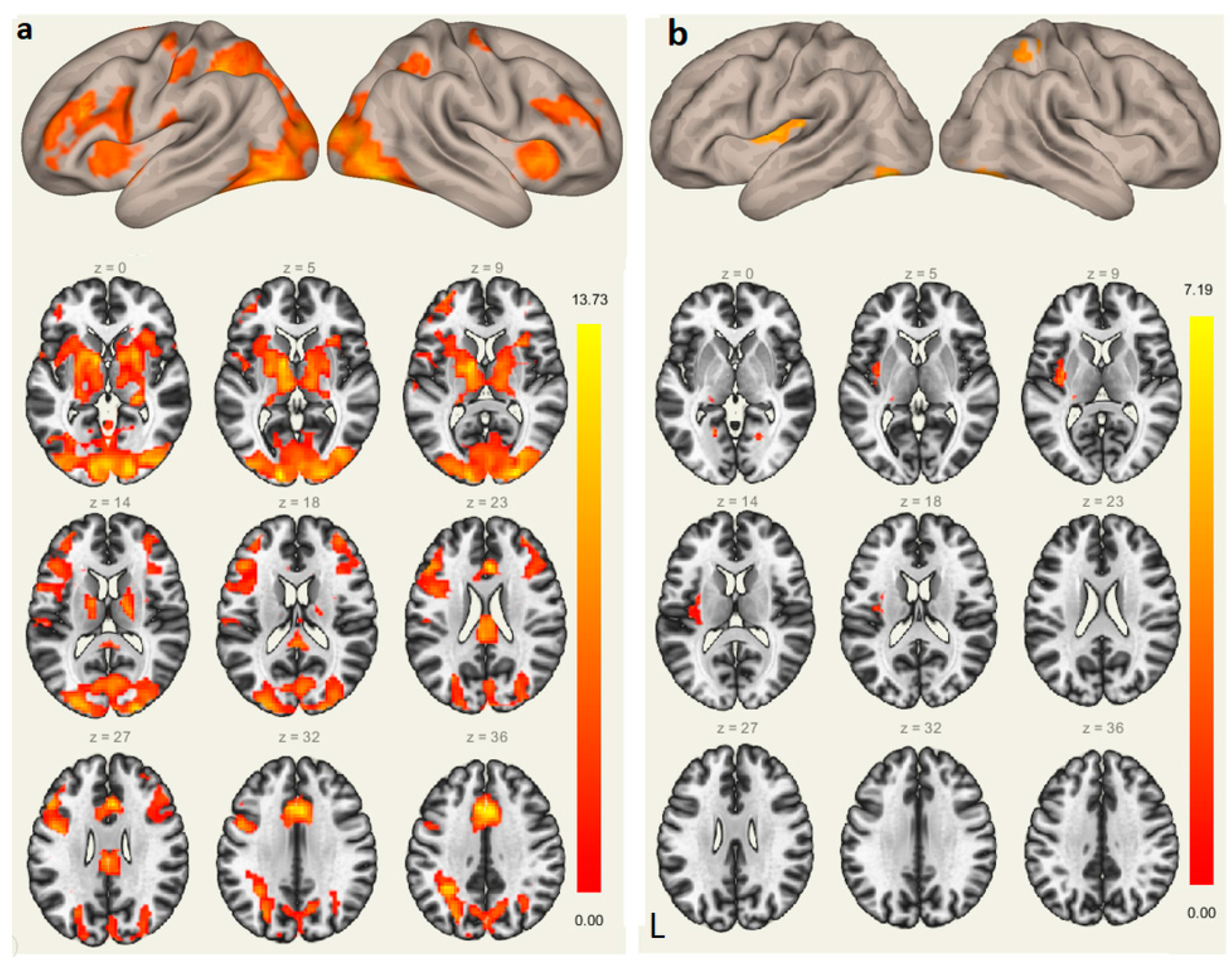
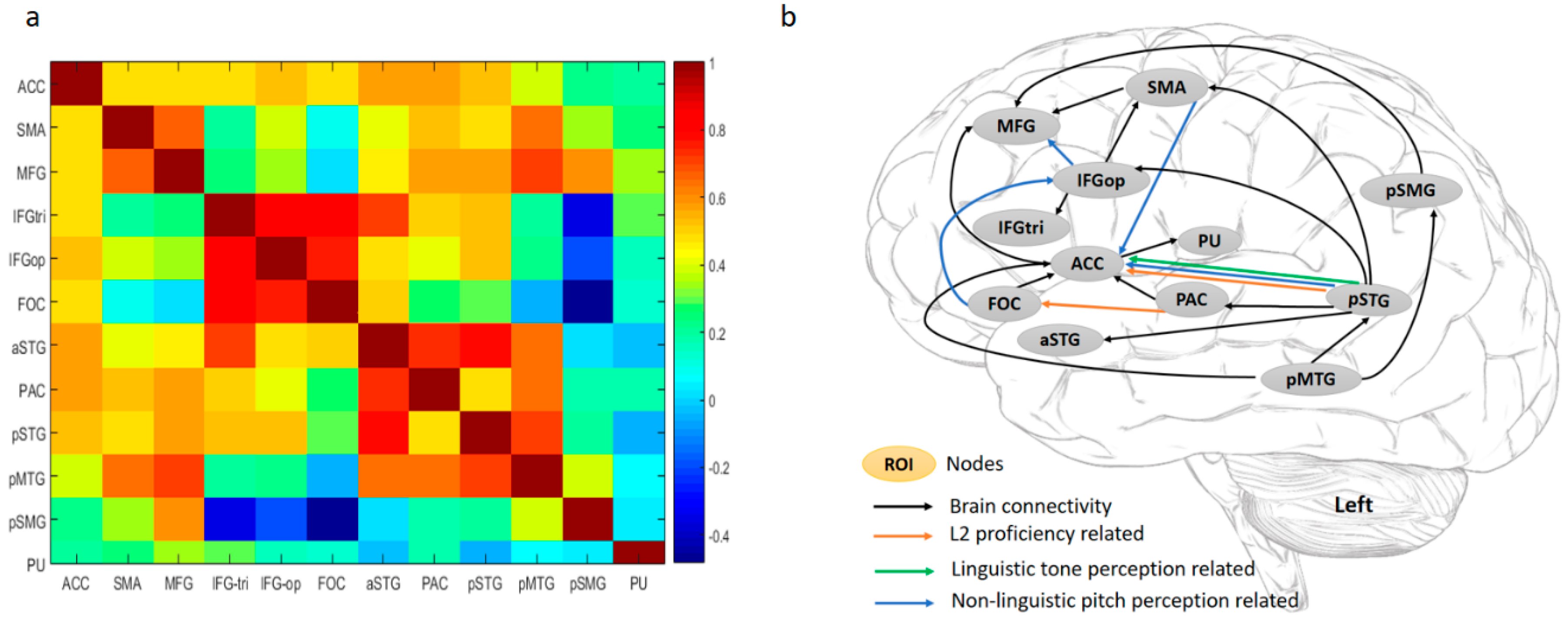
3.2. fMRI Results
4. Discussion
4.1. L2 Word Learning Depends on Auditory Perception Abilities in the Processing of Auditory Decoding, Articulatory Rehearsal, and Phonological Storage
4.2. L2 Word Learning Success Lies in the Collaboration of Dorsal and Ventral Streams of the L2 Language Network
5. Conclusions and Limitations
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Chandrasekaran, B.; Sampath, P.D.; Wong, P.C. Individual variability in cue-weighting and lexical tone learning. J. Acoust. Soc. Am. 2010, 128, 456–465. [Google Scholar] [CrossRef] [PubMed]
- Cumming, R.; Wilson, A.; Goswami, U. Basic auditory processing and sensitivity to prosodic structure in children with specific language impairments: A new look at a perceptual hypothesis. Front. Psychol. 2015, 6, 972. [Google Scholar] [CrossRef] [PubMed]
- Kuhl, P.K. Early Language Learning and Literacy: Neuroscience Implications for Education. Mind Brain Educ. 2011, 5, 128–142. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Silva-Pereyra, J.; Kuhl, P.K.; Rivera-Gaxiola, M.; Silva-Pereyra, J.; Rivera-Gaxiola, M.; Silva-Pereyra, J. Brain potentials to native and non-native speech contrasts in 7- and 11-month-old American infants. Dev. Sci. 2005, 8, 162–172. [Google Scholar]
- Wong, P.C.M.; Perrachione, T.K. Learning pitch patterns in lexical identification by native English-speaking adults. Appl. Psycholinguist. 2007, 28, 565–585. [Google Scholar] [CrossRef] [Green Version]
- Li, P.; Legault, J.; Litcofsky, K.A. Neuroplasticity as a function of second language learning: Anatomical changes in the human brain. Cortex 2014, 58, 301–324. [Google Scholar] [CrossRef] [PubMed]
- Golestani, N.; Molko, N.; Dehaene, S.; LeBihan, D.; Pallier, C. Brain Structure Predicts the Learning of Foreign Speech Sounds. Cereb. Cortex 2006, 17, 575–582. [Google Scholar] [CrossRef] [Green Version]
- Golestani, N.; Pallier, C. Anatomical Correlates of Foreign Speech Sound Production. Cereb. Cortex 2006, 17, 929–934. [Google Scholar] [CrossRef]
- Golestani, N.; Zatorre, R.J. Learning new sounds of speech: Reallocation of neural substrates. NeuroImage 2004, 21, 494–506. [Google Scholar] [CrossRef]
- Ventura-Campos, N.; Sanjuán, A.; González, J.; Palomar-García, M.-Á.; Rodríguez-Pujadas, A.; Sebastián-Gallés, N.; Deco, G.; Ávila, C. Spontaneous brain activity predicts learning ability of foreign sounds. J. Neurosci. 2013, 33, 9295–9305. [Google Scholar] [CrossRef]
- Sereno, J.A.; Wang, Y.; Jongman, A. Acoustic and perceptual evaluation of Mandarin tone productions before and after perceptual training. J. Acoust. Soc. Am. 2003, 113, 1033–1043. [Google Scholar]
- Mei, L.; Xue, G.; Lu, Z.-L.; Chen, C.; Zhang, M.; He, Q.; Wei, M.; Dong, Q. Learning to Read Words in a New Language Shapes the Neural Organization of the Prior Languages. Neuropsychologia 2014, 65, 156–168. [Google Scholar] [CrossRef] [PubMed]
- Raboyeau, G.; Marcotte, K.; Adrover-Roig, D.; Ansaldo, A. Brain activation and lexical learning: The impact of learning phase and word type. NeuroImage 2010, 49, 2850–2861. [Google Scholar] [CrossRef] [PubMed]
- Veroude, K.; Norris, D.G.; Shumskaya, E.; Gullberg, M.; Indefrey, P. Functional connectivity between brain regions involved in learning words of a new language. Brain Lang. 2010, 113, 21–27. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wong, P.C.; Perrachione, T.K.; Parrish, T.B. Neural characteristics of successful and less successful speech and word learning in adults. Hum. Brain Mapp. 2007, 28, 995–1006. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.; Gates, K.M.; Molenaar, P.; Li, P. Neural changes underlying successful second language word learning: An fMRI study. J. Neurolinguist. 2015, 33, 29–49. [Google Scholar] [CrossRef] [Green Version]
- Legault, J.; Fang, S.-Y.; Lan, Y.-J.; Li, P. Structural Brain Changes as a Function of Second Language Vocabulary Training: Effects of Learning Context. Brain Cogn. 2018, 11. [Google Scholar] [CrossRef] [PubMed]
- Prat, C.S.; Yamasaki, B.L.; Kluender, R.A.; Stocco, A. Resting-state qEEG predicts rate of second language learning in adults. Brain Lang. 2016, 157, 44–50. [Google Scholar] [CrossRef]
- Grant, A.M.; Fang, S.-Y.; Li, P. Second language lexical development and cognitive control: A longitudinal fMRI study. Brain Lang. 2015, 144, 35–47. [Google Scholar] [CrossRef]
- Stein, M.; Federspiel, A.; Koenig, T.; Wirth, M.; Strik, W.; Wiest, R.; Brandeis, D.; Dierks, T. Structural plasticity in the language system related to increased second language proficiency. Cortex 2012, 48, 458–465. [Google Scholar] [CrossRef]
- Mårtensson, J.; Eriksson, J.; Bodammer, N.C.; Lindgren, M.; Johansson, M.; Nyberg, L.; Lövdén, M. Growth of language-related brain areas after foreign language learning. NeuroImage 2012, 63, 240–244. [Google Scholar] [CrossRef] [PubMed]
- Xiang, H.; Dediu, D.; Roberts, L.; Van Oort, E.; Norris, D.G.; Hagoort, P. The Structural Connectivity Underpinning Language Aptitude, Working Memory, and IQ in the Perisylvian Language Network. Lang. Learn. 2012, 62, 110–130. [Google Scholar] [CrossRef] [Green Version]
- Qi, Z.; Han, M.; Garel, K.-L.; Chen, E.S.; Gabrieli, J.D. White-matter structure in the right hemisphere predicts Mandarin Chinese learning success. J. Neurolinguist. 2015, 33, 14–28. [Google Scholar] [CrossRef] [Green Version]
- McLaughlin, J.; Osterhout, L.; Kim, A. Neural correlates of second-language word learning: Minimal instruction produces rapid change. Nat. Neurosci. 2004, 7, 703–704. [Google Scholar] [CrossRef] [PubMed]
- Breitenstein, C.; Jansen, A.; Deppe, M.; Foerster, A.-F.; Sommer, J.; Wolbers, T.; Knecht, S. Hippocampus activity differentiates good from poor learners of a novel lexicon. NeuroImage 2005, 25, 958–968. [Google Scholar] [CrossRef] [PubMed]
- Mei, L.; Chen, C.; Xue, G.; He, Q.; Li, T.; Xue, F.; Yang, Q.; Dong, Q. Neural predictors of auditory word learning. NeuroReport 2008, 19, 215–219. [Google Scholar] [CrossRef] [Green Version]
- Chai, X.J.; Berken, J.A.; Barbeau, E.B.; Soles, J.; Callahan, M.; Chen, J.-K.; Klein, D. Intrinsic Functional Connectivity in the Adult Brain and Success in Second-Language Learning. J. Neurosci. 2016, 36, 755–761. [Google Scholar] [CrossRef] [PubMed]
- Qi, Z.; Han, M.; Wang, Y.; Angeles, C.D.L.; Liu, Q.; Garel, K.; Chen, E.S.; Whitfield-Gabrieli, S.; Gabrieli, J.D.; Perrachione, T.K. Speech processing and plasticity in the right hemisphere predict variation in adult foreign language learning. NeuroImage 2019, 192, 76–87. [Google Scholar] [CrossRef]
- Yang, J.; Li, P. Brain Networks of Explicit and Implicit Learning. PLoS ONE 2012, 7, e42993. [Google Scholar] [CrossRef]
- Sheppard, J.P.; Wang, J.-P.; Wong, P.C.M. Large-scale Cortical Network Properties Predict Future Sound-to-Word Learning Success. J. Cogn. Neurosci. 2012, 24, 1087–1103. [Google Scholar] [CrossRef] [Green Version]
- Majerus, S.; Belayachi, S.; De Smedt, B.; Leclercq, A.; Martinez, T.; Schmidt, C.; Weekes, B.; Maquet, P. Neural networks for short-term memory for order differentiate high and low proficiency bilinguals. NeuroImage 2008, 42, 1698–1713. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Golestani, N.; Zatorre, R.J. Individual differences in the acquisition of second language phonology. Brain Lang. 2009, 109, 55–67. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hu, X.; Ackermann, H.; Martin, J.A.; Erb, M.; Winkler, S.; Reiterer, S.M. Language aptitude for pronunciation in advanced second language (L2) Learners: Behavioural predictors and neural substrates. Brain Lang. 2013, 127, 366–376. [Google Scholar] [CrossRef] [PubMed]
- Golestani, N.; Moser-Mercer, B.; Hervais-Adelman, A.; Murray, M.; Frauenfelder, U.; Meuli, R.; Hageman, P.; Scott, S.; Michel, C. Experience-dependent brain structural plasticity in simultaneous language interpreters. In Proceedings of the Cognitive Neuroscience Society Meeting, Montreal, QC, Canada, 6–10 June 2010. [Google Scholar]
- Friederici, A.D. Pathways to language: fiber tracts in the human brain. Trends Cogn. Sci. 2009, 13, 175–181. [Google Scholar] [CrossRef] [PubMed]
- Friederici, A.D.; Gierhan, S.M. The language network. Curr. Opin. Neurobiol. 2013, 23, 250–254. [Google Scholar] [CrossRef] [PubMed]
- Hickok, G. The Functional Neuroanatomy of Language. Phys. Life Rev. 2009, 6, 121–143. [Google Scholar] [CrossRef] [PubMed]
- Hagoort, P.; Indefrey, P. The Neurobiology of Language Beyond Single Words. Annu. Rev. Neurosci. 2014, 37, 347–362. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Green, D.W.; Abutalebi, J. Language control in bilinguals: The adaptive control hypothesis. J. Cogn. Psychol. 2013, 25, 515–530. [Google Scholar] [CrossRef] [PubMed]
- Stocco, A.; Yamasaki, B.; Natalenko, R.; Prat, C.S. Bilingual brain training: A neurobiological framework of how bilingual experience improves executive function. Int. J. Bilingual. 2014, 18, 67–92. [Google Scholar] [CrossRef]
- Golestani, N. Neuroimaging of phonetic perception in bilinguals. Bilingual. Lang. Cogn. 2016, 19, 674–682. [Google Scholar] [CrossRef]
- Grant, A.; Legault, J.; Li, P. What do bilingual models tell us about the neurocognition of multiple languages? In The Handbook of the Neuroscience of Multilingualism; John Wiley & Sons: Hoboken, NJ, USA, 2019; pp. 48–74. [Google Scholar]
- Snyder, P.J.; Harris, L.J. Handedness, Sex, Familial Sinistrality Effects on Spatial Tasks. Cortex 1993, 29, 115–134. [Google Scholar] [CrossRef]
- Li, P.; Zhang, F.; Tsai, E.; Puls, B. Language history questionnaire (LHQ 2.0): A new dynamic web-based research tool. Bilingual. Lang. Cogn. 2014, 17, 673–680. [Google Scholar] [CrossRef]
- Li, P.; Zhang, F.; Yu, A.; Zhao, X. Language History Questionnaire (LHQ3): An enhanced tool for assessing multilingual experience. Biling. Lang. Cogn. 2019, 1–7. [Google Scholar] [CrossRef]
- Lee, Y.-S.; Vakoch, D.A.; Wurm, L.H. Tone perception in Cantonese and Mandarin: A cross-linguistic comparison. J. Psycholinguist. Res. 1996, 25, 527–542. [Google Scholar] [CrossRef]
- Klein, D.; Zatorre, R.J.; Milner, B.; Zhao, V. A Cross-Linguistic PET Study of Tone Perception in Mandarin Chinese and English Speakers. NeuroImage 2001, 13, 646–653. [Google Scholar] [CrossRef] [Green Version]
- Hao, Y.-C. Second language acquisition of Mandarin Chinese tones by tonal and non-tonal language speakers. J. Phon. 2012, 40, 269–279. [Google Scholar] [CrossRef]
- Zhang, L.; Shu, H.; Zhou, F.; Wang, X.; Li, P. Common and distinct neural substrates for the perception of speech rhythm and intonation. Hum. Brain Mapp. 2010, 31, 1106–1116. [Google Scholar] [CrossRef]
- Bates, E.; D’Amico, S.; Jacobsen, T.; Szekely, A.; Andonova, E.; Devescovi, A.; Herron, D.; Lu, C.C.; Pechmann, T.; Pléh, C.; et al. Timed picture naming in seven languages. Psychon. Bull. Rev. 2003, 10, 344–380. [Google Scholar] [CrossRef] [Green Version]
- Liu, Y.; Hao, M.; Li, P.; Shu, H. Timed Picture Naming Norms for Mandarin Chinese. PLoS ONE 2011, 6, e16505. [Google Scholar] [CrossRef]
- Penny, W.D.; Friston, K.J.; Ashburner, J.T.; Kiebel, S.J.; Nichols, T.E. Statistical Parametric Mapping: The Analysis of Functional Brain Images; Elsevier: Amsterdam, The Netherlands, 2011. [Google Scholar]
- Yan, C.-G.; Wang, X.-D.; Zuo, X.-N.; Zang, Y.-F. DPABI: Data Processing & Analysis for (Resting-State) Brain Imaging. Neuroinformatics 2016, 14, 339–351. [Google Scholar]
- Makris, N.; Goldstein, J.M.; Kennedy, D.; Hodge, S.M.; Caviness, V.S.; Faraone, S.V.; Tsuang, M.T.; Seidman, L.J. Decreased volume of left and total anterior insular lobule in schizophrenia. Schizophr. Res. 2006, 83, 155–171. [Google Scholar] [CrossRef]
- Gates, K.M.; Molenaar, P.C. Group search algorithm recovers effective connectivity maps for individuals in homogeneous and heterogeneous samples. NeuroImage 2012, 63, 310–319. [Google Scholar] [CrossRef]
- Lane, S.T.; Gates, K.M. Automated Selection of Robust Individual-Level Structural Equation Models for Time Series Data. Struct. Equ. Model. A Multidiscip. J. 2017, 24, 768–782. [Google Scholar] [CrossRef]
- Lane, S.; Gates, K.; Molenaar, P.; Hallquist, M.; Pike, H. Gimme: Group Iterative Multiple Model Estimation. (R Package Version 0.1-7). Available online: https://cran.r-project.org/web/packages/gimme/vignettes/gimme_vignette.html (accessed on 2 January 2019).
- Bates, D.; Mächler, M.; Bolker, B.; Walker, S. Fitting Linear Mixed-Effects Models Using lme4. Available online: https://arxiv.org/abs/1406.5823 (accessed on 2 January 2019).
- Kim, J.; Zhu, W.; Chang, L.; Bentler, P.M.; Ernst, T. Unified structural equation modeling approach for the analysis of multisubject, multivariate functional MRI data. Hum. Brain Mapp. 2007, 28, 85–93. [Google Scholar] [CrossRef]
- Gates, K.M.; Molenaar, P.C.; Hillary, F.G.; Ram, N.; Rovine, M.J. Automatic search for fMRI connectivity mapping: An alternative to Granger causality testing using formal equivalences among SEM path modeling, VAR, and unified SEM. NeuroImage 2010, 50, 1118–1125. [Google Scholar] [CrossRef]
- Wong, P.C.; Warrier, C.M.; Penhune, V.B.; Roy, A.K.; Sadehh, A.; Parrish, T.B.; Zatorre, R.J. Volume of left Heschl’s gyrus and linguistic pitch learning. Cereb. Cortex 2007, 18, 828–836. [Google Scholar] [CrossRef]
- Graves, W.W.; Grabowski, T.J.; Mehta, S.; Gupta, P. The left posterior superior temporal gyrus participates specifically in accessing lexical phonology. J. Cogn. Neurosci. 2008, 20, 1698–1710. [Google Scholar] [CrossRef]
- Chang, E.F.; Rieger, J.W.; Johnson, K.; Berger, M.S.; Barbaro, N.M.; Knight, R.T. Categorical speech representation in human superior temporal gyrus. Nat. Neurosci. 2010, 13, 1428–1432. [Google Scholar] [CrossRef]
- Kimppa, L.; Kujala, T.; Leminen, A.; Vainio, M.; Shtyrov, Y. Rapid and automatic speech-specific learning mechanism in human neocortex. NeuroImage 2015, 118, 282–291. [Google Scholar] [CrossRef] [Green Version]
- Whitney, C.; Kirk, M.; O’Sullivan, J.; Ralph, M.A.L.; Jefferies, E. The Neural Organization of Semantic Control: TMS Evidence for a Distributed Network in Left Inferior Frontal and Posterior Middle Temporal Gyrus. Cereb. Cortex 2010, 21, 1066–1075. [Google Scholar] [CrossRef] [Green Version]
- Davis, M.H.; Di Betta, A.M.; Macdonald, M.J.E.; Gaskell, M.G. Learning and Consolidation of Novel Spoken Words. J. Cogn. Neurosci. 2009, 21, 803–820. [Google Scholar] [CrossRef] [Green Version]
- Iacoboni, M. The role of premotor cortex in speech perception: Evidence from fMRI and rTMS. J. Physiol. 2008, 102, 31–34. [Google Scholar] [CrossRef]
- Friederici, A.D.; Meyer, M.; Von Cramon, D. Auditory Language Comprehension: An Event-Related fMRI Study on the Processing of Syntactic and Lexical Information. Brain Lang. 2000, 74, 289–300. [Google Scholar] [CrossRef] [Green Version]
- Mechelli, A.; Crinion, J.T.; Noppeney, U.; O’Doherty, J.; Ashburner, J.; Frackowiak, R.S.; Price, C.J. Structural plasticity in the bilingual brain: proficiency in a second language and age at acquisition affect grey-matter density. Nature 2004, 431, 757. [Google Scholar] [CrossRef]
- Della Rosa, P.A.; Videsott, G.; Borsa, V.M.; Canini, M.; Weekes, B.S.; Franceschini, R.; Abutalebi, J. A neural interactive location for multilingual talent. Cortex 2013, 49, 605–608. [Google Scholar] [CrossRef]
- Chandrasekaran, B.; Garibaldi, K.; Wong, F.C.K.; Wong, P.C.M. White matter anisotropy in the ventral language pathway predicts sound-to-word learning success. J. Neurosci. 2011, 31, 8780–8785. [Google Scholar]
- Schlegel, A.A.; Rudelson, J.J.; Tse, P.U. White Matter Structure Changes as Adults Learn a Second Language. J. Cogn. Neurosci. 2012, 24, 1664–1670. [Google Scholar] [CrossRef]
- Xiang, H.; Van Leeuwen, T.M.; Dediu, D.; Roberts, L.; Norris, D.G.; Hagoort, P. L2-Proficiency-Dependent Laterality Shift in Structural Connectivity of Brain Language Pathways. Brain Connect. 2015, 5, 349–361. [Google Scholar] [CrossRef]
- Kerns, J.G.; Cohen, J.D.; Macdonald, A.W.; Cho, R.Y.; Stenger, V.A.; Carter, C.S. Anterior Cingulate Conflict Monitoring and Adjustments in Control. Science 2004, 303, 1023–1026. [Google Scholar] [CrossRef] [Green Version]
- Abutalebi, J.; Green, D.W. Neuroimaging of language control in bilinguals: neural adaptation and reserve. Biling. Lang. Cogn. 2016, 19, 689–698. [Google Scholar] [CrossRef] [Green Version]
- Ranganath, C.; Johnson, M.K.; D’Esposito, M. Left Anterior Prefrontal Activation Increases with Demands to Recall Specific Perceptual Information. J. Neurosci. 2000, 20, RC108. [Google Scholar] [CrossRef] [PubMed]
- Saur, D.; Kreher, B.W.; Schnell, S.; Kümmerer, D.; Kellmeyer, P.; Vry, M.-S.; Umarova, R.; Musso, M.; Glauche, V.; Abel, S. Ventral and dorsal pathways for language. Proc. Natl. Acad. Sci. USA 2008, 105, 18035–18040. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schlauch, R.S.; Nelson, P. Pure tone evaluation. In Katz Handbook of Clinical Audiology, 7th ed.; Lipponcott Williams &Wilkins: Baltimore, MD, USA, 2015; pp. 29–47. [Google Scholar]
- Gierhan, S.M. Connections for auditory language in the human brain. Brain Lang. 2013, 127, 205–221. [Google Scholar] [CrossRef] [PubMed]




© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yang, J.; Li, P. Mechanisms for Auditory Perception: A Neurocognitive Study of Second Language Learning of Mandarin Chinese. Brain Sci. 2019, 9, 139. https://doi.org/10.3390/brainsci9060139
Yang J, Li P. Mechanisms for Auditory Perception: A Neurocognitive Study of Second Language Learning of Mandarin Chinese. Brain Sciences. 2019; 9(6):139. https://doi.org/10.3390/brainsci9060139
Chicago/Turabian StyleYang, Jing, and Ping Li. 2019. "Mechanisms for Auditory Perception: A Neurocognitive Study of Second Language Learning of Mandarin Chinese" Brain Sciences 9, no. 6: 139. https://doi.org/10.3390/brainsci9060139
APA StyleYang, J., & Li, P. (2019). Mechanisms for Auditory Perception: A Neurocognitive Study of Second Language Learning of Mandarin Chinese. Brain Sciences, 9(6), 139. https://doi.org/10.3390/brainsci9060139