
Microarray Selection of Cooperative Peptides for Modulating Enzyme Activities

Abstract
:1. Introduction
2. Materials and Approach
2.1. Materials
2.2. Microarray Fabrication
2.3. Formaldehyde Crosslinking
2.4. Microarray Binding
2.5. Enzyme Inhibition Assays
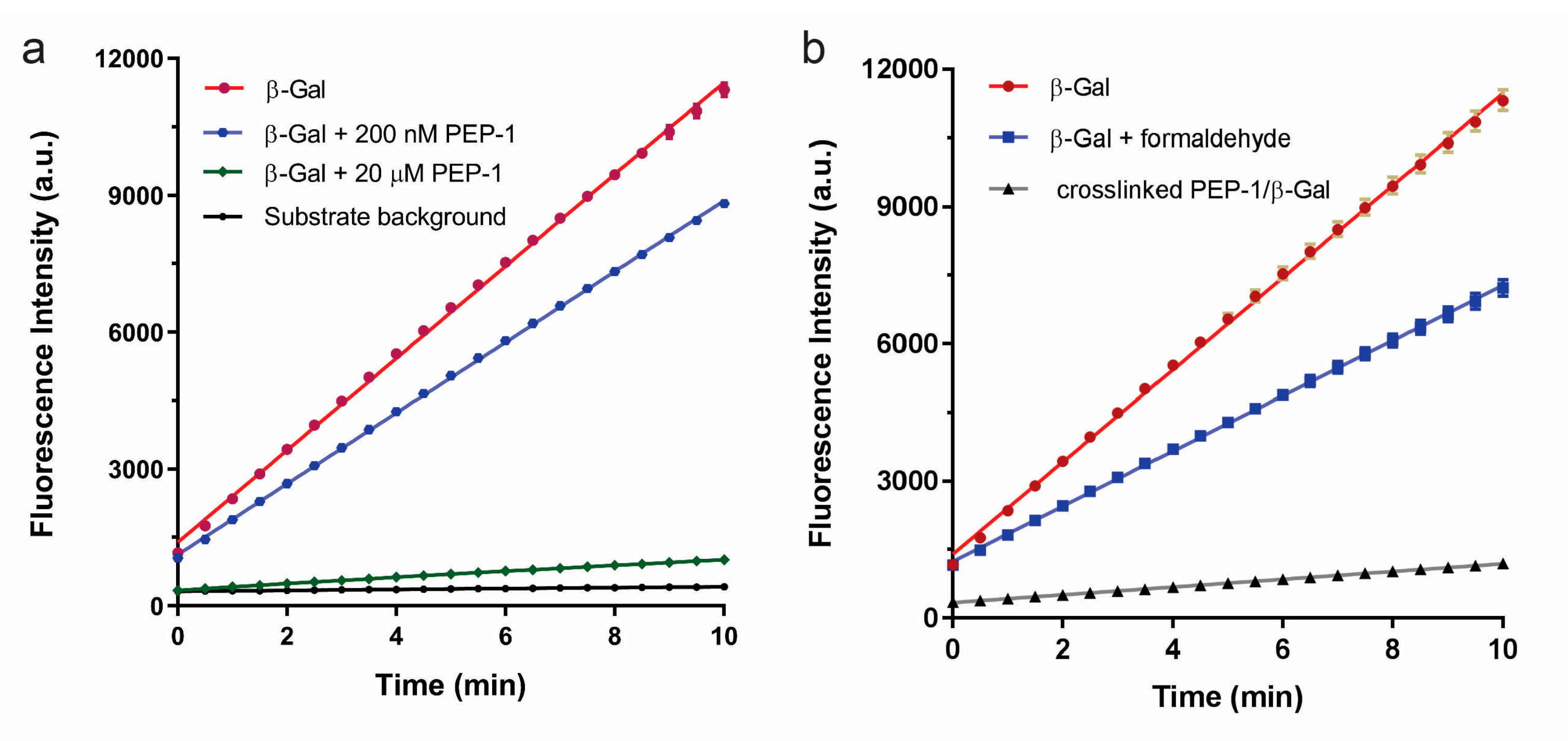
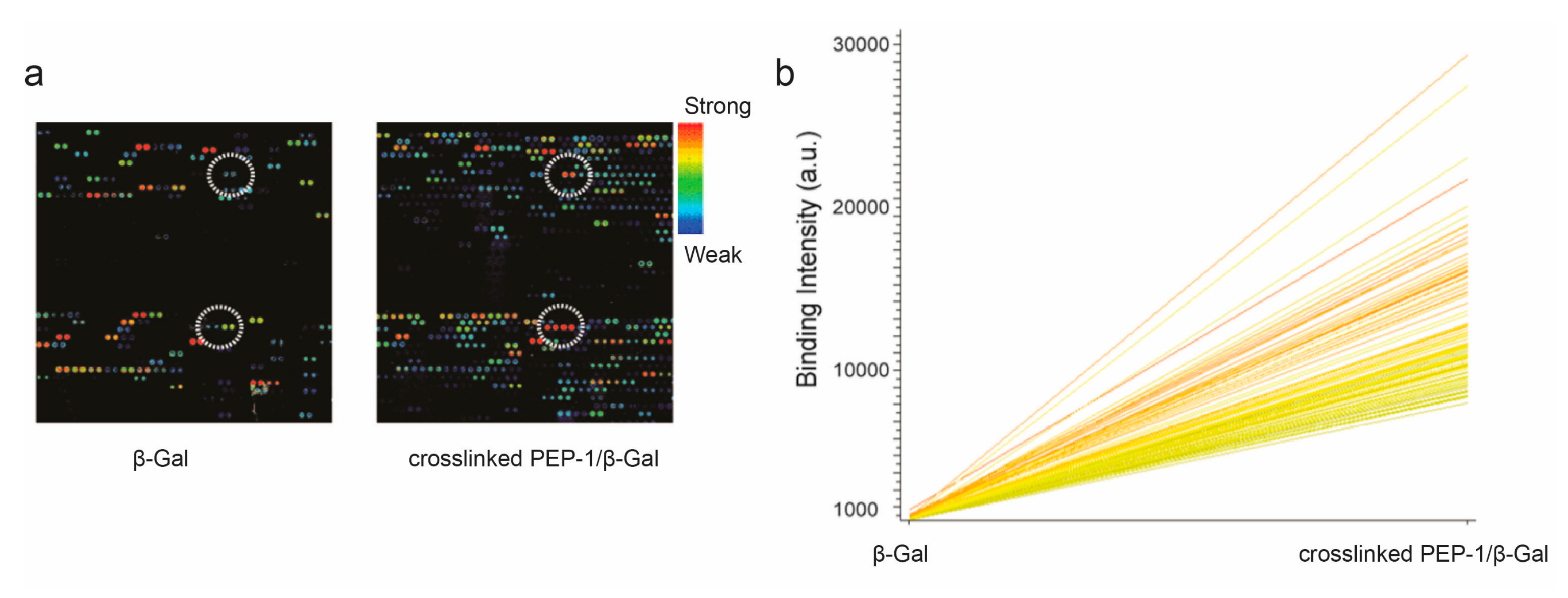
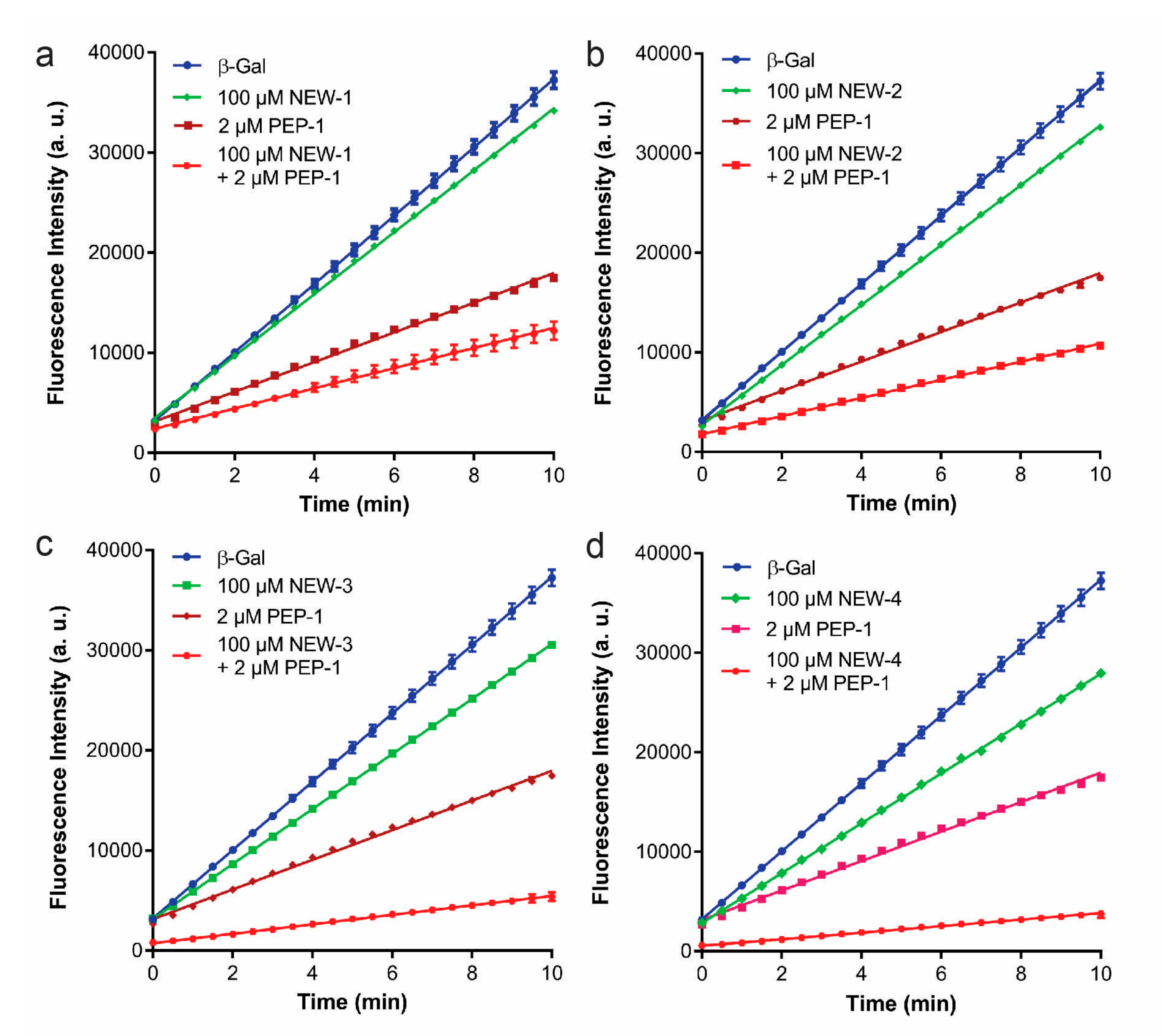
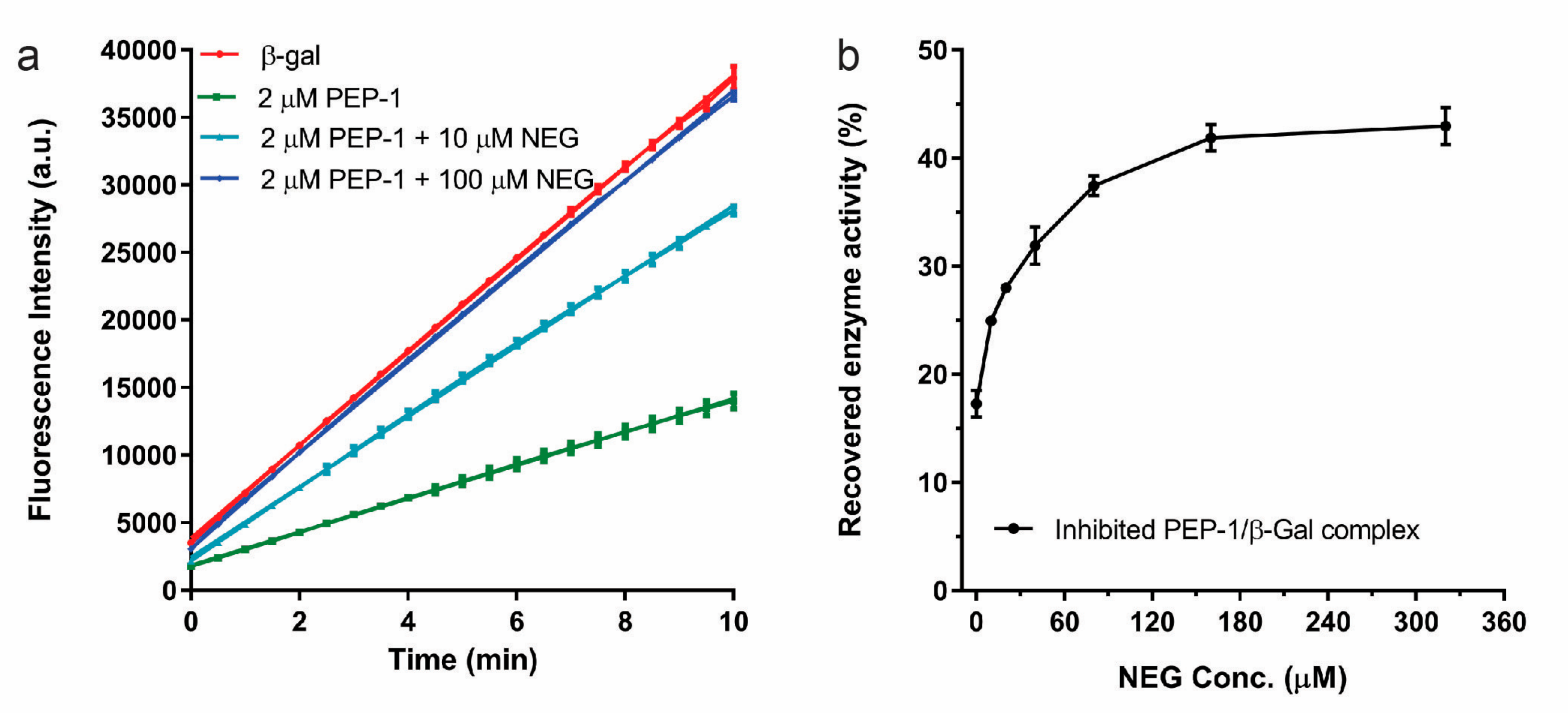
3. Results and Discussion
4. Conclusions
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Drews, J. Drug discovery: A historical perspective. Science 2000, 287, 1960–1964. [Google Scholar] [CrossRef] [PubMed]
- Gordon, G.H.; Wu, C.W. Regulation of enzyme activity. Science 1971, 172, 1205–1211. [Google Scholar]
- Vegas, A.J.; Fuller, J.H.; Koehler, A.N. Small-molecule microarrays as tools in ligand discovery. Chem. Soc. Rev. 2008, 37, 1385–1394. [Google Scholar] [CrossRef] [PubMed]
- Schreiber, S.L. The small-molecule approach to biology. Chem. Eng. News 2003, 81, 51–61. [Google Scholar]
- Troitskaya, L.A.; Kodadek, T. Peptides as modulators of enzymes and regulatory proteins. Methods 2004, 32, 406–415. [Google Scholar] [CrossRef] [PubMed]
- Devlin, J.; Panganiban, L.; Devlin, P. Random peptide libraries: A source of specific protein binding molecules. Science 1990, 249, 404–406. [Google Scholar] [CrossRef] [PubMed]
- Fodor, S.; Read, J.; Pirrung, M.; Stryer, L.; Lu, A.; Solas, D. Light-directed, spatially addressable parallel chemical synthesis. Science 1991, 251, 767–773. [Google Scholar] [CrossRef] [PubMed]
- Hyde-DeRuyscher, R.; Paige, L.A.; Christensen, D.J.; Hyde-DeRuyscher, N.; Lim, A.; Fredericks, Z.L.; Kranz, J.; Gallant, P.; Zhang, J.; Rocklage, S.M.; et al. Detection of small-molecule enzyme inhibitors with peptides isolated from phage-displayed combinatorial peptide libraries. Chem. Biol. 2000, 7, 17–25. [Google Scholar] [CrossRef]
- Yagi, Y.; Terada, K.; Noma, T.; Ikebukuro, K.; Sode, K. In silico panning for a non-competitive peptide inhibitor. BMC Bioinform. 2007, 8, 11. [Google Scholar] [CrossRef] [PubMed]
- Bogoyevitch, M.A.; Barr, R.K.; Ketterman, A.J. Peptide inhibitors of protein kinases—discovery, characterisation and use. Biochim. Biophys. Acta Proteins Proteom. 2005, 1754, 79–99. [Google Scholar] [CrossRef] [PubMed]
- Walker, B.; McCarthy, N.; Healy, A.; Ye, T.; McKervey, M.A. Peptide glyoxals: A novel class of inhibitor for serine and cysteine proteinases. Biochem. J. 1993, 293, 321–323. [Google Scholar] [CrossRef] [PubMed]
- Kaneto, H.; Nakatani, Y.; Miyatsuka, T.; Kawamori, D.; Matsuoka, T.; Matsuhisa, M.; Kajimoto, Y.; Ichijo, H.; Yamasaki, Y.; Hori, M. Possible novel therapy for diabetes with cell-permeable JNK-inhibitory peptide. Nat. Methods 2004, 10, 1128–1132. [Google Scholar] [CrossRef] [PubMed]
- Orange, J.; May, M. Cell penetrating peptide inhibitors of nuclear factor-kappa B. Cell. Mol. Life Sci. 2008, 65, 3564–3591. [Google Scholar] [CrossRef] [PubMed]
- Boltz, K.W.; Gonzalez-Moa, M.J.; Stafford, P.; Johnston, S.A.; Svarovsky, S.A. Peptide microarrays for carbohydrate recognition. Analyst 2009, 134, 650–652. [Google Scholar] [CrossRef] [PubMed]
- Frank, R. Spot-synthesis: An easy technique for the positionally addressable, parallel chemical synthesis on a membrane support. Tetrahedron 1992, 48, 9217–9232. [Google Scholar] [CrossRef]
- Northen, T.R.; Greving, M.P.; Woodbury, N.W. Combinatorial screening of biomimetic protein affinity materials. Adv. Mater. 2008, 20, 4691–4697. [Google Scholar] [CrossRef]
- Fu, J.; Cai, K.; Johnston, S.A.; Woodbury, N.W. Exploring peptide space for enzyme modulators. J. Am. Chem. Soc. 2010, 132, 6419–6424. [Google Scholar] [CrossRef] [PubMed]
- Fu, J.; Reinhold, J.; Woodbury, N.W. Peptide-modified surfaces for enzyme immobilization. PLoS ONE 2011, 6, e18692. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Peptide | Sequence | β-Gal Binding 1 (a.u.) | PEP-1/β-Gal Binding 2 (a.u.) |
|---|---|---|---|
| NEW-1 | GVSHLHWIKMLNETTVMGSC | 486.8 ± 81.3 | 27,417.3 ± 4933.1 |
| NEW-2 | HISPQHMMAYSPKAFDYGSC | 301.0 ± 53.8 | 21,378.3 ± 4007.7 |
| NEW-3 | YDTLHRNRQMMDWQFEPGSC | 334.7 ± 42.9 | 25,598.5 ± 2407.7 |
| NEW-4 | MHNHAFNDNHGRGPTAWGSC | 1210 ± 184.6 | 30,398.0 ± 3998.7 |
© 2017 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fu, J. Microarray Selection of Cooperative Peptides for Modulating Enzyme Activities. Microarrays 2017, 6, 8. https://doi.org/10.3390/microarrays6020008
Fu J. Microarray Selection of Cooperative Peptides for Modulating Enzyme Activities. Microarrays. 2017; 6(2):8. https://doi.org/10.3390/microarrays6020008
Chicago/Turabian StyleFu, Jinglin. 2017. "Microarray Selection of Cooperative Peptides for Modulating Enzyme Activities" Microarrays 6, no. 2: 8. https://doi.org/10.3390/microarrays6020008
APA StyleFu, J. (2017). Microarray Selection of Cooperative Peptides for Modulating Enzyme Activities. Microarrays, 6(2), 8. https://doi.org/10.3390/microarrays6020008