
The Protective Effect of Taurine on Oxidized Fish-Oil-Induced Liver Oxidative Stress and Intestinal Barrier-Function Impairment in Juvenile Ictalurus punctatus

Abstract
:1. Introduction
2. Materials and Methods
2.1. Preparation of Oxidized Fish Oil
2.2. Experimental Diets
2.3. Experimental Animals and Feeding Experiment
2.4. Sample Collection
2.5. Determination of Growth Parameters
2.6. Skin Pigment and Body Color Analysis
2.7. Biochemical Index Analysis
2.8. Histological Analysis
2.9. Real-Time Polymerase Chain Reaction
2.10. Statistical Analysis
3. Results
3.1. Growth Performance
3.2. Skin Pigment and Body Color
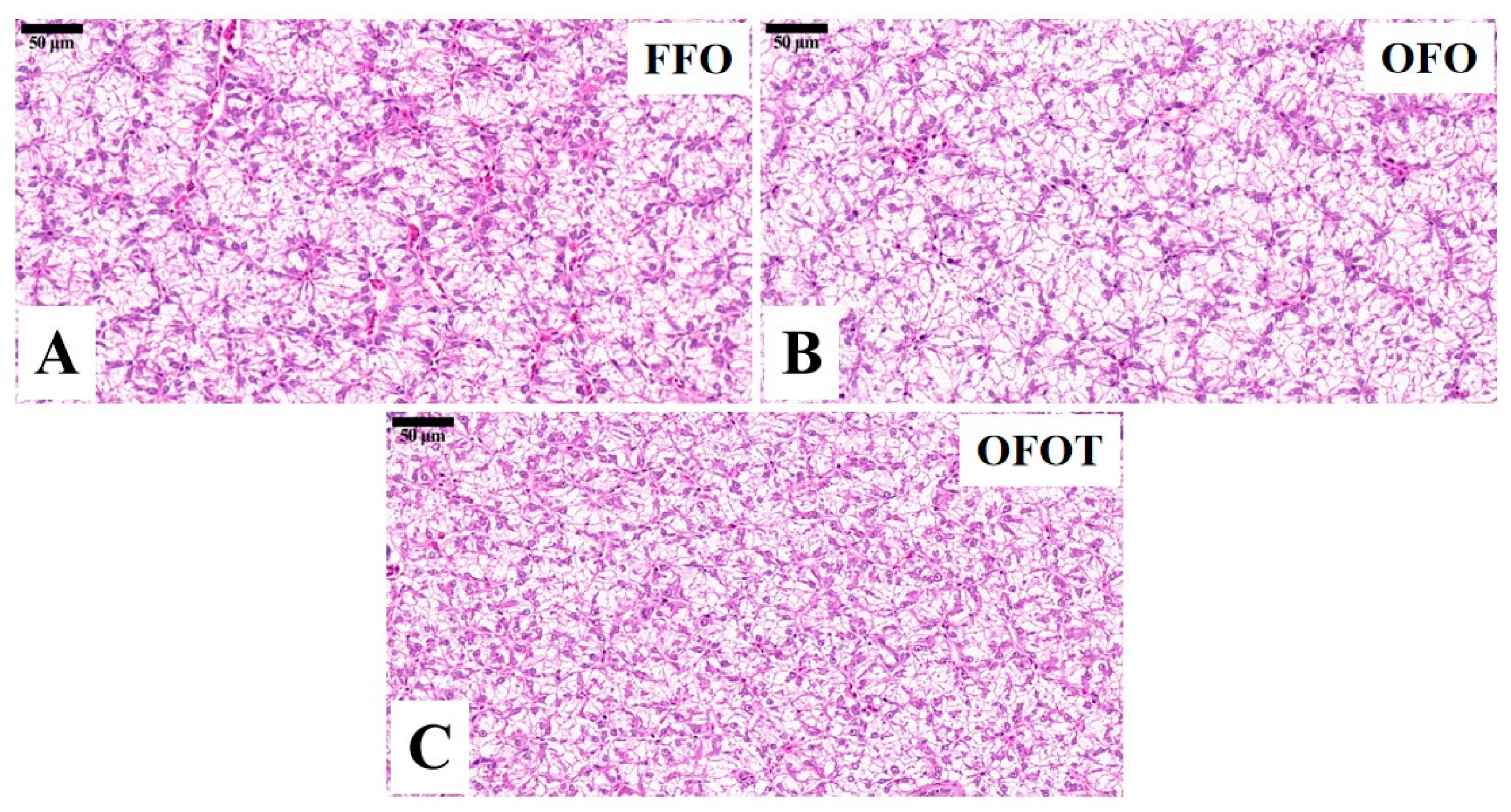
3.3. Lipid Deposition and Histological Structure of Liver
3.4. Serum Immune Indices
3.5. Antioxidant Indices in the Liver
3.6. Antioxidant-Related Gene Expression in the Liver
3.7. Histological Structure in the Intestine
3.8. Intestinal Physical-Barrier-Related Gene Expression
3.9. Intestinal Immune-Barrier-Related Gene Expression
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Sargent, J.; Bell, G.; McEvoy, L.; Tocher, D.; Estevez, A. Recent developments in the essential fatty acid nutrition of fish. Aquaculture 1999, 177, 191–199. [Google Scholar] [CrossRef]
- Zhou, L.; Han, D.; Zhu, X.; Yang, Y.; Jin, J.; Xie, S. Effects of total replacement of fish oil by pork lard or rapeseed oil and recovery by a fish oil finishing diet on growth, health and fish quality of gibel carp (Carassius auratus gibelio). Aquac. Res. 2016, 47, 2961–2975. [Google Scholar] [CrossRef]
- Gray, J.I. Measurement of lipid oxidation: A review. J. Am. Oil Chem. Soc. 1978, 55, 539–546. [Google Scholar] [CrossRef]
- Chen, Y.J.; Liu, Y.J.; Yang, H.J.; Yuan, Y.; Liu, F.J.; Tian, L.X.; Liang, G.Y.; Yuan, R.M. Effect of dietary oxidized fish oil on growth performance, body composition, antioxidant defence chanism and liver histology of juvenile largemouth bass Micropterus salmoides. Aquac. Nutr. 2012, 18, 321–331. [Google Scholar] [CrossRef]
- Liu, D.; Chi, S.; Tan, B.; Dong, X.; Yang, Q.; Liu, H.; Zhang, S.; Han, F.; He, Y. Effects of fish oil with difference oxidation degree on growth performance and expression abundance of antioxidant and fat metabolism genes in orange spotted grouper, Epinephelus coioides. Aquac. Res. 2019, 50, 188–197. [Google Scholar] [CrossRef] [Green Version]
- Yu, L.J.; Wen, H.; Jiang, M.; Wu, F.; Tian, J.; Lu, X.; Xiao, J.; Liu, W. Effects of ferulic acid on growth performance, immunity and antioxidant status in genetically improved farmed tilapia (Oreochromis niloticus) fed oxidized fish oil. Aquac. Nutr. 2020, 26, 1431–1442. [Google Scholar] [CrossRef]
- Song, C.; Liu, B.; Xu, P.; Ge, X.; Zhang, H. Emodin ameliorates metabolic and antioxidant capacity inhibited by dietary oxidized fish oil through PPARs and Nrf2-Keap1 signaling in Wuchang bream (Megalobrama amblycephala). Fish Shellfish Immunol. 2019, 94, 842–851. [Google Scholar]
- Yu, T.; Wang, Q.J.; Chen, X.M.; Chen, Y.K.; Ghonimy, A.; Zhang, D.M.; Wang, G.Q. Effect of dietary L-carnitine supplementation on growth performance and lipid metabolism in Rhynchocypris lagowski Dybowski fed oxidized fish oil. Aquac. Res. 2020, 51, 3698–3710. [Google Scholar] [CrossRef]
- Song, C.; Liu, B.; Xu, P.; Xie, J.; Ge, X.; Zhou, Q.; Sun, C.; Zhang, H.; Shan, F.; Yang, Z. Oxidized fish oil injury stress in Megalobrama amblycephala: Evaluated by growth, intestinal physiology, and transcriptome-based PI3K-Akt/NF-κB/TCR inflammatory signaling. Fish Shellfish Immunol. 2018, 81, 446–455. [Google Scholar] [CrossRef]
- Peng, M.; Luo, H.; Kumar, V.; Kajbaf, K.; Hu, Y.; Yang, G. Dysbiosis of intestinal microbiota induced by dietary oxidized fish oil and recovery of diet-induced dysbiosis via taurine supplementation in rice field eel (Monopterus albus). Aquaculture 2019, 512, 734288. [Google Scholar] [CrossRef]
- Huxtable, R. Physiological actions of taurine. Physiol. Rev. 1992, 72, 101–142. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oliveira, M.W.; Minotto, J.B.; de Oliveira, M.R.; Zanotto-Filho, A.; Behr, G.A.; Rocha, R.F.; Moreira, J.C.F.; Klamt, F. Scavenging and antioxidant potential of physiological taurine concentrations against different reactive oxygen/nitrogen species. Pharmacol. Rep. 2010, 62, 185–193. [Google Scholar] [CrossRef]
- Zhang, J.Z.; Hu, Y.; Ai, Q.H.; Mao, P.; Tian, Q.Q.; Zhong, L.; Xiao, T.Y.; Chu, W.Y. Effect of dietary taurine supplementation on growth performance, digestive enzyme activities and antioxidant status of juvenile black carp (Mylopharyngodon piceus) fed with low fish meal diet. Aquac. Res. 2018, 49, 3187–3195. [Google Scholar] [CrossRef]
- Hu, Y.; Yang, G.; Li, Z.; Hu, Y.; Zhong, L.; Zhou, Q.; Peng, M. Effect of dietary taurine supplementation on growth, digestive enzyme, immunity and resistant to dry stress of rice field eel (Monopterus albus) fed low fish meal diets. Aquac. Res. 2018, 49, 2108–2118. [Google Scholar]
- Martins, N.; Diógenes, A.F.; Magalhães, R.; Matas, I.; Oliva-Teles, A.; Peres, H. Dietary taurine supplementation affects lipid metabolism and improves the oxidative status of European seabass (Dicentrarchus labrax) juveniles. Aquaculture 2021, 531, 735820. [Google Scholar] [CrossRef]
- Yan, L.C.; Feng, L.; Jiang, W.D.; Wu, P.; Liu, Y.; Jiang, J.; Tang, L.; Tang, W.N.; Zhang, Y.A.; Yang, J.; et al. Dietary taurine supplementation to a plant protein source-based diet improved the growth and intestinal immune function of young grass carp (Ctenopharyngodon idella). Aquac. Nutr. 2019, 25, 873–896. [Google Scholar] [CrossRef]
- Garcia-Organista, A.A.; Mata-Sotres, J.A.; Viana, M.T.; Rombenso, A.N. The effects of high dietary methionine and taurine are not equal in terms of growth and lipid metabolism of juvenile California Yellowtail (Seriola dorsalis). Aquaculture 2019, 512, 734304. [Google Scholar] [CrossRef]
- FAO. Fishery and Aquaculture Statistics 2018/FAO Annuaire; Food and Agriculture Organization of the United Nations: Rome, Italy, 2020. [Google Scholar]
- Shi, Y.; Zhong, L.; Ma, X.; Liu, Y.; Tang, T.; Hu, Y. Effect of replacing fishmeal with stickwater hydrolysate on the growth, serum biochemical indexes, immune indexes, intestinal histology and microbiota of rice field eel (Monopterus albus). Aquac. Rep. 2019, 15, 100223. [Google Scholar] [CrossRef]
- Shi, Y.; Zhong, L.; Zhang, J.Z.; Ma, X.K.; Zhong, H.; Peng, M.; He, H.; Hu, Y. Substitution of fish meal with krill meal in rice field eel (Monopterus albus) diets: Effects on growth, immunity, muscle textural quality, and expression of myogenic regulation factors. Anim. Feed Sci. Technol. 2021, 280, 115047. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Schmittgen, Analysis of relative gene expression data using Real-Time Quantitative PCR and the 2−△△CT Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Zar, J.H. Biostatistical analysis. Q. Rev. Biol. 1999, 18, 797–799. [Google Scholar]
- Izquierdo, M.S.; Obach, A.; Arantzamendi, L.; Montero, D.; Robaina, L.; Rosenlund, G. Dietary lipid sources for seabream and seabass: Growth performance, tissue composition and flesh quality. Aquac. Nutr. 2003, 9, 397–407. [Google Scholar]
- Long, S.; Dong, X.; Tan, B.; Zhang, S.; Xie, S.; Yang, Q.; Chi, S.; Liu, H.; Deng, J.; Yang, Y.; et al. Growth performance, antioxidant ability, biochemical index in serum, liver histology and hepatic metabolomics analysis of juvenile hybrid grouper (♀ Epinephelus fuscoguttatus × ♂ Epinephelus lanceolatus) fed with oxidized fish oil. Aquaculture 2021, 545, 737261. [Google Scholar]
- Zhang, D.G.; Zhao, T.; Hogstrand, C.; Ye, H.M.; Xu, X.J.; Luo, Z. Oxidized fish oils increased lipid deposition via oxidative stress-mediated mitochondrial dysfunction and the CREB1-Bcl2-Beclin1 pathway in the liver tissues and hepatocytes of yellow catfish. Food Chem. 2021, 360, 129814. [Google Scholar] [CrossRef] [PubMed]
- Dong, G.F.; Huang, F.; Zhu, X.M.; Zhang, L.; Mei, M.X.; Hu, Q.W.; Liu, H.Y. Nutriphysiological and cytological responses of juvenile channel catfish (Ictalurus punctatus) to dietary oxidized fish oil. Aquac. Nutr. 2012, 18, 673–684. [Google Scholar] [CrossRef]
- Candebat, C.L.; Booth, M.; Codabaccus, M.B.; Pirozzi, I. Dietary methionine spares the requirement for taurine in juvenile Yellowtail Kingfish (Seriola lalandi). Aquaculture 2020, 522, 735090. [Google Scholar] [CrossRef]
- Zhang, Y.; Wei, Z.; Liu, G.; Deng, K.; Yang, M.; Pan, M.; Gu, Z.; Liu, D.; Zhang, W.; Mai, K. Synergistic effects of dietary carbohydrate and taurine on growth performance, digestive enzyme activities and glucose metabolism in juvenile turbot Scophthalmus maximus L. Aquaculture 2019, 499, 32–41. [Google Scholar] [CrossRef]
- Wei, Y.; Zhang, Q.; Xu, H.; Liang, M. Taurine requirement and metabolism response of tiger puffer Takifugu rubripes to graded taurine supplementation. Aquaculture 2020, 524, 735237. [Google Scholar] [CrossRef]
- Qi, G.; Ai, Q.; Mai, K.; Xu, W.; Liufu, Z.; Yun, B.; Zhou, H. Effects of dietary taurine supplementation to a casein-based diet on growth performance and taurine distribution in two sizes of juvenile turbot (Scophthalmus maximus L.). Aquaculture 2012, 358–359, 122–128. [Google Scholar] [CrossRef]
- Chen, K.; Ye, Y.; Cai, C.; Wu, P.; Huang, Y.; Wu, T.; Lin, X.; Luo, Q.; Zhang, B.; Xiao, P. Damage of oxidized fish oil on the structure and function of hepatopancreas of grass carp (Ctenopharyngodon idellus). Acta Hydrobiol. Sin. 2016, 40, 793–803. [Google Scholar]
- Prusińska, M.; Nowosad, J.; Jarmołowicz, S.; Mikiewicz, M.; Duda, A.; Wiszniewski, G.; Sikora, M.; Biegaj, M.; Samselska, A.; Arciuch-Rutkowska, M.; et al. Effect of feeding barbel larvae (Barbus barbus (L., 1758)) Artemia sp. nauplii enriched with PUFAs on their growth and survival rate, blood composition, alimentary tract histological structure and body chemical composition. Aquac. Rep. 2020, 18, 100492. [Google Scholar]
- Stejskal, V.; Gebauer, T.; Sebesta, R.; Nowosad, J.; Sikora, M.; Biegaj, M.; Kucharczyk, D. Effect of feeding strategy on survival, growth, intestine development, and liver status of maraena whitefish Coregonus maraena larvae. J. World Aquac. Soc. 2021, 52, 829–842. [Google Scholar] [CrossRef]
- Zhang, Y.; Li, Y.; Liang, X.; Cao, X.; Huang, L.; Yan, J.; Wei, Y.; Gao, J. Hepatic transcriptome analysis and identification of differentially expressed genes response to dietary oxidized fish oil in loach Misgurnus anguillicaudatus. PLoS ONE 2017, 12, e0172386. [Google Scholar] [CrossRef] [PubMed]
- Xie, S.; Yin, P.; Tian, L.; Liu, Y.; Niu, J. Lipid metabolism and plasma metabolomics of juvenile largemouth bass Micropterus salmoides were affected by dietary oxidized fish oil. Aquaculture 2020, 522, 735158. [Google Scholar] [CrossRef]
- Zhang, L.; Li, H.X.; Pan, W.S.; Khan, F.U.; Qian, C.; Qi-Li, F.R.; Xu, X. Novel hepatoprotective role of Leonurine hydrochloride against experimental non-alcoholic steatohepatitis mediated via AMPK/SREBP1 signaling pathway. Biomed. Pharmacother. 2019, 110, 571–581. [Google Scholar] [CrossRef] [PubMed]
- McPherson, R.; Gauthier, A. Molecular regulation of SREBP function: The Insig-SCAP connection and isoform-specific modulation of lipid synthesis. Biochem. Cell Biol. 2004, 82, 201–211. [Google Scholar] [CrossRef] [PubMed]
- Han, H.L.; Zhang, J.F.; Yan, E.F.; Shen, M.M.; Wu, J.M.; Gan, Z.D.; Wei, C.H.; Zhang, L.L.; Wang, T. Effects of taurine on growth performance, antioxidant capacity, and lipid metabolism in broiler chickens. Poult. Sci. 2020, 99, 5707–5717. [Google Scholar] [CrossRef] [PubMed]
- Magalhães, R.; Martins, N.; Martins, S.; Lopes, T.; Diáz-Rosales, P.; Pousão-Ferreira, P.; Oliva-Tele-S, A.; Peres, H. Is dietary taurine required for white seabream (Diplodus sargus) juveniles? Aquaculture 2018, 502, 296–302. [Google Scholar] [CrossRef]
- Hoseini, S.M.; Hosseini, S.A.; Eskandari, S.; Amirahmadi, M. Effect of dietary taurine and methionine supplementation on growth performance, body composition, taurine retention and lipid status of Persian sturgeon, Acipenser persicus (Borodin, 1897), fed with plant-based diet. Aquac. Nutr. 2018, 24, 324–331. [Google Scholar] [CrossRef]
- Tan, Y.; Kim, J.; Cheng, J.; Ong, M.; Lao, W.G.; Jin, X.L.; Lin, Y.G.; Xiao, L.; Zhu, X.Q.; Qu, X.Q. Green tea polyphenols ameliorate non-alcoholic fatty liver disease through upregulating AMPK activation in high fat fed Zucker fatty rats. World J. Gastroenterol. 2017, 23, 3805–3814. [Google Scholar] [CrossRef]
- Kim, S.J.; Tang, T.; Abbott, M.; Viscarra, J.A.; Wang, Y.; Sul, H.S. AMPK phosphorylates desnutrin/ATGL and hormone-sensitive lipase to regulate lipolysis and fatty acid oxidation within adipose tissue. Mol. Cell. Biol. 2016, 36, 1961–1976. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rahal, A.; Kumar, A.; Singh, V.; Yadav, B.; Tiwari, R.; Chakraborty, S.; Ghama, K. Oxidative stress, prooxidants, and antioxidants: The interplay. Biomed. Res. Int. 2014, 2014, 761264. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Giorgio, M.; Trinei, M.; Migliaccio, E.; Pelicci, P.G. Hydrogen peroxide: A metabolic by-product or a common mediator of ageing signals? Nat. Rev. Mol. Cell Biol. 2007, 8, 722–728. [Google Scholar] [CrossRef] [PubMed]
- Bondia-Pons, I.; Ryan, L.; Martinez, J.A. Oxidative stress and inflammation interactions in human obesity. J. Physiol. Biochem. 2012, 68, 701–711. [Google Scholar]
- Kanner, J.; Lapidot, T. The stomach as a bioreactor: Dietary lipid peroxidation in the gastric fluid and the effects of plant-derived antioxidants. Free Radic. Biol. Med. 2001, 31, 1388–1395. [Google Scholar] [CrossRef]
- Dinu, D.; Marinescu, D.; Munteanu, M.C.; Staicu, A.C.; Costache, M.; Dinischiotu, A. Modulatory effects of deltamethrin on antioxidant defense mechanisms and lipid peroxidation in Carassius auratus gibelio liver and intestine. Arch. Environ. Contam. Toxicol. 2010, 58, 757–764. [Google Scholar] [CrossRef] [PubMed]
- Ramalingam, M.; Kim, S.J. Reactive oxygen/nitrogen species and their functional correlations in neurodegenerative diseases. J. Neural Transm. 2012, 119, 891–910. [Google Scholar]
- Mehra, L.; Hasija, Y.; Mittal, G. Therapeutic potential of alpha-ketoglutarate against acetaminophen-induced hepatotoxicity in rats. J. Pharm. Bioallied Sci. 2016, 8, 296–299. [Google Scholar]
- Aly, H.A.; Khafagy, R.M. Taurine reverses endosulfan-induced oxidative stress and apoptosis in adult rat testis. Food Chem. Toxicol. 2014, 64, 1–9. [Google Scholar]
- Salze, G.P.; Davis, D.A. Taurine: A critical nutrient for future fish feeds. Aquaculture 2015, 437, 215–229. [Google Scholar] [CrossRef]
- Hayes, J.; Tipton, K.F.; Bianchi, L.; Corte, L.D. Complexities in the neurotoxic actions of 6-hydroxydopamine in relation to the cytoprotective properties of taurine. Brain. Res. Bull. 2001, 55, 239–245. [Google Scholar] [CrossRef]
- Morales, A.E.; Pérez-Jiménez, A.; Hidalgo, M.C.; Abellán, E.; Cardenete, G. Oxidative stress and antioxidant defenses after prolonged starvation in Dentex dentex liver. Comp. Biochem. Physiol. C Toxicol. Pharmacol. 2004, 139, 153–161. [Google Scholar] [CrossRef]
- Shi, Y.; Zhong, L.; Liu, Y.L.; Zhang, J.Z.; Lv, Z.; Li, Y.; Hu, Y. Effects of dietary andrographolide levels on growth performance, antioxidant capacity, intestinal immune function and microbioma of rice field eel (Monopterus albus). Animals 2020, 10, 1744. [Google Scholar] [CrossRef] [PubMed]
- Jung, K.A.; Kwak, M.K. The Nrf2 system as a potential target for the development of indirect antioxidants. Molecules 2010, 15, 7266–7291. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cheng, C.H.; Guo, Z.X.; Wang, A.L. The protective effects of taurine on oxidative stress, cytoplasmic free-Ca2+ and apoptosis of pufferfish (Takifugu obscurus) under low temperature stress. Fish Shellfish Immunol. 2018, 77, 457–464. [Google Scholar] [CrossRef]
- Zhang, M.; Li, M.; Wang, R.; Qian, Y. Effects of acute ammonia toxicity on oxidative stress, immune response and apoptosis of juvenile yellow catfish Pelteobagrus fulvidraco and the mitigation of exogenous taurine. Fish Shellfish Immunol. 2018, 79, 313–320. [Google Scholar] [CrossRef]
- Zhang, Y.; Wei, Z.; Yang, M.; Liu, D.; Pan, M.; Wu, C.; Zhang, W.; Mai, K. Dietary taurine modulates hepatic oxidative status, ER stress and inflammation in juvenile turbot (Scophthalmus maximus L.) fed high carbohydrate diets. Fish Shellfish Immunol. 2021, 109, 1–11. [Google Scholar] [CrossRef]
- Ni, P.J.; Jiang, W.D.; Wu, P.; Liu, Y.; Kuang, S.Y.; Tang, L.; Tang, W.N.; Zhang, Y.A.; Zhou, X.Q.; Feng, L. Dietary low or excess levels of lipids reduced growth performance, and impaired immune function and structure of head kidney, spleen and skin in young grass carp (Ctenopharyngodon idella) under the infection of Aeromonas hydrophila. Fish Shellfish Immunol. 2016, 55, 28–47. [Google Scholar] [CrossRef] [PubMed]
- Ai, Q.H.; Mai, K.S. Advance on nutritional immunity of fish. Acta Hydrobiol. Sin. 2007, 31, 425–430. [Google Scholar]
- Mori, K.; Nakanishi, T.; Suzuki, T.; Oohara, I. Defense mechanisms in invertebrates and fish. Tanpakushitsu Kakusan Koso 1989, 34, 214–223. [Google Scholar]
- Shi, Y.; Zhong, L.; Zhong, H.; Zhang, J.; Che, C.; Fu, G.; Hu, Y.; Mai, K. Taurine supplements in high-fat diets improve survival of juvenile Monopterus albus by reducing lipid deposition and intestinal damage. Aquaculture 2022, 547, 737431. [Google Scholar] [CrossRef]
- Dong, J.; Cheng, R.; Yang, Y.; Zhao, Y.; Wu, G.; Zhang, R.; Zhu, X.; Li, L.; Li, X. Effects of dietary taurine on growth, non-specific immunity, anti-oxidative properties and gut immunity in the Chinese mitten crab Eriocheir sinensis. Fish Shellfish Immunol. 2018, 82, 212–219. [Google Scholar] [CrossRef]
- Li, M.; Lai, H.; Li, Q.; Gong, S.; Wang, R. Effects of dietary taurine on growth, immunity and hyperammonemia in juvenile yellow catfish Pelteobagrus fulvidraco fed all-plant protein diets. Aquaculture 2016, 450, 349–355. [Google Scholar] [CrossRef]
- Kim, C.; Cha, Y.N. Taurine chloramine produced from taurine under inflammation provides anti-inflammatory and cytoprotective effects. Amino Acids. 2014, 46, 89–100. [Google Scholar] [CrossRef]
- Rymuszka, A.; Adaszek, Ł. Pro- and anti-inflammatory cytokine expression in carp blood and head kidney leukocytes exposed to cyanotoxin stress--an in vitro study. Fish Shellfish Immunol. 2012, 33, 382–388. [Google Scholar] [CrossRef]
- Zhang, D.M.; Guo, Z.X.; Zhao, Y.L.; Wang, Q.J.; Gao, Y.S.; Yu, T.; Chen, Y.K.; Chen, X.M.; Wang, G.Q. L-carnitine regulated Nrf2/Keap1 activation in vitro and in vivo and protected oxidized fish oil-induced inflammation response by inhibiting the NF-κB signaling pathway in Rhynchocypris lagowski Dybowski. Fish Shellfish Immunol. 2019, 93, 1100–1110. [Google Scholar] [CrossRef]
- Sen, R.; Baltimore, D. Multiple nuclear factors interact with the immunoglobulin enhancer sequences. Cell 1986, 46, 705–716. [Google Scholar] [CrossRef]
- Cario, E.; Gerken, G.; Podolsky, D.K. Toll-like receptor 2 controls mucosal inflammation by regulating epithelial barrier function. Gastroenterology 2007, 132, 1359–1374. [Google Scholar] [CrossRef]
- Pirarat, N.; Pinpimai, K.; Endo, M.; Katagiri, T.; Ponpornpisit, A.; Chansue, N.; Maita, M. Modulation of intestinal morphology and immunity in nile tilapia (Oreochromis niloticus) by Lactobacillus rhamnosus GG. Res. Vet. Sci. 2011, 91, e92–e97. [Google Scholar] [CrossRef]
- Farhangi, M.; Carter, C.G. Growth, physiological and immunological responses of rainbow trout (Oncorhynchus mykiss) to different dietary inclusion levels of dehuded lupin (Lupinus angustifolius). Aquac. Res. 2002, 32, 329–340. [Google Scholar] [CrossRef]
- Xu, H.J.; Jiang, W.D.; Feng, L.; Liu, Y.; Wu, P.; Jiang, J.; Kuang, S.Y.; Tang, L.; Tang, W.N.; Zhang, Y.A.; et al. Dietary vitamin C deficiency depresses the growth, head kidney and spleen immunity and structural integrity by regulating NF-κB, TOR, Nrf2, apoptosis and MLCK signaling in young grass carp (Ctenopharyngodon idella). Fish Shellfish Immunol. 2016, 52, 111–138. [Google Scholar] [CrossRef] [PubMed]
- Xu, M.; Che, L.; Gao, K.; Wang, L.; Yang, X.; Wen, X.; Jiang, Z.; Wu, D. Effects of dietary taurine supplementation to gilts during late gestation and lactation on offspring growth and oxidative stress. Animals 2019, 9, 220. [Google Scholar] [CrossRef] [PubMed] [Green Version]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Ingredients | FFO | OFO | OFOT |
|---|---|---|---|
| Fish meal | 10.00 | 10.00 | 10.00 |
| Soybean meal | 28.00 | 28.00 | 28.00 |
| Rapeseed meal | 20.00 | 20.00 | 20.00 |
| Rice bran | 3.00 | 3.00 | 3.00 |
| Wheat flour | 25.26 | 25.26 | 25.06 |
| Chicken meal | 9.00 | 9.00 | 9.00 |
| Fish oil | 2.00 | 0.00 | 0.00 |
| Oxidized fish oil | 0.00 | 2.00 | 2.00 |
| Premix 1 | 1.00 | 1.00 | 1.00 |
| Choline | 0.20 | 0.20 | 0.20 |
| Ca(H2PO4)2 | 1.50 | 1.50 | 1.50 |
| Mold inhibitor | 0.03 | 0.03 | 0.03 |
| Antioxidants | 0.01 | 0.01 | 0.01 |
| Taurine | 0.00 | 0.00 | 0.20 |
| Approximate composition (%) 2 | |||
| Crude protein | 36.27 | 36.14 | 36.31 |
| Crude lipid | 5.47 | 5.52 | 5.43 |
| Crude ash | 6.57 | 6.49 | 6.53 |
| POV (meq/kg) | 3.7 | 21.4 | 21.2 |
| Gene | Forward Sequences (5′→3′) | Reverse Sequences (5′→3′) | Accession No. |
|---|---|---|---|
| fas | CTGCTGTCTGAGGGCGTAA | CGATGGCGATGAGGTTCT | NM_001200193.1 |
| lpl | AGCAACATTACCCAACCTCAGC | CCAGCTACATGAGCACCCAAA | KF693235.1 |
| srebp1 | GTTGCGGAAGGCGATTGA | GCAGTGGGCTGTTGGGTTC | XM_017480901.1 |
| sod | GACTTGGGCAAAGGTGGAAA | CACTCAGCAATGCCTATCACG | NM_001200992.1 |
| gpx1 | TCTGAGGCACGACCACCA | GCGTCTTTCCCGTTCACAT | NM_001200741.1 |
| gpx8 | TCACTTCACCGTGTTGGCTT | CCCTCAGCACTCACCAGAAA | XM_017466944.1 |
| gr | GGATGTGAAGGATAAGCGAAAC | TTCGGCAACACGGGTATG | GU588318.1 |
| keap1 | CGGCAAGCATCTCAGTCG | TGCTCGGGTCCAACTGC | XM_017482237.1 |
| nrf2 | GGTCCACGCCTACCAACAA | CAGGGAGGAATGGAGGGAT | XM_017470076.1 |
| zo-1 | TACCAAACCGTGGATACAAACC | CTTCTATGGGTGGAGGAGGC | XM_017458510.1 |
| zo-2 | GAGGTCAAAGGGCAGCAAA | GAAATCTTCGGGCAGGTCA | XM_017488926.1 |
| claudin-12 | GCTGGGATGTTCCTCTTGATAG | AGAGCGGCGAACTCAAGG | XM_017453476.1 |
| claudin-15 | GTGGTTCTCGGCACATTCG | CAAGCCCTGTAGGATGAAGAAG | XM_017471911.1 |
| occludin | GCATCGGTAGCGGGTCAT | GACTTGGTTGAGTTCTGCCTTG | XM_017451558.1 |
| tnf-α | CGCCAGCGGTAAACACG | CCGTTGAATGTCCGAAAGG | XM_017464718.1 |
| il-1β | CTGAAGGGTGGAAACAAGGAT | GGAGTCACCAGTGCCGTTT | AJ586102.1 |
| il-6 | GAAGATTGATACTCCGCTCCTG | GATTAAATGTAACAGCCTGGTGG | XM_017455306.1 |
| il-8 | TCCAAGTGCCTCCTGTTCAA | CCCTTCTTCCCTTGGACTTTAT | KP701473.1 |
| il-10 | GCAGGCTTACGAAAGGGTTA | CGGCGTATGAAGAACGAAGT | XM_017450800.1 |
| tgf-β1 | GGAACGGCTGAGTGGGTCT | TGCTTACTGAGGCGGCTATG | XM_017483625.1 |
| tgf-β2 | TGAAGCGGTCAGCGAATG | CTCACTCTTGTTTGGGATGATGTA | XM_017476217.1 |
| tgf-β3 | TCGGTGCCCTGTCCTATTG | GCGGAGAACGAGGCTTACA | XM_017476492.1 |
| nf-κb | CTCAGCCCATCTACGACAACA | CGTCAGGTTCGTATCGCAGT | KF572025.1 |
| gapdh | TGTCCGTTTGGAGAAGCCT | ATCAGGTCACAGACACGGTTG | NM_001201199.1 |
| FFO | OFO | OFOT | p-Value | |
|---|---|---|---|---|
| Initial weight (g) | 6.00 ± 0.01 | 6.00 ± 0.01 | 5.99 ± 0.00 | 0.702 |
| Final weight (g) | 26.20 ± 0.14 b | 23.43 ± 0.72 a | 25.62 ± 0.40 b | 0.036 |
| WGR | 336.53 ± 1.38 b | 290.49 ± 11.93 a | 327.27 ± 6.55 b | 0.034 |
| SR | 97.14 ± 2.86 | 94.29 ± 2.86 | 93.33 ± 6.67 | 0.829 |
| FCR | 1.16 ± 0.04 | 1.35 ± 0.09 | 1.20 ± 0.08 | 0.223 |
| HSI | 2.26 ± 0.05 a | 2.74 ± 0.12 b | 2.46 ± 0.07 a | 0.003 |
| VSI | 13.20 ± 0.66 | 13.67 ± 0.47 | 13.28 ± 0.30 | 0.781 |
| CF | 1.52 ± 0.04 | 1.57 ± 0.03 | 1.49 ± 0.01 | 0.216 |
| FFO | OFO | OFOT | p-Value | |
|---|---|---|---|---|
| Backside | ||||
| Carotenoids (μg/mL) | 16.00 ± 0.47 b | 14.04 ± 0.32 a | 18.42 ± 0.57 c | 0.002 |
| Lutein (pg/mL) | 1746.8 ± 84.5 a | 2331.8 ± 26.0 b | 2243.5 ± 17.3 b | <0.001 |
| Tyrosinase (ng/mL) | 4986.3 ± 43.3 a | 4699.7 ± 87.4 a | 5583.0 ± 120.0 b | 0.001 |
| L* | 51.14 ± 0.49 | 53.24 ± 1.25 | 47.64 ± 2.87 | 0.125 |
| a* | −5.10 ± 0.06 a | −5.44 ± 0.08 a | −4.54 ± 0.29 b | 0.008 |
| b* | 1.60 ± 0.23 a | 2.49 ± 0.15 ab | 2.07 ± 0.25 b | 0.031 |
| Abdomen | ||||
| Carotenoids (μg/mL) | 13.18 ± 0.23 a | 13.61 ± 0.07 ab | 14.91 ± 0.64 b | 0.049 |
| Lutein (pg/mL) | 2216.0 ± 21.7 b | 2391.8 ± 22.4 c | 2083.5 ± 40.1 a | 0.001 |
| Tyrosinase (ng/mL) | 4448.0 ± 31.8 a | 4309.7 ± 57.0 a | 4693.0 ± 75.5 b | 0.009 |
| L* | 82.41 ± 0.20 b | 81.52 ± 0.25 a | 81.55 ± 0.14 a | 0.011 |
| a* | −3.47 ± 0.06 ab | −3.55 ± 0.06 a | −3.31 ± 0.03 b | 0.017 |
| b* | 8.19 ± 0.05 | 8.67 ± 0.12 | 8.01 ± 0.42 | 0.200 |
| FFO | OFO | OFOT | p-Value | |
|---|---|---|---|---|
| Size of nuclei (μm) | 6.57 ± 0.02 | 5.35 ± 0.03 | 6.61 ± 0.02 | <0.001 |
| Size of hepatocytes (μm) | 17.03 ± 0.05 | 21.66 ± 0.26 | 17.02 ± 0.05 | <0.001 |
| FFO | OFO | OFOT | p-Value | |
|---|---|---|---|---|
| C3 (g/L) | 1.04 ± 0.03 c | 0.72 ± 0.02 a | 0.87 ± 0.03 b | <0.001 |
| C4 (g/L) | 0.65 ± 0.02 b | 0.5 ± 0.02 a | 0.69 ± 0.03 b | <0.001 |
| IgM (g/L) | 1.42 ± 0.08 b | 1.01 ± 0.03 a | 1.37 ± 0.07 b | 0.003 |
| AST (U/L) | 33.02 ± 6.31 a | 56.45 ± 0.97 b | 35.2 ± 0.42 a | 0.008 |
| ALT (U/L) | 8.65 ± 0.58 a | 13.33 ± 0.77 b | 7.57 ± 0.62 a | <0.001 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Shi, Y.; Hu, Y.; Wang, Z.; Zhou, J.; Zhang, J.; Zhong, H.; Fu, G.; Zhong, L. The Protective Effect of Taurine on Oxidized Fish-Oil-Induced Liver Oxidative Stress and Intestinal Barrier-Function Impairment in Juvenile Ictalurus punctatus. Antioxidants 2021, 10, 1690. https://doi.org/10.3390/antiox10111690
Shi Y, Hu Y, Wang Z, Zhou J, Zhang J, Zhong H, Fu G, Zhong L. The Protective Effect of Taurine on Oxidized Fish-Oil-Induced Liver Oxidative Stress and Intestinal Barrier-Function Impairment in Juvenile Ictalurus punctatus. Antioxidants. 2021; 10(11):1690. https://doi.org/10.3390/antiox10111690
Chicago/Turabian StyleShi, Yong, Yi Hu, Ziqin Wang, Jiancheng Zhou, Junzhi Zhang, Huan Zhong, Guihong Fu, and Lei Zhong. 2021. "The Protective Effect of Taurine on Oxidized Fish-Oil-Induced Liver Oxidative Stress and Intestinal Barrier-Function Impairment in Juvenile Ictalurus punctatus" Antioxidants 10, no. 11: 1690. https://doi.org/10.3390/antiox10111690
APA StyleShi, Y., Hu, Y., Wang, Z., Zhou, J., Zhang, J., Zhong, H., Fu, G., & Zhong, L. (2021). The Protective Effect of Taurine on Oxidized Fish-Oil-Induced Liver Oxidative Stress and Intestinal Barrier-Function Impairment in Juvenile Ictalurus punctatus. Antioxidants, 10(11), 1690. https://doi.org/10.3390/antiox10111690