
Fermentation Affects the Antioxidant Activity of Plant-Based Food Material through the Release and Production of Bioactive Components

 ,
, 
 , ,
, ,
Abstract
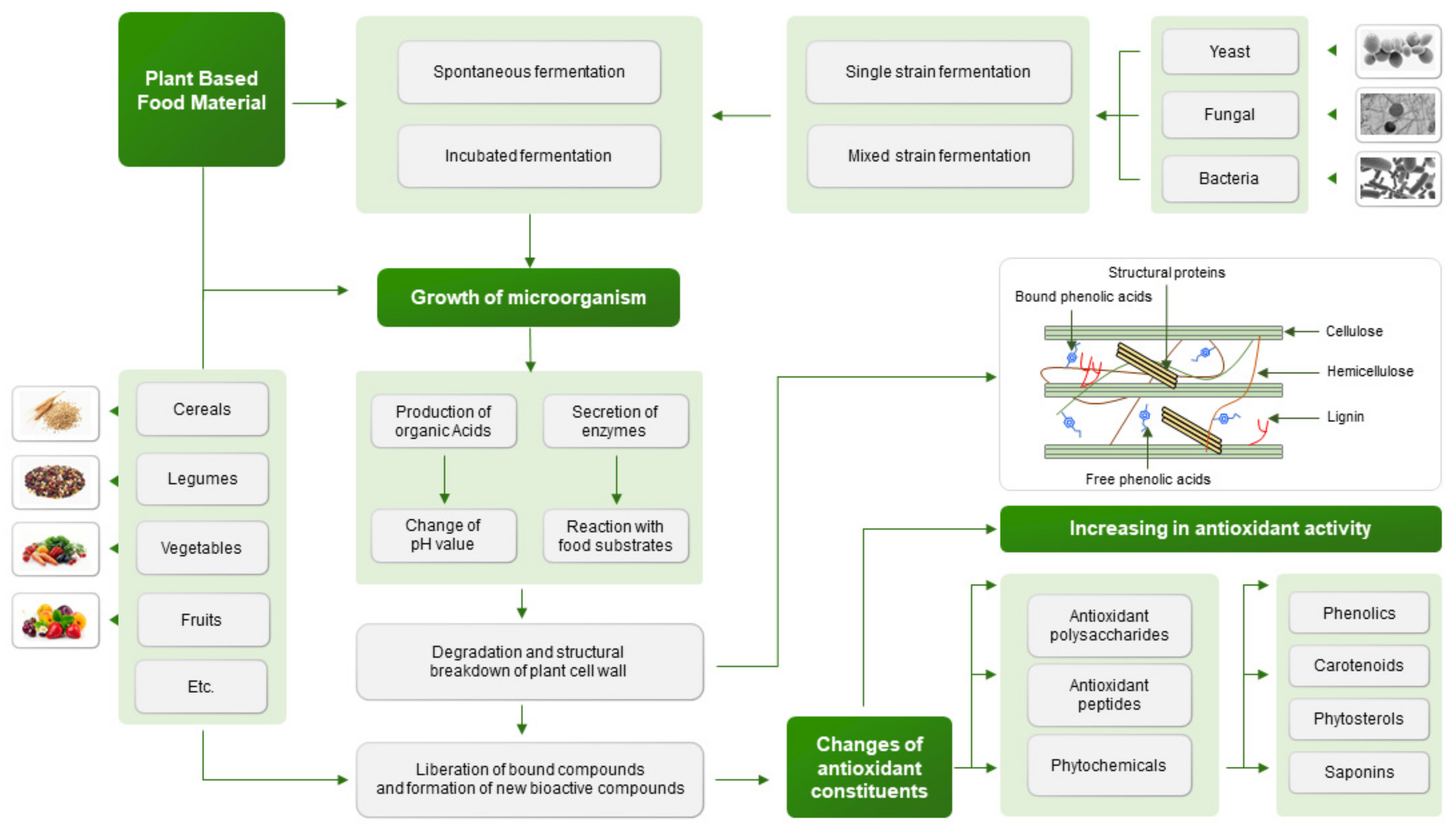
:1. Introduction
2. Impact of Fermentation Strain on Antioxidant Activity of Plant-Based Food Material
2.1. Single Strain Fermentation
2.1.1. Yeast Fermentation
2.1.2. Fungal Fermentation
2.1.3. Bacterial Fermentation
2.2. Mixed Strain Fermentation
3. Impact of Fermentation on Antioxidant Activity of Various Plant-Based Food Materials
3.1. Cereals
3.2. Legumes
3.3. Vegetables
3.4. Fruits
4. Changes of Antioxidant Constituents in Plant-Based Food Material during Fermentation
4.1. Phytochemicals
4.1.1. Phenolic Compounds
4.1.2. Carotenoids
4.1.3. Phytosterols
4.1.4. Saponins
4.2. Antioxidant Peptides
4.3. Antioxidant Polysaccharides
4.4. Conversion Effect of Fermentation on Antioxidant Components
5. Effects of Various Factors on Changes to Antioxidant Components and Activities during Fermentation
5.1. Effect of Enzyme on Fermentation Results
5.2. Effect of Fermentation Conditions on Fermentation Results
5.3. Selection of Liquid Fermentation and Solid-State Fermentation
6. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Jiménez-López, J.; Ruiz-Medina, A.; Ortega-Barrales, P.; Llorent-Martínez, E.J. Phytochemical profile and antioxidant activity of caper berries (Capparis spinosa L.): Evaluation of the influence of the fermentation process. Food Chem. 2018, 250, 54–59. [Google Scholar] [CrossRef] [PubMed]
- Chung, I.-M.; Seo, S.-H.; Ahn, J.-K.; Kim, S.-H. Effect of processing, fermentation, and aging treatment to content and profile of phenolic compounds in soybean seed, soy curd and soy paste. Food Chem. 2011, 127, 960–967. [Google Scholar] [CrossRef] [PubMed]
- Ravichandran, K.; Saw, N.M.M.T.; Mohdaly, A.A.A.; Gabr, A.M.M.; Kastell, A.; Riedel, H.; Cai, Z.; Knorr, D.; Smetanska, I. Impact of processing of red beet on betalain content and antioxidant activity. Food Res. Int. 2013, 50, 670–675. [Google Scholar] [CrossRef]
- Frias, J.; Miranda, M.L.; Doblado, R.; Vidal-Valverde, C. Effect of germination and fermentation on the antioxidant vitamin content and antioxidant capacity of Lupinus albus L. var. Multolupa. Food Chem. 2005, 92, 211–220. [Google Scholar] [CrossRef]
- Septembre-Malaterre, A.; Remize, F.; Poucheret, P. Fruits and vegetables, as a source of nutritional compounds and phytochemicals: Changes in bioactive compounds during lactic fermentation. Food Res. Int. 2018, 104, 86–99. [Google Scholar] [CrossRef]
- Alrahmany, R.; Avis, T.J.; Tsopmo, A. Treatment of oat bran with carbohydrases increases soluble phenolic acid content and influences antioxidant and antimicrobial activities. Food Res. Int. 2013, 52, 568–574. [Google Scholar] [CrossRef]
- Budaraju, S.; Mallikarjunan, K.; Annor, G.; Schoenfuss, T.; Raun, R. Effect of pre-treatments on the antioxidant potential of phenolic extracts from barley malt rootlets. Food Chem. 2018, 266, 31–37. [Google Scholar] [CrossRef]
- López-Gámez, G.; Elez-Martínez, P.; Martín-Belloso, O.; Soliva-Fortuny, R. Enhancing phenolic content in carrots by pulsed electric fields during post-treatment time: Effects on cell viability and quality attributes. Innov. Food Sci. Emerg. Technol. 2020, 59, 102252. [Google Scholar] [CrossRef]
- Goldsmith, C.D.; Vuong, Q.V.; Stathopoulos, C.E.; Roach, P.D.; Scarlett, C.J. Ultrasound increases the aqueous extraction of phenolic compounds with high antioxidant activity from olive pomace. LWT Food Sci. Technol. 2018, 89, 284–290. [Google Scholar] [CrossRef] [Green Version]
- Peinado, J.; López de Lerma, N.; Peralbo-Molina, A.; Priego-Capote, F.; de Castro, C.; McDonagh, B. Sunlight exposure increases the phenolic content in postharvested white grapes. An evaluation of their antioxidant activity in Saccharomyces cerevisiae. J. Func. Foods 2013, 5, 1566–1575. [Google Scholar] [CrossRef]
- Maseko, I.; Mabhaudhi, T.; Ncube, B.; Tesfay, S.; Araya, H.T.; Fessehazion, M.K.; Chimonyo, V.G.P.; Ndhlala, A.R.; Du Plooy, C.P. Postharvest drying maintains phenolic, flavonoid and gallotannin content of some cultivated African leafy vegetables. Sci. Hortic. 2019, 255, 70–76. [Google Scholar] [CrossRef]
- El Khalifa, A.O.; El Tinay, A.H. Effect of fermentation on protein fractions and tannin content of low- and high-tannin cultivars of sorghum. Food Chem. 1994, 49, 265–269. [Google Scholar] [CrossRef]
- Hur, S.J.; Lee, S.Y.; Kim, Y.-C.; Choi, I.; Kim, G.-B. Effect of fermentation on the antioxidant activity in plant-based foods. Food Chem. 2014, 160, 346–356. [Google Scholar] [CrossRef]
- Ricci, A.; Cirlini, M.; Calani, L.; Bernini, V.; Neviani, E.; Del Rio, D.; Galaverna, G.; Lazzi, C. In vitro metabolism of elderberry juice polyphenols by lactic acid bacteria. Food Chem. 2019, 276, 692–699. [Google Scholar] [CrossRef]
- Đorđević, T.M.; Šiler-Marinković, S.S.; Dimitrijević-Branković, S.I. Effect of fermentation on antioxidant properties of some cereals and pseudo cereals. Food Chem. 2010, 119, 957–963. [Google Scholar] [CrossRef]
- Maria John, K.M.; Thiruvengadam, M.; Enkhtaivan, G.; Kim, D.H. Variation in major phenolic compounds and quality potential of CTC black tea elicited by Saccharomyces cercevisiae and its correlation with antioxidant potential. Ind. Crop. Prod. 2014, 55, 289–294. [Google Scholar] [CrossRef]
- Ooi, T.S.; Ting, A.S.Y.; Siow, L.F. Influence of selected native yeast starter cultures on the antioxidant activities, fermentation index and total soluble solids of Malaysia cocoa beans: A simulation study. LWT Food Sci. Technol. 2020, 122, 108977. [Google Scholar] [CrossRef]
- Laaksonen, O.; Kuldjärv, R.; Paalme, T.; Virkki, M.; Yang, B. Impact of apple cultivar, ripening stage, fermentation type and yeast strain on phenolic composition of apple ciders. Food Chem. 2017, 233, 29–37. [Google Scholar] [CrossRef] [PubMed]
- Puškaš, V.S.; Miljić, U.D.; Djuran, J.J.; Vučurović, V.M. The aptitude of commercial yeast strains for lowering the ethanol content of wine. Food Sci. Nutr. 2020, 8, 1489–1498. [Google Scholar] [CrossRef]
- Kim, M.J.; John, K.M.M.; Choi, J.N.; Lee, S.; Kim, A.J.; Kim, Y.M.; Lee, C.H. Changes in secondary metabolites of green tea during fermentation by Aspergillus oryzae and its effect on antioxidant potential. Food Res. Int. 2013, 53, 670–677. [Google Scholar] [CrossRef]
- Verni, M.; Verardo, V.; Rizzello, C.G. How fermentation affects the antioxidant properties of cereals and legumes. Foods 2019, 8, 362. [Google Scholar] [CrossRef] [Green Version]
- Reis, B.; Gnazzo, A.K.-C.; Schmitt, R.; Andlauer, W. Fermentation of plant material—Effect on sugar content and stability of bioactive compounds. Pol. J. Food Nutr. Sci. 2014, 64, 235–241. [Google Scholar] [CrossRef] [Green Version]
- Anson, N.M.; Selinheimo, E.; Havenaar, R.; Aura, A.M.; Mattila, I.; Lehtinen, P.; Bast, A.; Poutanen, K.; Haenen, G.R. Bioprocessing of wheat bran improves in vitro bioaccessibility and colonic metabolism of phenolic compounds. J. Agric. Food Chem. 2009, 57, 6148–6155. [Google Scholar] [CrossRef]
- Ajila, C.M.; Gassara, F.; Brar, S.K.; Verma, M.; Tyagi, R.D.; Valéro, J.R. Polyphenolic antioxidant mobilization in apple pomace by different methods of solid-state fermentation and evaluation of its antioxidant activity. Food Bioprocess Tech. 2012, 5, 2697–2707. [Google Scholar] [CrossRef]
- Dulf, F.V.; Vodnar, D.C.; Socaciu, C. Effects of solid-state fermentation with two filamentous fungi on the total phenolic contents, flavonoids, antioxidant activities and lipid fractions of plum fruit (Prunus domestica L.) by-products. Food Chem. 2016, 209, 27–36. [Google Scholar] [CrossRef] [PubMed]
- Hifney, A.F.; Fawzy, M.A.; Abdel-Gawad, K.M.; Gomaa, M. Upgrading the antioxidant properties of fucoidan and alginate from Cystoseira trinodis by fungal fermentation or enzymatic pretreatment of the seaweed biomass. Food Chem. 2018, 269, 387–395. [Google Scholar] [CrossRef]
- Dachery, B.; Hernandes, K.C.; Veras, F.F.; Schmidt, L.; Augusti, P.R.; Manfroi, V.; Zini, C.A.; Welke, J.E. Effect of Aspergillus carbonarius on ochratoxin a levels, volatile profile and antioxidant activity of the grapes and respective wines. Food Res. Int. 2019, 126, 108687. [Google Scholar] [CrossRef]
- Handa, C.L.; de Lima, F.S.; Guelfi, M.F.G.; Fernandes, M.d.S.; Georgetti, S.R.; Ida, E.I. Parameters of the fermentation of soybean flour by Monascus purpureus or Aspergillus oryzae on the production of bioactive compounds and antioxidant activity. Food Chem. 2019, 271, 274–283. [Google Scholar] [CrossRef]
- Lee, J.H.; Hwang, S.-R.; Lee, Y.-H.; Kim, K.; Cho, K.M.; Lee, Y.B. Changes occurring in compositions and antioxidant properties of healthy soybean seeds [Glycine max (L.) Merr.] and soybean seeds diseased by Phomopsis longicolla and Cercospora kikuchii fungal pathogens. Food Chem. 2015, 185, 205–211. [Google Scholar] [CrossRef]
- Tkacz, K.; Chmielewska, J.; Turkiewicz, I.P.; Nowicka, P.; Wojdyło, A. Dynamics of changes in organic acids, sugars and phenolic compounds and antioxidant activity of sea buckthorn and sea buckthorn-apple juices during malolactic fermentation. Food Chem. 2020, 332, 127382. [Google Scholar] [CrossRef] [PubMed]
- Rizzello, C.G.; Lorusso, A.; Russo, V.; Pinto, D.; Marzani, B.; Gobbetti, M. Improving the antioxidant properties of quinoa flour through fermentation with selected autochthonous lactic acid bacteria. Int. J. Food Microbiol. 2017, 241, 252–261. [Google Scholar] [CrossRef]
- Li, S.; Jin, Z.; Hu, D.; Yang, W.; Yan, Y.; Nie, X.; Lin, J.; Zhang, Q.; Gai, D.; Ji, Y.; et al. Effect of solid-state fermentation with Lactobacillus casei on the nutritional value, isoflavones, phenolic acids and antioxidant activity of whole soybean flour. LWT Food Sci. Technol. 2020, 125, 109264. [Google Scholar] [CrossRef]
- Hashemi, S.M.B.; Jafarpour, D. Fermentation of bergamot juice with Lactobacillus plantarum strains in pure and mixed fermentations: Chemical composition, antioxidant activity and sensorial properties. LWT Food Sci. Technol. 2020, 131, 109803. [Google Scholar] [CrossRef]
- Wang, L.; Luo, Y.; Wu, Y.; Liu, Y.; Wu, Z. Fermentation and complex enzyme hydrolysis for improving the total soluble phenolic contents, flavonoid aglycones contents and bio-activities of guava leaves tea. Food Chem. 2018, 264, 189–198. [Google Scholar] [CrossRef]
- Zhang, J.; Deng, H.; Bai, J.; Zhou, X.; Zhao, Y.; Zhu, Y.; McClements, D.J.; Xiao, X.; Sun, Q. Health-promoting properties of barley: A review of nutrient and nutraceutical composition, functionality, bioprocessing, and health benefits. Crit. Rev. Food Sci. 2021. Early Access. [Google Scholar]
- Liu, L.; Zhang, R.; Deng, Y.; Zhang, Y.; Xiao, J.; Huang, F.; Wen, W.; Zhang, M. Fermentation and complex enzyme hydrolysis enhance total phenolics and antioxidant activity of aqueous solution from rice bran pretreated by steaming with α-amylase. Food Chem. 2017, 221, 636–643. [Google Scholar] [CrossRef]
- Huang, Q.; Zhang, H.; Xue, D. Enhancement of antioxidant activity of Radix Puerariae and red yeast rice by mixed fermentation with Monascus purpureus. Food Chem. 2017, 226, 89–94. [Google Scholar] [CrossRef]
- Zhao, Y.; Wu, C.; Zhu, Y.; Zhou, C.; Xiong, Z.; Samy Eweys, A.; Zhou, H.; Dong, Y.; Xiao, X. Metabolomics strategy for revealing the components in fermented barley extracts with Lactobacillus plantarum dy-1. Food Res. Int. 2021, 139, 109808. [Google Scholar] [CrossRef]
- Limón, R.I.; Peñas, E.; Torino, M.I.; Martínez-Villaluenga, C.; Dueñas, M.; Frias, J. Fermentation enhances the content of bioactive compounds in kidney bean extracts. Food Chem. 2015, 172, 343–352. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dueñas, M.; Fernández, D.; Hernández, T.; Estrella, I.; Muñoz, R. Bioactive phenolic compounds of cowpeas (Vigna sinensis L). Modifications by fermentation with natural microflora and with Lactobacillus plantarum ATCC 14917. J. Sci. Food Agr. 2005, 85, 297–304. [Google Scholar] [CrossRef]
- Starzyńska-Janiszewska, A.; Stodolak, B.; Mickowska, B. Effect of controlled lactic acid fermentation on selected bioactive and nutritional parameters of tempeh obtained from unhulled common bean (Phaseolus vulgaris) seeds. J. Sci. Food Agr. 2014, 94, 3068. [Google Scholar] [CrossRef]
- Crujeiras, A.B.; Parra, D.; Abete, I.; Martínez, J.A. A hypocaloric diet enriched in legumes specifically mitigates lipid peroxidation in obese subjects. Free Radic. Res. 2007, 41, 498–506. [Google Scholar] [CrossRef] [PubMed]
- Xiao, Y.; Xing, G.; Rui, X.; Li, W.; Chen, X.; Jiang, M.; Dong, M. Enhancement of the antioxidant capacity of chickpeas by solid state fermentation with Cordyceps militaris SN-18. J. Funct. Foods 2014, 10, 210–222. [Google Scholar] [CrossRef]
- Marazza, J.A.; Nazareno, M.A.; de Giori, G.S.; Garro, M.S. Enhancement of the antioxidant capacity of soymilk by fermentation with Lactobacillus rhamnosus. J. Funct. Foods 2012, 4, 594–601. [Google Scholar] [CrossRef]
- Yang, J.-H.; Mau, J.-L.; Ko, P.-T.; Huang, L.-C. Antioxidant properties of fermented soybean broth. Food Chem. 2000, 71, 249–254. [Google Scholar] [CrossRef]
- Lee, I.H.; Hung, Y.-H.; Chou, C.-C. Solid-state fermentation with fungi to enhance the antioxidative activity, total phenolic and anthocyanin contents of black bean. Int. J. Food Microbiol. 2008, 121, 150–156. [Google Scholar] [CrossRef] [PubMed]
- Sawicki, T.; Wiczkowski, W. The effects of boiling and fermentation on betalain profiles and antioxidant capacities of red beetroot products. Food Chem. 2018, 259, 292–303. [Google Scholar] [CrossRef] [PubMed]
- Wiczkowski, W.; Szawara-Nowak, D.; Topolska, J. Changes in the content and composition of anthocyanins in red cabbage and its antioxidant capacity during fermentation, storage and stewing. Food Chem. 2015, 167, 115–123. [Google Scholar] [CrossRef]
- Olennikov, D.N.; Kirillina, C.S.; Chirikova, N.K. Water-soluble melanoidin pigment as a new antioxidant component of fermented willowherb leaves (Epilobium angustifolium). Antioxidants 2021, 10, 1300. [Google Scholar] [CrossRef]
- Antognoni, F.; Mandrioli, R.; Potente, G.; Taneyo Saa, D.L.; Gianotti, A. Changes in carotenoids, phenolic acids and antioxidant capacity in bread wheat doughs fermented with different lactic acid bacteria strains. Food Chem. 2019, 292, 211–216. [Google Scholar] [CrossRef] [PubMed]
- Bei, Q.; Wu, Z.; Chen, G. Dynamic changes in the phenolic composition and antioxidant activity of oats during simultaneous hydrolysis and fermentation. Food Chem. 2020, 305, 125269. [Google Scholar] [CrossRef]
- Makkar, H.P.S.; Sidhuraju, P.; Becker, K. Plant Secondary Metabolites; Humana Press: Totowa, NJ, USA, 2007. [Google Scholar]
- Zieliński, H.; Szawara-Nowak, D.; Bączek, N.; Wronkowska, M. Effect of liquid-state fermentation on the antioxidant and functional properties of raw and roasted buck wheat flours. Food Chem. 2019, 271, 291–297. [Google Scholar] [CrossRef]
- Chung, Y.; Park, J.-Y.; Lee, J.-E.; Kim, K.-T.; Paik, H.-D. Antioxidant Activity and Inhibitory Effect on Nitric Oxide Production of Hydroponic Ginseng Fermented with Lactococcus lactis KC24. Antioxidants 2021, 10, 1614. [Google Scholar] [CrossRef] [PubMed]
- Mapelli-Brahm, P.; Barba, F.J.; Remize, F.; Garcia, C.; Fessard, A.; Mousavi Khaneghah, A.; Sant’Ana, A.S.; Lorenzo, J.M.; Montesano, D.; Meléndez-Martínez, A.J. The impact of fermentation processes on the production, retention and bioavailability of carotenoids: An overview. Trends Food Sci. Tech. 2020, 99, 389–401. [Google Scholar] [CrossRef]
- Oloo, B.O.; Shitandi, A.A.; Mahungu, S.; Malinga, J.; Ogata, R. Effects of lactic acid fermentation on the retention of β-carotene content in orange fleshed sweet potatoes. Int. J. Food Stud. 2014, 3, 13–33. [Google Scholar] [CrossRef]
- Hubert, J.; Berger, M.; Nepveu, F.; Paul, F.; Daydé, J. Effects of fermentation on the phytochemical composition and antioxidant properties of soy germ. Food Chem. 2008, 109, 709–721. [Google Scholar] [CrossRef]
- Adeyeye, E. The effect of fermentation on the dietary quality of lipids from African locust bean (Parkia biglobosa) seeds. Elixir Food Sci. 2013, 58, 14912–14922. [Google Scholar]
- Chen, L.; Hui, Y.; Gao, T.; Shu, G.; Chen, H. Function and characterization of novel antioxidant peptides by fermentation with a wild Lactobacillus plantarum 60. LWT Food Sci. Technol. 2021, 135, 110162. [Google Scholar] [CrossRef]
- Muhialdin, B.J.; Abdul Rani, N.F.; Meor Hussin, A.S. Identification of antioxidant and antibacterial activities for the bioactive peptides generated from bitter beans (Parkia speciosa) via boiling and fermentation processes. LWT Food Sci. Technol. 2020, 131, 109776. [Google Scholar] [CrossRef]
- Miao, J.; Shi, W.; Zhang, J.; Zhang, X.; Zhang, H.; Wang, Z.; Qiu, J. Response surface methodology for the fermentation of polysaccharides from Auricularia auricula using Trichoderma viride and their antioxidant activities. Int. J. Biol. Macromol. 2020, 155, 393–402. [Google Scholar] [CrossRef] [PubMed]
- He, R.; Ju, X.; Yuan, J.; Wang, L.; Girgih, A.T.; Aluko, R.E. Antioxidant activities of rapeseed peptides produced by solid state fermentation. Food Res. Int. 2012, 49, 432–438. [Google Scholar] [CrossRef]
- Liu, X.; Hou, R.; Xu, K.; Chen, L.; Wu, X.; Lin, W.; Zheng, M.; Fu, J. Extraction, characterization and antioxidant activity analysis of the polysaccharide from the solid-state fermentation substrate of Inonotus hispidus. Int. J. Biol. Macromol. 2019, 123, 468–476. [Google Scholar] [CrossRef] [PubMed]
- Cho, K.M.; Lee, J.H.; Yun, H.D.; Ahn, B.Y.; Kim, H.; Seo, W.T. Changes of phytochemical constituents (isoflavones, flavanols, and phenolic acids) during cheonggukjang soybeans fermentation using potential probiotics Bacillus subtilis CS90. J. Food Compos. Anal. 2011, 24, 402–410. [Google Scholar] [CrossRef]
- Koistinen, V.M.; Katina, K.; Nordlund, E.; Poutanen, K.; Hanhineva, K. Changes in the phytochemical profile of rye bran induced by enzymatic bioprocessing and sourdough fermentation. Food Res. Int. 2016, 89, 1106–1115. [Google Scholar] [CrossRef]
- Sheih, I.C.; Fang, T.J.; Wu, T.-K.; Chen, R.-Y. Effects of fermentation on antioxidant properties and phytochemical composition of soy germ. J. Sci. Food Agr. 2014, 94, 3163–3170. [Google Scholar] [CrossRef]
- Rodríguez, H.; Landete, J.M.; de las Rivas, B.; Muñoz, R. Metabolism of food phenolic acids by Lactobacillus plantarum CECT 748T. Food Chem. 2008, 107, 1393–1398. [Google Scholar] [CrossRef] [Green Version]
- Rodríguez, H.; Curiel, J.A.; Landete, J.M.; de las Rivas, B.; de Felipe, F.L.; Gómez-Cordovés, C.; Mancheño, J.M.; Muñoz, R. Food phenolics and lactic acid bacteria. Int. J. Food Microbiol. 2009, 132, 79–90. [Google Scholar] [CrossRef] [Green Version]
- Acin-Albiac, M.; Filannino, P.; Arora, K.; Da Ros, A.; Gobbetti, M.; Di Cagno, R. Role of lactic acid bacteria phospho-β-glucosidases during the fermentation of cereal by-products. Foods. 2021, 10, 97. [Google Scholar] [CrossRef]
- de Araújo, J.A.; Ferreira, N.R.; da Silva, S.H.M.; Oliveira, G.; Monteiro, R.C.; Alves, Y.F.M.; Lopes, A.S. Filamentous fungi diversity in the natural fermentation of Amazonian cocoa beans and the microbial enzyme activities. Ann. Microbiol. 2019, 69, 975–987. [Google Scholar] [CrossRef]
- Costa, R.d.S.; de Almeida, S.S.; Cavalcanti, E.d.A.C.; Freire, D.M.G.; Moura-Nunes, N.; Monteiro, M.; Perrone, D. Enzymes produced by solid state fermentation of agro-industrial by-products release ferulic acid in bioprocessed whole-wheat breads. Food Res. Int. 2021, 140, 109843. [Google Scholar] [CrossRef] [PubMed]
- Yao, Y.; Wu, M.; Huang, Y.; Li, C.; Pan, X.; Zhu, W.; Huang, Y. Appropriately raising fermentation temperature beneficial to the increase of antioxidant activity and gallic acid content in Eurotium cristatum-fermented loose tea. LWT Food Sci. Technol. 2017, 82, 248–254. [Google Scholar] [CrossRef]
- Nguyen, T.T.T.; Loiseau, G.; Icard-Vernière, C.; Rochette, I.; Trèche, S.; Guyot, J.-P. Effect of fermentation by amylolytic lactic acid bacteria, in process combinations, on characteristics of rice/soybean slurries: A new method for preparing high energy density complementary foods for young children. Food Chem. 2007, 100, 623–631. [Google Scholar] [CrossRef]
- Darwesh, O.M.; El-Latief, A.H.A.; Abuarab, M.E.; Kasem, M.A. Enhancing the efficiency of some agricultural wastes as low-cost absorbents to remove textile dyes from their contaminated solutions. Biomass Convers. Biorefinery. 2021, 1–10. [Google Scholar] [CrossRef]
- Manan, M.; Webb, C. Design aspects of solid state fermentation as applied to microbial bioprocessing. J. Appl. Biotechnol. Bioeng. 2017, 4, 1–25. [Google Scholar]


{kind=link}
| Food Material | The Used Microorganism | Effect on Bioactive Compounds of Plant-Based Food Material | Fermentation Type | Literature |
|---|---|---|---|---|
| Wheat | Lactobacillus fermentum (MR13), L. rhamnosus (C249, C1272), L. plantarum (LB102, LB124, LB126, LB245, 29DAN, 83DAN, 6BHI, 98A) and L. brevis (3BHI) |
| Liquid and solid | [50] |
| Oats | Monascus anka GIM 3.592 |
| Liquid and solid | [51] |
| Goat milk | Lactobacillus plantarum 60 |
| Liquid | [52] |
| Buckwheat, wheat germ, barley, and rye | Lactobacillus rhamnosus, and Saccharomyces cerevisiae |
| Liquid and solid | [15] |
| Defatted soybean flour | Monascus purpureus or Aspergillus oryzae |
| Solid | [28] |
| Bergamot juice | Pure and mixed cultures of L. plantarum subsp. plantarum PTCC 1896, L. plantarum AF1 and L. plantarum LP3 |
| Liquid | [33] |
| Whole soybean flour | Lactobacillus casei |
| Liquid and solid | [32] |
| Kidney beans | Solid-state fermentation was carried out by Bacillus subtilis, whilst liquid state fermentation was performed either by natural fermentation (NF) or by Lactobacillus plantarum strain (LPF) |
| Liquid and solid | [39] |
| Rice bran | It steamed with α-amylase, fermented with lactic acid bacteria, and hydrolyzed with complex enzymes |
| Liquid and solid | [36] |
| Cocoa beans | 13 naturally existing yeast strains |
| Liquid and solid | [17] |
| Elderberry juices | Ten strains of Lactobacillus |
| Liquid | [14] |
| Sea buckthornberries | Lactobacillus plantarum, Lactobacillus plantarum subsp. argentoratensis and Oenococcus oeni strains |
| Liquid | [30] |
| Guava leaves tea | It first fermented with Monascus anka and Saccharomyces cerevisiae and then hydrolyzed with complex enzymes |
| Liquid and solid | [34] |
| Chickpeas | Cordyceps militaris SN-18 |
| Solid | [43] |
| Buckwheat flours | Selected lactic acid bacteria (LAB) and Rhizopus oligosporus fungi |
| Liquid | [53] |
| Factor for Comparison | Solid-State Fermentation | Liquid Fermentation |
|---|---|---|
| Advantages |
|
|
| Limitations |
|
|
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhao, Y.-S.; Eweys, A.S.; Zhang, J.-Y.; Zhu, Y.; Bai, J.; Darwesh, O.M.; Zhang, H.-B.; Xiao, X. Fermentation Affects the Antioxidant Activity of Plant-Based Food Material through the Release and Production of Bioactive Components. Antioxidants 2021, 10, 2004. https://doi.org/10.3390/antiox10122004
Zhao Y-S, Eweys AS, Zhang J-Y, Zhu Y, Bai J, Darwesh OM, Zhang H-B, Xiao X. Fermentation Affects the Antioxidant Activity of Plant-Based Food Material through the Release and Production of Bioactive Components. Antioxidants. 2021; 10(12):2004. https://doi.org/10.3390/antiox10122004
Chicago/Turabian StyleZhao, Yan-Sheng, Aya Samy Eweys, Jia-Yan Zhang, Ying Zhu, Juan Bai, Osama M. Darwesh, Hai-Bo Zhang, and Xiang Xiao. 2021. "Fermentation Affects the Antioxidant Activity of Plant-Based Food Material through the Release and Production of Bioactive Components" Antioxidants 10, no. 12: 2004. https://doi.org/10.3390/antiox10122004
APA StyleZhao, Y. -S., Eweys, A. S., Zhang, J. -Y., Zhu, Y., Bai, J., Darwesh, O. M., Zhang, H. -B., & Xiao, X. (2021). Fermentation Affects the Antioxidant Activity of Plant-Based Food Material through the Release and Production of Bioactive Components. Antioxidants, 10(12), 2004. https://doi.org/10.3390/antiox10122004