
NAD(P)H Drives the Ascorbate–Glutathione Cycle and Abundance of Catalase in Developing Beech Seeds Differently in Embryonic Axes and Cotyledons



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Material
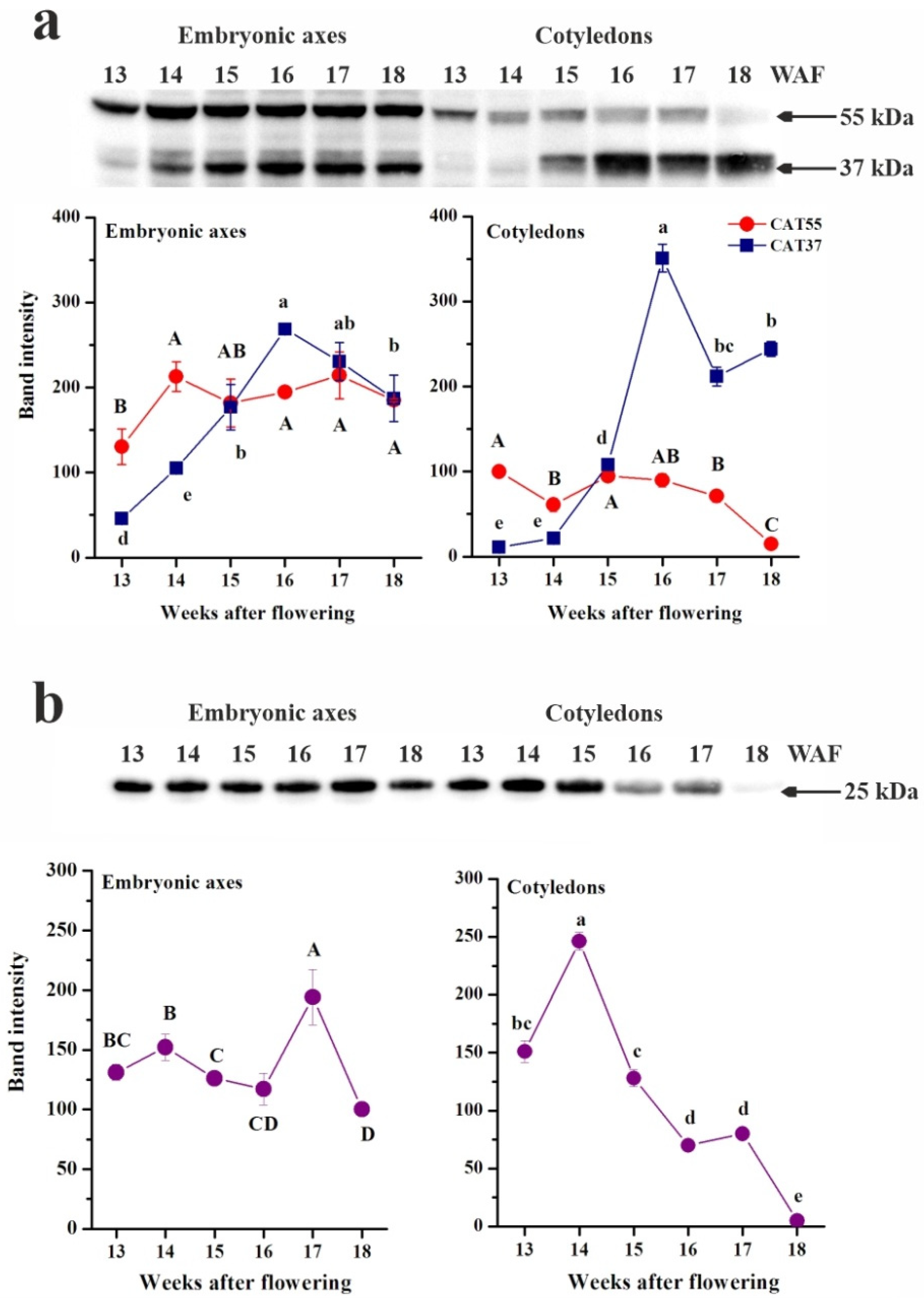
2.2. Protein Extraction, Electrophoresis and Western Blot Analysis
2.3. Determination of the Contents of Redox Forms
2.3.1. Determination of Ascorbate
2.3.2. Determination of Glutathione
2.3.3. Determination of NAD(P)
2.3.4. Determination of NAD(P)-Originated Physiological Indices
2.4. Statistical Analyses
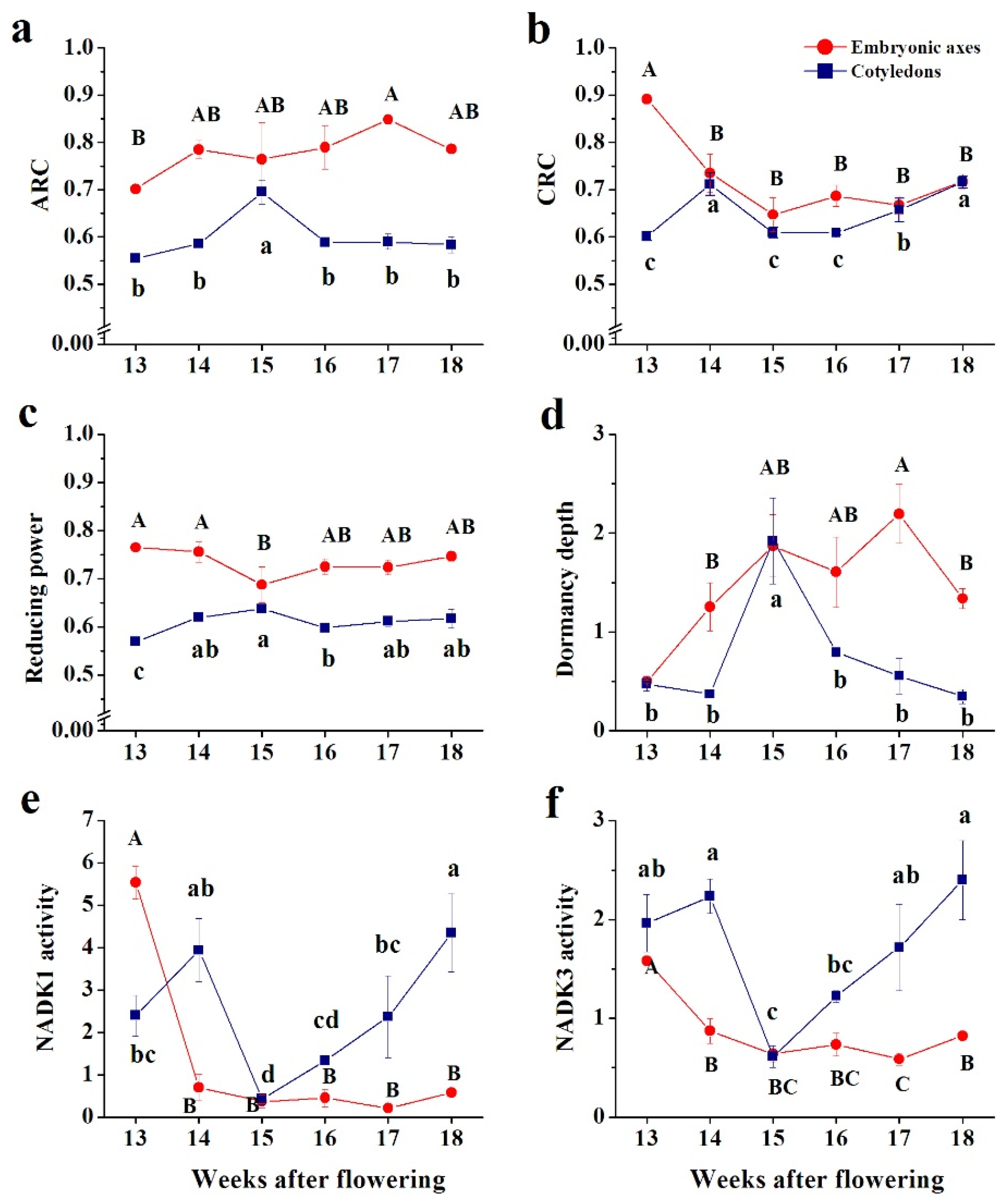
3. Results
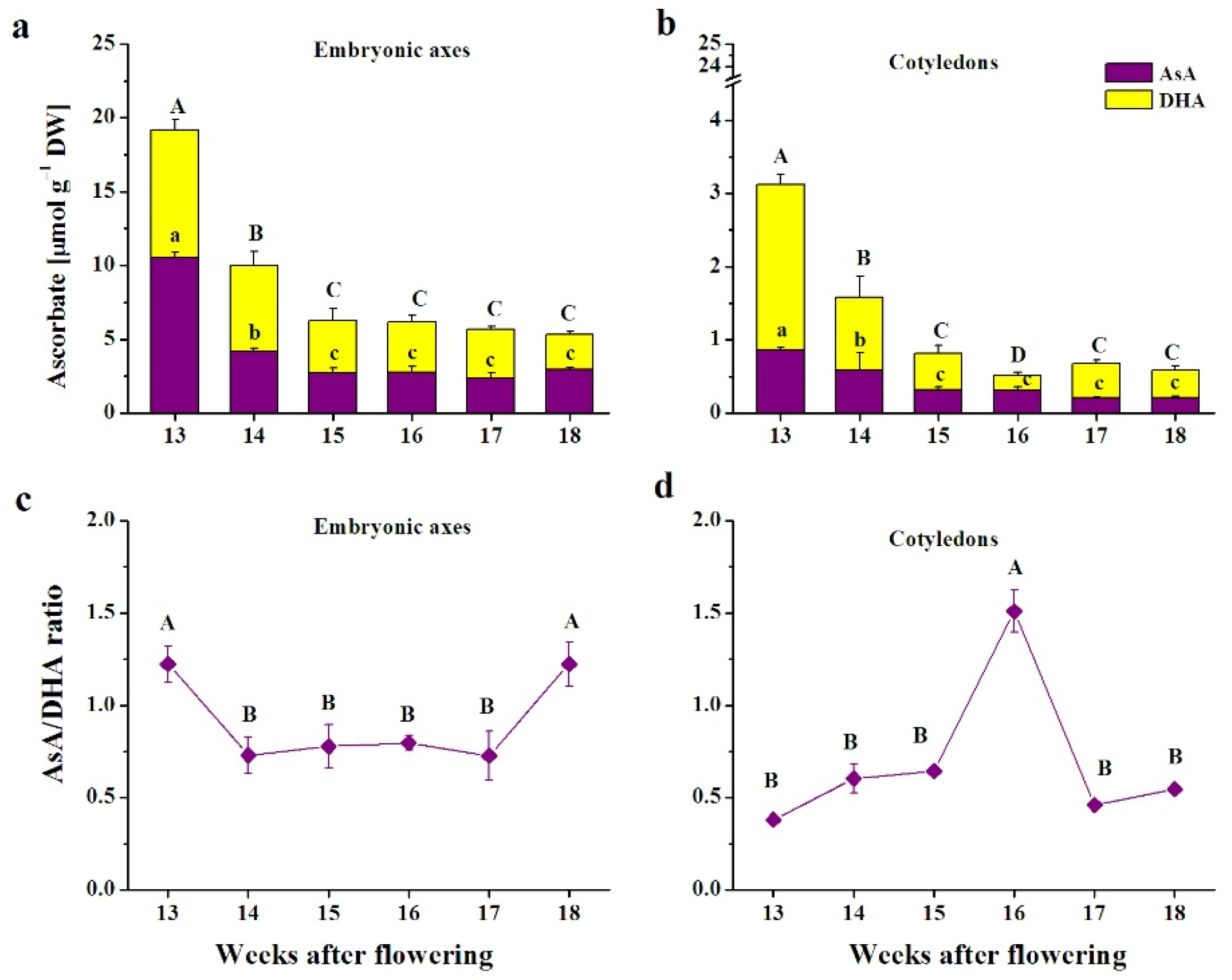
3.1. Ascorbate
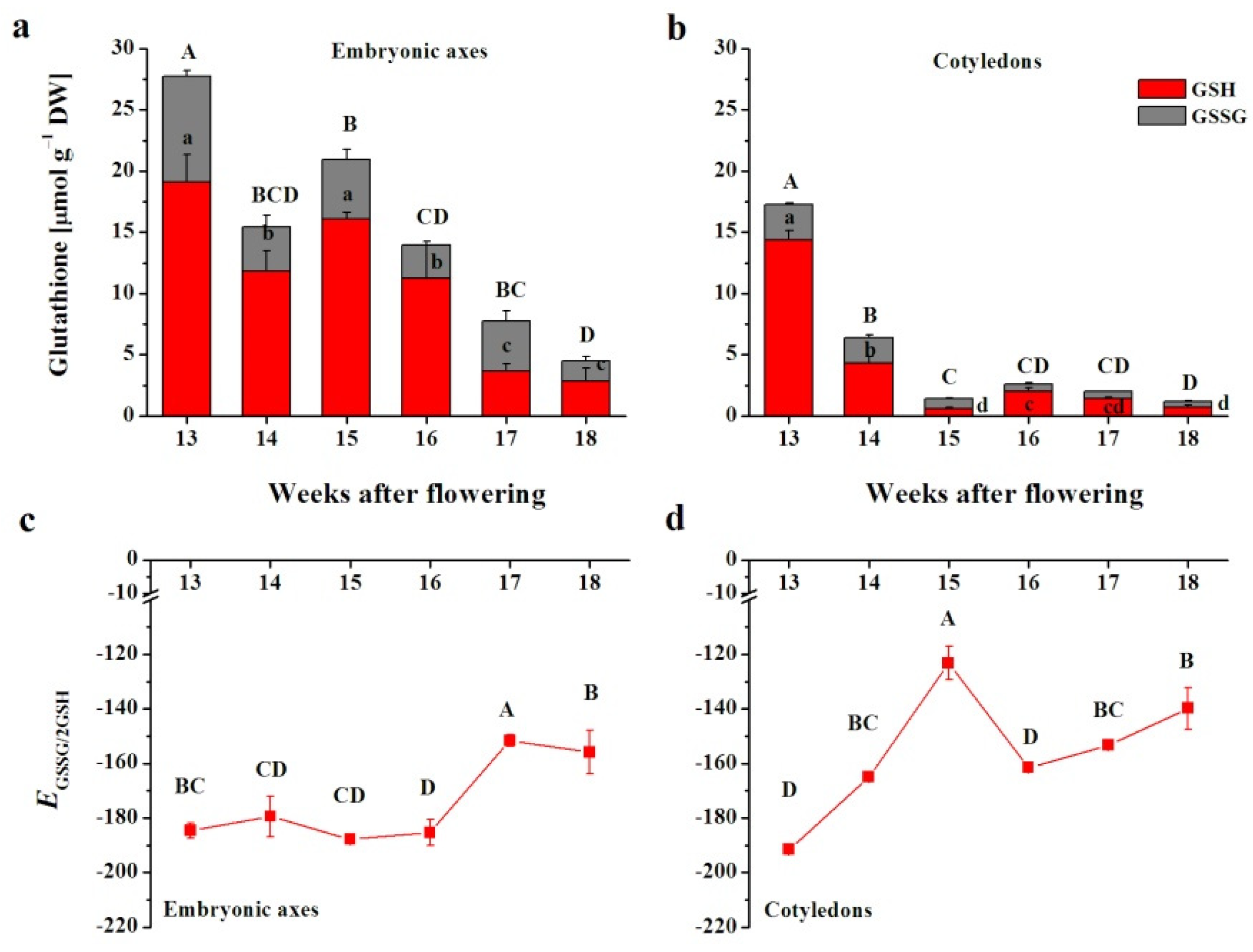
3.2. Glutathione
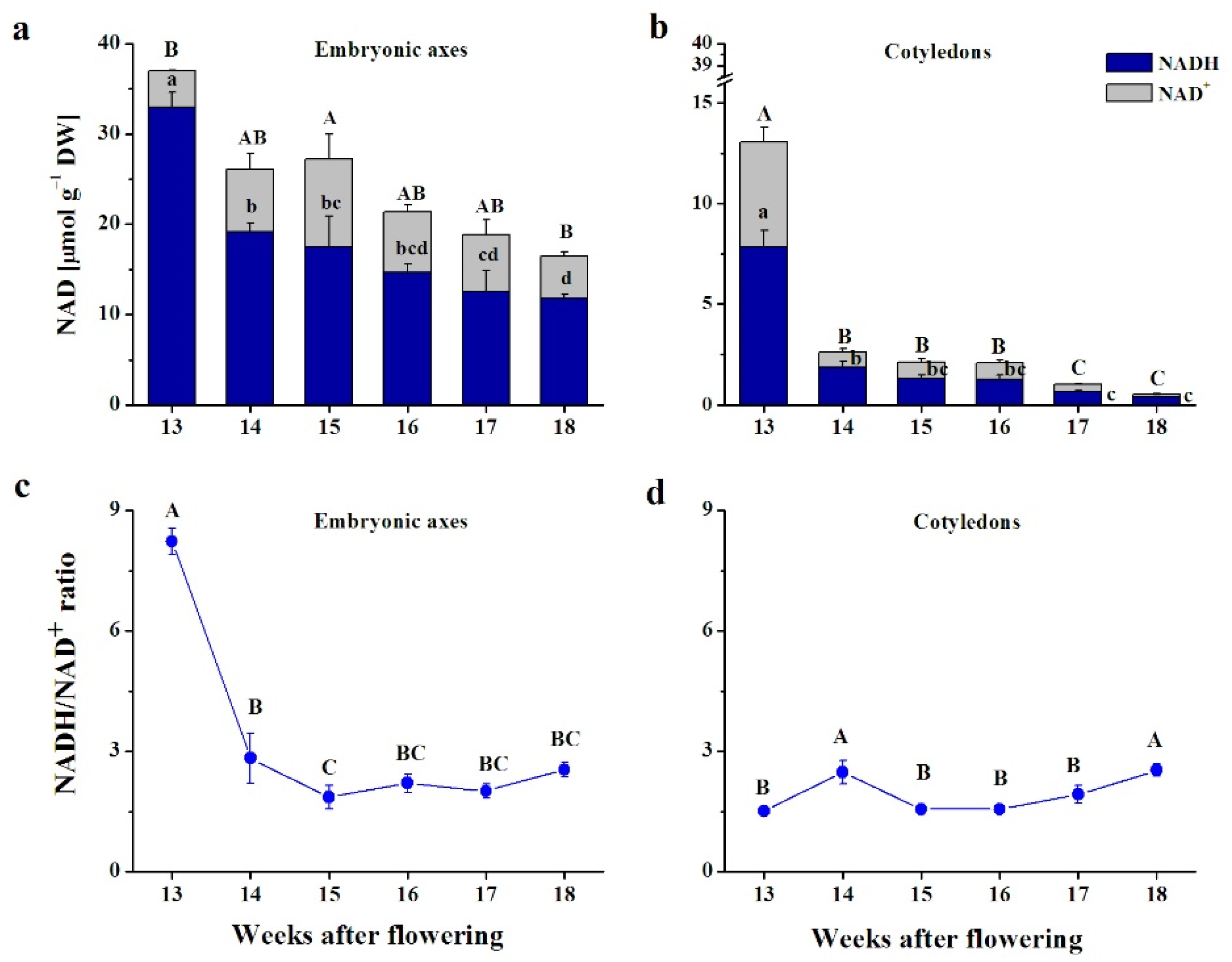
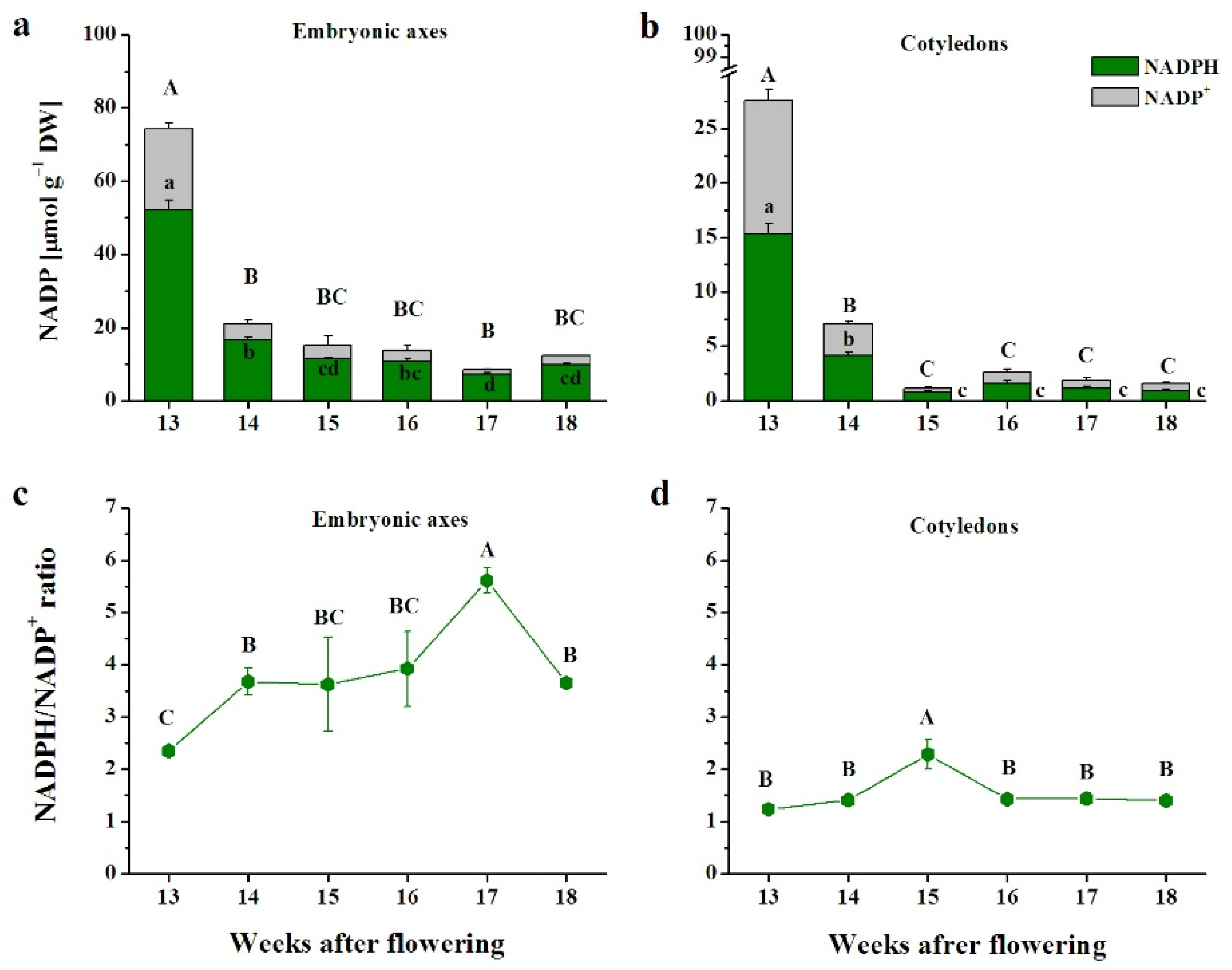
3.3. Pyridine Nucleotides
3.4. Enzymes Involved in H2O2 Removal
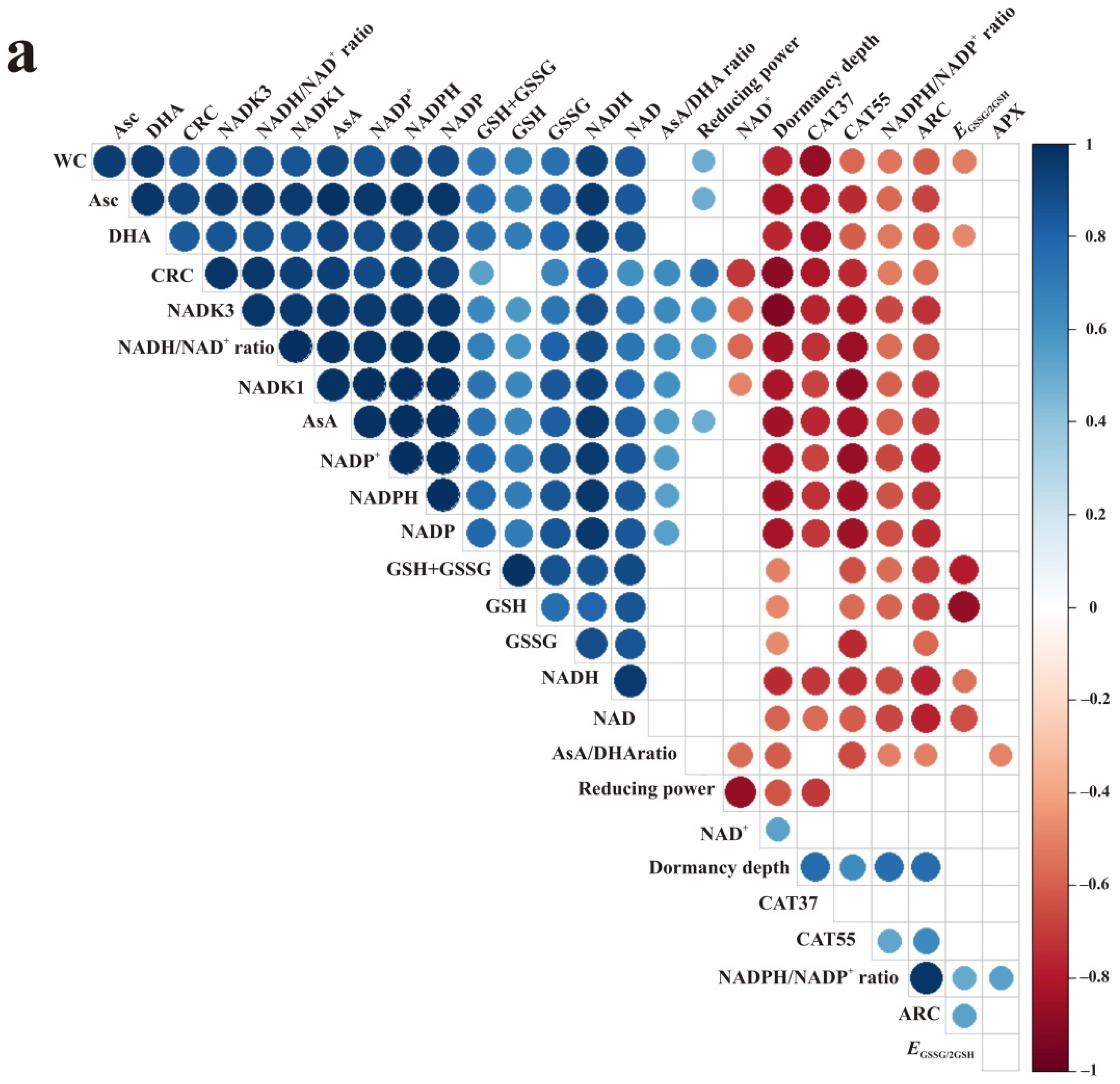
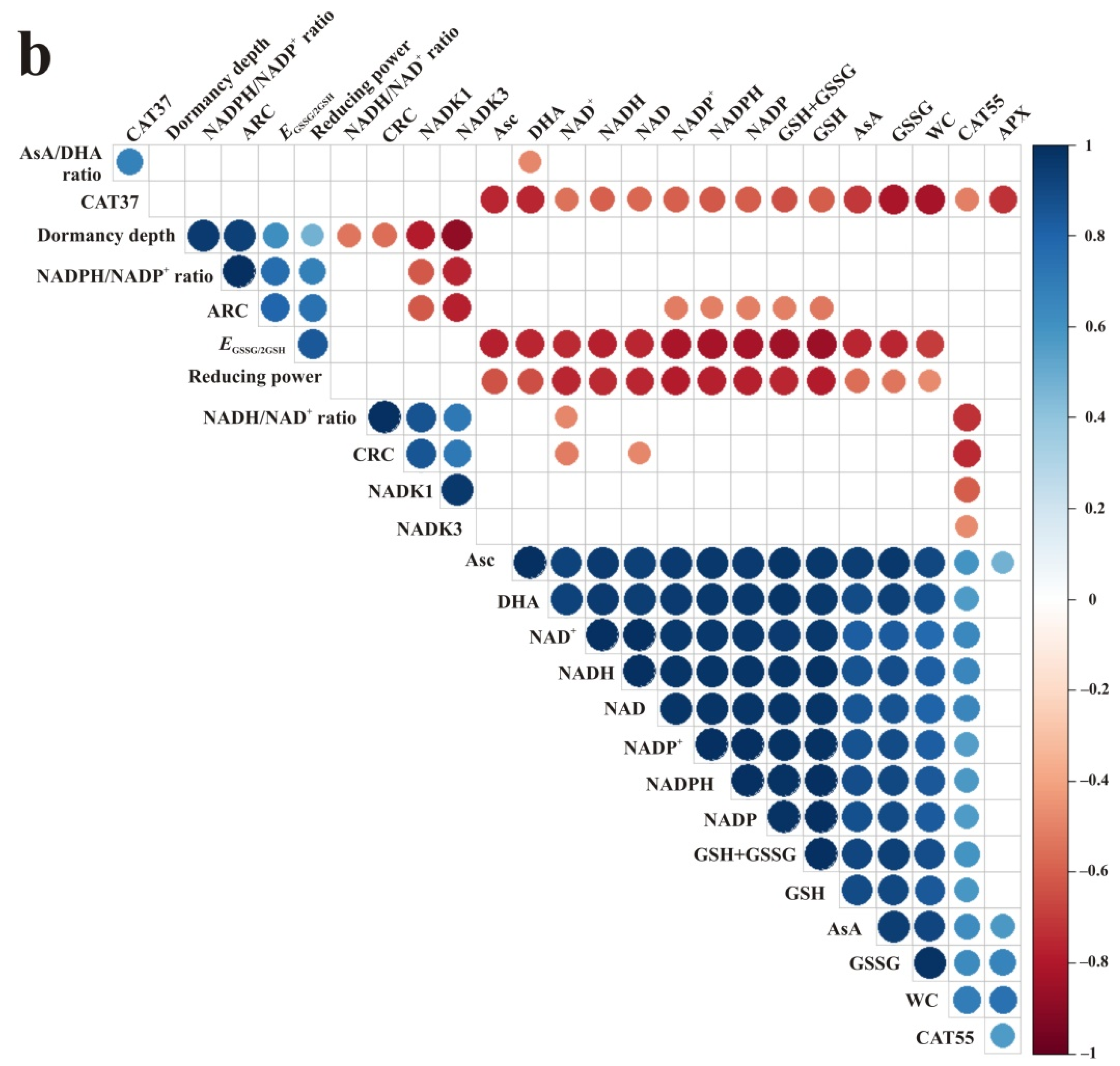
3.5. Correlations
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Packham, J.R.; Thomas, P.A.; Atkinson, M.D.; Degen, T. Biological Flora of the British Isles: Fagus sylvatica. J. Ecol. 2012, 100, 1557–1608. [Google Scholar] [CrossRef]
- Suszka, B.; Muller, C.; Bonnet-Masimbert, M.; Gordon, A. Seeds of Forest Broadleaves: From Harvest to Sowing; Institute National de la Recherche Agronomique: Paris, France, 1996; ISBN 978-2-7380-0659-2. [Google Scholar]
- Bogdziewicz, M.; Szymkowiak, J.; Kasprzyk, I.; Grewling, Ł.; Borowski, Z.; Borycka, K.; Kantorowicz, W.; Myszkowska, D.; Piotrowicz, K.; Ziemianin, M.; et al. Masting in Wind-Pollinated Trees: System-Specific Roles of Weather and Pollination Dynamics in Driving Seed Production. Ecology 2017, 98, 2615–2625. [Google Scholar] [CrossRef]
- Drobyshev, I.; Övergaard, R.; Saygin, I.; Niklasson, M.; Hickler, T.; Karlsson, M.; Sykes, M.T. Masting Behaviour and Dendrochronology of European Beech (Fagus sylvatica L.) in Southern Sweden. For. Ecol. Manag. 2010, 259, 2160–2171. [Google Scholar] [CrossRef]
- Augustaitis, A.; Jasineviciene, D.; Girgzdiene, R.; Kliucius, A.; Marozas, V. Sensitivity of Beech Trees to Global Environmental Changes at Most North-Eastern Latitude of Their Occurrence in Europe. Sci. World J. 2012, 2012, 743926. [Google Scholar] [CrossRef] [Green Version]
- Growth and Quality of Fagus sylvatica Saplings Depend on Seed Source, Site, and Browsing Intensity—Frank—2019—Ecosphere—Wiley Online Library. Available online: https://esajournals.onlinelibrary.wiley.com/doi/full/10.1002/ecs2.2580 (accessed on 5 November 2021).
- Bogdziewicz, M.; Kelly, D.; Thomas, P.A.; Lageard, J.G.A.; Hacket-Pain, A. Climate Warming Disrupts Mast Seeding and Its Fitness Benefits in European Beech. Nat. Plants 2020, 6, 88–94. [Google Scholar] [CrossRef]
- Kalemba, E.M.; Janowiak, F.; Pukacka, S. Desiccation Tolerance Acquisition in Developing Beech (Fagus sylvatica L.) Seeds: The Contribution of Dehydrin-like Protein. Trees 2008, 23, 305. [Google Scholar] [CrossRef]
- León-Lobos, P.; Ellis, R.H. Seed Storage Behaviour of Fagus sylvatica and Fagus crenata. Seed Sci. Res. 2002, 12, 31–37. [Google Scholar] [CrossRef]
- Goldberg, R.B.; de Paiva, G.; Yadegari, R. Plant Embryogenesis: Zygote to Seed. Science 1994, 266, 605–614. [Google Scholar] [CrossRef] [PubMed]
- Baud, S.; Dubreucq, B.; Miquel, M.; Rochat, C.; Lepiniec, L. Storage Reserve Accumulation in Arabidopsis: Metabolic and Developmental Control of Seed Filling. Arab. Book 2008, 6, e0113. [Google Scholar] [CrossRef] [Green Version]
- Leprince, O.; Pellizzaro, A.; Berriri, S.; Buitink, J. Late Seed Maturation: Drying without Dying. J. Exp. Bot. 2017, 68, 827–841. [Google Scholar] [CrossRef] [Green Version]
- Pereira Lima, J.J.; Buitink, J.; Lalanne, D.; Rossi, R.F.; Pelletier, S.; da Silva, E.A.A.; Leprince, O. Molecular Characterization of the Acquisition of Longevity during Seed Maturation in Soybean. PLoS ONE 2017, 12, e0180282. [Google Scholar] [CrossRef] [PubMed]
- Ellis, R.H. Temporal Patterns of Seed Quality Development, Decline, and Timing of Maximum Quality during Seed Development and Maturation. Seed Sci. Res. 2019, 29, 135–142. [Google Scholar] [CrossRef] [Green Version]
- Dante, R.A.; Larkins, B.A.; Sabelli, P.A. Cell Cycle Control and Seed Development. Front. Plant Sci. 2014, 5, 493. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sehgal, A.; Sita, K.; Siddique, K.H.M.; Kumar, R.; Bhogireddy, S.; Varshney, R.K.; HanumanthaRao, B.; Nair, R.M.; Prasad, P.V.V.; Nayyar, H. Drought or/and Heat-Stress Effects on Seed Filling in Food Crops: Impacts on Functional Biochemistry, Seed Yields, and Nutritional Quality. Front. Plant Sci. 2018, 9, 1705. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sano, N.; Rajjou, L.; North, H.M.; Debeaujon, I.; Marion-Poll, A.; Seo, M. Staying Alive: Molecular Aspects of Seed Longevity. Plant Cell Physiol. 2016, 57, 660–674. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ratajczak, E.; Staszak, A.M.; Wojciechowska, N.; Bagniewska-Zadworna, A.; Dietz, K.J. Regulation of Thiol Metabolism as a Factor That Influences the Development and Storage Capacity of Beech Seeds. J. Plant Physiol. 2019, 239, 61–70. [Google Scholar] [CrossRef]
- Meyer, A.J.; Dreyer, A.; Ugalde, J.M.; Feitosa-Araujo, E.; Dietz, K.-J.; Schwarzländer, M. Shifting Paradigms and Novel Players in Cys-Based Redox Regulation and ROS Signaling in Plants-and Where to Go Next. Biol. Chem. 2021, 402, 399–423. [Google Scholar] [CrossRef]
- de Simone, A.; Hubbard, R.; de la Torre, N.V.; Velappan, Y.; Wilson, M.; Considine, M.J.; Soppe, W.J.J.; Foyer, C.H. Redox Changes During the Cell Cycle in the Embryonic Root Meristem of Arabidopsis thaliana. Antioxid. Redox Signal. 2017, 27, 1505–1519. [Google Scholar] [CrossRef] [Green Version]
- Noctor, G.; Reichheld, J.-P.; Foyer, C.H. ROS-Related Redox Regulation and Signaling in Plants. Semin. Cell Dev. Biol. 2018, 80, 3–12. [Google Scholar] [CrossRef] [Green Version]
- Stolarska, E.; Bilska, K.; Wojciechowska, N.; Bagniewska-Zadworna, A.; Rey, P.; Kalemba, E.M. Integration of MsrB1 and MsrB2 in the Redox Network during the Development of Orthodox and Recalcitrant Acer Seeds. Antioxidants 2020, 9, 1250. [Google Scholar] [CrossRef]
- Alipour, S.; Wojciechowska, N.; Stolarska, E.; Bilska, K.; Kalemba, E.M. NAD(P)-Driven Redox Status Contributes to Desiccation Tolerance in Acer Seeds. Plant Cell Physiol. 2020, 61, 1158–1167. [Google Scholar] [CrossRef] [PubMed]
- Wojciechowska, N.; Alipour, S.; Stolarska, E.; Bilska, K.; Rey, P.; Kalemba, E.M. Peptide-Bound Methionine Sulfoxide (MetO) Levels and MsrB2 Abundance Are Differentially Regulated during the Desiccation Phase in Contrasted Acer Seeds. Antioxidants 2020, 9, 391. [Google Scholar] [CrossRef] [PubMed]
- Alipour, S.; Bilska, K.; Stolarska, E.; Wojciechowska, N.; Kalemba, E.M. Nicotinamide Adenine Dinucleotides Are Associated with Distinct Redox Control of Germination in Acer Seeds with Contrasting Physiology. PLoS ONE 2021, 16, e0245635. [Google Scholar] [CrossRef] [PubMed]
- Wojciechowska, N.; Alipour, S.; Stolarska, E.; Bilska, K.; Rey, P.; Kalemba, E.M. Involvement of the MetO/Msr System in Two Acer Species That Display Contrasting Characteristics during Germination. Int. J. Mol. Sci. 2020, 21, 9197. [Google Scholar] [CrossRef]
- Considine, M.J.; Foyer, C.H. Redox Regulation of Plant Development. Antioxid. Redox Signal. 2014, 21, 1305–1326. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kapoor, D.; Singh, S.; Kumar, V.; Romero, R.; Prasad, R.; Singh, J. Antioxidant Enzymes Regulation in Plants in Reference to Reactive Oxygen Species (ROS) and Reactive Nitrogen Species (RNS). Plant Gene 2019, 19, 100182. [Google Scholar] [CrossRef]
- Noctor, G.; Foyer, C.H. ASCORBATE AND GLUTATHIONE: Keeping Active Oxygen Under Control. Annu. Rev. Plant Physiol. Plant Mol. Biol. 1998, 49, 249–279. [Google Scholar] [CrossRef] [PubMed]
- Foyer, C.H.; Noctor, G. Ascorbate and Glutathione: The Heart of the Redox Hub. Plant Physiol. 2011, 155, 2–18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hasanuzzaman, M.; Bhuyan, M.H.M.B.; Anee, T.I.; Parvin, K.; Nahar, K.; Mahmud, J.A.; Fujita, M. Regulation of Ascorbate-Glutathione Pathway in Mitigating Oxidative Damage in Plants under Abiotic Stress. Antioxidants 2019, 8, 384. [Google Scholar] [CrossRef] [Green Version]
- Cairns, N.G.; Pasternak, M.; Wachter, A.; Cobbett, C.S.; Meyer, A.J. Maturation of Arabidopsis Seeds Is Dependent on Glutathione Biosynthesis within the Embryo. Plant Physiol. 2006, 141, 446–455. [Google Scholar] [CrossRef] [Green Version]
- Pukacka, S.; Ratajczak, E. Ascorbate and Glutathione Metabolism during Development and Desiccation of Orthodox and Recalcitrant Seeds of the Genus Acer. Funct. Plant Biol. FPB 2007, 34, 601–613. [Google Scholar] [CrossRef]
- Tommasi, F.; Paciolla, C.; de Pinto, M.C.; De Gara, L. A Comparative Study of Glutathione and Ascorbate Metabolism during Germination of Pinus pinea L. Seeds. J. Exp. Bot. 2001, 52, 1647–1654. [Google Scholar] [CrossRef] [Green Version]
- Gerna, D.; Roach, T.; Stöggl, W.; Wagner, J.; Vaccino, P.; Limonta, M.; Kranner, I. Changes in Low-Molecular-Weight Thiol-Disulphide Redox Couples Are Part of Bread Wheat Seed Germination and Early Seedling Growth. Free Radic. Res. 2017, 51, 568–581. [Google Scholar] [CrossRef] [Green Version]
- Kranner, I.; Birtić, S.; Anderson, K.M.; Pritchard, H.W. Glutathione Half-Cell Reduction Potential: A Universal Stress Marker and Modulator of Programmed Cell Death? Free Radic. Biol. Med. 2006, 40, 2155–2165. [Google Scholar] [CrossRef] [PubMed]
- Kocsy, G. Die or Survive? Redox Changes as Seed Viability Markers. Plant Cell Environ. 2015, 38, 1008–1010. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xia, F.; Cheng, H.; Chen, L.; Zhu, H.; Mao, P.; Wang, M. Influence of Exogenous Ascorbic Acid and Glutathione Priming on Mitochondrial Structural and Functional Systems to Alleviate Aging Damage in Oat Seeds. BMC Plant Biol. 2020, 20, 104. [Google Scholar] [CrossRef] [Green Version]
- Arrigoni, O.; De Tullio, M.C. Ascorbic Acid: Much More than Just an Antioxidant. Biochim. Biophys. Acta 2002, 1569, 1–9. [Google Scholar] [CrossRef]
- Bilska, K.; Wojciechowska, N.; Alipour, S.; Kalemba, E.M. Ascorbic Acid-The Little-Known Antioxidant in Woody Plants. Antioxidants 2019, 8, 645. [Google Scholar] [CrossRef] [Green Version]
- Hoang, M.T.T.; Doan, M.T.A.; Nguyen, T.; Tra, D.-P.; Chu, T.N.; Dang, T.P.T.; Quach, P.N.D. Phenotypic Characterization of Arabidopsis Ascorbate and Glutathione Deficient Mutants under Abiotic Stresses. Agronomy 2021, 11, 764. [Google Scholar] [CrossRef]
- Hunt, L.; Lerner, F.; Ziegler, M. NAD—New Roles in Signalling and Gene Regulation in Plants. New Phytol. 2004, 163, 31–44. [Google Scholar] [CrossRef]
- Mahalingam, R. Pyridine Nucleotide Homeostasis in Plant Development and Stress. Int. J. Plant Dev. Biol. 2007, 1, 194–201. [Google Scholar]
- Pétriacq, P.; de Bont, L.; Tcherkez, G.; Gakière, B. NAD: Not Just a Pawn on the Board of Plant-Pathogen Interactions. Plant Signal. Behav. 2013, 8, e22477. [Google Scholar] [CrossRef] [Green Version]
- Gakière, B.; Hao, J.; de Bont, L.; Pétriacq, P.; Nunes-Nesi, A.; Fernie, A.R. NAD+ Biosynthesis and Signaling in Plants. Crit. Rev. Plant Sci. 2018, 37, 259–307. [Google Scholar] [CrossRef]
- Xia, X.-J.; Zhou, Y.-H.; Shi, K.; Zhou, J.; Foyer, C.H.; Yu, J.-Q. Interplay between Reactive Oxygen Species and Hormones in the Control of Plant Development and Stress Tolerance. J. Exp. Bot. 2015, 66, 2839–2856. [Google Scholar] [CrossRef] [Green Version]
- Queval, G.; Noctor, G. A Plate Reader Method for the Measurement of NAD, NADP, Glutathione, and Ascorbate in Tissue Extracts: Application to Redox Profiling during Arabidopsis Rosette Development. Anal. Biochem. 2007, 363, 58–69. [Google Scholar] [CrossRef]
- Hashida, S.; Takahashi, H.; Takahara, K.; Kawai-Yamada, M.; Kitazaki, K.; Shoji, K.; Goto, F.; Yoshihara, T.; Uchimiya, H. NAD+ Accumulation during Pollen Maturation in Arabidopsis Regulating Onset of Germination. Mol. Plant 2013, 6, 216–225. [Google Scholar] [CrossRef] [Green Version]
- Sghaier-Hammami, B.; BM Hammami, S.; Baazaoui, N.; Gómez-Díaz, C.; Jorrín-Novo, J.V. Dissecting the Seed Maturation and Germination Processes in the Non-Orthodox Quercus ilex Species Based on Protein Signatures as Revealed by 2-DE Coupled to MALDI-TOF/TOF Proteomics Strategy. Int. J. Mol. Sci. 2020, 21, 4870. [Google Scholar] [CrossRef] [PubMed]
- Bradford, M.M. A Rapid and Sensitive Method for the Quantitation of Microgram Quantities of Protein Utilizing the Principle of Protein-Dye Binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Hewitt, E.J.; Dickes, G.J. Spectrophotometric Measurements on Ascorbic Acid and Their Use for the Estimation of Ascorbic Acid and Dehydroascorbic Acid in Plant Tissues. Biochem. J. 1961, 78, 384–391. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Griffith, O.W. Determination of Glutathione and Glutathione Disulfide Using Glutathione Reductase and 2-Vinylpyridine. Anal. Biochem. 1980, 106, 207–212. [Google Scholar] [CrossRef]
- Schafer, F.Q.; Buettner, G.R. Redox Environment of the Cell as Viewed through the Redox State of the Glutathione Disulfide/Glutathione Couple. Free Radic. Biol. Med. 2001, 30, 1191–1212. [Google Scholar] [CrossRef]
- Monéger, R.; Vermeersch, J.; Lechevallier, D.; Richard, D. Micro-analyse du NADP et du NAD réduits et oxydés dans les tissus foliaires et dans les plastes isolés de spirodèle et de blé. 1. Problèmes posés par le dosage séparé du extraits végétaux. Physiol. Vég. 1997, 15, 29–62. [Google Scholar]
- Lorenc-Plucińska, G.; Karolewski, P. Aluminium Effects on Pyridine Nucleotide Redox State in Roots of Scots Pine. Acta Soc. Bot. Pol. 1994, 63, 167–171. [Google Scholar] [CrossRef] [Green Version]
- Quebedeaux, B. Adenylate and Nicotinamide Nucleotides in Developing Soybean Seeds during Seed-Fill. Plant Physiol. 1981, 68, 23–27. [Google Scholar] [CrossRef] [PubMed]
- Hunt, L.; Holdsworth, M.J.; Gray, J.E. Nicotinamidase Activity Is Important for Germination. Plant J. Cell Mol. Biol. 2007, 51, 341–351. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2018. [Google Scholar]
- Wei, T.; Simko, V. R Package “Corrplot”: Visualization of a Correlation Matrix (Version 0.84). 2017. Available online: https://github.com/taiyun/corrplot (accessed on 21 June 2021).
- Xiao, W.; Wang, R.-S.; Handy, D.E.; Loscalzo, J. NAD(H) and NADP(H) Redox Couples and Cellular Energy Metabolism. Antioxid. Redox Signal. 2018, 28, 251–272. [Google Scholar] [CrossRef]
- Blacker, T.S.; Duchen, M.R. Investigating Mitochondrial Redox State Using NADH and NADPH Autofluorescence. Free Radic. Biol. Med. 2016, 100, 53–65. [Google Scholar] [CrossRef] [Green Version]
- Lunt, S.Y.; Vander Heiden, M.G. Aerobic Glycolysis: Meeting the Metabolic Requirements of Cell Proliferation. Annu. Rev. Cell Dev. Biol. 2011, 27, 441–464. [Google Scholar] [CrossRef] [Green Version]
- Chapman, A.G.; Fall, L.; Atkinson, D.E. Adenylate Energy Charge in Escherichia coli during Growth and Starvation. J. Bacteriol. 1971, 108, 1072–1086. [Google Scholar] [CrossRef] [Green Version]
- Gupta, M.; Cuypers, A.; Vangronsveld, J.; Clijsters, H. Copper Affects the Enzymes of the Ascorbate-Glutathione Cycle and Its Related Metabolites in the Roots of Phaseolus vulgaris. Physiol. Plant. Den. 1999, 106, 262–267. [Google Scholar] [CrossRef]
- Pukacka, S.; Ratajczak, E. Ascorbate and Glutathione Metabolism during Development and Desiccation of Beech (Fagus sylvatica L.) Seeds. Plant Growth Regul. 2010, 62, 77–83. [Google Scholar] [CrossRef]
- Jung, H.-I.; Lee, B.-R.; Chae, M.-J.; Lee, E.-J.; Lee, T.-G.; Jung, G.-B.; Kim, M.-S.; Lee, J. Ascorbate-Mediated Modulation of Cadmium Stress Responses: Reactive Oxygen Species and Redox Status in Brassica napus. Front. Plant Sci. 2020, 11, 586547. [Google Scholar] [CrossRef] [PubMed]
- Roach, T.; Beckett, R.P.; Minibayeva, F.V.; Colville, L.; Whitaker, C.; Chen, H.; Bailly, C.; Kranner, I. Extracellular Superoxide Production, Viability and Redox Poise in Response to Desiccation in Recalcitrant Castanea sativa Seeds. Plant Cell Environ. 2010, 33, 59–75. [Google Scholar] [CrossRef]
- Renard, J.; Niñoles, R.; Martínez-Almonacid, I.; Gayubas, B.; Mateos-Fernández, R.; Bissoli, G.; Bueso, E.; Serrano, R.; Gadea, J. Identification of Novel Seed Longevity Genes Related to Oxidative Stress and Seed Coat by Genome-Wide Association Studies and Reverse Genetics. Plant Cell Environ. 2020, 43, 2523–2539. [Google Scholar] [CrossRef]
- Reeg, S.; Grune, T. Protein Oxidation in Aging: Does It Play a Role in Aging Progression? Antioxid. Redox Signal. 2015, 23, 239–255. [Google Scholar] [CrossRef] [Green Version]
- Hashida, S.-N.; Kawai-Yamada, M.; Uchimiya, H. NAD+ Accumulation as a Metabolic off Switch for Orthodox Pollen. Plant Signal. Behav. 2013, 8, e23937. [Google Scholar] [CrossRef] [Green Version]
- Ying, W. NAD+/NADH and NADP+/NADPH in Cellular Functions and Cell Death: Regulation and Biological Consequences. Antioxid. Redox Signal. 2008, 10, 179–206. [Google Scholar] [CrossRef] [Green Version]
- Tommasi, F.; Paciolla, C.; Arrigoni, O. The Ascorbate System in Recalcitrant and Orthodox Seeds. Physiol. Plant. 1999, 105, 193–198. [Google Scholar] [CrossRef]
- Ding, H.; Wang, B.; Han, Y.; Li, S. The Pivotal Function of Dehydroascorbate Reductase in Glutathione Homeostasis in Plants. J. Exp. Bot. 2020, 71, 3405–3416. [Google Scholar] [CrossRef] [PubMed]
- Turner, W.L.; Waller, J.C.; Snedden, W.A. Identification, Molecular Cloning and Functional Characterization of a Novel NADH Kinase from Arabidopsis thaliana (Thale Cress). Biochem. J. 2005, 385, 217–223. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Corpas, F.J.; Barroso, J.B. Peroxisomal Plant Metabolism—An Update on Nitric Oxide, Ca2+ and the NADPH Recycling Network. J. Cell Sci. 2018, 131, jcs202978. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pan, R.; Liu, J.; Hu, J. Peroxisomes in Plant Reproduction and Seed-Related Development. J. Integr. Plant Biol. 2019, 61, 784–802. [Google Scholar] [CrossRef] [Green Version]
- Palsson, B.Ø. Systems Biology: Simulation of Dynamic Network States; Cambridge University Press: Cambridge, UK, 2011; ISBN 978-1-107-00159-6. [Google Scholar]
- Karasov, T.L.; Chae, E.; Herman, J.J.; Bergelson, J. Mechanisms to Mitigate the Trade-Off between Growth and Defense. Plant Cell 2017, 29, 666–680. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Leprince, O.; Buitink, J.; Hoekstra, F.A. Axes and Cotyledons of Recalcitrant Seeds of Castanea sativa Mill. Exhibit Contrasting Responses of Respiration to Drying in Relation to Desiccation Sensitivity. J. Exp. Bot. 1999, 50, 1515–1524. [Google Scholar] [CrossRef]
- Anjum, N.A.; Sharma, P.; Gill, S.S.; Hasanuzzaman, M.; Khan, E.A.; Kachhap, K.; Mohamed, A.A.; Thangavel, P.; Devi, G.D.; Vasudhevan, P.; et al. Catalase and Ascorbate Peroxidase-Representative H2O2-Detoxifying Heme Enzymes in Plants. Environ. Sci. Pollut. Res. Int. 2016, 23, 19002–19029. [Google Scholar] [CrossRef] [PubMed]
- Palma, J.M.; Mateos, R.M.; López-Jaramillo, J.; Rodríguez-Ruiz, M.; González-Gordo, S.; Lechuga-Sancho, A.M.; Corpas, F.J. Plant Catalases as NO and H2S Targets. Redox Biol. 2020, 34, 101525. [Google Scholar] [CrossRef]
- The UniProt Consortium. UniProt: The Universal Protein Knowledgebase in 2021. Nucleic Acids Res. 2021, 49, D480–D489. [Google Scholar] [CrossRef] [PubMed]
- Mhamdi, A.; Noctor, G.; Baker, A. Plant Catalases: Peroxisomal Redox Guardians. Arch. Biochem. Biophys. 2012, 525, 181–194. [Google Scholar] [CrossRef]
- Huan, C.; Jiang, L.; An, X.; Yu, M.; Xu, Y.; Ma, R.; Yu, Z. Potential Role of Reactive Oxygen Species and Antioxidant Genes in the Regulation of Peach Fruit Development and Ripening. Plant Physiol. Biochem. PPB 2016, 104, 294–303. [Google Scholar] [CrossRef]
- López, A.P.; Gochicoa, M.T.N.; Franco, A.R. Activities of Antioxidant Enzymes during Strawberry Fruit Development and Ripening. Biol. Plant. 2010, 54, 349–352. [Google Scholar] [CrossRef]
- Bailly, C.; Leymarie, J.; Lehner, A.; Rousseau, S.; Côme, D.; Corbineau, F. Catalase Activity and Expression in Developing Sunflower Seeds as Related to Drying. J. Exp. Bot. 2004, 55, 475–483. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Eising, R.; Trelease, R.N.; Ni, W.T. Biogenesis of Catalase in Glyoxysomes and Leaf-Type Peroxisomes of Sunflower Cotyledons. Arch. Biochem. Biophys. 1990, 278, 258–264. [Google Scholar] [CrossRef]
- Kalemba, E.M.; Bagniewska-Zadworna, A.; Suszka, J.; Pukacka, S. Dehydration Sensitivity at the Early Seedling Establishment Stages of the European Beech (Fagus sylvatica L.). Forests 2019, 10, 900. [Google Scholar] [CrossRef] [Green Version]








Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kalemba, E.M.; Alipour, S.; Wojciechowska, N. NAD(P)H Drives the Ascorbate–Glutathione Cycle and Abundance of Catalase in Developing Beech Seeds Differently in Embryonic Axes and Cotyledons. Antioxidants 2021, 10, 2021. https://doi.org/10.3390/antiox10122021
Kalemba EM, Alipour S, Wojciechowska N. NAD(P)H Drives the Ascorbate–Glutathione Cycle and Abundance of Catalase in Developing Beech Seeds Differently in Embryonic Axes and Cotyledons. Antioxidants. 2021; 10(12):2021. https://doi.org/10.3390/antiox10122021
Chicago/Turabian StyleKalemba, Ewa Marzena, Shirin Alipour, and Natalia Wojciechowska. 2021. "NAD(P)H Drives the Ascorbate–Glutathione Cycle and Abundance of Catalase in Developing Beech Seeds Differently in Embryonic Axes and Cotyledons" Antioxidants 10, no. 12: 2021. https://doi.org/10.3390/antiox10122021
APA StyleKalemba, E. M., Alipour, S., & Wojciechowska, N. (2021). NAD(P)H Drives the Ascorbate–Glutathione Cycle and Abundance of Catalase in Developing Beech Seeds Differently in Embryonic Axes and Cotyledons. Antioxidants, 10(12), 2021. https://doi.org/10.3390/antiox10122021