
Molybdenum-Induced Regulation of Antioxidant Defense-Mitigated Cadmium Stress in Aromatic Rice and Improved Crop Growth, Yield, and Quality Traits

 ,
,  ,
,  ,
,
Abstract
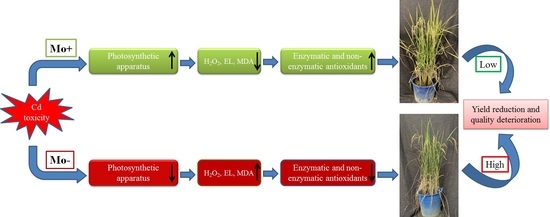
:
1. Introduction
2. Materials and Methods
2.1. Experimental Location, Treatments, and Crop Husbandry
2.2. Sampling
2.3. Estimation of Photosynthetic Efficiency, Pigment Contents, and Transmission Electron Microscopy
2.4. Measurement of Soluble Protein and Proline Contents
2.5. Measurement of Reactive Oxygen Species and Lipid Peroxidation
2.6. Measurement of Enzymatic and Non-Enzymatic Antioxidant
2.7. Total RNA Extraction and qRT-PCR Analysis
2.8. Estimation of Molybdenum and Cadmium Concentration in Different Plant Parts
2.9. Estimation of 2-acetyl-1-pyrroline (2AP) Content
2.10. Measurement of Grain Yield, Yield Attributes, and Grain Quality Characters of Aromatic Rice
2.11. Statistical Analysis
3. Results
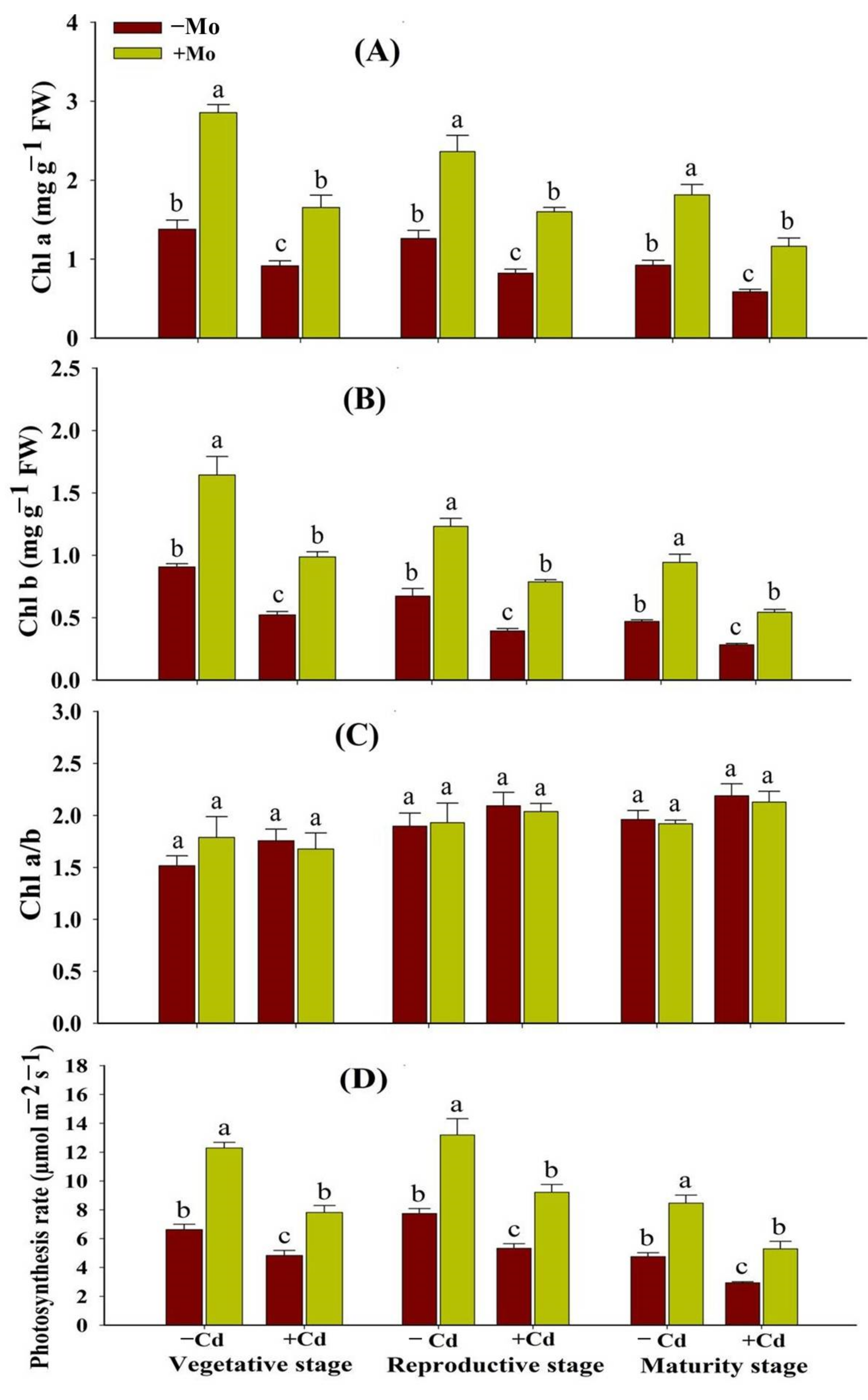
3.1. Effect of Mo Supply on Photosynthetic Pigments and Photosynthetic Efficiency and Chloroplast Configuration of Aromatic Rice Plants under Cd Toxicity
3.2. Influence of Mo Supply on Osmo-Regulation under Cd Toxicity
3.3. Effect of Mo Supply on Membrane Integrity in Aromatic Rice under Cd Stress
3.4. Effect of Mo Supply on the Activities and Transcript Abundance of Enzymatic Antioxidants under Cd Toxicity in Aromatic Rice
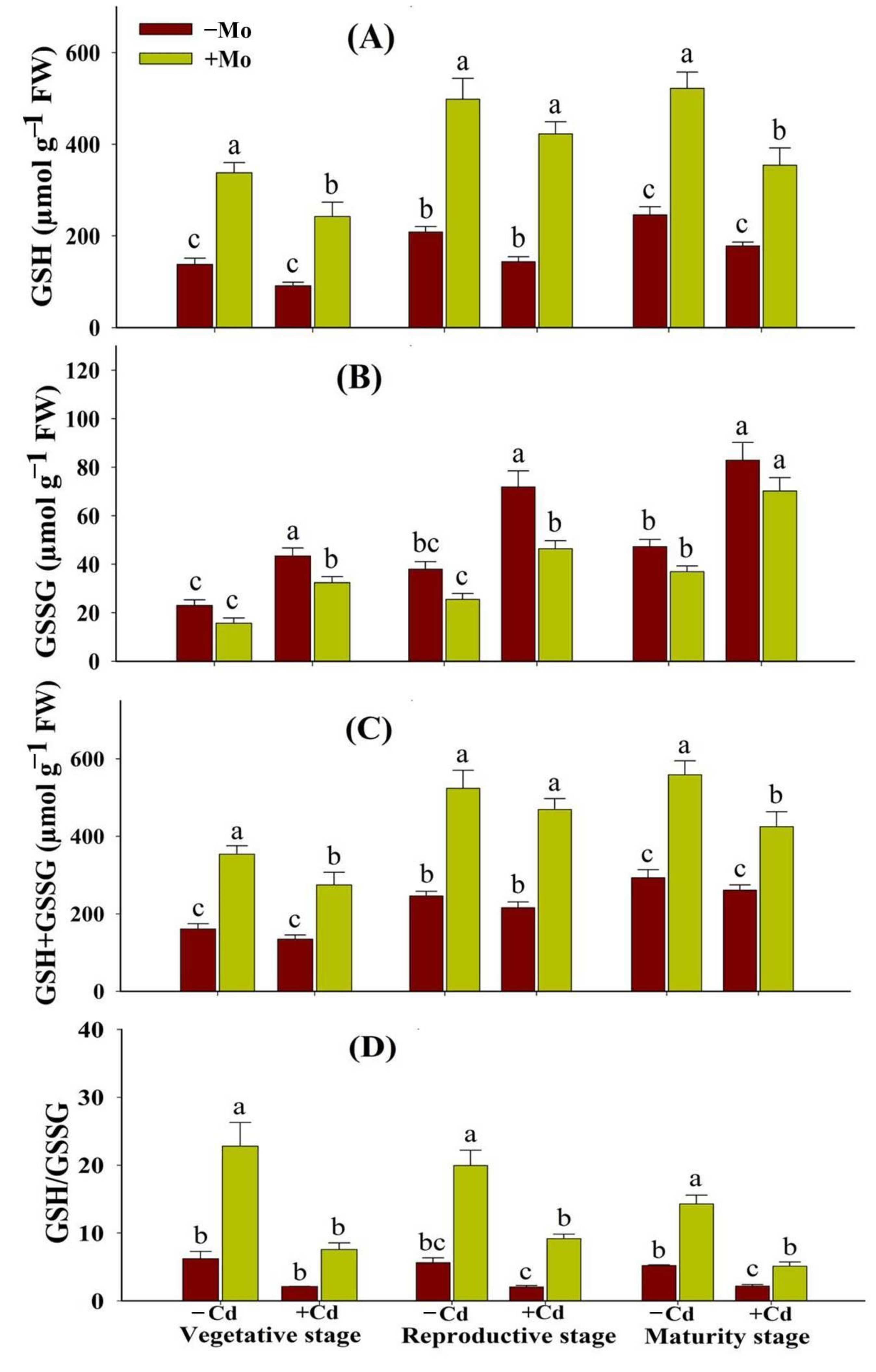
3.5. Influence of Mo Supply on Non-Enzymatic Antioxidants in Aromatic Rice under Cd-Stress
3.6. Mo and Cd Concentrations in Different Plant Parts of Aromatic Rice Plants
3.7. Influence of Mo Supply on Yield Traits and Grain Quality Characteristics of Aromatic Rice under Cd Stress
3.8. Relationships
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Bryant, R.; McClung, A. Volatile profiles of aromatic and non-aromatic rice cultivars using SPME/GC–MS. Food Chem. 2011, 124, 501–513. [Google Scholar] [CrossRef]
- Ashraf, U.; Kanu, A.S.; Deng, Q.; Mo, Z.; Pan, S.; Tian, H.; Tang, X. Lead (Pb) toxicity; physio-biochemical mechanisms, grain yield, quality, and Pb distribution proportions in scented rice. Front. Plant Sci. 2017, 8, 259. [Google Scholar] [CrossRef] [Green Version]
- Grant, C.; Clarke, J.; Duguid, S.; Chaney, R. Selection and breeding of plant cultivars to minimize cadmium accumulation. Sci. Total Environ. 2008, 390, 301–310. [Google Scholar] [CrossRef]
- Ismael, M.A.; Elyamine, A.M.; Moussa, M.G.; Cai, M.; Zhao, X.; Hu, C. Cadmium in plants: Uptake, toxicity, and its interactions with selenium fertilizers. Metallomics 2019, 11, 255–277. [Google Scholar] [CrossRef]
- Uraguchi, S.; Mori, S.; Kuramata, M.; Kawasaki, A.; Arao, T.; Ishikawa, S. Root-to-shoot Cd translocation via the xylem is the major process determining shoot and grain cadmium accumulation in rice. J. Exp. Bot. 2009, 60, 2677–2688. [Google Scholar] [CrossRef] [Green Version]
- Hussain, S.; Khaliq, A.; Noor, M.A.; Tanveer, M.; Hussain, H.A.; Hussain, S.; Shah, T.; Mehmood, T. Metal Toxicity and Nitrogen Metabolism in Plants: An Overview. In Carbon and Nitrogen Cycling in Soil; Springer: Berlin/Heidelberg, Germany, 2020; pp. 221–248. [Google Scholar]
- Cao, F.; Wang, R.; Cheng, W.; Zeng, F.; Ahmed, I.M.; Hu, X.; Zhang, G.; Wu, F. Genotypic and environmental variation in cadmium, chromium, lead and copper in rice and approaches for reducing the accumulation. Sci. Total Environ. 2014, 496, 275–281. [Google Scholar] [CrossRef]
- Imran, M.; Hussain, S.; Rana, M.S.; Saleem, M.H.; Rasul, F.; Ali, K.H.; Potcho, M.P.; Pan, S.; Duan, M.; Tang, X. Molybdenum improves 2-acetyl-1-pyrroline, grain quality traits and yield attributes in fragrant rice through efficient nitrogen assimilation under cadmium toxicity. Ecotoxicol. Environ. Saf. 2021, 211, 111911. [Google Scholar] [CrossRef] [PubMed]
- Rana, M.S.; Hu, C.X.; Shaaban, M.; Imran, M.; Afzal, J.; Moussa, M.G.; Elyamine, A.M.; Bhantana, P.; Saleem, M.H.; Syaifudin, M. Soil phosphorus transformation characteristics in response to molybdenum supply in leguminous crops. J. Environ. Manag. 2020, 268, 110610. [Google Scholar] [CrossRef] [PubMed]
- Rana, M.S.; Sun, X.; Imran, M.; Ali, S.; Shaaban, M.; Moussa, M.G.; Khan, Z.; Afzal, J.; Binyamin, R.; Bhantana, P. Molybdenum-induced effects on leaf ultra-structure and rhizosphere phosphorus transformation in Triticum aestivum L. Plant Physiol. Biochem. 2020, 153, 20–29. [Google Scholar] [CrossRef] [PubMed]
- Zhang, M.; Hu, C.; Zhao, X.; Tan, Q.; Sun, X.; Cao, A.; Cui, M.; Zhang, Y. Molybdenum improves antioxidant and osmotic-adjustment ability against salt stress in Chinese cabbage (Brassica campestris L. ssp. Pekinensis). Plant Soil 2012, 355, 375–383. [Google Scholar] [CrossRef]
- Wu, S.; Hu, C.; Tan, Q.; Xu, S.; Sun, X. Nitric oxide mediates molybdenum-induced antioxidant defense in wheat under drought stress. Front. Plant Sci. 2017, 8, 1085. [Google Scholar] [CrossRef] [PubMed]
- Sun, X.; Hu, C.; Tan, Q.; Liu, J.; Liu, H. Effects of molybdenum on expression of cold-responsive genes in abscisic acid (ABA)-dependent and ABA-independent pathways in winter wheat under low-temperature stress. Ann. Bot. 2009, 104, 345–356. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Imran, M.; Sun, X.; Hussain, S.; Ali, U.; Rana, M.S.; Rasul, F.; Saleem, M.H.; Moussa, M.G.; Bhantana, P.; Afzal, J. Molybdenum-Induced Effects on Nitrogen Metabolism Enzymes and Elemental Profile of Winter Wheat (Triticum aestivum L.) Under Different Nitrogen Sources. Int. J. Mol. Sci. 2019, 20, 3009. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ismael, M.A.; Elyamine, A.M.; Zhao, Y.Y.; Moussa, M.G.; Rana, M.S.; Afzal, J.; Imran, M.; Zhao, X.H.; Hu, C.X. Can selenium and molybdenum restrain cadmium toxicity to pollen grains in Brassica napus? Int. J. Mol. Sci. 2018, 19, 2163. [Google Scholar] [CrossRef] [Green Version]
- Ali, N.; Hadi, F.; Ali, M. Growth stage and molybdenum treatment affect cadmium accumulation, antioxidant defence and chlorophyll contents in Cannabis sativa plant. Chemosphere 2019, 236, 124360. [Google Scholar] [CrossRef]
- Kanu, A.S.; Ashraf, U.; Mo, Z.; Fuseini, I.; Mansaray, L.R.; Duan, M.; Pan, S.; Tang, X. Cadmium uptake and distribution in fragrant rice genotypes and related consequences on yield and grain quality traits. J. Chem. 2017, 2017, 1405878. [Google Scholar] [CrossRef]
- Ihtisham, M.; Liu, S.; Shahid, M.O.; Khan, N.; Lv, B.; Sarraf, M.; Ali, S.; Chen, L.; Liu, Y.; Chen, Q. The Optimized N, P, and K Fertilization for Bermudagrass Integrated Turf Performance during the Establishment and Its Importance for the Sustainable Management of Urban Green Spaces. Sustainability 2020, 12, 294. [Google Scholar] [CrossRef]
- Ihtisham, M.; Fahad, S.; Luo, T.; Larkin, R.M.; Yin, S.; Chen, L. Optimization of nitrogen, phosphorus, and potassium fertilization rates for overseeded perennial ryegrass turf on dormant bermudagrass in a transitional climate. Front. Plant Sci. 2018, 9, 487. [Google Scholar] [CrossRef] [Green Version]
- Saleem, M.H.; Ali, S.; Rehman, M.; Rana, M.S.; Rizwan, M.; Kamran, M.; Imran, M.; Riaz, M.; Soliman, M.H.; Elkelish, A. Influence of phosphorus on copper phytoextraction via modulating cellular organelles in two jute (Corchorus capsularis L.) varieties grown in a copper mining soil of Hubei Province, China. Chemosphere 2020, 248, 126032. [Google Scholar] [CrossRef]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Bates, L.S.; Waldren, R.P.; Teare, I. Rapid determination of free proline for water-stress studies. Plant Soil 1973, 39, 205–207. [Google Scholar] [CrossRef]
- Valentovic, P.; Luxova, M.; Kolarovic, L.; Gasparikova, O. Effect of osmotic stress on compatible solutes content, membrane stability and water relations in two maize cultivars. Plant Soil Environ. 2006, 52, 184. [Google Scholar]
- Imran, M.; Sun, X.; Hussain, S.; Ali, U.; Rana, M.S.; Rasul, F.; Shaukat, S.; Hu, C. Molybdenum application regulates oxidative stress tolerance in winter wheat under different nitrogen sources. J. Soil Sci. Plant Nutr. 2020, 20, 1827–1837. [Google Scholar] [CrossRef]
- Imran, M.; Sun, X.; Hussain, S.; Rana, M.S.; Saleem, M.H.; Riaz, M.; Tang, X.; Khan, I.; Hu, C. Molybdenum supply increases root system growth of winter wheat by enhancing nitric oxide accumulation and expression of NRT genes. Plant Soil 2021, 459, 235–248. [Google Scholar] [CrossRef]
- Pfaffl, M.W. A new mathematical model for relative quantification in real-time RT–PCR. Nucleic Acids Res. 2001, 29, e45. [Google Scholar] [CrossRef] [PubMed]
- Filipiak-Szok, A.; Kurzawa, M.; Szłyk, E. Determination of toxic metals by ICP-MS in Asiatic and European medicinal plants and dietary supplements. J. Trace Elem. Med. Biol. 2015, 30, 54–58. [Google Scholar] [CrossRef]
- Mo, Z.; Li, Y.; Nie, J.; He, L.; Pan, S.; Duan, M.; Tian, H.; Xiao, L.; Zhong, K.; Tang, X. Nitrogen application and different water regimes at booting stage improved yield and 2-acetyl-1-pyrroline (2AP) formation in fragrant rice. Rice 2019, 12, 74. [Google Scholar] [CrossRef] [Green Version]
- Mo, Z.; Ashraf, U.; Tang, Y.; Li, W.; Pan, S.; Duan, M.; Tian, H.; Tang, X. Nitrogen application at the booting stage affects 2-acetyl-1-pyrroline, proline, and total nitrogen contents in aromatic rice. Chil. J. Agric. Res. 2018, 78, 165–172. [Google Scholar] [CrossRef] [Green Version]
- Cao, F.; Cai, Y.; Liu, L.; Zhang, M.; He, X.; Zhang, G.; Wu, F. Differences in photosynthesis, yield and grain cadmium accumulation as affected by exogenous cadmium and glutathione in the two rice genotypes. Plant Growth Regul. 2015, 75, 715–723. [Google Scholar] [CrossRef]
- Fahad, S.; Rehman, A.; Shahzad, B.; Tanveer, M.; Saud, S.; Kamran, M.; Ihtisham, M.; Khan, S.U.; Turan, V.; ur Rahman, M.H. Rice responses and tolerance to metal/metalloid toxicity. In Advances in Rice Research for Abiotic Stress Tolerance; Elsevier: Amsterdam, The Netherlands, 2019; pp. 299–312. [Google Scholar]
- Saleem, M.H.; Rehman, M.; Zahid, M.; Imran, M.; Xiang, W.; Liu, L. Morphological changes and antioxidative capacity of jute (Corchorus capsularis, Malvaceae) under different color light-emitting diodes. Braz. J. Bot. 2019, 42, 581–590. [Google Scholar] [CrossRef]
- Najeeb, U.; Jilani, G.; Ali, S.; Sarwar, M.; Xu, L.; Zhou, W. Insights into cadmium induced physiological and ultra-structural disorders in Juncus effusus L. and its remediation through exogenous citric acid. J. Hazard. Mater. 2011, 186, 565–574. [Google Scholar] [CrossRef] [PubMed]
- Padmaja, K.; Prasad, D.; Prasad, A. Inhibition of chlorophyll synthesis in Phaseolus vulgaris L. seedlings by cadmium acetate. Photosynthetica 1990, 24, 399–405. [Google Scholar]
- Liu, Z.; Gao, J.; Gao, F.; Liu, P.; Zhao, B.; Zhang, J. Photosynthetic characteristics and chloroplast ultrastructure of summer maize response to different nitrogen supplies. Front. Plant Sci. 2018, 9, 576. [Google Scholar] [CrossRef] [PubMed]
- Imran, M.; Hu, C.; Hussain, S.; Rana, M.S.; Riaz, M.; Afzal, J.; Aziz, O.; Elyamine, A.M.; Ismael, M.A.F.; Sun, X. Molybdenum-induced effects on photosynthetic efficacy of winter wheat (Triticum aestivum L.) under different nitrogen sources are associated with nitrogen assimilation. Plant Physiol. Biochem. 2019, 141, 154–163. [Google Scholar] [CrossRef] [PubMed]
- Liu, L.; Xiao, W.; Li, L.; Li, D.-M.; Gao, D.-S.; Zhu, C.-y.; Fu, X.-L. Effect of exogenously applied molybdenum on its absorption and nitrate metabolism in strawberry seedlings. Plant Physiol. Biochem. 2017, 115, 200–211. [Google Scholar] [CrossRef]
- Hasanuzzaman, M.; Inafuku, M.; Nahar, K.; Fujita, M.; Oku, H. Nitric Oxide Regulates Plant Growth, Physiology, Antioxidant Defense, and Ion Homeostasis to Confer Salt Tolerance in the Mangrove Species, Kandelia obovata. Antioxidants 2021, 10, 611. [Google Scholar] [CrossRef]
- Palma, J.M.; Sandalio, L.M.; Corpas, F.J.; Romero-Puertas, M.C.; McCarthy, I.; Luis, A. Plant proteases, protein degradation, and oxidative stress: Role of peroxisomes. Plant Physiol. Biochem. 2002, 40, 521–530. [Google Scholar] [CrossRef]
- Romero-Puertas, M.; Palma, J.; Gómez, M.; Del Rio, L.; Sandalio, L. Cadmium causes the oxidative modification of proteins in pea plants. Plant Cell Environ. 2002, 25, 677–686. [Google Scholar] [CrossRef]
- Li, L.; Wei, X.; Ji, M.-L.; Chao, Y.; Ling, L.; Gao, D.-S.; Fu, X.-L. Effects of molybdenum on nutrition, quality, and flavour compounds of strawberry (Fragaria× ananassa Duch. cv. Akihime) fruit. J. Integr. Agric. 2017, 16, 1502–1512. [Google Scholar] [CrossRef]
- Wu, Z.; Zhao, X.; Sun, X.; Tan, Q.; Tang, Y.; Nie, Z.; Qu, C.; Chen, Z.; Hu, C. Antioxidant enzyme systems and the ascorbate–glutathione cycle as contributing factors to cadmium accumulation and tolerance in two oilseed rape cultivars (Brassica napus L.) under moderate cadmium stress. Chemosphere 2015, 138, 526–536. [Google Scholar] [CrossRef]
- Moustafa-Farag, M.; Mahmoud, A.; Arnao, M.B.; Sheteiwy, M.S.; Dafea, M.; Soltan, M.; Elkelish, A.; Hasanuzzaman, M.; Ai, S. Melatonin-induced water stress tolerance in plants: Recent advances. Antioxidants 2020, 9, 809. [Google Scholar] [CrossRef] [PubMed]
- Hasanuzzaman, M.; Bhuyan, M.; Zulfiqar, F.; Raza, A.; Mohsin, S.M.; Mahmud, J.A.; Fujita, M.; Fotopoulos, V. Reactive oxygen species and antioxidant defense in plants under abiotic stress: Revisiting the crucial role of a universal defense regulator. Antioxidants 2020, 9, 681. [Google Scholar] [CrossRef] [PubMed]
- Sachdev, S.; Ansari, S.A.; Ansari, M.I.; Fujita, M.; Hasanuzzaman, M. Abiotic stress and reactive oxygen species: Generation, signaling, and defense mechanisms. Antioxidants 2021, 10, 277. [Google Scholar] [CrossRef] [PubMed]
- Hussain, S.; Khan, F.; Cao, W.; Wu, L.; Geng, M. Seed priming alters the production and detoxification of reactive oxygen intermediates in rice seedlings grown under sub-optimal temperature and nutrient supply. Front. Plant Sci. 2016, 7, 439. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Imran, M.; Hussain, S.; El-Esawi, M.A.; Rana, M.S.; Saleem, M.H.; Riaz, M.; Ashraf, U.; Potcho, M.P.; Duan, M.; Rajput, I.A. Molybdenum Supply Alleviates the Cadmium Toxicity in Fragrant Rice by Modulating Oxidative Stress and Antioxidant Gene Expression. Biomolecules 2020, 10, 1582. [Google Scholar] [CrossRef]
- Sharma, S.S.; Dietz, K.-J. The relationship between metal toxicity and cellular redox imbalance. Trends Plant Sci. 2009, 14, 43–50. [Google Scholar] [CrossRef]
- Monostori, P.; Wittmann, G.; Karg, E.; Túri, S. Determination of glutathione and glutathione disulfide in biological samples: An in-depth review. J. Chromatogr. B 2009, 877, 3331–3346. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Cd Toxicity | Mo Application | Tillers Pot−1 | Productive Tillers Pot−1 | Grains Panicle−1 | Filled Grain Percentage | 1000-Grain Weight (g) | Grain Yield Pot−1 (g) |
|---|---|---|---|---|---|---|---|
| −Cd | −Mo | 26.97 ± 1.36 b | 21.31 ± 1.48 c | 128.91 ± 4.59 b | 68.24 ± 2.23 b | 18.11 ± 0.42 b | 46.11 ± 2.47 c |
| +Mo | 40.08 ± 2.70 a | 32.46 ± 1.47 a | 165.21 ± 8.61 a | 83.35 ± 5.15 a | 20.30 ± 0.85 a | 73.36 ± 4.32 a | |
| +Cd | −Mo | 19.89 ± 0.61 c | 16.02 ± 1.40 d | 103.72 ± 3.48 c | 57.76 ± 2.81 c | 16.97 ± 0.67 b | 35.26 ± 1.43 d |
| +Mo | 32.05 ± 1.59 b | 27.20 ± 1.18 b | 145.28 ± 5.67 b | 72.25 ± 1.88 b | 18.70 ± 0.64 ab | 58.08 ± 4.19 b | |
| LSD (p < 0.05) | 5.35 | 4.27 | 18.20 | 10.09 | 2.05 | 10.26 |
| Cd Toxicity | Mo Application | Brown Rice Rate (%) | Milled Rice Rate (%) | Milling Degree (%) | Head Rice Rate (%) | Chalkiness Rate (%) | Chalkiness Degree (%) | Moisture Content (%) | Protein Content (%) | 2AP Content (ng g−1 FW) |
|---|---|---|---|---|---|---|---|---|---|---|
| −Cd | −Mo | 73.89 ± 3.78 a | 60.14 ± 1.52 bc | 77.91 ± 1.59 b | 56.89 ± 3.82 bc | 33.66 ± 2.44 b | 16.94 ± 0.53 b | 12.27 ± 0.08 b | 6.47 ± 0.14 c | 144.35 ± 7.40 b |
| +Mo | 77.81 ± 1.87 a | 66.42 ± 1.70 a | 85.83 ± 1.46 a | 68.53 ± 5.26 a | 24.06 ± 1.28 c | 11.75 ± 0.58 c | 12.68 ± 0.18 a | 8.49 ± 0.22 a | 297.08 ± 23.91 a | |
| +Cd | −Mo | 71.24 ± 1.78 a | 55.20 ± 1.71 c | 71.55 ± 1.82 c | 49.57 ± 2.04 c | 39.78 ± 1.70 a | 20.62 ± 0.94 a | 12.12 ± 0.13 b | 5.50 ± 0.09 d | 97.01 ± 6.39 c |
| +Mo | 75.22 ± 2.31 a | 62.02 ± 2.35 ab | 81.74 ± 2.05 ab | 62.34 ± 2.94 ab | 30.74 ± 1.71 b | 15.25 ± 0.56 b | 12.45 ± 0.07 ab | 7.12 ± 0.12 b | 171.79 ± 14.58 b | |
| LSD (p < 0.05) | 7.89 | 5.69 | 5.38 | 11.42 | 5.50 | 2.07 | 0.38 | 0.47 | 45.69 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Imran, M.; Hussain, S.; He, L.; Ashraf, M.F.; Ihtisham, M.; Warraich, E.A.; Tang, X. Molybdenum-Induced Regulation of Antioxidant Defense-Mitigated Cadmium Stress in Aromatic Rice and Improved Crop Growth, Yield, and Quality Traits. Antioxidants 2021, 10, 838. https://doi.org/10.3390/antiox10060838
Imran M, Hussain S, He L, Ashraf MF, Ihtisham M, Warraich EA, Tang X. Molybdenum-Induced Regulation of Antioxidant Defense-Mitigated Cadmium Stress in Aromatic Rice and Improved Crop Growth, Yield, and Quality Traits. Antioxidants. 2021; 10(6):838. https://doi.org/10.3390/antiox10060838
Chicago/Turabian StyleImran, Muhammad, Saddam Hussain, Longxin He, Muhammad Furqan Ashraf, Muhammad Ihtisham, Ejaz Ahmad Warraich, and Xiangru Tang. 2021. "Molybdenum-Induced Regulation of Antioxidant Defense-Mitigated Cadmium Stress in Aromatic Rice and Improved Crop Growth, Yield, and Quality Traits" Antioxidants 10, no. 6: 838. https://doi.org/10.3390/antiox10060838
APA StyleImran, M., Hussain, S., He, L., Ashraf, M. F., Ihtisham, M., Warraich, E. A., & Tang, X. (2021). Molybdenum-Induced Regulation of Antioxidant Defense-Mitigated Cadmium Stress in Aromatic Rice and Improved Crop Growth, Yield, and Quality Traits. Antioxidants, 10(6), 838. https://doi.org/10.3390/antiox10060838