
Investigating the Drought and Salinity Effect on the Redox Components of Sulla coronaria (L.) Medik

 ,
,  , and
, and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Material and Stress Application
2.2. Biometric Analysis
2.3. Water Content
2.4. Levels of Photosynthetic Pigments
2.5. Photosynthetic Activity
2.6. Trypan Blue Staining
2.7. Proline Concentration
2.8. Extraction and Quantification of ABA and Ascorbate by High-Performance Liquid Chromatography with Diode-Array Detection (HPLC-DAD)
2.9. Reactive Oxygen Species Detection
2.10. Lipid Peroxidation
2.11. Total Phenolic Content
2.12. Glutathione Quantitation
2.13. Antioxidant Enzyme Extraction and Activity Assays
2.14. Quantitative Real-Time PCR (qRT-PCR)
2.15. Statistical Analysis
3. Results
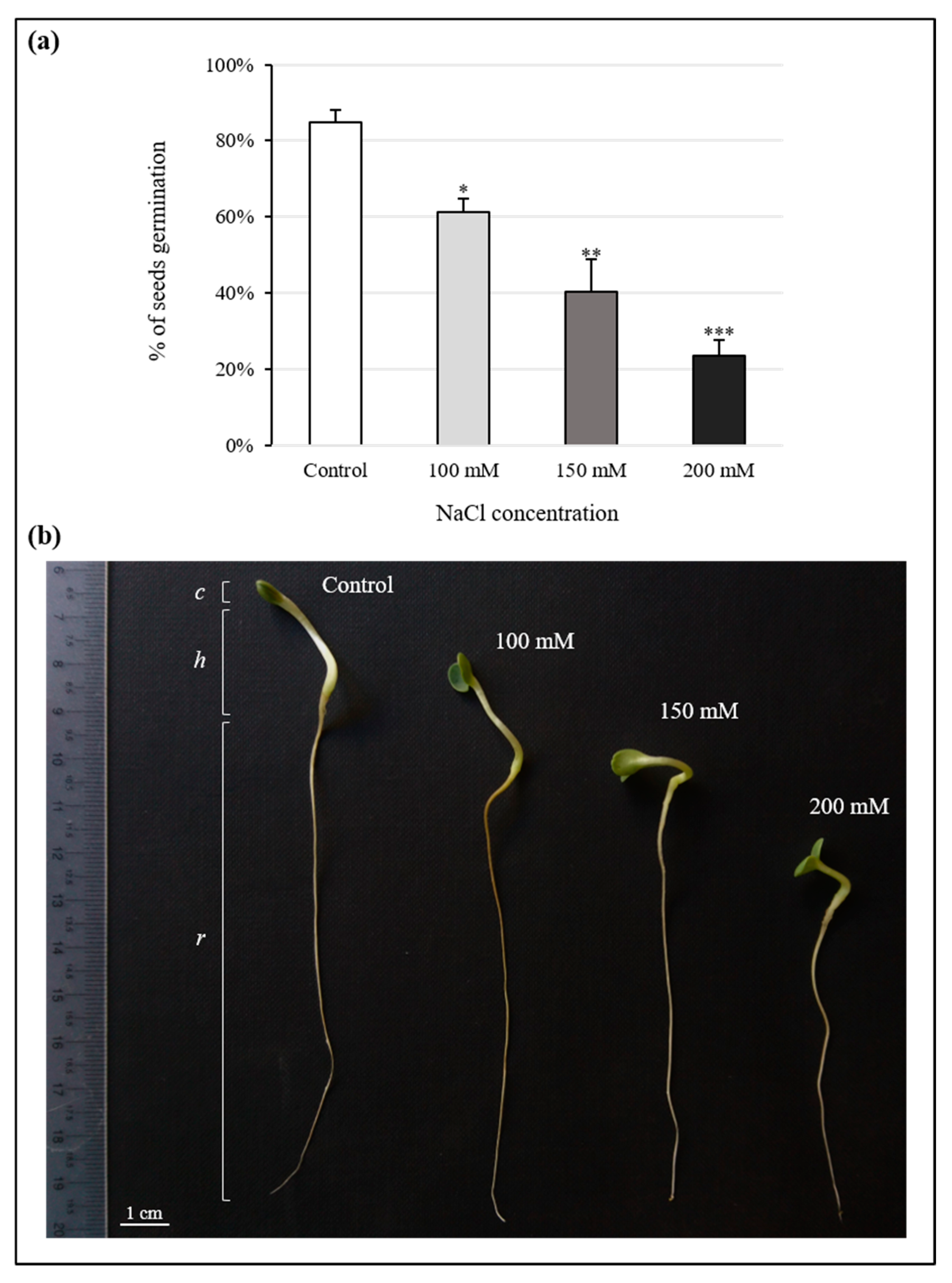
3.1. Effects of NaCl on Seed Germination
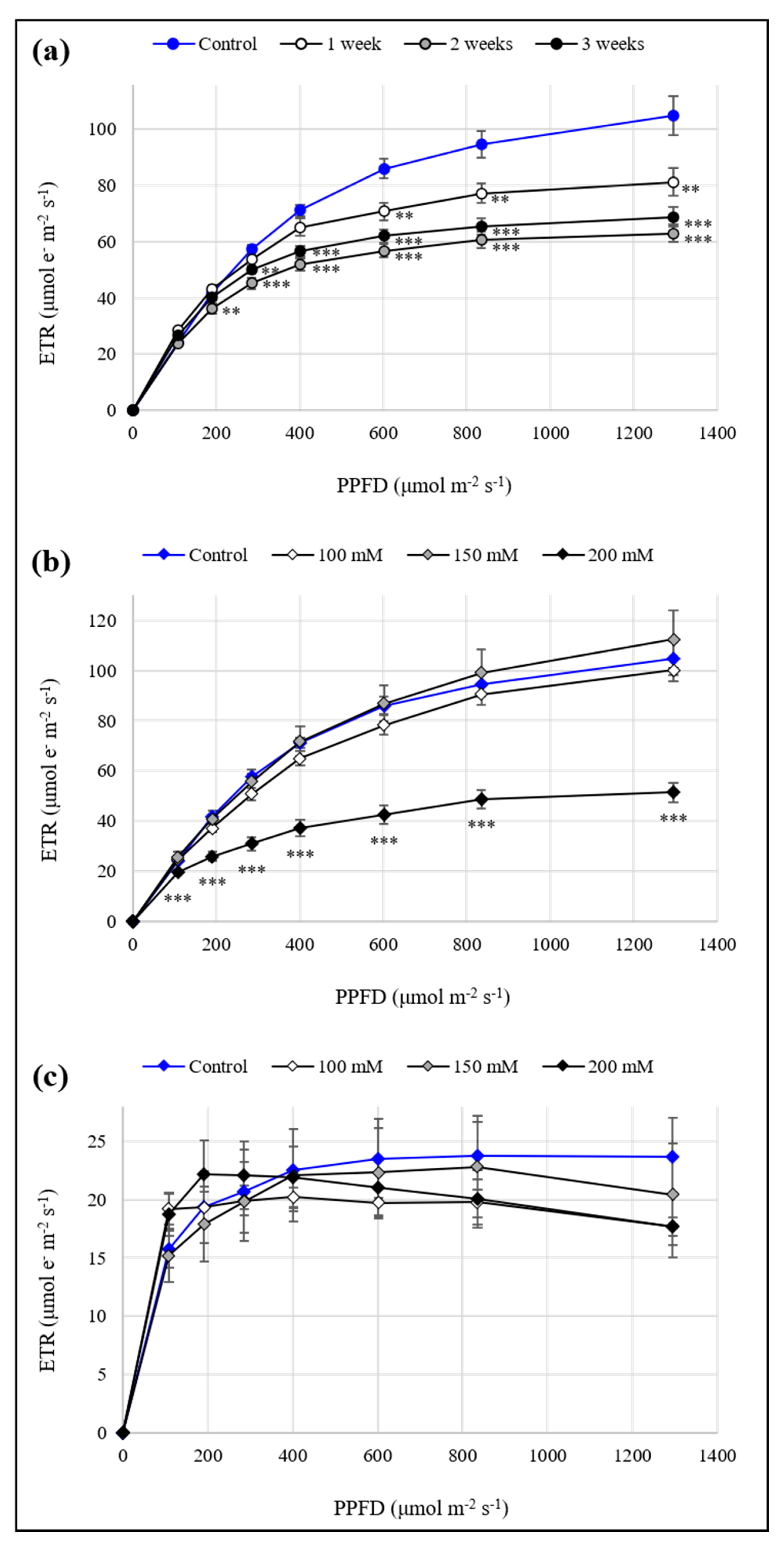
3.2. Evaluation of Plant Physiological State after Exposure to Abiotic Stressors
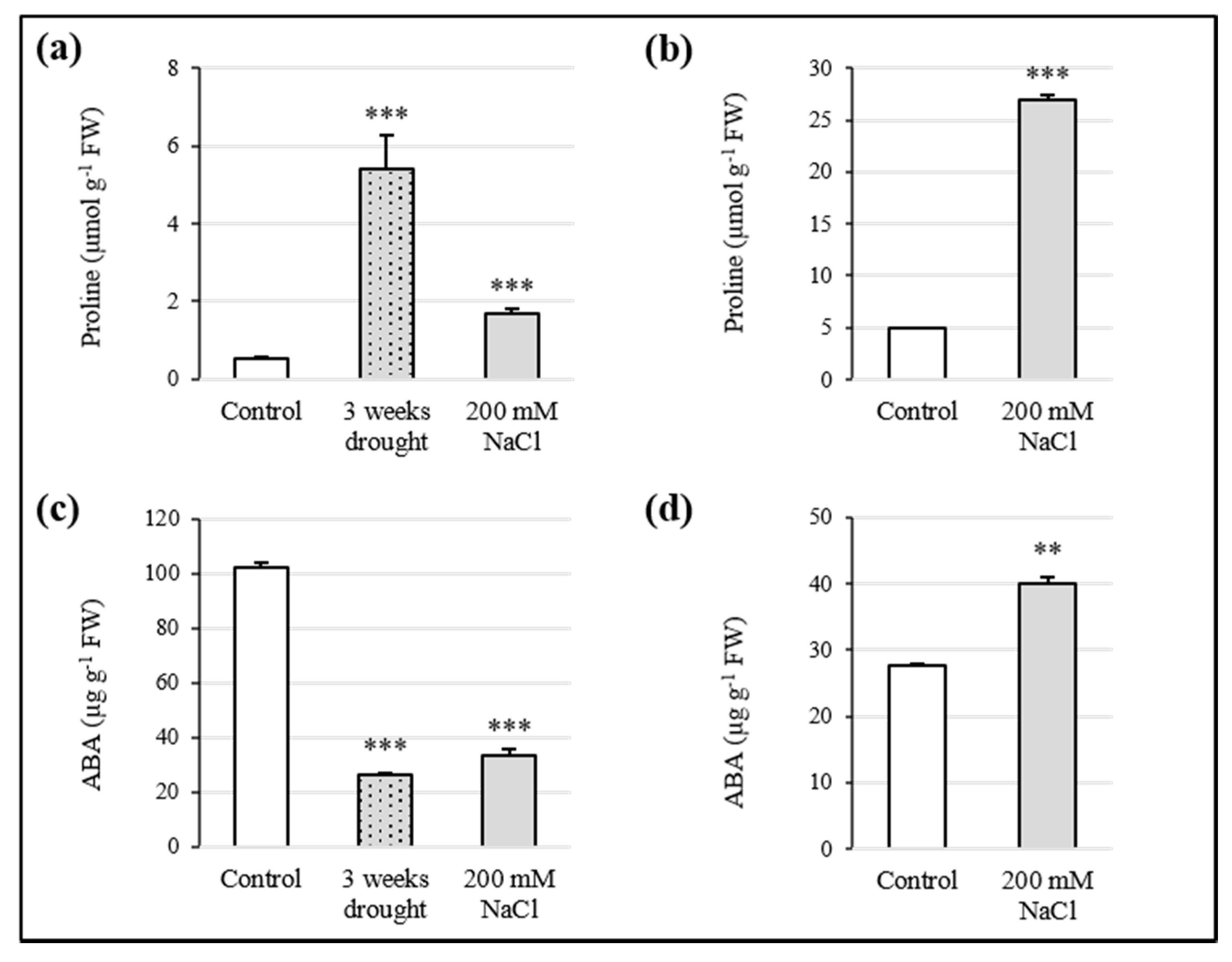
3.3. Proline and ABA Contribution to Osmotic Adjustment
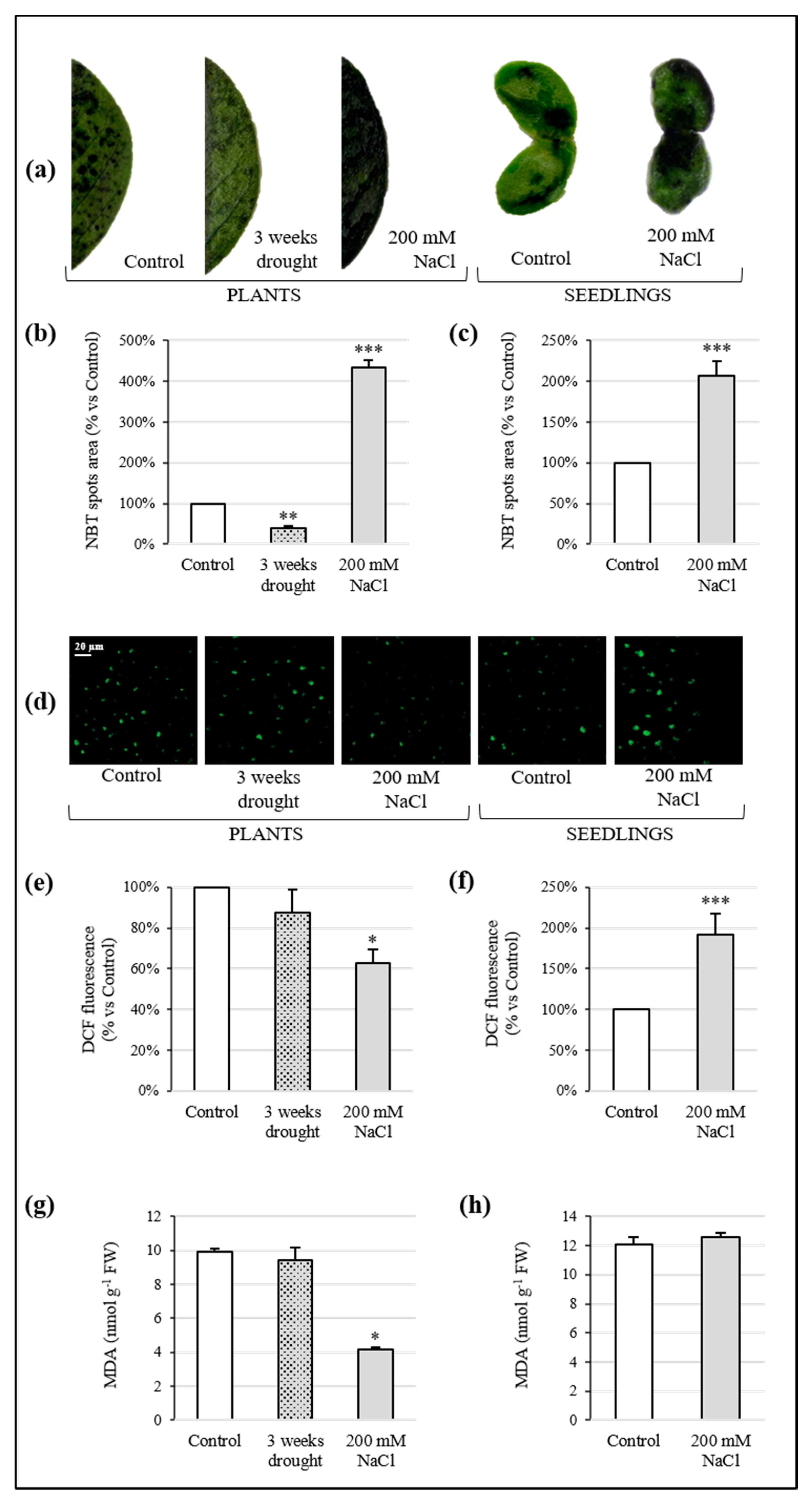
3.4. ROS Detection and Cell Membrane Integrity
3.5. Regulation of Non-Enzymatic Antioxidant Defence
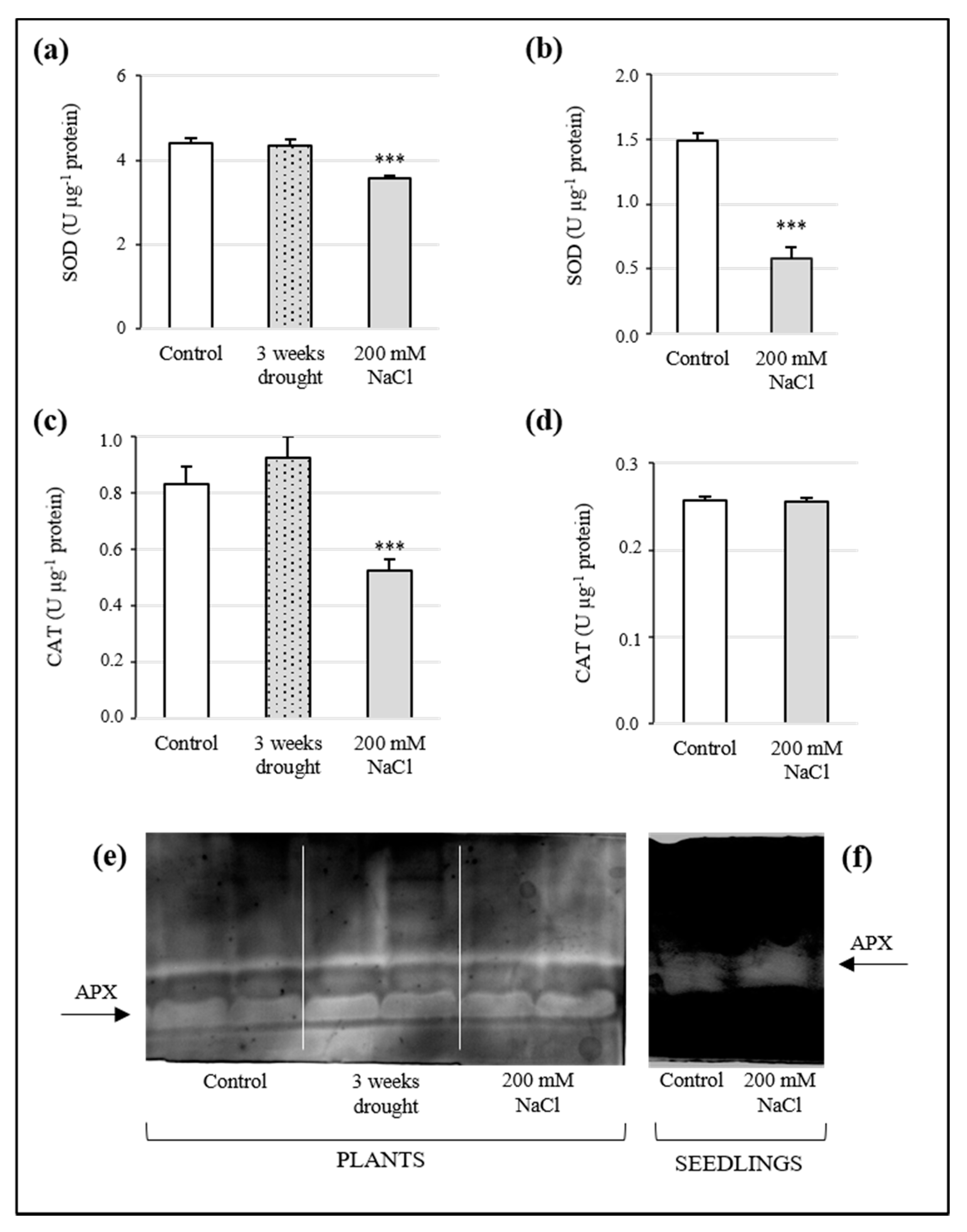
3.6. Analysis of Antioxidant Enzyme Activity
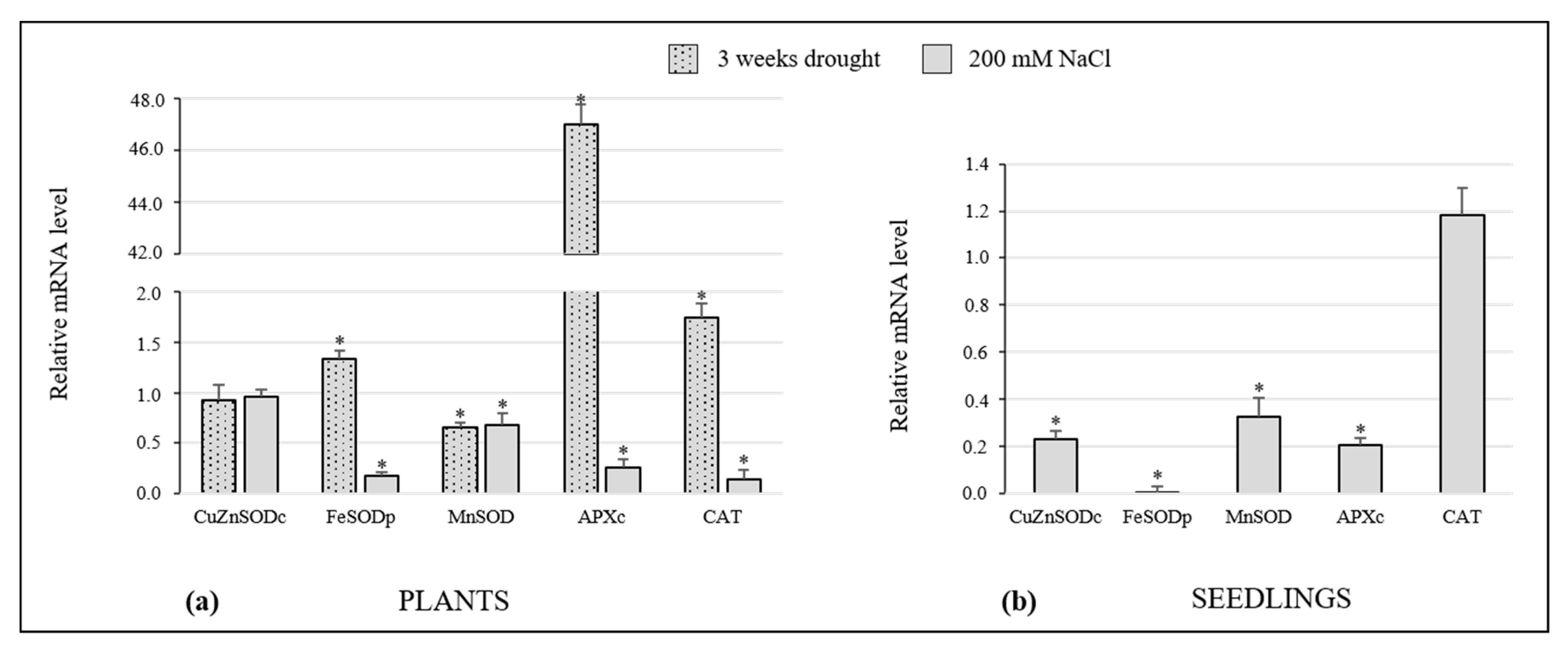
3.7. Expression of Genes Encoding for Antioxidant Enzymes
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- IPCC. Climate Change 2014: Synthesis Report. Contribution of Working Groups I, II and III to the Fifth Assessment Report of the Intergovernmental Panel on Climate Change; Core Writing Team, Pachauri, R.K., Meyer, L.A., Eds.; IPCC: Geneva, Switzerland, 2014. [Google Scholar]
- Montanarella, L. Trends in land degradation in Europe. In Climate and Land Degradation; Sivakumar, M.V., N’diangui, N., Eds.; Springer: Berlin, Germany, 2007; pp. 83–104. [Google Scholar]
- Salvati, L.; Ferrara, C. The local-scale impact of soil salinization on the socioeconomic context: An exploratory analysis in Italy. Catena 2015, 127, 312–322. [Google Scholar] [CrossRef]
- Fang, Y.; Xiong, L. General mechanisms of drought response and their application in drought resistance improvement in plants. Cell. Mol. Life Sci. 2015, 72, 673–689. [Google Scholar] [CrossRef] [PubMed]
- Munns, R. Comparative physiology of salt and water stress. Plant Cell Environ. 2002, 25, 239–250. [Google Scholar] [CrossRef]
- Chaves, M.M.; Flexas, J.; Pinheiro, C. Photosynthesis under drought and salt stress: Regulation mechanisms from whole plant to cell. Ann. Bot. 2009, 103, 551–560. [Google Scholar] [CrossRef] [PubMed]
- Hasegawa, P.; Bressan, R.; Zhu, J.; Bohnert, H. Plant cellular and molecular response to high salinity. Annu. Rev. Plant Biol. 2000, 51, 463–499. [Google Scholar] [CrossRef] [PubMed]
- Apel, K.; Hirt, H. Reactive oxygen species: Metabolism, oxidative stress, and signal transduction. Annu. Rev. Plant. Biol. 2004, 55, 373–399. [Google Scholar] [CrossRef] [PubMed]
- Gill, S.S.; Tuteja, N. Reactive oxygen species and antioxidant machinery in abiotic stress tolerance in crop plants. Plant Physiol. Biochem. 2010, 48, 909–930. [Google Scholar] [CrossRef]
- Tsang, E.W.T.; Bowler, C.; Hérouart, D.; Van Camp, W.; Villarroel, R.; Genetello, C.; Van Montagu, M.; Inzé, D. Differential regulation of superoxide dismutases in plants exposed to environmental stress. Plant Cell 1991, 3, 783–792. [Google Scholar] [PubMed]
- Mittler, R.; Zilinskas, B.A. Regulation of pea cytosolic ascorbate peroxidase and other antioxidant enzymes during the progression of drought stress and following recovery from drought. Plant J. 1994, 5, 397–405. [Google Scholar] [CrossRef]
- Hernández, J.A.; Jiménez, A.; Mullineaux, P.; Sevilla, F. Tolerance of pea (Pisum sativum L.) to long-term salt stress is associated with induction of antioxidant defences. Plant Cell Environ. 2000, 23, 853–862. [Google Scholar] [CrossRef]
- Luna, C.M.; Pastori, G.M.; Driscoll, S.; Groten, K.; Bernard, S.; Foyer, C.H. Drought controls on H2O2 accumulation, catalase (CAT) activity and CAT gene expression in wheat. J. Exp. Bot. 2005, 56, 417–423. [Google Scholar] [CrossRef]
- Rubio, M.C.; Bustos-Sanmamed, P.; Clemente, M.R.; Becana, M. Effects of salt stress on the expression of antioxidant genes and proteins in the model legume Lotus japonicus. New Phytol. 2009, 181, 851–859. [Google Scholar] [CrossRef] [PubMed]
- Araújo, S.S.; Beebe, S.; Crespi, M.; Delbreil, B.; González, E.M.; Gruber, V.; Lejeune-Henaut, I.; Link, W.; Monteros, M.J.; Prats, E.; et al. Abiotic stress responses in legumes: Strategies used to cope with environmental challenges. Crit. Rev. Plant. Sci. 2015, 34, 237–280. [Google Scholar] [CrossRef]
- Sulas, L.; Piluzza, G.; Salis, M.; Deligios, P.A.; Ledda, L.; Canu, S. Cropping systems sustainability: Inoculation and fertilisation effect on sulla performances in a new cultivation area. Ital. J. Agron. 2017, 12, 315–324. [Google Scholar] [CrossRef]
- Annicchiarico, P.; Abdelguerfi, A.; Ben Younes, M.; Bouzerzour, H.; Carroni, A.M.; Pecetti, L.; Tibaoui, G. Adaptation of sulla cultivars to contrasting Mediterranean environments. Aust. J. Agric. Res. 2008, 59, 702–706. [Google Scholar] [CrossRef]
- Nuti, M.P.; Casella, S.; Filippi, C.; Lepidi, A.A.; Galluzzi, R. Rhizobia as inoculant for field trials in marginal soils of middle- and northern-Italy. In Current Perspectives on Nitrogen Fixation; Newton, W., Gibson, A.H., Eds.; Academy of Science: Canberra, Australia, 1981; p. 539. [Google Scholar]
- Squartini, A.; Dazz, F.B.; Casella, S.; Nuti, M.P. The root nodule symbiosis between Rhizobium ’hedysari’ and its drought-tolerant host Hedysarum coronarium. Symbiosis 1993, 15, 227–238. [Google Scholar]
- Sulas, L.; Seddaiu, G.; Muresu, R.; Roggero, P.P. Nitrogen fixation of sulla under mediterranean conditions. Agron. J. 2009, 101, 1470–1478. [Google Scholar] [CrossRef]
- Di Marco, G.; Gismondi, A.; Canuti, L.; Scimeca, M.; Volpe, A.; Canini, A. Tetracycline accumulates in Iberis sempervirens L. through apoplastic transport inducing oxidative stress and growth inhibition. Plant Biol. 2014, 16, 792–800. [Google Scholar] [CrossRef]
- Luo, N.; Liu, J.; Yu, X.; Jiang, Y. Natural variation of drought response in Brachypodium distachyon. Physiol. Plant. 2011, 141, 19–29. [Google Scholar] [CrossRef] [PubMed]
- Lichtenthaler, H.K. Chlorophylls and carotenoids: Pigments of photosynthetic biomembranes. Methods Enzymol. 1987, 148, 350–382. [Google Scholar]
- Genty, B.; Briantais, J.M.; Baker, N.R. The relationship between the quantum yield of photosynthetic electron transport and quenching of chlorophyll fluorescence. Biochim. Biophys. Acta Gen. Subj. 1989, 990, 87–92. [Google Scholar] [CrossRef]
- Duan, Y.; Zhang, W.; Li, B.; Wang, Y.; Li, K.; Sodmergen, T.; Han, C.; Zhang, Y.; Li, X. An endoplasmic reticulum response pathway mediates programmed cell death of root tip induced by water stress in Arabidopsis. New Phytol. 2010, 186, 681–695. [Google Scholar] [CrossRef] [PubMed]
- Carillo, P.; Gibon, Y.; PrometheusWiki Contributors. Extraction and Determination of Proline. 2011. Available online: http://prometheuswiki.org/tiki-index.php?page=Extraction+and+determination+of+proline (accessed on 6 April 2020).
- Nakurte, I.; Keisa, A.; Rostoks, N. Development and validation of a reversed-phase liquid chromatography method for the simultaneous determination of indole-3-acetic acid, indole-3-pyruvic acid, and abscisic acid in Barley (Hordeum vulgare L.). J. Anal. Methods Chem. 2012, 2012, 103575. [Google Scholar] [CrossRef]
- Gismondi, A.; De Rossi, S.; Canuti, L.; Novelli, S.; Di Marco, G.; Fattorini, L.; Canini, A. From Robinia pseudoacacia L. nectar to Acacia monofloral honey: Biochemical changes and variation of biological properties. J. Sci. Food Agric. 2018, 98, 4312–4322. [Google Scholar] [CrossRef] [PubMed]
- Wohlgemuth, H.; Mittelstrass, K.; Kschieschan, S.; Bender, J.; Weigel, H.J.; Overmyer, K.; Kangasjärvi, J.; Sandermann, H.; Langebartels, C. Activation of an oxidative burst is a general feature of sensitive plants exposed to the air pollutant ozone. Plant Cell Environ. 2002, 25, 717–726. [Google Scholar] [CrossRef]
- Rodríguez-Serrano, M.; Romero-Puertas, M.C.; Zabalza, A.; Corpas, F.J.; Gómez, M.; Del Río, L.A.; Sandalio, L.M. Cadmium effect on oxidative metabolism of pea (Pisum sativum L.) roots. Imaging of reactive oxygen species and nitric oxide accumulation in vivo. Plant Cell Environ. 2006, 29, 1532–1544. [Google Scholar] [CrossRef] [PubMed]
- Heath, R.L.; Packer, L. Photoperoxidation in isolated chloroplasts. I. Kinetics and stoichiometry of fatty acid peroxidation. Arch. Biochem. Biophys. 1968, 125, 189–198. [Google Scholar] [CrossRef]
- Gismondi, A.; Serio, M.; Canuti, L.; Canini, A. Biochemical, antioxidant and antineoplastic properties of Italian saffron (Crocus sativus L.). Am. J. Plant Sci. 2012, 3, 1573–1580. [Google Scholar] [CrossRef]
- Sahoo, S.; Awasthi, J.P.; Sunkar, R.; Panda, S.K. Determining glutathione levels in plants. Plant Stress Toler. 2017, 1631, 273–277. [Google Scholar]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Beauchamp, C.; Fridovich, I. Superoxide dismutase: Improved assays and an assay applicable to acrylamide gels. Anal. Biochem. 1971, 44, 276–287. [Google Scholar] [CrossRef]
- Iwase, T.; Tajima, A.; Sugimoto, S.; Okuda, K.I.; Hironaka, I.; Kamata, Y.; Takada, K.; Mizunoe, Y. A simple assay for measuring catalase activity: A visual approach. Sci. Rep. 2013, 3, 3–6. [Google Scholar] [CrossRef]
- Mittler, R.; Zilinskas, B.A. Detection of ascorbate peroxidase activity in native gels by inhibition of the ascorbate-dependent reduction of nitroblue tetrazolium. Anal. Biochem. 1993, 212, 540–546. [Google Scholar] [CrossRef]
- Novelli, S.; Gismondi, A.; Di Marco, G.; Canuti, L.; Nanni, V.; Canini, A. Plant defense factors involved in Olea europaea resistance against Xylella fastidiosa infection. J. Plant Res. 2019, 132, 439–455. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCT method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Asada, K. Ascorbate peroxidase—A hydrogen peroxide-scavenging enzyme in plants. Physiol. Plantarum 1992, 85, 235–241. [Google Scholar] [CrossRef]
- Willekens, H.; Chamnongpol, S.; Davey, M.; Schraudner, M.; Langebartels, C.; Van Montagu, M.; Inzé, D.; Van Camp, W. Catalase is a sink for H2O2 and is indispensable for stress defence in C3 plants. EMBO J. 1997, 16, 4806–4816. [Google Scholar] [CrossRef] [PubMed]
- Farooq, M.; Gogoi, N.; Hussain, M.; Barthakur, S.; Paul, S.; Bharadwaj, N.; Migdadi, H.M.; Alghamdi, S.S.; Siddique, K.H.M. Effects, tolerance mechanisms and management of salt stress in grain legumes. Plant Physiol. Biochem. 2017, 118, 199–217. [Google Scholar] [CrossRef]
- Kaddour, R.; Sellami, N.; Chennaoui, H.; Nasri, N.; Baatour, O.; Mahmoudi, H.; Tarchoun, I.; Lachaâl, M.; Gruber, M.; Trifi, N. Correlation between salt tolerance and genetic diversity between Sulla carnosa and Sulla coronaria. Afr. J. Biotechnol. 2011, 10, 14355–14365. [Google Scholar]
- Parihar, P.; Singh, S.; Singh, R.; Singh, V.P.; Prasad, S.M. Effect of salinity stress on plants and its tolerance strategies: A review. Environ. Sci. Pollut. Res. 2015, 22, 4056–4075. [Google Scholar] [CrossRef]
- Li, H.; Li, X.; Zhang, D.; Liu, H.; Guan, K. Effects of drought stress on the seed germination and early seedling growth of the endemic desert plant Eremosparton songoricum (Fabaceae). EXCLI J. 2013, 12, 89–101. [Google Scholar]
- Ashraf, M.; Foolad, M.R. Roles of glycine betaine and proline in improving plant abiotic stress resistance. Environ. Exp. Bot. 2007, 59, 206–216. [Google Scholar] [CrossRef]
- Munns, R.; Tester, M. Mechanisms of salinity tolerance. Annu. Rev. Plant Biol. 2008, 59, 651–681. [Google Scholar] [CrossRef]
- Sharma, P.; Jha, A.B.; Dubey, R.S.; Pessarakli, M. Reactive oxygen species, oxidative damage, and antioxidative defense mechanism in plants under stressful conditions. J. Bot. 2012, 2012, 1–26. [Google Scholar] [CrossRef]
- Sharma, A.; Kumar, V.; Shahzad, B.; Ramakrishnan, M.; Singh Sidhu, G.P.; Bali, A.S.; Handa, N.; Kapoor, D.; Yadav, P.; Khanna, K.; et al. Photosynthetic response of plants under different abiotic stresses: A review. J. Plant Growth Regul. 2020, 39, 509–531. [Google Scholar] [CrossRef]
- Tuteja, N. Abscisic acid and abiotic stress signaling. Plant Signal. Behav. 2007, 2, 135–138. [Google Scholar] [CrossRef]
- He, T.; Cramer, G.R. Abscisic acid concentrations are correlated with leaf area reductions in two salt-stressed rapid-cycling Brassica species. Plant Soil 1996, 179, 25–33. [Google Scholar] [CrossRef]
- Montillet, J.L.; Chamnongpol, S.; Rustérucci, C.; Dat, J.; Van De Cotte, B.; Agnel, J.P.; Battesti, C.; Inzé, D.; Van Breusegem, F.; Triantaphylidès, C. Fatty acid hydroperoxides and H2O2 in the execution of hypersensitive cell death in tobacco leaves. Plant Physiol. 2005, 138, 1516–1526. [Google Scholar] [CrossRef]
- Bejaoui, F.; Salas, J.J.; Nouairi, I.; Smaoui, A.; Abdelly, C.; Martínez-Force, E.; Youssef, N.B. Changes in chloroplast lipid contents and chloroplast ultrastructure in Sulla carnosa and Sulla coronaria leaves under salt stress. J. Plant Physiol. 2016, 198, 32–38. [Google Scholar] [CrossRef]
- Mittler, R.; Vanderauwera, S.; Gollery, M.; Van Breusegem, F. Reactive oxygen gene network of plants. Trends Plant Sci. 2004, 9, 490–498. [Google Scholar] [CrossRef]
- Choudhury, S.; Panda, P.; Sahoo, L.; Panda, S.K. Reactive oxygen species signaling in plants under abiotic stress. Plant Signal. Behav. 2013, 8, e23681. [Google Scholar] [CrossRef]
- Foyer, C.H.; Noctor, G. Oxidant and antioxidant signalling in plants: A re-evaluation of the concept of oxidative stress in a physiological context. Plant Cell Environ. 2005, 28, 1056–1071. [Google Scholar] [CrossRef]
- Nascimento, N.C.; Fett-Neto, A.G. Plant secondary metabolism and challenges in modifying its operation: An overview. Methods Mol. Biol. 2010, 643, 1–13. [Google Scholar]
- May, M.J.; Vernoux, T.; Leaver, C.; Van Montagu, M.; Inzé, D. Glutathione homeostasis in plants: Implications for environmental sensing and plant development. J. Exp. Bot. 1998, 49, 649–667. [Google Scholar] [CrossRef]
- Gill, S.S.; Anjum, N.A.; Hasanuzzaman, M.; Gill, R.; Trivedi, D.K.; Ahmad, I.; Pereira, E.; Tuteja, N. Glutathione and glutathione reductase: A boon in disguise for plant abiotic stress defense operations. Plant Physiol. Biochem. 2013, 70, 204–212. [Google Scholar] [CrossRef]
- Sewelam, N.; Jaspert, N.; Van Der Kelen, K.; Tognetti, V.B.; Schmitz, J.; Frerigmann, H.; Stahl, E.; Zeier, J.; Van Breusegem, F.; Maurino, V.G. Spatial H2O2 signaling specificity: H2O2 from chloroplasts and peroxisomes modulates the plant transcriptome differentially. Mol. Plant 2014, 7, 1191–1210. [Google Scholar] [CrossRef]
- Caverzan, A.; Passaia, G.; Rosa, S.B.; Ribeiro, C.W.; Lazzarotto, F.; Margis-Pinheiro, M. Plant responses to stresses: Role of ascorbate peroxidase in the antioxidant protection. Genet. Mol. Biol. 2012, 35, 1011–1019. [Google Scholar] [CrossRef] [PubMed]
- Hasanuzzaman, M.; Fujita, M. Selenium pretreatment upregulates the antioxidant defense and methylglyoxal detoxification system and confers enhanced tolerance to drought stress in rapeseed seedlings. Biol. Trace Elem. Res. 2011, 143, 1758–1776. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Treatment | Root Length (cm) | Hypocotyl Length (cm) | Cotyledon Width (mm) |
|---|---|---|---|
| Control | 6.68 ± 0.33 | 2.03 ± 0.07 | 4.74 ± 0.15 |
| 100 mM | 5.78 ± 0.42 | 1.85 ± 0.08 | 4.77 ± 0.15 |
| 150 mM | 3.51 ± 0.51 *** | 1.32 ± 0.12 *** | 4.39 ± 0.16 |
| 200 mM | 3.26 ± 0.47 *** | 0.87 ± 0.11 *** | 4.52 ± 0.17 |
| Pigment (µg 100 mg−1 FW) | |||
|---|---|---|---|
| Plants (drought) | Chl a | Chl b | Carotenoids |
| Control | 81.58 ± 7.34 | 23.17 ± 2.40 | 19.95 ± 2.02 |
| 1 week | 94.03 ± 2.02 | 22.69 ± 2.84 | 27.06 ± 1.27 * |
| 2 weeks | 103.93 ± 1.77 * | 25.48 ± 1.77 | 28.57 ± 0.94 ** |
| 3 weeks | 43.55 ± 8.54 ** | 10.86 ± 1.95 ** | 9.04 ± 2.30 ** |
| Plants (NaCl) | |||
| Control | 81.58 ± 7.34 | 23.17 ± 2.40 | 19.95 ± 2.02 |
| 100 mM | 31.05 ± 3.42 *** | 7.61 ± 1.14 *** | 7.05 ± 0.63 *** |
| 150 mM | 31.56 ± 0.18 ** | 7.63 ± 0.18 ** | 9.16 ± 0.07 * |
| 200 mM | 30.10 ± 1.29 *** | 7.91 ± 0.12 *** | 6.75 ± 0.62 *** |
| Seedlings (NaCl) | |||
| Control | 2.56 ± 0.46 | 1.46 ± 0.20 | 0.89 ± 0.12 |
| 100 mM | 4.86 ± 0.54 ** | 1.98 ± 0.17 | 1.44 ± 0.13 ** |
| 150 mM | 4.80 ± 0.74 * | 1.80 ± 0.20 | 1.35 ± 0.19 * |
| 200 mM | 5.79 ± 0.67 *** | 2.15 ± 0.20 * | 1.58 ± 0.16 ** |
| Phenolics (μg GAE 100 mg−1 FW) | ||
|---|---|---|
| Plants | ||
| Control | 30.80 ± 5.09 | |
| 3 weeks drought | 38.21 ± 9.88 | |
| 200 mM NaCl | 12.96 ± 1.96 ** | |
| Seedlings | ||
| Control | 19.73 ± 1.50 | |
| 200 mM NaCl | 45.91 ± 4.85 *** | |
| Glutathione (μM 50 mg−1 FW) | ||
| Total glutathione | GSSG/GSH | |
| Plants | ||
| Control | 137.56 ± 14.49 | 0.21 |
| 3 weeks drought | 140.28 ± 13.05 | 0.66 * |
| 200 mM NaCl | 130.16 ± 9.68 | 0.80 * |
| Seedlings | ||
| Control | 156.09 ± 12.13 | 0.14 |
| 200 mM NaCl | 96.38 ± 6.31 ** | 0.63 ** |
| Ascorbic acid (μg g−1 FW) | ||
| Plants | ||
| Control | 36.7 ± 1.55 | |
| 3 weeks drought | 41.2 ± 2.12 | |
| 200 mM NaCl | 43.7 ± 2.33 | |
| Seedlings | ||
| Control | 30.2 ± 1.52 | |
| 200 mM NaCl | 27.9 ± 1.32 | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
De Rossi, S.; Di Marco, G.; Bruno, L.; Gismondi, A.; Canini, A. Investigating the Drought and Salinity Effect on the Redox Components of Sulla coronaria (L.) Medik. Antioxidants 2021, 10, 1048. https://doi.org/10.3390/antiox10071048
De Rossi S, Di Marco G, Bruno L, Gismondi A, Canini A. Investigating the Drought and Salinity Effect on the Redox Components of Sulla coronaria (L.) Medik. Antioxidants. 2021; 10(7):1048. https://doi.org/10.3390/antiox10071048
Chicago/Turabian StyleDe Rossi, Silvia, Gabriele Di Marco, Laura Bruno, Angelo Gismondi, and Antonella Canini. 2021. "Investigating the Drought and Salinity Effect on the Redox Components of Sulla coronaria (L.) Medik" Antioxidants 10, no. 7: 1048. https://doi.org/10.3390/antiox10071048
APA StyleDe Rossi, S., Di Marco, G., Bruno, L., Gismondi, A., & Canini, A. (2021). Investigating the Drought and Salinity Effect on the Redox Components of Sulla coronaria (L.) Medik. Antioxidants, 10(7), 1048. https://doi.org/10.3390/antiox10071048