
Antioxidant, Anti-Inflammatory, and Cytotoxic Activity of Extracts from Some Commercial Apple Cultivars in Two Colorectal and Glioblastoma Human Cell Lines


 ,
, 

Abstract
:
1. Introduction
2. Materials and Methods
2.1. Plant Materials
2.2. Cell Cultures
2.3. Chemicals and Solvents
2.4. Preparation of Apple Samples
2.5. Preparation of Dry Phenolic and Triterpene Extracts
2.6. Determination of Phenolic Compounds by HPLC-PDA Method
2.7. Determination of Triterpene Compounds by HPLC-PDA Method
2.8. Antioxidant Activity Assays
2.9. Anti-Inflammatory Activities Assays
2.10. Cell Viability Assay
2.11. Cell Colony Formation
2.12. Spheroid Growth Assay
2.13. Statistical Analysis
3. Results
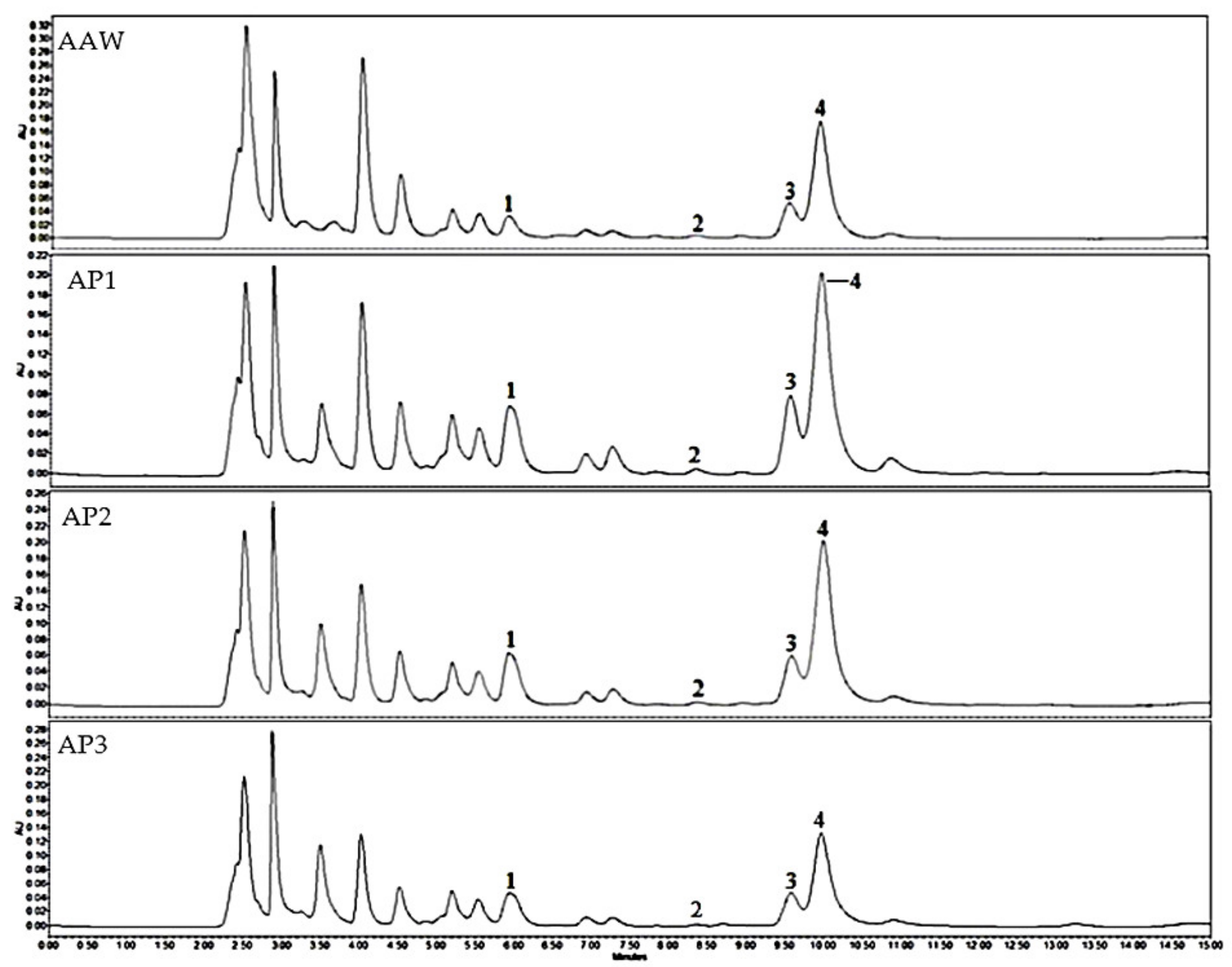
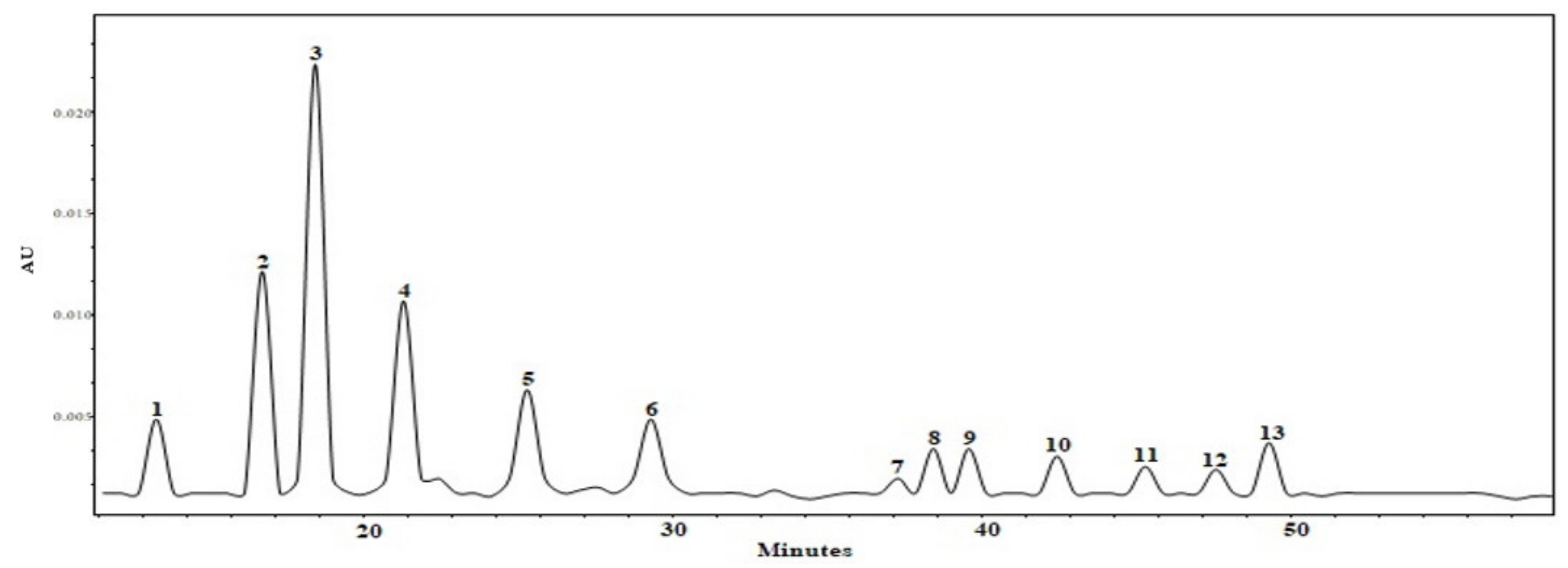
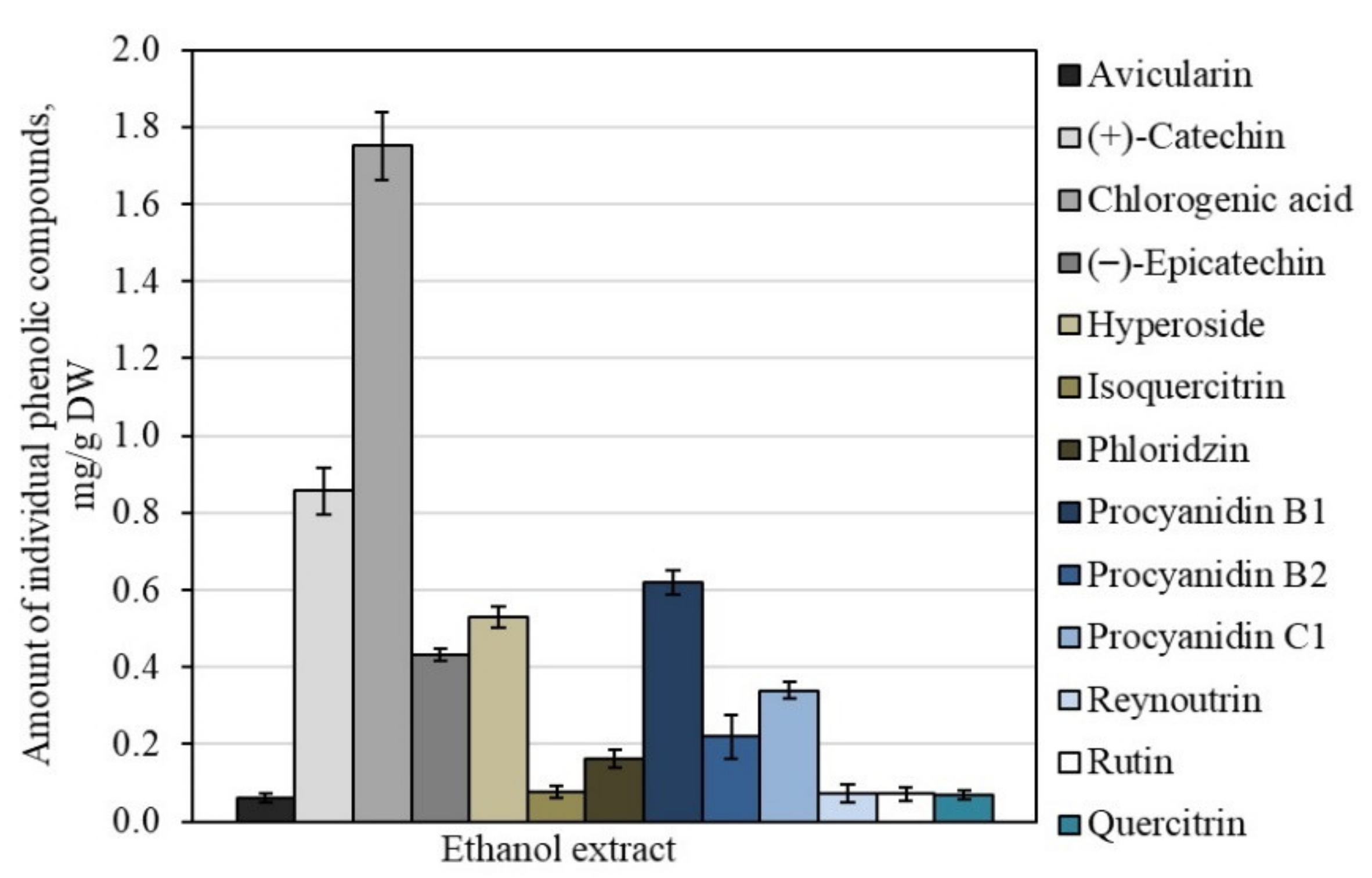
3.1. Qualitative and Quantitative Analysis of Triterpene and Phenolic Compounds
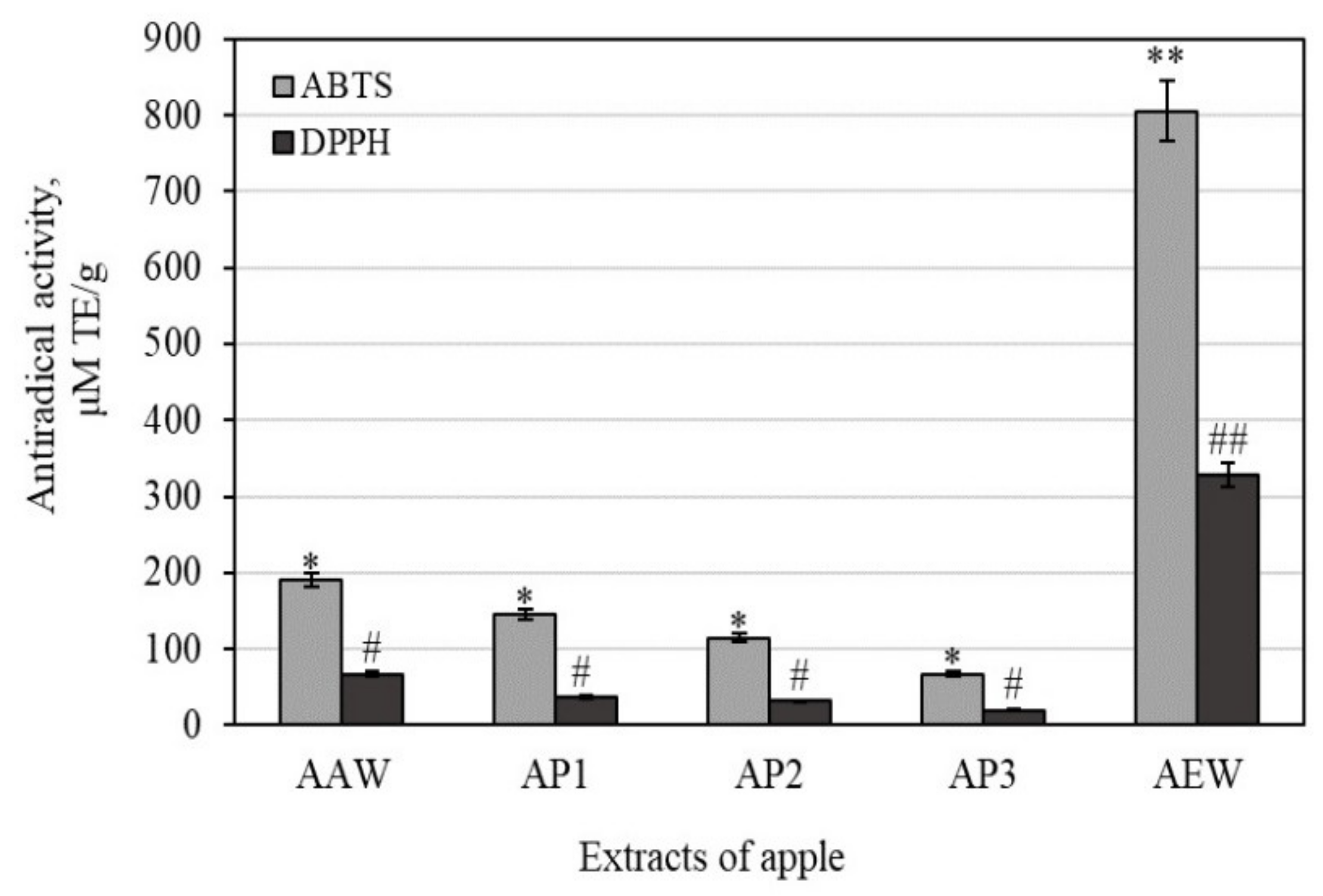
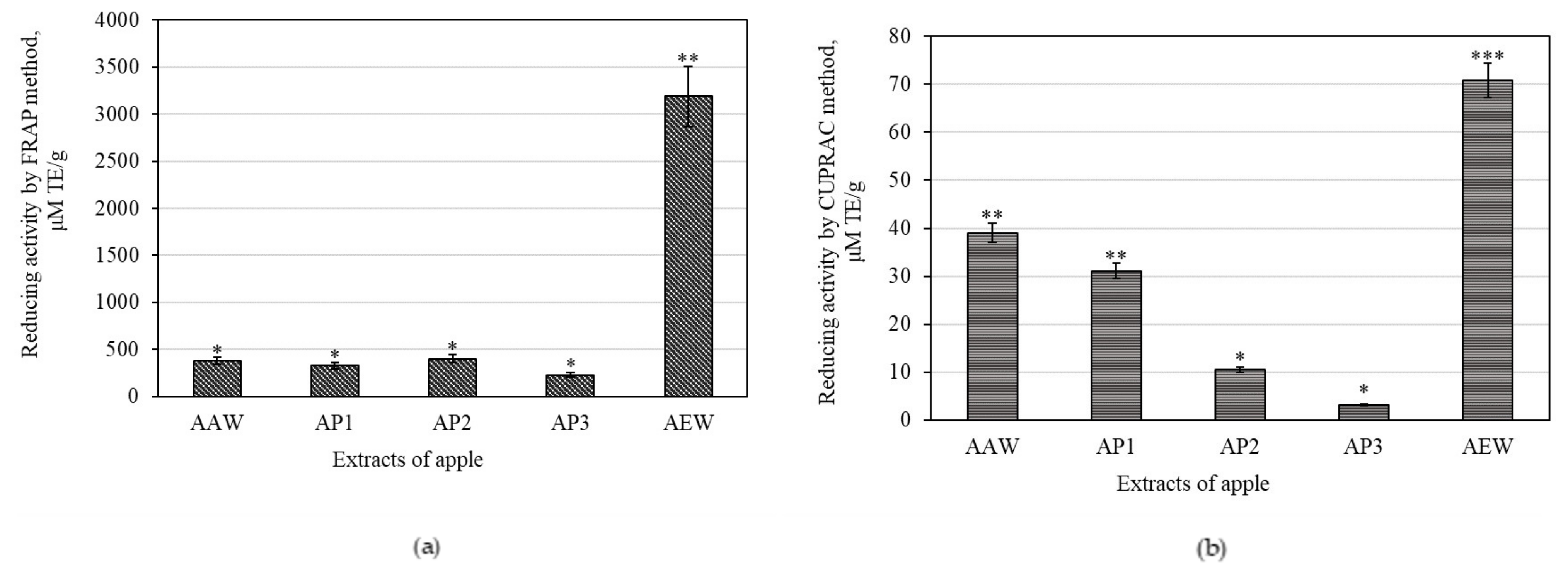
3.2. Antioxidant Activities of Whole Apple and Apple Peel Extracts
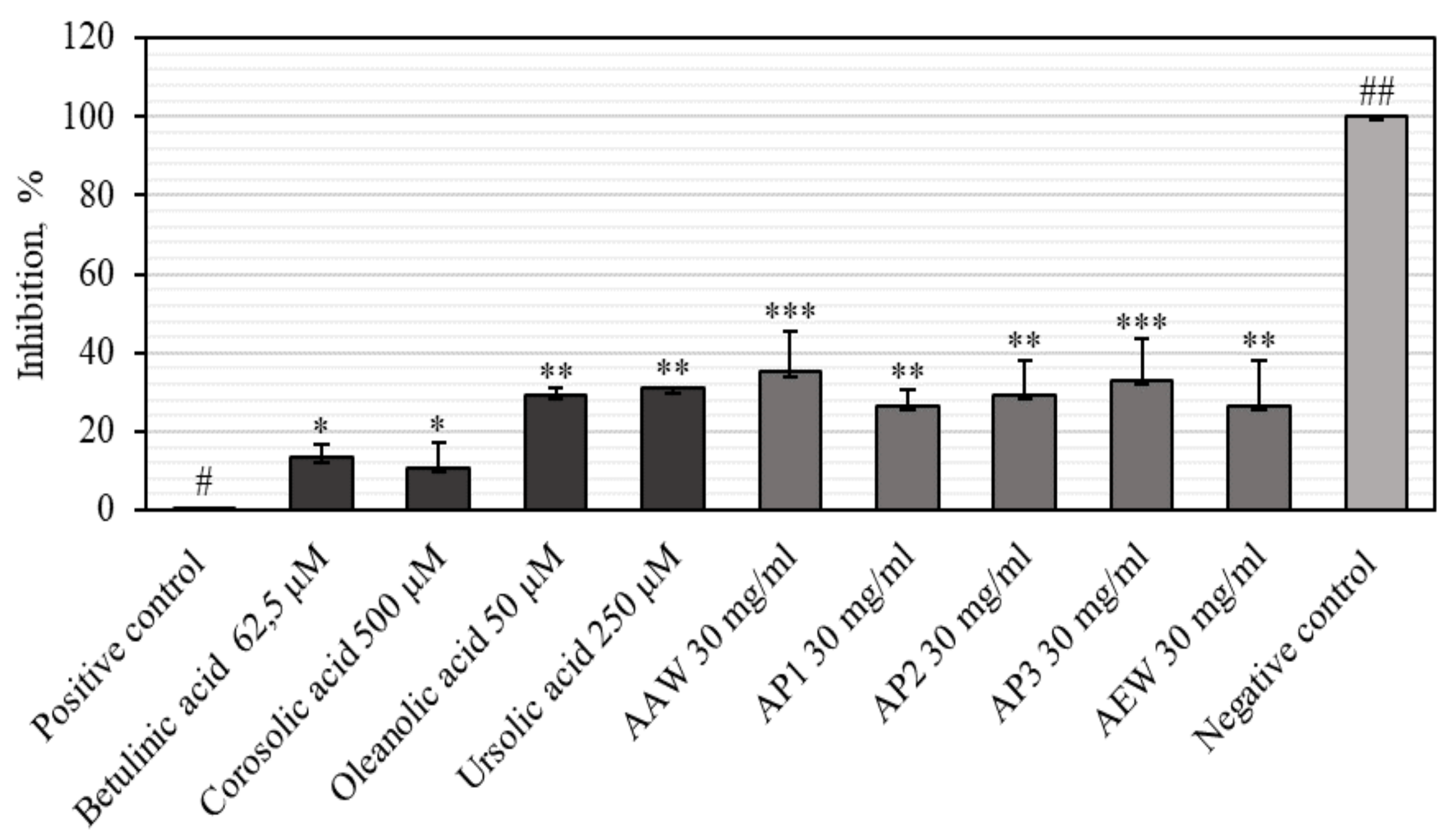
3.3. Anti-Inflammatory Activities of Apple Extracts and Individual Triterpenes
3.4. Cytotoxic Activity of Apple Extracts and Individual Triterpenes
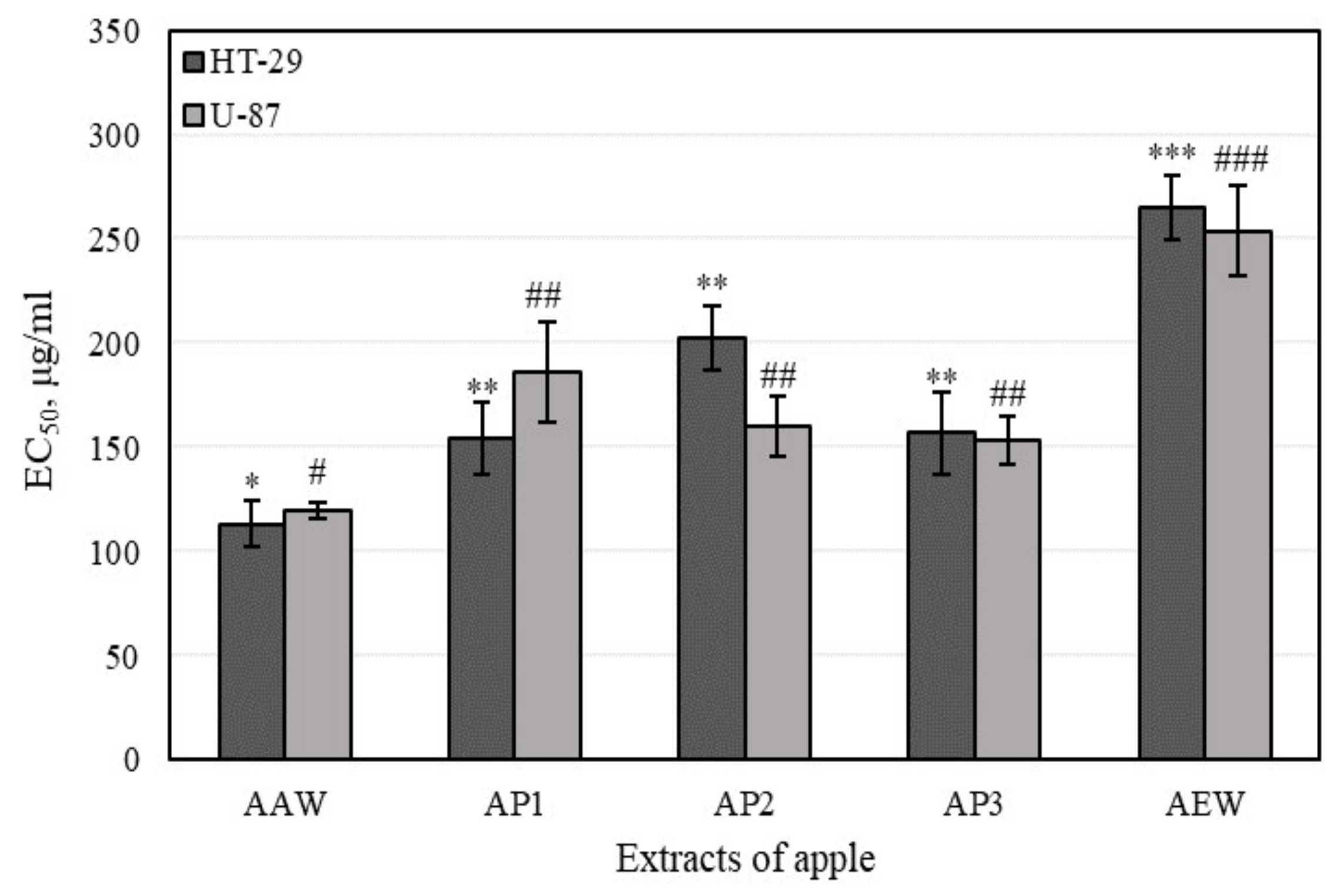
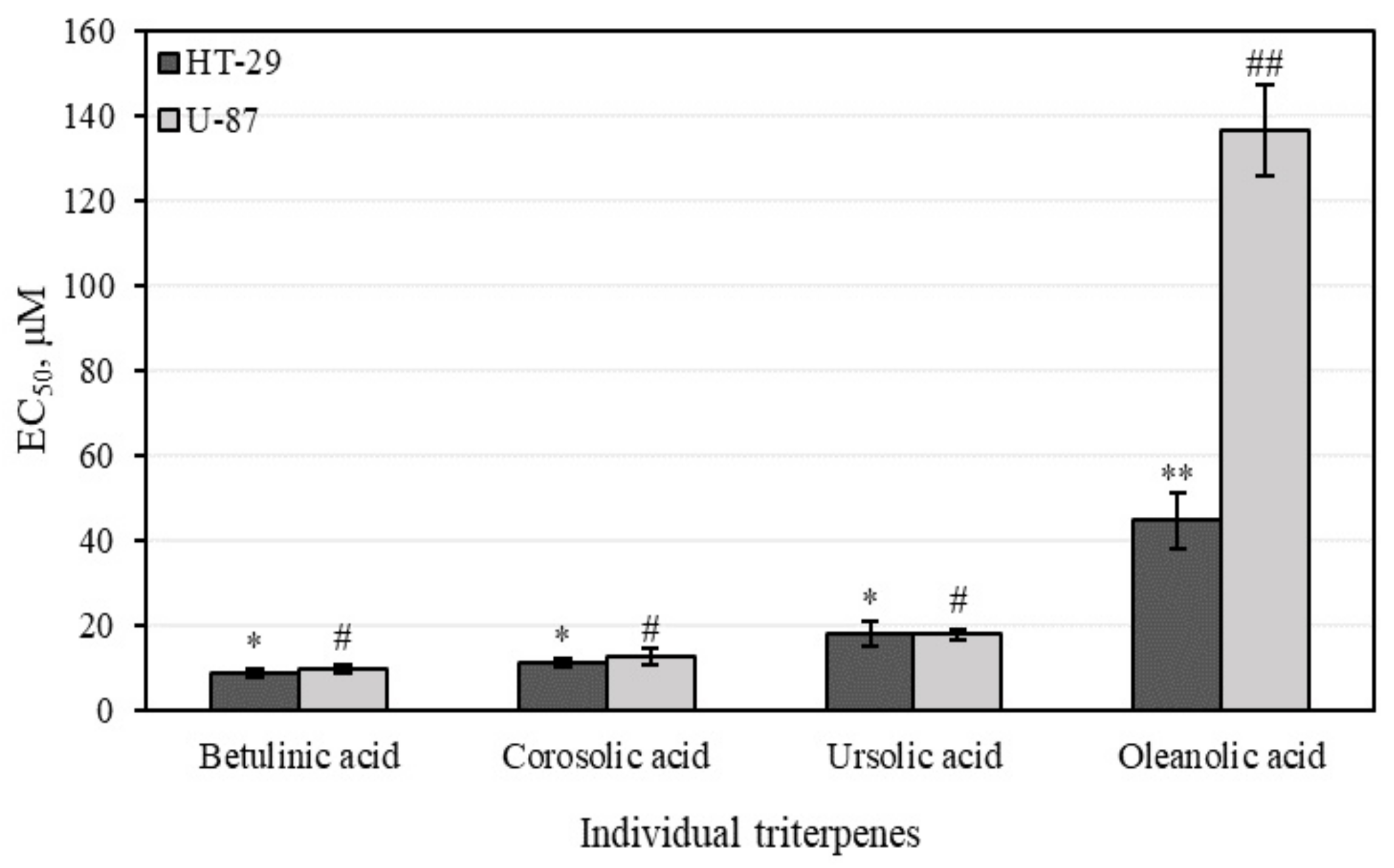
3.4.1. Cell Viability Determined by MTT Assays
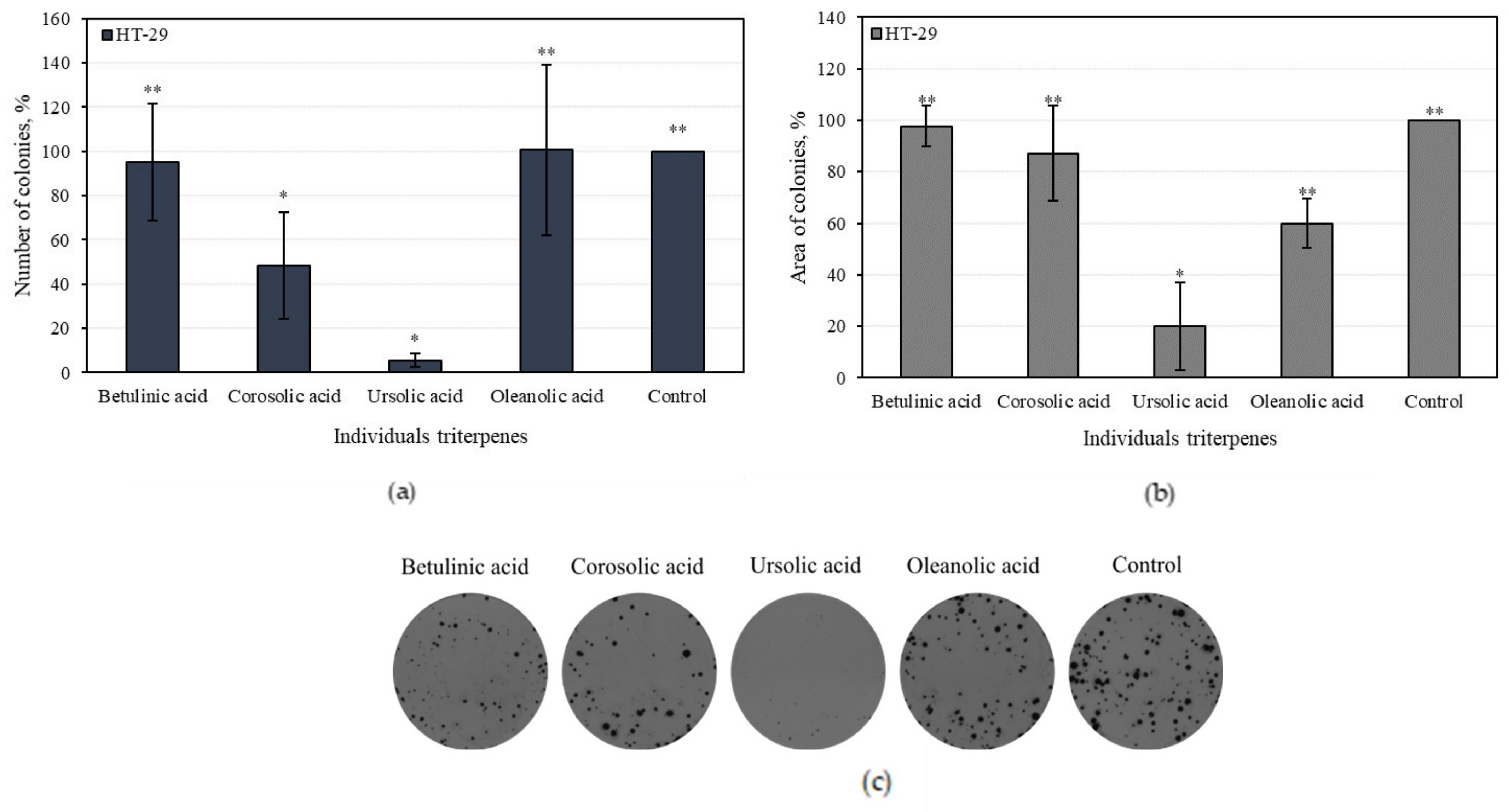
3.4.2. Effect of Individual Triterpenes on HT-29 Colony-Forming
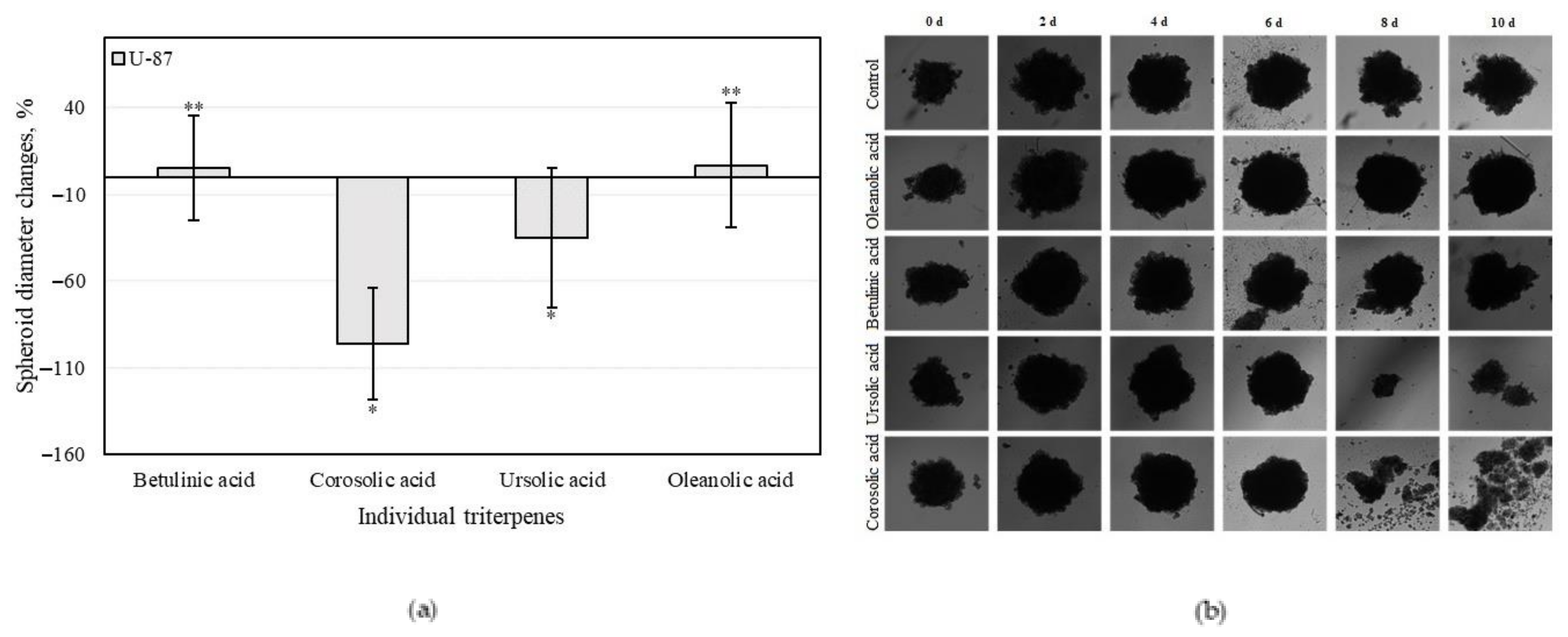
3.4.3. Triterpenes on U-87 Cell 3D Spheroid Growth
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Food and Agriculture Organization. FAOSTAT. Available online: http://www.fao.org/faostat/en/#data/QL (accessed on 23 May 2021).
- Wu, J.; Gao, H.; Zhao, L.; Liao, X.; Chen, F.; Wang, Z.; Hu, X. Chemical compositional characterization of some apple cultivars. Food Chem. 2007, 103, 88–93. [Google Scholar] [CrossRef]
- Morresi, C.; Cianfruglia, L.; Armeni, T.; Mancini, F.; Tenore, G.C.; D’Urso, E.; Micheletti, A.; Ferretti, G.; Bacchetti, T. Polyphenolic compounds and nutraceutical properties of old and new apple cultivars. J. Food Biochem. 2018, 42, e12641. [Google Scholar] [CrossRef]
- Oszmianski, J.; Lachowicz, S.; Gławdel, E.; Cebulak, T.; Ochmian, I. Determination of phytochemical composition and antioxidant capacity of 22 old apple cultivars grown in Poland. Eur. Food Res. Technol. 2017, 244, 647–662. [Google Scholar] [CrossRef] [Green Version]
- Ferretti, G.; Turco, I.; Bacchetti, T. Apple as a source of dietary phytonutrients: Bioavailability and evidence of protective effects against human cardiovascular disease. Food Nutr. Sci. 2014, 5, 1234–1246. [Google Scholar] [CrossRef] [Green Version]
- Nour, V.; Trandafir, I.; Ionica, M.E. Compositional characteristics of fruits of several apple (Malus domestica Borkh.) cultivars. Not. Bot. Hort. Agrobot. 2010, 38, 228–233. [Google Scholar]
- Berni, R.; Cantini, C.; Guarnieri, M.; Nepi, M.; Hausman, J.-F.; Guerriero, G.; Romi, M.; Cai, G. Nutraceutical characteristics of ancient Malus × domestica Borkh. fruits recovered across Siena in Tuscany. Medicines 2019, 6, 27. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Boyer, J.; Liu, R.H. Apple phytochemicals and their health benefits. Nutr. J. 2004, 3, 5. [Google Scholar] [CrossRef] [Green Version]
- Pandey, K.B.; Rizvi, S.I. Plant polyphenols as dietary antioxidants in human health and disease. Oxid. Med. Cell. Longev. 2009, 2, 270–278. [Google Scholar] [CrossRef] [Green Version]
- Allouche, Y.; Beltrán, G.; Gaforio, J.J.; Uceda, M.; Mesa, M.D. Antioxidant and antiatherogenic activities of pentacyclic triterpenic diols and acids. Food Chem. Toxicol. 2010, 48, 2885–2890. [Google Scholar] [CrossRef] [PubMed]
- Jeong, J.W.; Shim, J.J.; Choi, I.D.; Kim, S.H.; Ra, J.; Ku, H.K.; Lee, D.E.; Kim, T.Y.; Jeung, W.; Lee, J.H.; et al. Apple pomace extract improves endurance in exercise performance by increasing strength and weight of skeletal muscle. J. Med. Food 2015, 18, 1380–1386. [Google Scholar] [CrossRef] [PubMed]
- Waldbauer, K.; Seiringer, G.; Nguyen, D.L.; Winkler, J.; Blaschke, M.; McKinnon, R.; Urban, E.; Ladurner, A.; Dirsch, V.M.; Zehl, M.; et al. Triterpenoic acids from apple pomace enhance the activity of the endothelial nitric oxide synthase (eNOS). J. Agric. Food Chem. 2016, 64, 185–194. [Google Scholar] [CrossRef]
- Gerhauser, C. Cancer chemopreventive potential of apples, apple juice, and apple components. Planta Med. 2008, 74, 1608–1624. [Google Scholar] [CrossRef] [Green Version]
- Zhang, C.; Wang, C.; Li, W.; Wu, R.; Guo, Y.; Cheng, D.; Yang, Y.; Androulakis, I.P.; Kong, A.N. Pharmacokinetics and pharmacodynamics of the triterpenoid ursolic acid in regulating the anti-oxidant, anti-inflammatory and epigenetic gene responses in rat leukocytes. Mol. Pharm. 2017, 14, 3709–3717. [Google Scholar] [CrossRef] [Green Version]
- Chen, C.; Kong, A.N. Dietary chemopreventive compounds and ARE/EpRE signaling. Free Radic. Biol. Med. 2004, 36, 1505–1516. [Google Scholar] [CrossRef] [PubMed]
- Baldelli, G.; De Santi, M.; Fraternale, D.; Brandi, G.; Fanelli, M.; Schiavano, G.F. Chemopreventive potential of apple pulp callus against colorectal cancer cell proliferation and tumorigenesis. J. Med. Food 2019, 22, 614–622. [Google Scholar] [CrossRef] [Green Version]
- Hartheimer, J.S.; Park, S.; Rao, S.S.; Kim, Y. Targeting hyaluronan interactions for glioblastoma stem cell therapy. Cancer Microenviron. 2019, 12, 47–56. [Google Scholar] [CrossRef]
- Chen, J.W.E.; Pedron, S.; Shyu, P.; Hu, Y.; Sarkaria, J.N.; Harley, B.A.C. Influence of hyaluronic acid transitions in tumor microenvironment on glioblastoma malignancy and invasive behavior. Front. Mater. Sci. 2018, 5, 1–11. [Google Scholar] [CrossRef]
- Kimlin, C.; Casagrande, G.; Virador, V.M. In vitro threedimensional (3D) models in cancer research: An update. Mol. Carcinog. 2013, 52, 167–182. [Google Scholar] [CrossRef]
- Herrera-Perez, H.M.; Voytik-Harbin, S.L.; Rickus, J.L. Extracellular matrix properties regulate the migratory response of glioblastoma stem cells in three-dimensional culture. Tissue Eng. Part A 2015, 21, 2572–2582. [Google Scholar] [CrossRef] [PubMed]
- Lv, D.; Yu, S.C.; Ping, Y.F.; Wu, H.; Zhao, X.; Zhang, H.; Cui, Y.; Chen, B.; Zhang, X.; Dai, J.; et al. A three-dimensional collagen scaffold cell culture system for screening anti-glioma therapeutics. Oncotarget 2016, 7, 56904–56914. [Google Scholar] [CrossRef] [Green Version]
- Kohi, S.; Sato, N.; Koga, A.; Hirata, K.; Harunari, E.; Igarashi, Y. Hyaluromycin, a novel hyaluronidase inhibitor, attenuates pancreatic cancer cell migration and proliferation. J. Oncol. 2016, 2016, 9063087. [Google Scholar] [CrossRef] [PubMed]
- Stern, R.; Asari, A.A.; Sugahara, K.N. Hyaluronan fragments: An informationrich system. Eur. J. Cell Biol. 2006, 85, 699–715. [Google Scholar] [CrossRef]
- Bouga, H.; Tsouros, I.; Bounias, D.; Kyriakopoulou, D.; Stavropoulos, M.S.; Papageorgakopoulou, N.; Theocharis, D.A.; Vynios, D.H. Involvement of hyaluronidases in colorectal cancer. BMC Cancer 2010, 10, 499. [Google Scholar] [CrossRef] [Green Version]
- Girish, K.S.; Kemparaju, K.; Nagaraju, S.; Vishwanath, B.S. Hyaluronidase inhibitors: A biological and therapeutic perspective. Curr. Med. Chem. 2009, 16, 2261–2288. [Google Scholar] [CrossRef] [PubMed]
- Butkevičiūtė, A.; Liaudanskas, M.; Kviklys, D.; Zymonė, Z.; Raudonis, R.; Viškelis, J.; Uselis, N.; Janulis, V. Detection and analysis of triterpenic compounds in apple extracts. Int. J. Food Prop. 2018, 21, 1716–1727. [Google Scholar] [CrossRef] [Green Version]
- Liaudanskas, M.; Viskelis, P.; Kviklys, D.; Raudonis, R.; Janulis, V. A comparative study of phenolic content in apple fruits. Int. J. Food Prop. 2015, 18, 945–953. [Google Scholar] [CrossRef]
- Brand-Williams, W.; Cuvelier, M.E.; Berset, C. Use of a free radical method to evaluate antioxidant activity. Food Sci. Technol. 1995, 28, 25–30. [Google Scholar] [CrossRef]
- Benzie, I.F.F.; Strain, J.J. Ferric reducing/antioxidant power assay: Direct measure of total antioxidant activity of biological fluids and modified version for simultaneous measurement of total antioxidant power and ascorbic acid concentration. Methods Enzymol. 1998, 299, 15–27. [Google Scholar]
- Özyürek, M.; GüçLü, K.; Tütem, E.; Başkan, K.S.; Erçağ, E.; Çelik, S.E.; Baki, S.; Yıldız, L.; Karaman, Ş.; Apak, R. A comprehensive review of CUPRAC methodology. Anal. Methods 2011, 3, 2439. [Google Scholar] [CrossRef]
- El-Guendouz, S.; Aazza, S.; Lyoussi, B.; Antunes, M.D.; Faleiro, M.L.; Miguel, M.G. Anti-acetylcholinesterase, antidiabetic, anti-inflammatory, antityrosinase and antixanthine oxidase activities of Moroccan propolis. Int. J. Food Sci. Technol. 2016, 51, 1762–1773. [Google Scholar] [CrossRef] [Green Version]
- Grigalius, I.; Petrikaite, V. Relationship between antioxidant and anticancer Activity of trihydroxyflavones. Molecules 2017, 22, 2169. [Google Scholar] [CrossRef] [Green Version]
- Čekanavičius, V.; Murauskas, G. Applied Regression Analysis in Social Research; Vilnius University Press: Vilnius, Lithuania, 2014; p. 124. [Google Scholar]
- Hyson, D.A. A comprehensive review of apples and apple components and their relationship to human health. Adv. Nutr. 2011, 2, 408–420. [Google Scholar] [CrossRef]
- Siani, A.C.; Nakamura, M.J.; Santos, D.S.; Mazzei, J.L.; Nascimento, A.C.; Valente, L.M.M. Efficiency and selectivity of triterpenic acid extraction from decoctions and tinctures prepared from apple peels. Pharmacogn. Mag. 2014, 10, 225–231. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Farneti, B.; Masuero, D.; Costa, F.; Magnago, P.; Malnoy, M.; Costa, G.; Vrhovsek, U.; Mattivi, F. Is there room for improving the nutraceutical composition of apple? J. Agric. Food Chem. 2015, 63, 2750–2759. [Google Scholar] [CrossRef] [PubMed]
- Sut, S.; Zengin, G.; Maggi, F.; Malagoli, M.; Dall’Acqua, S. Triterpene acid and phenolics from ancient apples of Friuli Venezia Giulia as nutraceutical ingredients: LC-MS study and in vitro activities. Molecules 2019, 24, 1109. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Poirier, B.C.; Buchanan, D.A.; Mattheis, J.; Rudell, D. Differential partitioning of triterpenes and triterpene esters in apple peel. J. Agric. Food Chem. 2018, 66, 1800–1806. [Google Scholar] [CrossRef] [PubMed]
- Contessa, C.; Botta, R. Comparison of physicochemical traits of red-fleshed, commercial and ancient apple cultivars. Hort. Sci. 2016, 43, 159–166. [Google Scholar] [CrossRef] [Green Version]
- Tsao, R.; Yang, R.; Young, A.J.C.; Zhu, H. Polyphenolic profiles in eight apple cultivars using high-performance liquid chromatography (HPLC). J. Agric. Food Chem. 2003, 51, 6347–6353. [Google Scholar] [CrossRef]
- Duda-Chodak, A.; Tarko, T.; Satora, P.; Sroka, P.; Tuszy’nski, T. The profile of polyphenols and antioxidant properties of selected apple cultivars grown in Poland. J. Fruit Ornam. Plant Res. 2010, 18, 39–50. [Google Scholar]
- De Paepe, D.; Valkenborg, D.; Noten, B.; Servaes, K.; Diels, L.; De Loose, M. Variability of the phenolic profiles in the fruits from old, recent and new apple cultivars cultivated in Belgium. Metabolomics 2015, 11, 739–752. [Google Scholar] [CrossRef]
- Simmonds, M.S.J.; Howes, M.J.R. Profile of compounds in different cultivars of apple (Malus × domestica). In Nutritional Composition of Fruit Cultivars; Elsevier: Amsterdam, The Netherlands, 2016; Chapter 1; pp. 1–18. [Google Scholar]
- Raudonė, L.; Raudonis, R.; Liaudanskas, M.; Janulis, V.; Viskelis, P. Phenolic antioxidant profiles in the whole fruit, flesh and peel of apple cultivars grown in Lithuania. Sci. Hortic. 2017, 216, 186. [Google Scholar] [CrossRef]
- Liaudanskas, M.; Zymone, K.; Viskelis, J.; Kviklys, D.; Viskelis, P.; Janulis, V. Seasonal variation of the qualitative and quantitative composition of phenolic compounds in Malus domestica leaves. Chem. Nat. Comp. 2018, 54, 348. [Google Scholar] [CrossRef]
- Faramarzi, S.; Pacifico, S.; Yadollahi, A.; Lettieri, A.; Nocera, P.; Piccolella, S. Red-fleshed apples: Old autochthonous fruits as a novel source of anthocyanin antioxidants. Plant Foods Hum. Nutr. 2015, 70, 324–330. [Google Scholar] [CrossRef] [PubMed]
- Han, M.; Li, G.; Liu, X.; Li, A.; Mao, P.; Liu, P.; Li, H. Phenolic profile, antioxidant activity and anti-proliferative activity of crabapple fruits. Hort. Plant J. 2019, 5, 155–163. [Google Scholar] [CrossRef]
- Chen, F.; Li, F.; Lu, L.; Zhang, X.; Xu, X.; Li, D. Phenolic profile and changes in the antioxidant activity of crabapple (Malus domestica cv Royalty) fruit during maturation on the tree. Int. J. Food Sci. Technol. 2014, 49, 1680–1688. [Google Scholar] [CrossRef]
- Zheng, H.Z.; Kim, Y.; Chung, S.K. A profile of physicochemical and antioxidant changes during fruit growth for the utilisation of unripe apples. Food Chem. 2012, 131, 106. [Google Scholar] [CrossRef]
- Sethi, S.; Joshi, A.; Arora, B.; Bhowmik, A.; Sharma, R.R.; Kumar, P. Significance of FRAP, DPPH, and CUPRAC assays for antioxidant activity determination in apple fruit extracts. Eur. Food Res. Technol. 2020, 246, 591–598. [Google Scholar] [CrossRef]
- Ma, Y.; Huang, H. Characterisation and comparison of phenols, flavonoids and isoflavones of soymilk and their correlations with antioxidant activity. Int. J. Food Sci. Technol. 2014, 49, 2290–2298. [Google Scholar] [CrossRef]
- Wang, X.H.; Zhou, S.Y.; Qian, Z.Z.; Zhang, H.L.; Qiu, L.H.; Song, Z.; Zhao, J.; Wang, P.; Hao, X.S.; Wang, H.Q. Evaluation of toxicity and single-dose pharmacokinetics of intravenous ursolic acid liposomes in healthy adult volunteers and patients with advanced solid tumors. Expert Opin. Drug Metab. Toxicol. 2013, 9, 117–125. [Google Scholar] [CrossRef]
- Moon, S.H.; Kim, K.T.; Lee, N.K.; Han, Y.S.; Nah, S.Y.; Cho, S.G.; Park, Y.S.; Paik, H.D. Inhibitory effects of naringenin and its novel derivatives on hyaluronidase. Food Sci. Biotechnol. 2009, 18, 267–270. [Google Scholar]
- Abdullah, N.H.; Thomas, N.F.; Sivasothy, Y.; Lee, V.S.; Liew, S.Y.; Noorbatcha, I.A.; Awang, K. Hyaluronidase inhibitory activity of pentacylic triterpenoids from Prismatomeris tetrandra (Roxb.) K. Schum: Isolation, synthesis and qsar study. Int. J. Mol. Sci. 2016, 17, 143. [Google Scholar] [CrossRef]
- Omar, R.; Galala, A.; Badria, F. Multi-target inhibitory activity of some medicinal plants on α-amylase, tyrosinase and hyaluronidase: Potential therapy for treatment diabetes and diabetic complications. ASNH 2019, 3, 68–75. [Google Scholar]
- Lee, E.H.; Cho, F.B.; Kim, B.O.; Jung, H.Y.; Lee, S.Y.; Yoo, J.; Kang, I.K.; Cho, Y.J. Functional properties of newly-bred ‘Summer king’ apples. J. Hortic. Sci. Technol. 2020, 38, 405–417. [Google Scholar]
- Piwowarskia, J.P.; Kissa, A.K.; Wojciechowskab, M.K. Anti-hyaluronidase and anti-elastase activity screening of tannin-rich plant materials used in traditional Polish medicine for external treatment of diseases with inflammatory background. J. Ethnopharmacol. 2011, 137, 937–941. [Google Scholar] [CrossRef]
- Félix-Silva, J.; Gomes, J.A.S.; Xavier-Santos, J.B.; Passos, J.G.R.; Silva-Junior, A.A.; Tambourgi, D.V.; Fernandes-Pedrosa, M.F. Inhibition of local effects induced by Bothrops erythromelas snake venom: Assessment of the effectiveness of Brazilian polyvalent bothropic antivenom and aqueous leaf extract of Jatropha gossypiifolia. Toxicon 2017, 125, 74–83. [Google Scholar] [CrossRef]
- Shukla, S.; Gupta, S. Apigenin: A promising molecule for cancer prevention. Pharm. Res. 2010, 27, 962–978. [Google Scholar] [CrossRef]
- Stump, T.; Santee, B.; Williams, L.; Heinze, C.; Kunze, R. The effects of apigenin on cell proliferation and apoptosis in glioblastoma multiforme. J. Pharm. Pharmacol. 2017, 69, 907–916. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Juan, M.E.; Planas, J.M.; Ruiz-Gutierrez, V.; Daniel, H.; Wenzel, U. Antiproliferative and apoptosis-inducing effects of maslinic and oleanolic acids, two pentacyclic triterpenes from olives, on HT-29 colon cancer cells. Br. J. Nutr. 2008, 100, 36–43. [Google Scholar] [CrossRef] [Green Version]
- Bache, M.; Hein, A.; Petrenko, M.; Güttler, A.; Keßler, J.; Wichmann, H.; Kappler, M.; Emmerich, D.; Paschke, R.; Vordermark, D. Evaluation of the betulinic acid–cisplatin conjugate APC and its precursor DE9B for the treatment of human malignant glioma. Chem. Biol. Interact. 2019, 1, 108841. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Li, Y.; Wang, X.; Jiang, C. Ursolic acid inhibits proliferation and induces apoptosis in human glioblastoma cell lines U251 by suppressing TGF-b1/miR-21/PDCD4 pathway. Basic Clin. Pharmacol. Toxicol. 2012, 111, 106–112. [Google Scholar] [PubMed]
- Shen, S.; Zhang, Y.; Zhang, R.; Tu, X.; Gong, X. Ursolic acid induces autophagy in U87MG cells via ROS-dependent endoplasmic reticulum stress. Chem. Biol. Interact. 2014, 25, 28–41. [Google Scholar] [CrossRef]
- Tan, J.; Shen, Z.X.; Gen, W. Ursolic acid induces apoptosis in colon cancer HT-29 cells. Chin. J. Oncog. 2006, 28, 99–102. [Google Scholar]
- Andersson, D.; Liu, J.J.; Nilsson, A.; Duan, R.D. Ursolic acid inhibits proliferation and stimulates apoptosis in HT29 cells following activation of alkaline sphingomyelinase. Anticancer Res. 2003, 23, 3317–3322. [Google Scholar]
- Shan, J.Z.; Xuan, Y.Y.; Zheng, S.; Dong, Q.; Zhang, S.Z. Ursolic acid inhibits proliferation and induces apoptosis of HT-29 colon cancer cells by inhibiting the EGFR/MAPK pathway. J. Zhejiang Univ. Sci. B 2009, 10, 668–674. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Apak, R.; Güçlü, K.; Demirata, B.; Özyürek, M.; Çelik, S.E.; Bektaşoğlu, B.; Berker, K.I.; Özyurt, D. Comparative evaluation of various total antioxidant capacity assays applied to phenolic compounds with the CUPRAC assay. Molecules 2007, 12, 1496–1547. [Google Scholar] [CrossRef] [Green Version]
- Lee, F.H.; Park, H.J.; Kim, B.O.; Choi, H.W.; Park, K.I.; Kang, I.K.; Cho, Y.J. Anti-inflammatory effect of Malus domestica cv. Green ball apple peel extract on raw 264.7 macrophages. Appl. Biol. Chem. 2020, 63, 117–123. [Google Scholar] [CrossRef]
- Checker, R.; Sandur, S.K.; Sharma, D.; Patwardhan, R.S.; Jayakumar, S.; Kohli, V.; Sethi, G.; Aggarwal, B.B.; Sainis, K.B. Potent anti-inflammatory activity of ursolic acid, a triterpenoid antioxidant, is mediated through suppression of NF-kB, AP-1 and NF-AT. PLoS ONE 2012, 7, e31318. [Google Scholar]
- Whatcott, C.; Han, H.; Posner, R.G.; Hostetter, G.; Von Hoff, D.D. Targeting the tumor microenvironment in cancer: Why hyaluronidase deserves a second look. Cancer Discov. 2011, 1, 291–296. [Google Scholar] [CrossRef] [Green Version]
- Liyanaarachchia, G.D.; Samarasekeraa, J.K.R.R.; Mahanama, K.R.R.; Hemalal, K.D.P. Tyrosinase, elastase, hyaluronidase, inhibitory and antioxidant activity of Sri Lankan medicinal plants for novel cosmeceuticals. Ind. Crop. Prod. 2018, 111, 597–605. [Google Scholar] [CrossRef]
- Michel, P.; Owczarek, A.; Matczak, M.; Kosno, M.; Szymanski, P.; Mikiciuk-Olasik, E.; Kilanowicz, A.; Wesołowski, W.; Olszewska, M.A. Metabolite profiling of eastern teaberry (Gaultheria procumbens L.) lipophilic leaf extracts with hyaluronidase and lipoxygenase inhibitory activity. Molecules 2017, 22, 412. [Google Scholar] [CrossRef] [Green Version]
- Cargnin, S.T.; Gnoatto, S.B. Ursolic acid from apple pomace and traditional plants: A valuable triterpenoid with functional properties. Food Chem. 2017, 220, 477–489. [Google Scholar] [CrossRef]
- Han, N.; Bakovic, M. Biologically active triterpenoids and their cardioprotective and anti-inflammatory effects. J. Bioanal. Biomed. 2015, 12, 005. [Google Scholar]
- Yadav, V.R.; Prasad, S.; Sung, B.; Kannappan, R.; Aggarwal, B.B. Targeting inflammatory pathways by triterpenoids for prevention and treatment of cancer. Molecules 2010, 2, 2428–2466. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wozniak, L.; Skapska, S.; Marszałek, K. Ursolic acid—A pentacyclic terpenoid with a wide spectrum of pharmacological activities. Molecules 2015, 20, 20614–20641. [Google Scholar] [CrossRef] [Green Version]
- Lipinski, C.A. Drug-like properties and the causes of poor solubility and poor permeability. J. Pharmacol. Toxicol. Methods 2000, 44, 235–249. [Google Scholar] [CrossRef]
- Gareth, T. Medicinal Chemistry, an Introduction, 2nd ed.; John Wiley & Sons: Chichester, UK, 2007; p. 621. [Google Scholar]
- Furtado, N.A.J.C.; Pirson, L.; Edelberg, H.; Miranda, L.M.; Loira-Pastoriza, C.; Preat, V.; Larondelle, Y.; André, C.M. Pentacyclic triterpene bioavailability: An overview of in vitro and in vivo studies. Molecules 2017, 22, 400. [Google Scholar] [CrossRef] [PubMed] [Green Version]












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| No. | Cultivars | Dry Extracts | Abbreviation |
|---|---|---|---|
| 1. | ‘Kostele’ | Whole apple | AAW |
| 2. | ‘Ligol’ | Apple peel | AP1 |
| 3. | ‘Rubin’ | Apple peel | AP2 |
| 4. | ‘Auksis’ | Apple peel | AP3 |
| 5. | ‘Paprastasis antaninis’ | Whole apple | AEW |
| DPPH | ABTS | CUPRAC | FRAP | HT-29 | U-87 | ||
|---|---|---|---|---|---|---|---|
| Anti-HYAL | Pearson Correlation | 0.397 | 0.422 | 0.420 | 0.449 | 0.331 | 0.669 |
| Sig. (2-tailed) | 0.508 | 0.479 | 0.482 | 0.448 | 0.587 | 0.217 | |
| HT-29 | Pearson Correlation | 0.728 | 0.729 | 0.427 | 0.796 | 1 | 0.904 * |
| Sig. (2-tailed) | 0.163 | 0.162 | 0.473 | 0.107 | 0.035 | ||
| U-87 | Pearson Correlation | 0.835 | 0.846 | 0.658 | 0.881 * | 0.904 * | 1 |
| Sig. (2-tailed) | 0.079 | 0.071 | 0.227 | 0.049 | 0.035 | ||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Butkeviciute, A.; Petrikaite, V.; Jurgaityte, V.; Liaudanskas, M.; Janulis, V. Antioxidant, Anti-Inflammatory, and Cytotoxic Activity of Extracts from Some Commercial Apple Cultivars in Two Colorectal and Glioblastoma Human Cell Lines. Antioxidants 2021, 10, 1098. https://doi.org/10.3390/antiox10071098
Butkeviciute A, Petrikaite V, Jurgaityte V, Liaudanskas M, Janulis V. Antioxidant, Anti-Inflammatory, and Cytotoxic Activity of Extracts from Some Commercial Apple Cultivars in Two Colorectal and Glioblastoma Human Cell Lines. Antioxidants. 2021; 10(7):1098. https://doi.org/10.3390/antiox10071098
Chicago/Turabian StyleButkeviciute, Aurita, Vilma Petrikaite, Vidmante Jurgaityte, Mindaugas Liaudanskas, and Valdimaras Janulis. 2021. "Antioxidant, Anti-Inflammatory, and Cytotoxic Activity of Extracts from Some Commercial Apple Cultivars in Two Colorectal and Glioblastoma Human Cell Lines" Antioxidants 10, no. 7: 1098. https://doi.org/10.3390/antiox10071098
APA StyleButkeviciute, A., Petrikaite, V., Jurgaityte, V., Liaudanskas, M., & Janulis, V. (2021). Antioxidant, Anti-Inflammatory, and Cytotoxic Activity of Extracts from Some Commercial Apple Cultivars in Two Colorectal and Glioblastoma Human Cell Lines. Antioxidants, 10(7), 1098. https://doi.org/10.3390/antiox10071098