

Bioactive Compounds, Nutritional Quality and Antioxidant Capacity of the Red-Fleshed Kirkwood Navel and Ruby Valencia Oranges

 , and
, and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Material
2.2. Fruit Weight, Size, Color Index and Internal Maturity Determination
2.3. Sugar and Organic Acid Determination
2.4. L-Ascorbic-Acid Determination
2.5. Tocopherol Determination
2.6. Carotenoid Determination
2.7. Analysis of Total Phenolics and Flavonoids
2.8. Flavonoid Determination
2.9. Determination of the Antioxidant Capacity
2.10. Analysis of Singlet Oxygen Absorption Capacity (SOAC)
2.11. Statistical and Principal Component Analysis (PCA)
3. Results and Discussion
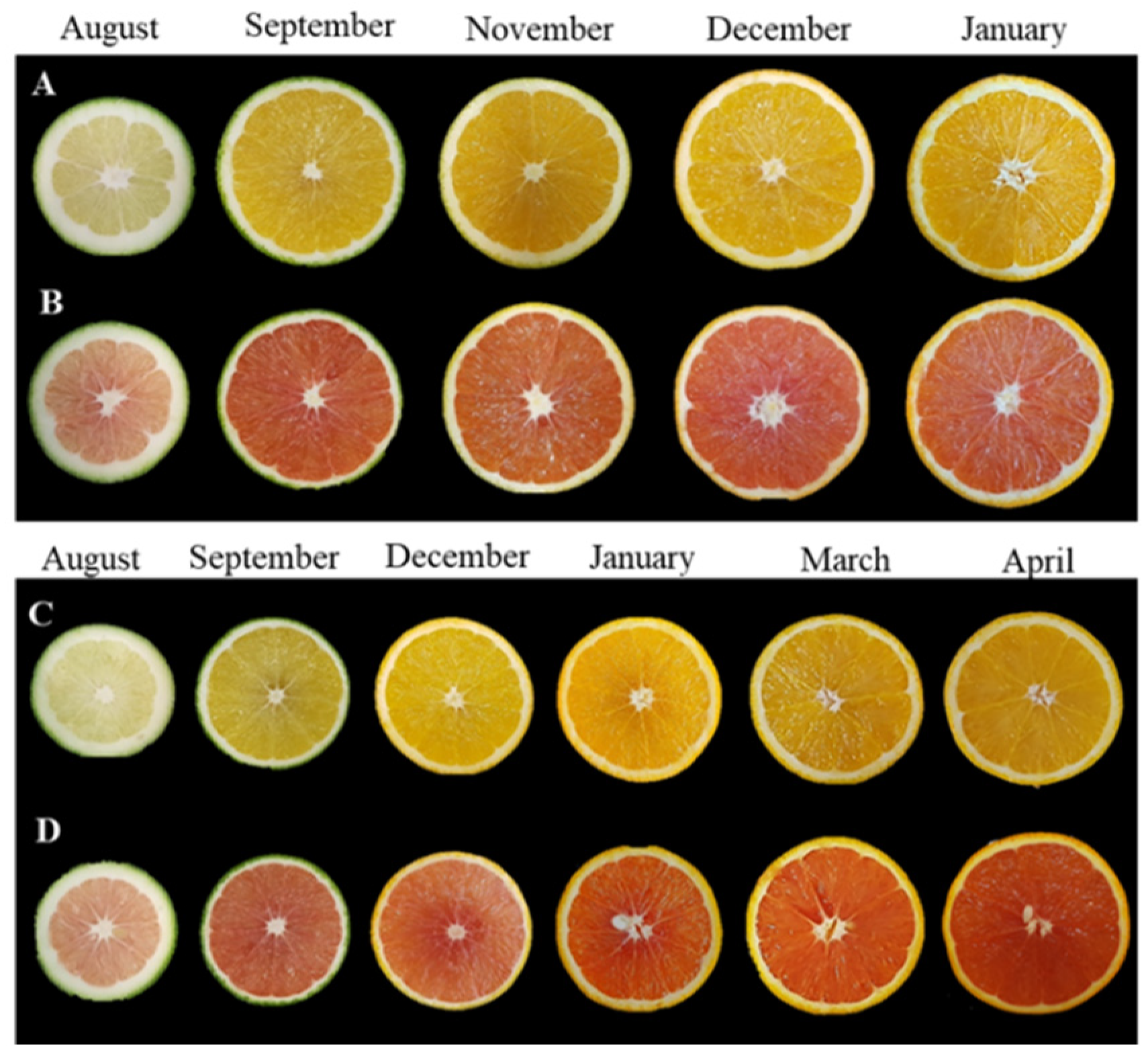
3.1. Physiological Parameters, Color and Internal Quality in the Red-Fleshed Kirkwood Navel and Ruby Valencia, and the Ordinary Navel and Valencia Oranges
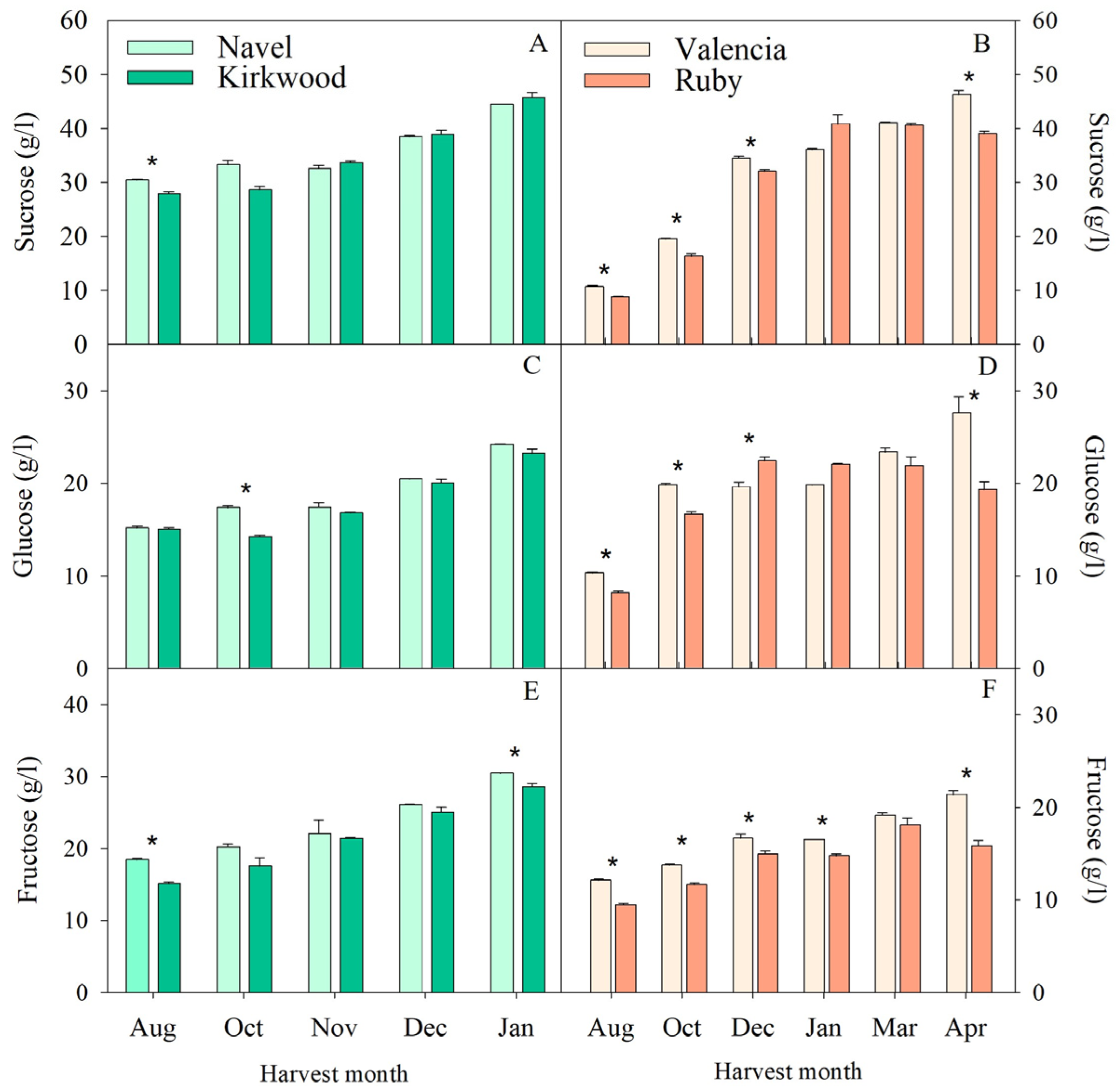
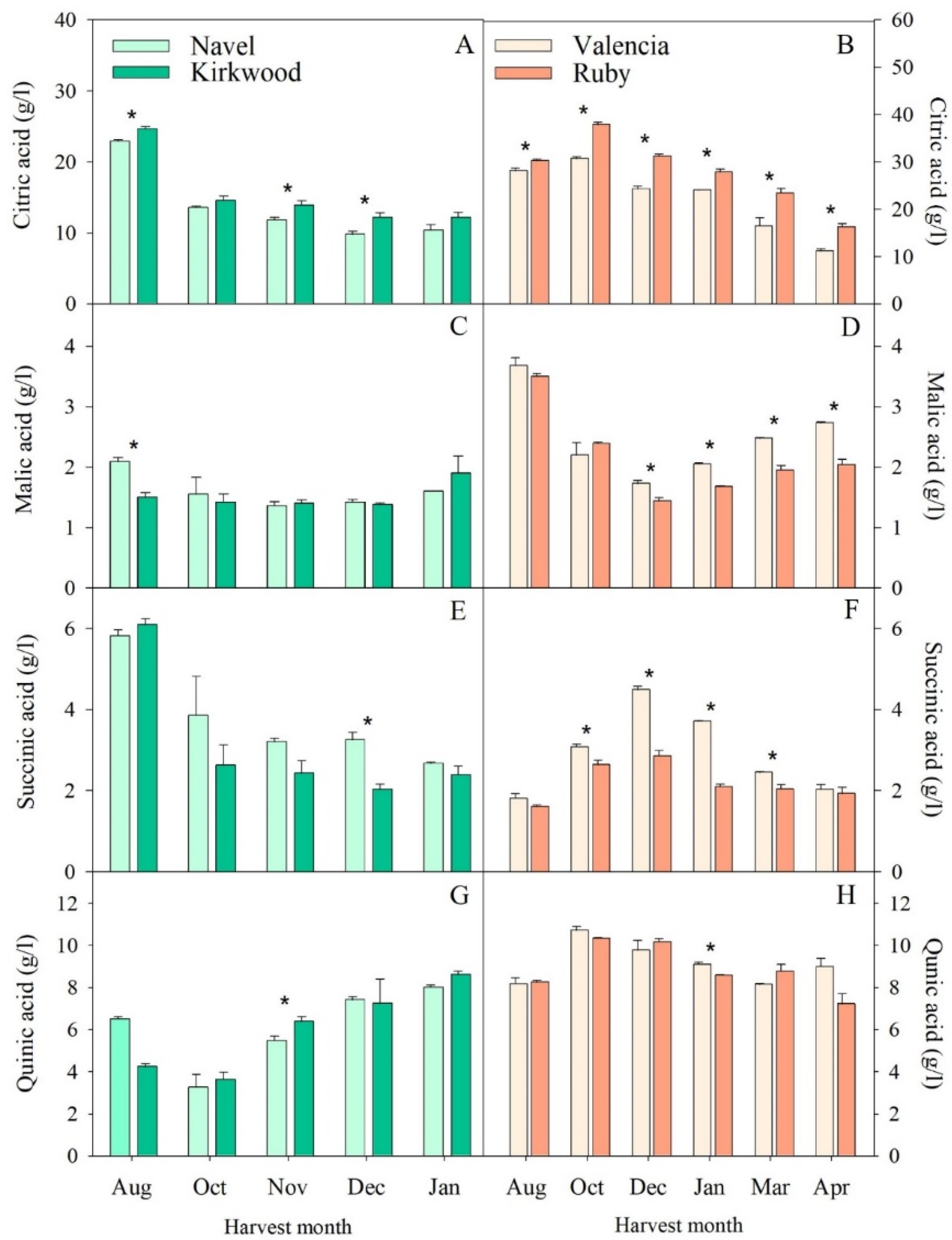
3.2. Changes in Sugars and Organic Acids in the Red-Fleshed Kirkwood Navel and Ruby Valencia and in the Ordinary Navel and Valencia Oranges
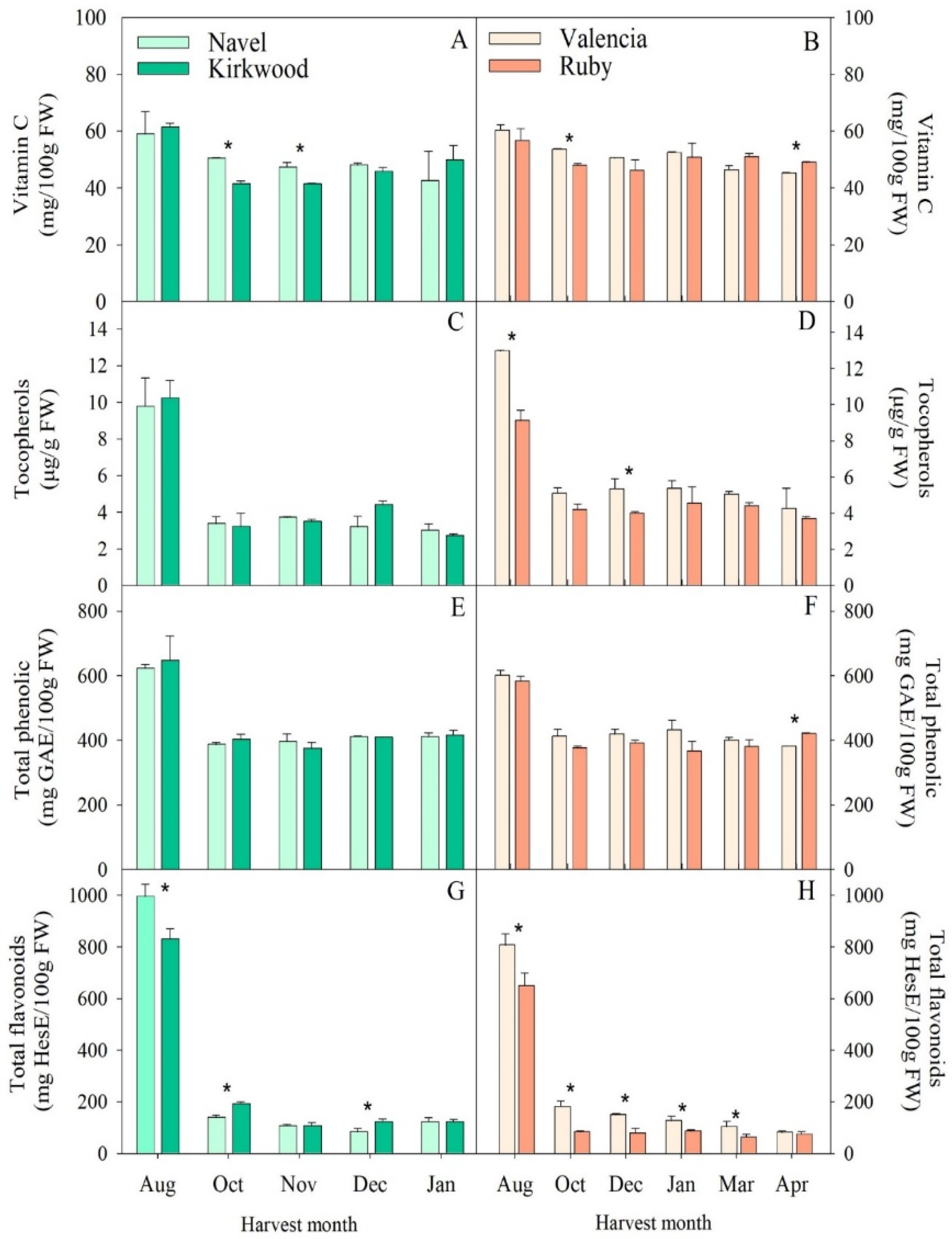
3.3. Accumulation of Ascorbic Acid, Tocopherols, Phenolic and Flavonoids Compounds in the Red-Fleshed Kirkwood Navel and Ruby Valencia and the Ordinary Navel and Valencia Oranges
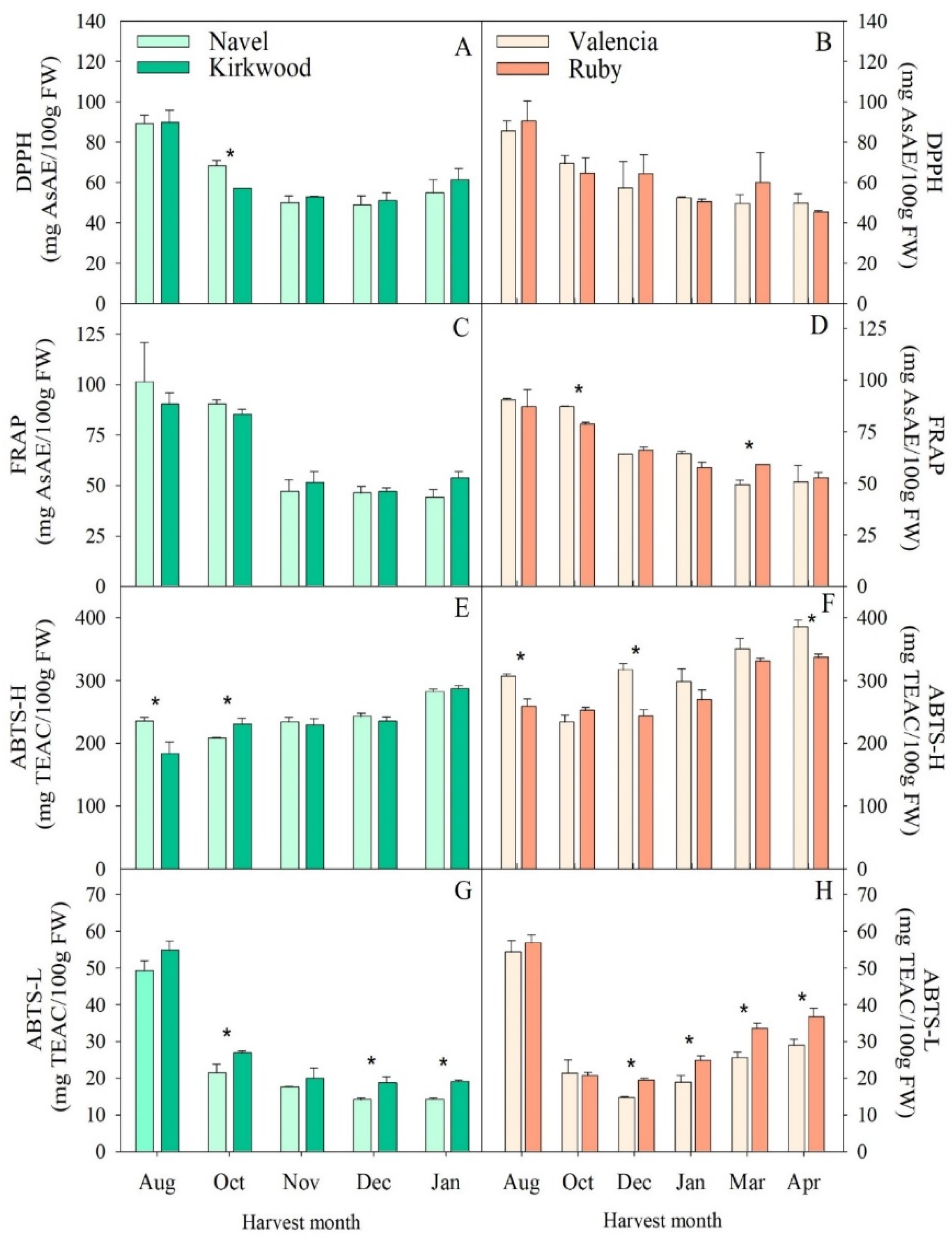
3.4. Hydrophilic and Lipophilic Antioxidant Capacity in the Red-Fleshed Kirkwood Navel and Ruby Valencia and the Ordinary Navel and Valencia Oranges
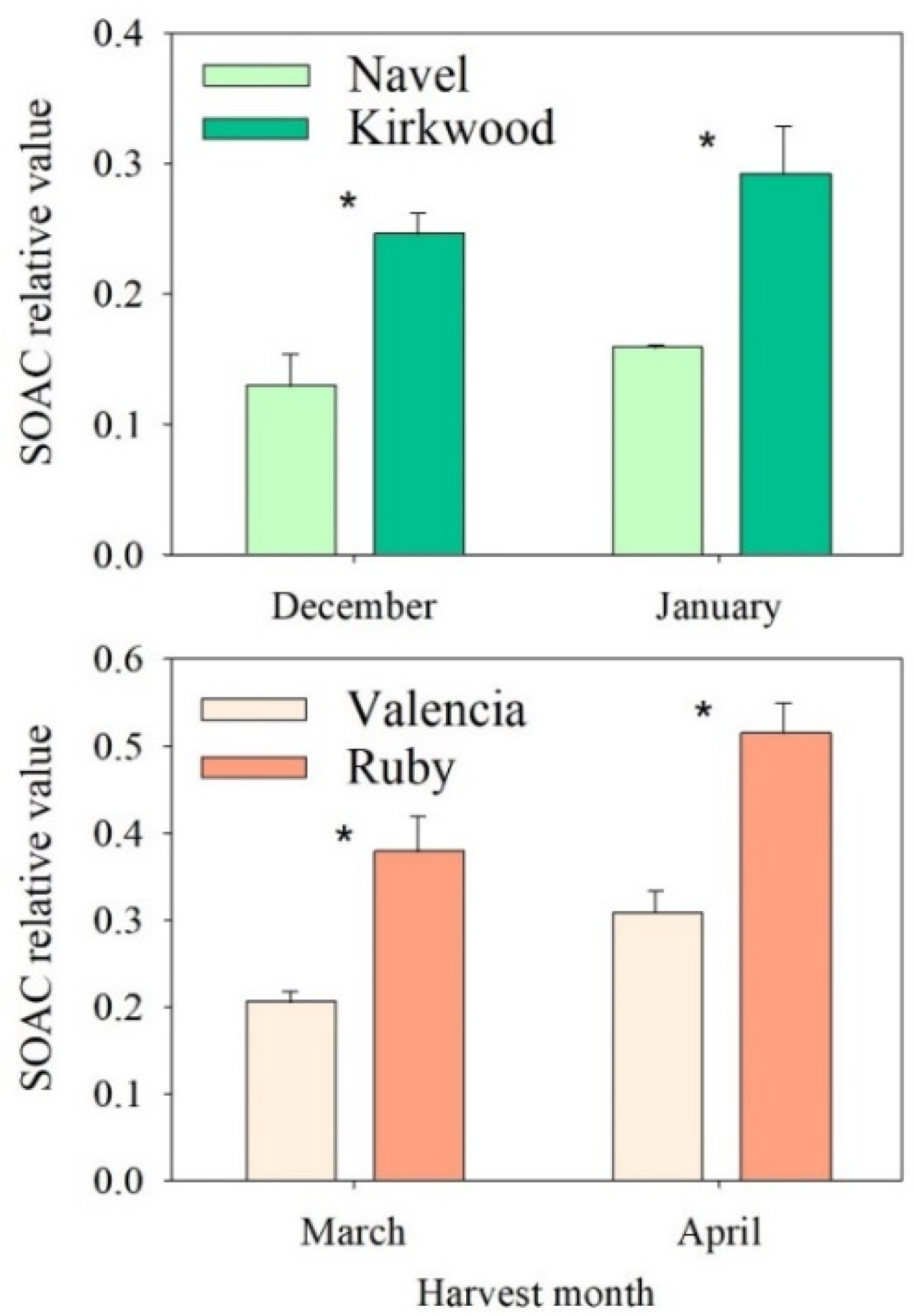
3.5. Singlet Oxygen Absorption Capacity (SOAC) and Its Relationship with Carotenoid Concentrations in the Red-Fleshed Kirkwood Navel and Ruby Valencia and the Ordinary Navel and Valencia Oranges
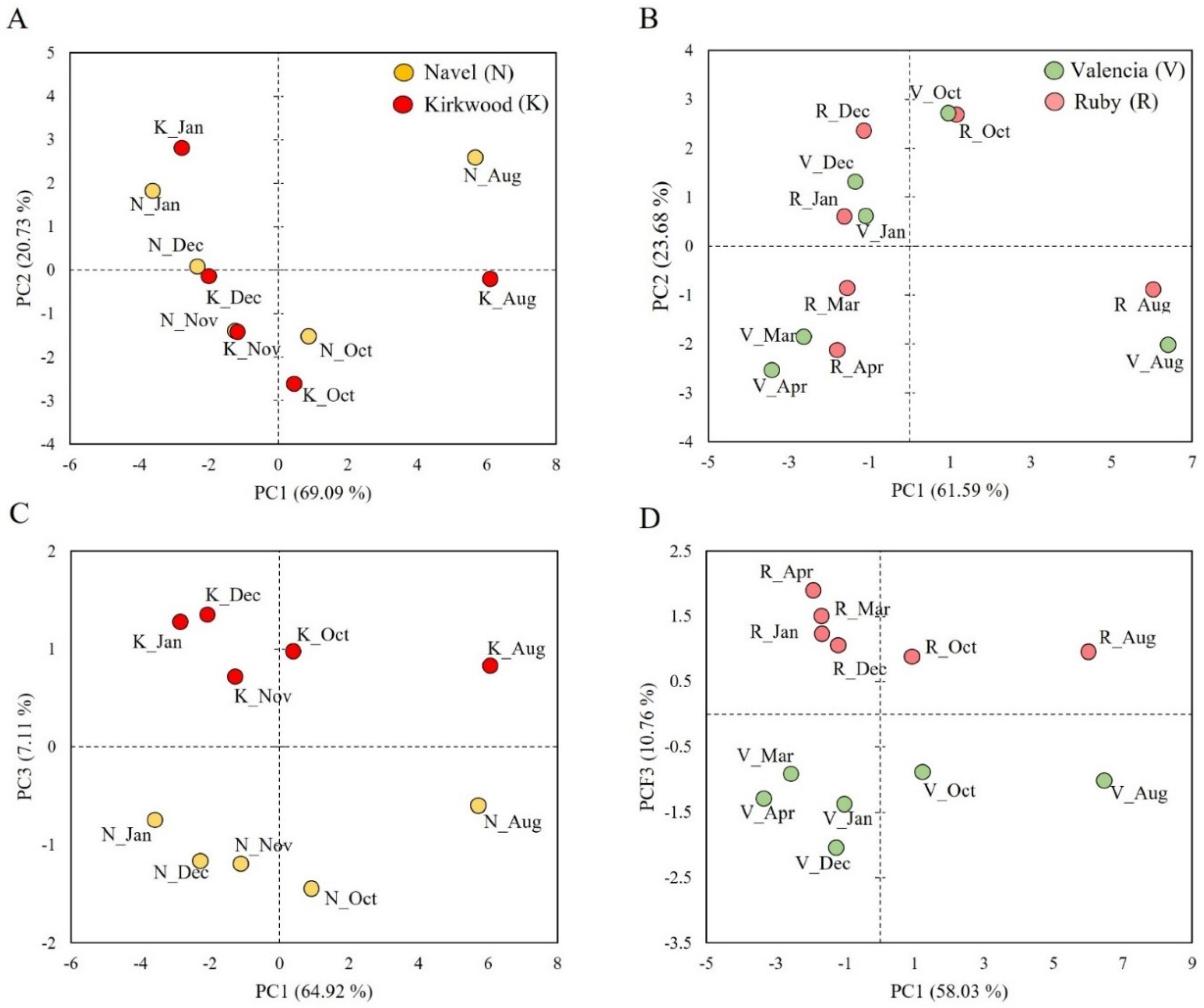
3.6. Multivariate Analysis (PCA) of Bioactive Compounds, Sugars, Organic Acids and Antioxidant Capacity in the Red-Fleshed Kirkwood Navel and Ruby Valencia and the Ordinary Navel and Valencia Oranges during Fruit Development and Maturation
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ma, G.; Zhang, L.; Sugiura, M.; Kato, M. Citrus and health. In The Genus Citrus; Talón, M., Caruso, M., Gmitter, F., Eds.; Woodhead Publishing: Cambridge, UK, 2020; pp. 495–511. [Google Scholar]
- Lv, X.; Zhao, S.; Ning, Z.; Zeng, H.; Shu, Y.; Tao, O.; Xiao, C.; Lu, C.; Liu, Y. Citrus fruits as a treasure trove of active natural metabolites that potentially provide benefits for human health. Chem. Cent. J. 2015, 9, 68. [Google Scholar] [CrossRef] [PubMed]
- Rampersaud, G.C.; Valim, M.F. 100% citrus juice: Nutritional contribution, dietary benefits, and association with anthropometric measures. Crit. Rev. Food. Sci. Nutr. 2017, 57, 129–140. [Google Scholar] [CrossRef] [PubMed]
- Saini, R.K.; Ranjit, A.; Sharma, K.; Prasad, P.; Shang, X.; Gowda, K.G.M.; Keum, Y.S. Bioactive compounds of Citrus fruits: A review of composition and health benefits of carotenoids, flavonoids, limonoids, and terpenes. Antioxidants 2022, 11, 239. [Google Scholar] [CrossRef] [PubMed]
- Rajendran, P.; Nandakumar, N.; Rengarajan, T.; Palaniswami, R.; Gnanadhas, E.N.; Lakshminarasaiah, U.; Gopas, J.; Nishigaki, I. Antioxidants and human diseases. Clin. Chim. Acta 2014, 436, 332–334. [Google Scholar] [CrossRef]
- Zou, Z.; Xi, W.; Hu, Y.; Nie, C.; Zhiquin, Z. Antioxidant activity of Citrus fruits. Food Chem. 2016, 196, 885–896. [Google Scholar] [CrossRef]
- Lu, X.; Zhao, C.; Shi, H.; Liao, Y.; Xu, F.; Du, H.; Xiao, H.; Zheng, J. Nutrients and bioactives in citrus fruits: Different citrus varieties, fruit parts, and growth stages. Crit. Rev. Food Sci. Nutr. 2021, 5, 1–24. [Google Scholar] [CrossRef]
- Roussos, P.A. Oranges (Citrus sinensis L. Osbeck). In Nutritional Composition of Fruit Cultivars; Simmods, M.S.J., Preedy, V.R., Eds.; Academic Press: Cambridge, MA, USA, 2016; pp. 469–496. [Google Scholar]
- Tadeo, F.R.; Terol, J.; Rodrigo, M.J.; Licciardelo, C.; Sadka, A. Fruit growth and development. In The Genus Citrus; Talón, M., Caruso, M., Gmitter, F., Eds.; Woodhead Publishing: Cambridge, UK, 2020; pp. 495–508. [Google Scholar]
- Rodrigo, M.J.; Alquézar, B.; Alòs, E.; Lado, J.; Zacarías, L. Biochemical bases and molecular regulation of pigmentation in the peel of Citrus fruit. Sci. Hortic. 2013, 163, 46–62. [Google Scholar] [CrossRef]
- Multari, S.; Licciardello, C.; Caruso, M.; Martens, S. Monitoring the changes in phenolic compounds and carotenoids occurring during fruit development in the tissues of four citrus fruits. Food Res. Int. 2020, 134, 109228. [Google Scholar] [CrossRef]
- Zhao, C.; Wang, F.; Lian, Y.; Xiao, H.; Zheng, J. Biosynthesis of citrus flavonoids and their health effects. Crit. Rev. Food Sci. Nutr. 2020, 60, 566–583. [Google Scholar] [CrossRef]
- Martí, N.; Mena, P.; Cánovas, J.A.; Micol, V.; Saura, D. Vitamin C and the role of citrus juices as functional food. Nat. Prod. Commun. 2009, 4, 677–700. [Google Scholar] [CrossRef] [Green Version]
- Zacarías-García, J.; Rey, F.; Gil, J.V.; Rodrigo, M.J.; Zacarías, L. Antioxidant capacity in fruit of Citrus cultivars with marked differences in pulp coloration: Contribution of carotenoids and vitamin C. Food Sci. Technol. Int. 2021, 27, 210–222. [Google Scholar] [CrossRef] [PubMed]
- Fenech, M.; Amaya, I.; Valpuesta, V.; Botella, M.A. Vitamin C Content in Fruits: Biosynthesis and Regulation. Front. Plant Sci. 2019, 9, 2006. [Google Scholar] [CrossRef] [PubMed]
- Rey, F.; Rodrigo, M.J.; Zacarias, L. Accumulation of tocopherols and transcriptional regulation of their biosynthesis during cold storage of mandarin fruit. Postharvest Biol. Technol. 2021, 180, 111594. [Google Scholar] [CrossRef]
- Traber, M.G.; Sies, H. Vitamin E in humans: Demand and delivery. Annu. Rev. Nutr. 1996, 16, 321–347. [Google Scholar] [CrossRef]
- van Breda, S.G.; de Kok, T.M. Smart combinations of bioactive compounds in fruits and vegetables may guide new strategies for personalized prevention of chronic diseases. Mol. Nutr. Food Res. 2018, 62, 1700597. [Google Scholar] [CrossRef]
- Natarajan, T.D.; Ramasamy, J.R.; Palanisamy, K. Nutraceutical potentials of synergic foods: A systematic review. J. Ethn. Food 2019, 6, 27. [Google Scholar] [CrossRef]
- Zhanga, L.; Virgous, C.; Si, H. Synergistic anti-inflammatory effects and mechanisms of combined phytochemicals. J. Nutr. Biochem. 2019, 69, 19–30. [Google Scholar] [CrossRef]
- Niki, E.; Noguchi, N.; Tsuchihashi, H.; Gotoh, N. Interaction among vitamin C, vitamin E, and β-carotene. Am. J. Clin. Nutr. 1995, 62, 1322S–1326S. [Google Scholar] [CrossRef]
- Stahl, W.; Junghans, A.; de Boer, B.; Driomina, E.S.; Briviba, K.; Sies, H. Carotenoid mixtures protect multilamellar liposomes against oxidative damage: Synergistic effects of lycopene and lutein. FEBS Lett. 1998, 427, 305–308. [Google Scholar] [CrossRef]
- Milde, J.; Elstner, E.F.; Grassmann, J. Synergistic effects of phenolics and carotenoids on human low-density lipoprotein oxidation. Mol. Nutr. Food Res. 2007, 51, 956–961. [Google Scholar] [CrossRef]
- Liu, K.; Juan, X.; Liu, Y.; Zhao, X. A novel bud mutation that confers abnormal patterns of lycopene accumulation in sweet orange fruit (Citrus sinensis L Osbeck). J. Exp. Bot. 2007, 58, 4161–4171. [Google Scholar] [CrossRef] [PubMed]
- Alquézar, B.; Rodrigo, M.J.; Zacarías, L. Regulation of carotenoid biosynthesis during fruit maturation in the red-fleshed orange mutant Cara Cara. Phytochemistry 2008, 69, 1997–2007. [Google Scholar] [CrossRef] [PubMed]
- Rodrigo, M.J.; Cilla, A.; Barberá, R.; Zacarías, L. Carotenoid bioaccessibility in pulp and fresh juice from carotenoid-rich sweet oranges and mandarins. Food Funct. 2015, 6, 1950–1959. [Google Scholar] [CrossRef]
- Lu, P.J.; Wang, C.Y.; Yin, T.T.; Zhong, S.L.; Grierson, D.; Chen, K.S.; Xu, C.J. Cytological and molecular characterization of carotenoid accumulation in normal and high-lycopene mutant oranges. Sci. Rep. 2017, 7, 761. [Google Scholar] [CrossRef]
- Chen, J.; Zhang, H.; Pang, Y.; Cheng, Y.; Deng, X.; Xu, J. Comparative study of flavonoid production in lycopene-accumulated and blonde-flesh sweet oranges (Citrus sinensis) during fruit development. Food Chem. 2015, 184, 238–246. [Google Scholar] [CrossRef]
- Brasili, E.; Chaves, D.F.; Xavier, A.A.; Mercadante, A.Z.; Hassimotto, N.M.; Lajolo, F.M. Effect of Pasteurization on Flavonoids and Carotenoids in Citrus sinensis (L.) Osbeck cv. ‘Cara Cara’ and ‘Bahia’ Juices. J. Agric. Food Chem. 2017, 65, 1371–1377. [Google Scholar] [CrossRef] [PubMed]
- De Ancos, B.; Cilla, A.; Barberá, R.; Sánchez-Moreno, C.; Cano, M.P. Influence of orange cultivar and mandarin postharvest storage on polyphenols, ascorbic acid and antioxidant activity during gastrointestinal digestion. Food Chem. 2017, 225, 114–124. [Google Scholar] [CrossRef] [PubMed]
- Cilla, A.; Rodrigo, M.J.; Zacarías, L.; de Ancos, B.; Sánchez-Moreno, C.; Barberá, R.; Alegría, A. Protective effect of bioaccessible fractions of citrus fruit pulps against H2O2-induced oxidative stress in Caco-2 cells. Food Res. Int. 2018, 103, 335–344. [Google Scholar] [CrossRef]
- de Oliveira, R.; Orlando, C.; Cadavid, M.; Carmona, L.; Peña, L.; De, R.; Oliveira, P.; Scotti, L. Pasteurized Orange Juice Rich in Carotenoids Protects Caenorhabditis elegans against Oxidative Stress and β-Amyloid Toxicity through Direct and Indirect Mechanisms. Oxid. Med. Cell. Longev. 2019, 2019, 5046280. [Google Scholar]
- Pan, Z.; Liu, Q.; Yun, Z.; Guan, R.; Zeng, W.; Xu, Q.; Deng, X. Comparative proteomics of a lycopene-accumulating mutant reveals the important role of oxidative stress on carotenogenesis in sweet orange (Citrus sinensis [L.] osbeck). Proteomics 2009, 9, 5455–5470. [Google Scholar] [CrossRef]
- Xu, Q.; Yu, K.; Zhu, A.; Ye, J.; Liu, Q.; Zhang, J.; Deng, X. Comparative transcripts profiling reveals new insight into molecular processes regulating lycopene accumulation in a sweet orange (Citrus sinensis) red-flesh mutant. BMC Genet. 2019, 10, 540. [Google Scholar] [CrossRef] [PubMed]
- Kafkas, E.; Polatöz, S.; Koç, N.K. Quantification and comparison of sugars, carboxylic acids and vitamin c components of various citrus species by HPLC techniques. J. Agric. Sci. Technol. 2011, 5, 175–180. [Google Scholar]
- Yu, K.; Xu, Q.; Da, X.; Guo, F.; Ding, Y.; Deng, X. Transcriptome changes during fruit development and ripening of sweet orange (Citrus sinensis). BMC Genet. 2012, 13, 10. [Google Scholar] [CrossRef] [PubMed]
- Liu, C.; He, M.; Wang, Z.; Xu, J. Integrative analysis of terpenoid profiles and hormones from fruits of red-flesh citrus mutants and their wild types. Molecules 2019, 24, 3456. [Google Scholar] [CrossRef]
- Zacarías-García, J.; Cronje, P.; Diretto, G.; Zacarias, L.; Rodrigo, M.J. A comprehensive analysis of carotenoids metabolism in two red-fleshed mutants of Navel and Valencia oranges (Citrus sinensis). Front. Plant Sci. 2022; accepted. [Google Scholar]
- Barry, G.H.; Caruso, M.; Gmiter, F.G. Commercial Scion Varieties. In The Genus Citrus; Talón, M., Caruso, M., Gmitter, F., Eds.; Woodhead Publishing: Cambridge, UK, 2020; pp. 83–104. [Google Scholar]
- Pérez-Través, L.; Lopes, C.A.; González, R.; Barrio, E.; Querol, A. Physiological and genomic characterisation of Saccharomyces cerevisiae hybrids with improved fermentation performance and mannoprotein release capacity. Int. J. Food Microbiol. 2015, 205, 30–40. [Google Scholar] [CrossRef]
- Alòs, E.; Rodrigo, M.J.; Zacarías, L. Differential transcriptional regulation of L-ascorbic acid content in peel and pulp of citrus fruits during development and maturation. Planta 2014, 239, 1113–1128. [Google Scholar] [CrossRef]
- Singleton, V.L.; Rossi, J.A. Colorimetry of total phenolics with phosphomolybdic-phosphotungstic acid reagents. Am. J. Enol. Vitic. 1965, 16, 144–158. [Google Scholar]
- Lafuente, M.T.; Alférez, F.; Romero, P. Postharvest ethylene conditioning as a tool to reduce quality loss of stored mature sweet oranges. Postharvest Biol. Technol. 2014, 94, 104–111. [Google Scholar] [CrossRef]
- Rey, F.; Zacarías, L.; Rodrigo, M.J. Carotenoids, vitamin c, and antioxidant capacity in the peel of mandarin fruit in relation to the susceptibility to chilling injury during postharvest cold storage. Antioxidants 2020, 9, 1296. [Google Scholar] [CrossRef]
- Benzie, I.F.F.; Strain, J.J. The Ferric Reducing Ability of Plasma (FRAP) as a Measure of ‘Antioxidant Power’: The FRAP Assay. Anal. Biochem. 1996, 239, 70–76. [Google Scholar] [CrossRef] [PubMed]
- Legua, P.; Hernández, F.; Díaz-Mula, H.M.; Valero, D.; Serrano, M. Quality, bioactive compounds, and antioxidant activity of new flat-type peach and nectarine cultivars: A Comparative Study. J. Food Sci. 2011, 76, 729–736. [Google Scholar] [CrossRef] [PubMed]
- Bain, J.M. Morphological anatomical and physiological changes in the developing fruit of the Valencia orange, Citrus sinensis L. Osbeck. Aust. J. Bot. 1968, 6, 1–23. [Google Scholar] [CrossRef]
- Lado, J.; Rodrigo, M.J.; Zacarías, L. Maturity indicators and citrus fruit quality. Stewart Postharvest Rev. 2014, 10, 1–6. [Google Scholar]
- Ladaniya, M.S. Citrus Fruit: Biology, Technology and Evaluation; Elsevier Inc.: Atlanta, GA, USA, 2008. [Google Scholar]
- Bermejo, A.; Cano, A. Analysis of nutritional constituents in twenty citrus cultivars from the mediterranean area at different stages of ripening. Food Sci. Nutr. 2012, 03, 639–650. [Google Scholar] [CrossRef]
- Guo, L.-X.; Shi, C.-Y.; Liu, X.; Ning, D.-Y.; Jing, L.-F.; Yang, H.; Liu, Y.Z. Citrate accumulation-related gene expression and/or enzyme activity analysis combined with metabolomics provide a novel insight for an orange mutant. Sci. Rep. 2016, 6, 29343. [Google Scholar] [CrossRef]
- Hussain, S.B.; Shi, C.-Y.; Guo, L.-X.; Kamran, H.M.; Sadka, A.; Liu, Y.-Z. Recent advances in the regulation of citric acid metabolism in citrus fruit. Crit. Rev. Plant Sci. 2017, 36, 241–256. [Google Scholar] [CrossRef]
- Rey, F.; Rodrigo, M.J.; Diretto, G.; Zacarías, L. Effect of fruit shading and cold storage on tocopherol biosynthesis and its involvement in the susceptibility of Star Ruby grapefruit to chilling injury. Food Chem. 2021, 3, 100037. [Google Scholar] [CrossRef]
- Rey, F.; Rodrigo, M.J.; Zacarias, L. Regulation of tocopherol biosynthesis during fruit maturation of different Citrus species. Front. Plant Sci. 2021, 12, 743993. [Google Scholar] [CrossRef]
- Dong, X.; Hu, Y.; Li, Y.; Zhou, Z. The maturity degree, phenolic compounds and antioxidant activity of Eureka lemon [Citrus limon (L.) Burm. f.]: A negative correlation between total phenolic content, antioxidant capacity and soluble solid content. Sci. Hortic. 2019, 243, 281–289. [Google Scholar] [CrossRef]
- De Ancos, B.; Rodrigo, M.J.; Sánchez-Moreno, C.; Cano, M.P.; Zacarías, L. Effect of high-pressure processing applied as pretreatment on carotenoids, flavonoids and vitamin C in juice of the sweet oranges ‘Navel’ and the red-fleshed ‘Cara Cara’. Food Res Int. 2020, 132, 109105. [Google Scholar] [CrossRef] [PubMed]
- Gironés-Vilaplana, A.; Moreno, D.A.; García-Viguera, C. Phytochemistry and biological activity of Spanish Citrus fruits. Food Funct. 2014, 5, 764–772. [Google Scholar] [CrossRef] [PubMed]
- Rodríguez-Amaya, D.B. Quantitative analysis in vitro assessment of bioavailability and antioxidant activity of food carotenoids—A review. J. Food Compos. Anal. 2010, 23, 726–740. [Google Scholar] [CrossRef]
- Yoo, K.M.; Lee, K.W.; Park, J.B.; Lee, H.J.; Hwang, I.K. Variation in major antioxidants and total antioxidant activity of Yuzu (Citrus Junos Sieb ex Tanaka) during maturation and between cultivars. J. Agric. Food Chem. 2004, 52, 5907–5913. [Google Scholar] [CrossRef] [PubMed]
- Huang, R.; Xia, R.; Hu, L.; Lu, H.; Wang, M. Antioxidant activity and oxygen-scavenging system in orange pulp during fruit ripening and maturation. Sci. Hortic. 2007, 113, 166–172. [Google Scholar] [CrossRef]
- Sánchez-Moreno, C.; Plaza, L.; De Ancos, B.; Cano, P. Quantitative bioactive compounds assessment and their relative contribution to the antioxidant capacity of commercial orange juices. J. Sci. Food Agric. 2003, 83, 430–439. [Google Scholar] [CrossRef]
- Long, X.; Zeng, X.; Yan, H.; Xu, M.; Zeng, Q.; Xu, C.; Xu, Q.; Liang, T.; Zhang, J. Flavonoids composition and antioxidant potential assessment of extracts from Gannanzao Navel Orange (Citrus sinensis Osbeck Cv. Gannanzao) peel. Nat. Prod. Res. 2021, 35, 702–706. [Google Scholar] [CrossRef]
- Gulo, K.N.; Suhartomi, S.; Saragih, A.D.; Raif, M.A.; Ikhtiari, R. Antioxidant Activity of flavonoid compounds in ethanol and ethyl acetate extract from Citrus sinensis. In Proceedings of the 2021 International Conference on Artificial Intelligence and Mechatronics Systems (AIMS), Jakarta, Indonesia, 28–30 April 2021; pp. 1–6. [Google Scholar]
- Di Mascio, P.; Kaiser, S.; Sies, H. Lycopene as the most efficient biological carotenoid singlet oxygen quencher. Arch. Biochem. Biophys. 1989, 274, 532–538. [Google Scholar] [CrossRef]
- Martínez, A.; Stinco, C.M.; Meléndez-Martínez, A.J. Free radical scavenging properties of phytofluene and phytoene isomers as compared to lycopene: A combined experimental and theoretical study. J. Phys. Chem. 2014, 118, 9819–9825. [Google Scholar] [CrossRef]
- Arnao, M.B.; Cano, A.; Acosta, M. The hydrophilic and lipophilic contribution to total antioxidant activity. Food Chem. 2001, 73, 239–244. [Google Scholar] [CrossRef]
- Liu, D.; Shi, J.; Ibarra, A.C.; Kakuda, Y.; Xue, S.J. The scavenging capacity and synergistic effects of lycopene, vitamin E, vitamin C, and b-carotene mixtures on the DPPH free radical. LWT 2008, 41, 1344–1349. [Google Scholar] [CrossRef]
- Chen, X.; Deng, Z.; Zheng, L.; Zhang, B.; Luo, T.; Li, H. Interaction between flavonoids and carotenoids on ameliorating oxidative stress and cellular uptake in different cells. Foods 2021, 10, 3096. [Google Scholar] [CrossRef] [PubMed]
- Ouchi, A.; Aizawa, K.; Iwasaki, Y.; Inakuma, T.; Terao, J.; Nagaoka, S.I.; Mukai, K. Kinetic study of the quenching reaction of singlet oxygen by carotenoids and food extracts in solution. Development of a singlet oxygen absorption capacity (SOAC) assay method. J. Agric. Food Chem. 2010, 58, 9967–9978. [Google Scholar] [CrossRef] [PubMed]
- Stahl, W.; Sies, H. Antioxidant activity of carotenoids. Mol. Asp. Med. 2003, 24, 345–351. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameter | Navel | ||||
| Aug | Oct | Nov | Dec | Jan | |
| Diameter (cm) | 4.63 ± 0.14 | 7.27 ± 0.16 | 7.18 ± 0.14 | 7.39 ± 0.27 | 7.70 ± 0.19 |
| Height (cm) | 4.58 ± 0.15 | 6.81 ± 0.18 | 6.81 ± 0.15 | 7.20 ± 0.27 * | 7.55 ± 0.21 |
| Peel color (a/b) | - | −0.72 ± 0.02 | 0.49 ± 0.04 | 0.66 ± 0.04 | 0.76 ± 0.03 |
| Pulp color (a/b) | - | −0.24 ± 0.01 * | −0.12 ± 0.07 * | −0.06 ± 0.01 * | 0.00 ± 0.01 * |
| TSS (°Brix) | - | 10.65 ± 0.07 * | 11.55 ± 0.07 | 12.55 ± 0.21 | 12.75 ± 12.8 |
| TA (mg CA/100 mL) | - | 1.28 ± 0.01 * | 1.01 ± 0.04 | 0.86 ± 0.05 | 0.84 ± 0.02 |
| MI (TSS/TA) | - | 8.32 ± 0.06 * | 11.47 ± 0.52 * | 14.59 ± 0.03 | 15.11 ± 0.44 |
| Parameter | Kirkwood | ||||
| Aug | Oct | Nov | Dec | Jan | |
| Diameter (cm) | 4.76 ± 0.14 | 7.00 ± 0.24 | 6.84 ± 0.11 | 7.11 ± 0.14 | 7.54 ± 0.18 |
| Height (cm) | 4.66 ± 0.19 | 6.43 ± 0.12 | 6.49 ± 0.11 | 6.53 ± 0.18 | 7.16 ± 0.11 |
| Peel color (a/b) | - | −0.72 ± 0.02 | 0.37 ± 0.02 | 0.57 ± 0.03 | 0.77 ± 0.02 |
| Pulp color (a/b) | - | 0.66 ± 0.07 | 0.41 ± 0.05 | 0.46 ± 0.05 | 0.54 ± 0.03 |
| TSS (°Brix) | - | 10.15 ± 0.07 | 11.45 ± 0.35 | 12.55 ± 0.11 | 12.80 ± 0.01 |
| TA (mg CA/100 mL) | - | 1.46 ± 0.01 | 1.14 ± 0.02 | 0.87 ± 0.03 | 0.83 ± 0.04 |
| MI (TSS/TA) | - | 6.95 ± 0.01 | 9.98 ± 0.21 | 14.42 ± 0.10 | 15.29 ± 0.16 |
| Parameters | Valencia | |||||
| Aug | Oct | Dec | Jan | Mar | Apr | |
| Diameter (cm) | 4.28 ± 0.09 | 6.18 ± 0.09 * | 6.44 ± 0.08 * | 6.59 ± 0.17 | 6.91 ± 0.11 | 6.95 ± 0.13 |
| Height (cm) | 4.24 ± 0.10 * | 6.10 ± 0.09 | 6.25 ± 0.12 | 6.46 ± 0.11 | 6.81 ± 0.09 | 7.08 ± 0.14 |
| Peel color (a/b) | - | −0.75 ± 0.01 | 0.39 ± 0.03 * | 0.66 ± 0.03 | 0.46 ± 0.02 | 0.66 ± 0.04 |
| Pulp color (a/b) | - | −0.34 ± 0.01 * | −0.16 ± 0.01 * | −0.07 ± 0.01 * | 0.00 ± 0.01 * | 0.06 ± 0.01 * |
| TSS (°Brix) | - | 7.80 ± 0.02 | 8.66 ± 0.05 | 9.02 ± 0.03 * | 10.70 ± 0.21 | 11.50 ± 0.10 |
| TA (mg CA/100 mL) | - | 1.92 ± 0.09 | 1.44 ± 0.17 | 1.24 ± 0.03 | 1.10 ± 0.06 | 0.99 ± 0.07 |
| MI (TSS/TA) | - | 4.06 ± 0.08 * | 6.01 ± 0.21 | 7.27 ± 0.15 | 9.72 ± 0.16 | 11.60 ± 0.87 |
| Parameters | Ruby | |||||
| Aug | Oct | Dec | Jan | Mar | Apr | |
| Diameter (cm) | 4.13 ± 0.11 | 5.73 ± 0.09 | 5.83 ± 0.27 | 6.32 ± 0.08 | 6.52 ± 0.09 | 6.51 ± 0.10 |
| Height (cm) | 4.15 ± 0.11 | 5.83 ± 0.25 | 6.03 ± 0.26 | 6.11 ± 0.11 | 6.32 ± 0.10 | 6.59 ± 0.11 |
| Peel color (a/b) | - | −0.79 ± 0.01 | 0.16 ± 0.03 | 0.59 ± 0.02 | 0.49 ± 0.03 | 0.67 ± 0.02 |
| Pulp color (a/b) | - | 0.72 ± 0.06 | 0.55 ± 0.05 | 0.55 ± 0.03 | 0.65 ± 0.03 | 0.60 ± 0.03 |
| TSS (°Brix) | - | 7.85 ± 0.07 | 9.05 ± 0.11 | 9.80 ± 0.03 | 10.86 ± 0.05 | 11.95 ± 0.20 |
| TA (mg CA/100 mL) | - | 1.75 ± 0.09 | 1.59 ± 0.06 | 1.50 ± 0.08 | 1.30 ± 0.17 | 1.05 ± 0.04 |
| MI (TSS/TA) | - | 4.48 ± 0.08 | 5.69 ± 0.30 | 6.53 ± 0.34 | 8.35 ± 1.10 | 11.38 ± 0.54 |
| Flavonoid | Navel | Kirkwood | Valencia | Ruby |
| Rutin | 2.23 ± 0.17 | 2.69 ± 0.60 | 2.21 ± 0.02 * | 2.04 ± 0.02 |
| Eriocitrin | 0.98 ± 0.09 | 1.08 ± 0.13 | 0.84 ± 0.12 | 0.76 ± 0.01 |
| Narirutin | 7.58 ± 1.05 | 9.08 ± 1.90 | 3.76 ± 0.15 * | 4.45 ± 0.07 |
| Naringin | 0.28 ± 0.03 | 0.30 ±0.07 | 0.63 ± 0.05 * | 0.45 ± 0.02 |
| Hesperidin | 46.05 ± 9.02 | 60.11 ± 4.72 | 53.93 ± 5.72 | 53.48 ± 1.35 |
| Dydimin | 0.85 ± 0.13 | 1.17 ± 0.26 | 0.46 ± 0.12 | 0.72 ± 0.07 |
| Total | 57.96 ± 10.43 | 74.43 ± 7.41 | 61.84 ± 6.17 | 61.90 ± 1.52 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zacarías-García, J.; Pérez-Través, L.; Gil, J.-V.; Rodrigo, M.-J.; Zacarías, L. Bioactive Compounds, Nutritional Quality and Antioxidant Capacity of the Red-Fleshed Kirkwood Navel and Ruby Valencia Oranges. Antioxidants 2022, 11, 1905. https://doi.org/10.3390/antiox11101905
Zacarías-García J, Pérez-Través L, Gil J-V, Rodrigo M-J, Zacarías L. Bioactive Compounds, Nutritional Quality and Antioxidant Capacity of the Red-Fleshed Kirkwood Navel and Ruby Valencia Oranges. Antioxidants. 2022; 11(10):1905. https://doi.org/10.3390/antiox11101905
Chicago/Turabian StyleZacarías-García, Jaime, Laura Pérez-Través, José-Vicente Gil, María-Jesús Rodrigo, and Lorenzo Zacarías. 2022. "Bioactive Compounds, Nutritional Quality and Antioxidant Capacity of the Red-Fleshed Kirkwood Navel and Ruby Valencia Oranges" Antioxidants 11, no. 10: 1905. https://doi.org/10.3390/antiox11101905
APA StyleZacarías-García, J., Pérez-Través, L., Gil, J. -V., Rodrigo, M. -J., & Zacarías, L. (2022). Bioactive Compounds, Nutritional Quality and Antioxidant Capacity of the Red-Fleshed Kirkwood Navel and Ruby Valencia Oranges. Antioxidants, 11(10), 1905. https://doi.org/10.3390/antiox11101905