
Characterization of SOD and GPX Gene Families in the Soybeans in Response to Drought and Salinity Stresses

 , ,
, ,  , ,
, ,  ,
, 

Abstract
:1. Introduction
2. Materials and Methods
2.1. Database Searches and Retrieval of SOD and GPX Genes
2.2. Physicochemical Characteristics and Subcellular Localization
2.3. Multiple Sequence Alignment and Phylogenetic Analysis
2.4. Gene Structure and Promoter Analysis
2.5. Chromosomal Localization and Gene Duplication Analysis
2.6. Syntenic and Evolutionary Relationship
2.7. Expression Pattern Detected by Transcriptome Data
2.8. Plant Materials, Stress Treatments and Tissue Sampling
2.9. RNA Isolation and Quantitative Real-Time PCR Analysis
3. Results
3.1. Identification and Distribution of SOD and GPX Genes
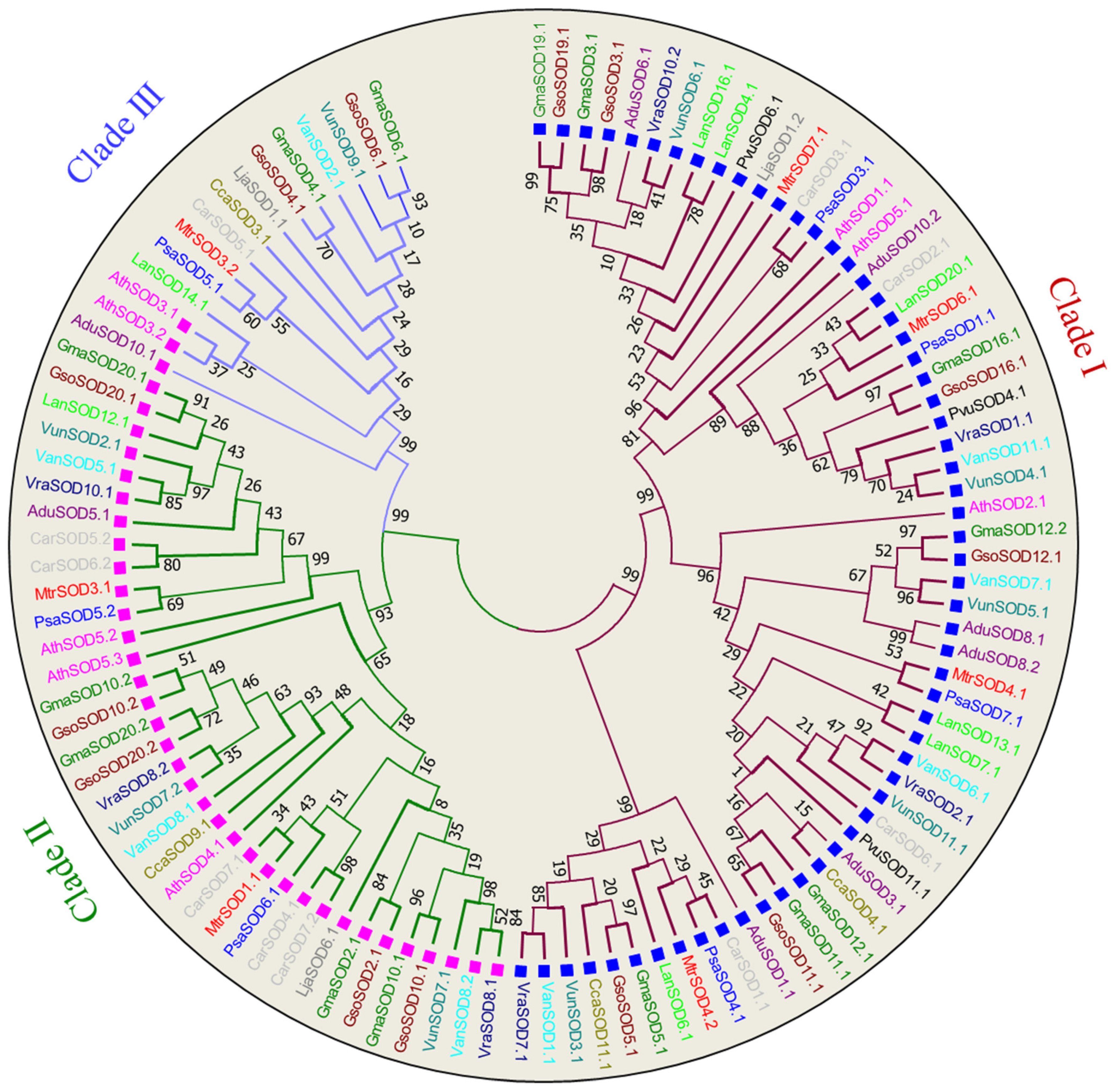
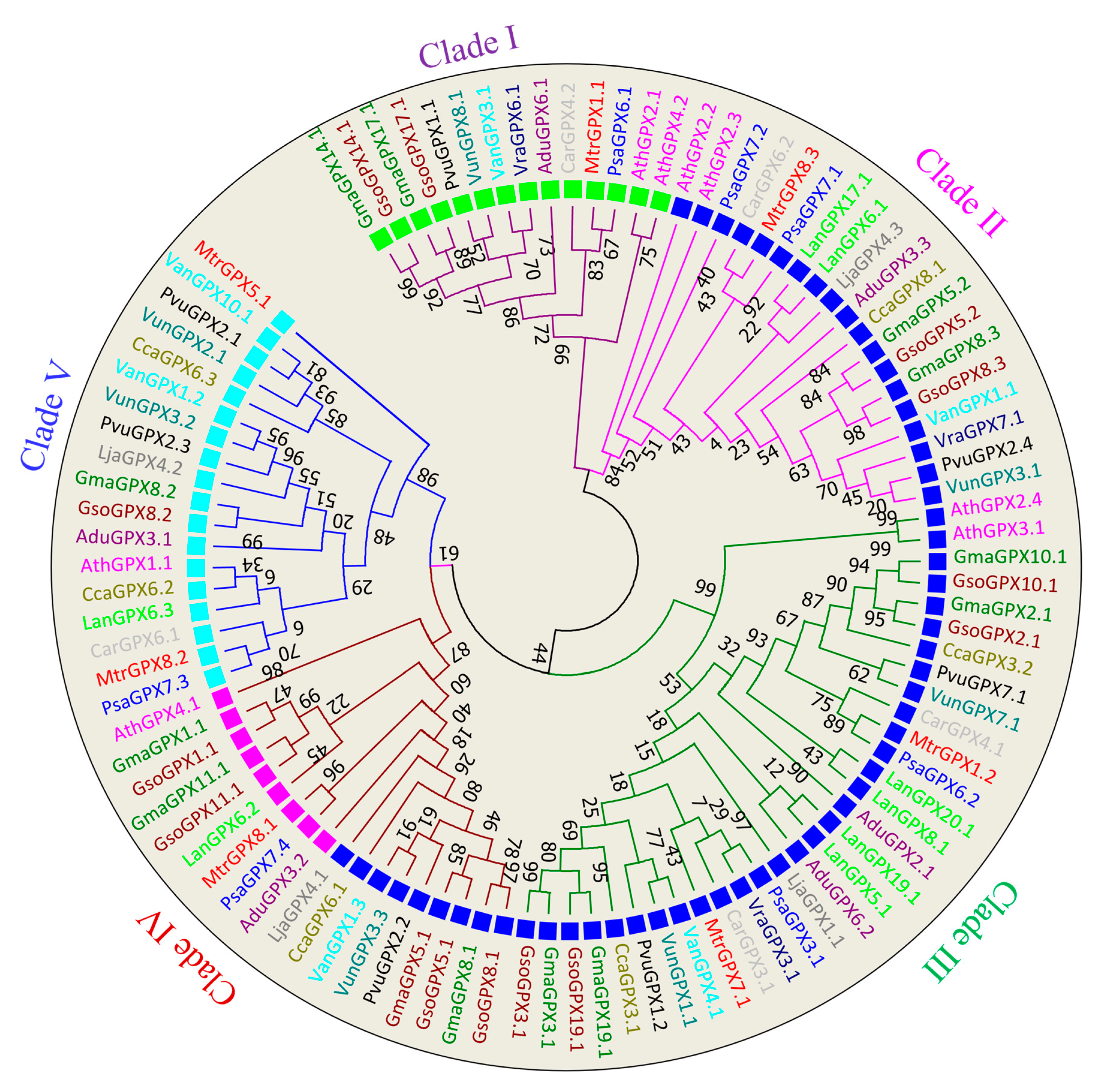
3.2. Phylogenetic Analysis of SOD and GPX Genes
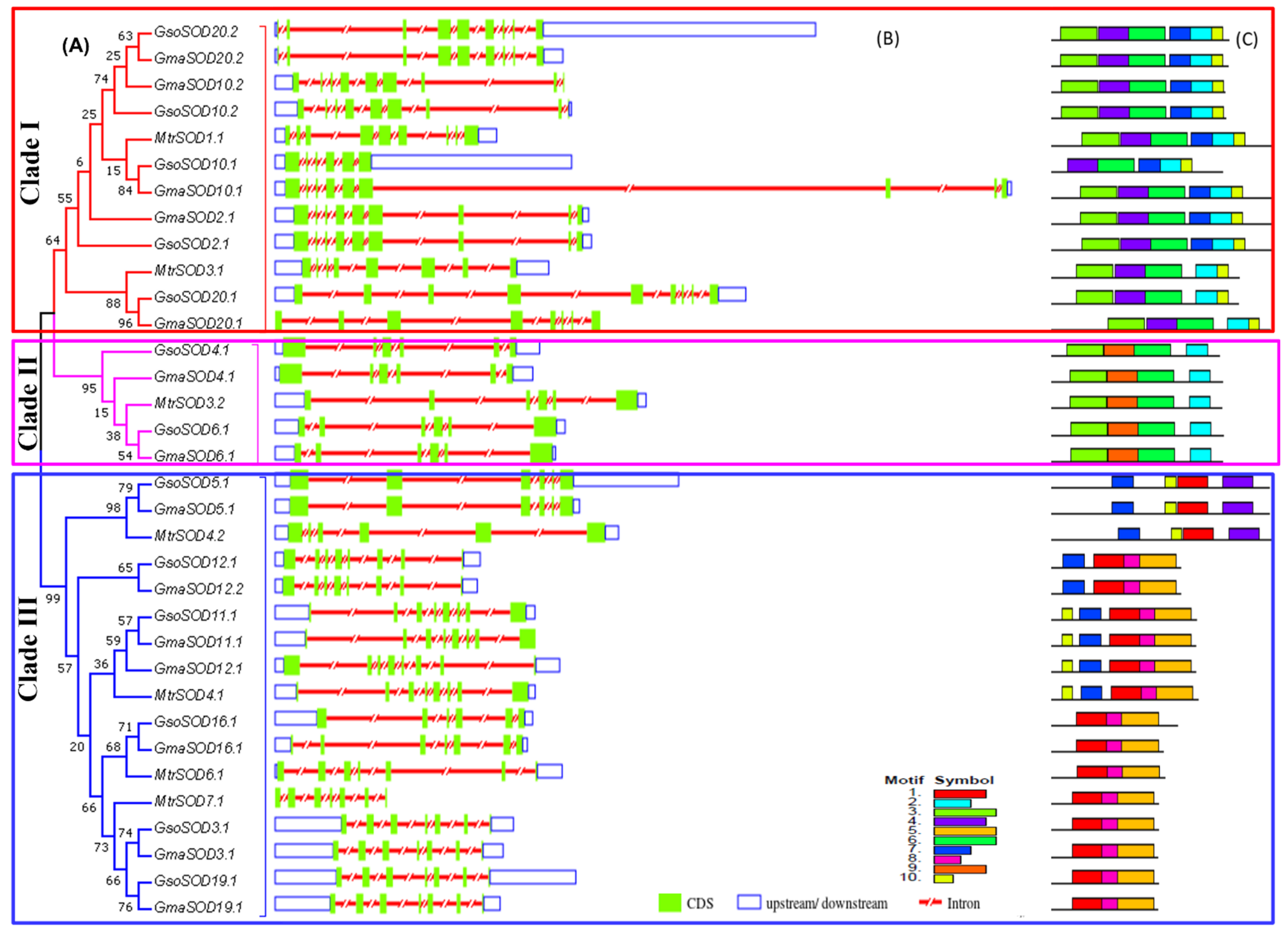
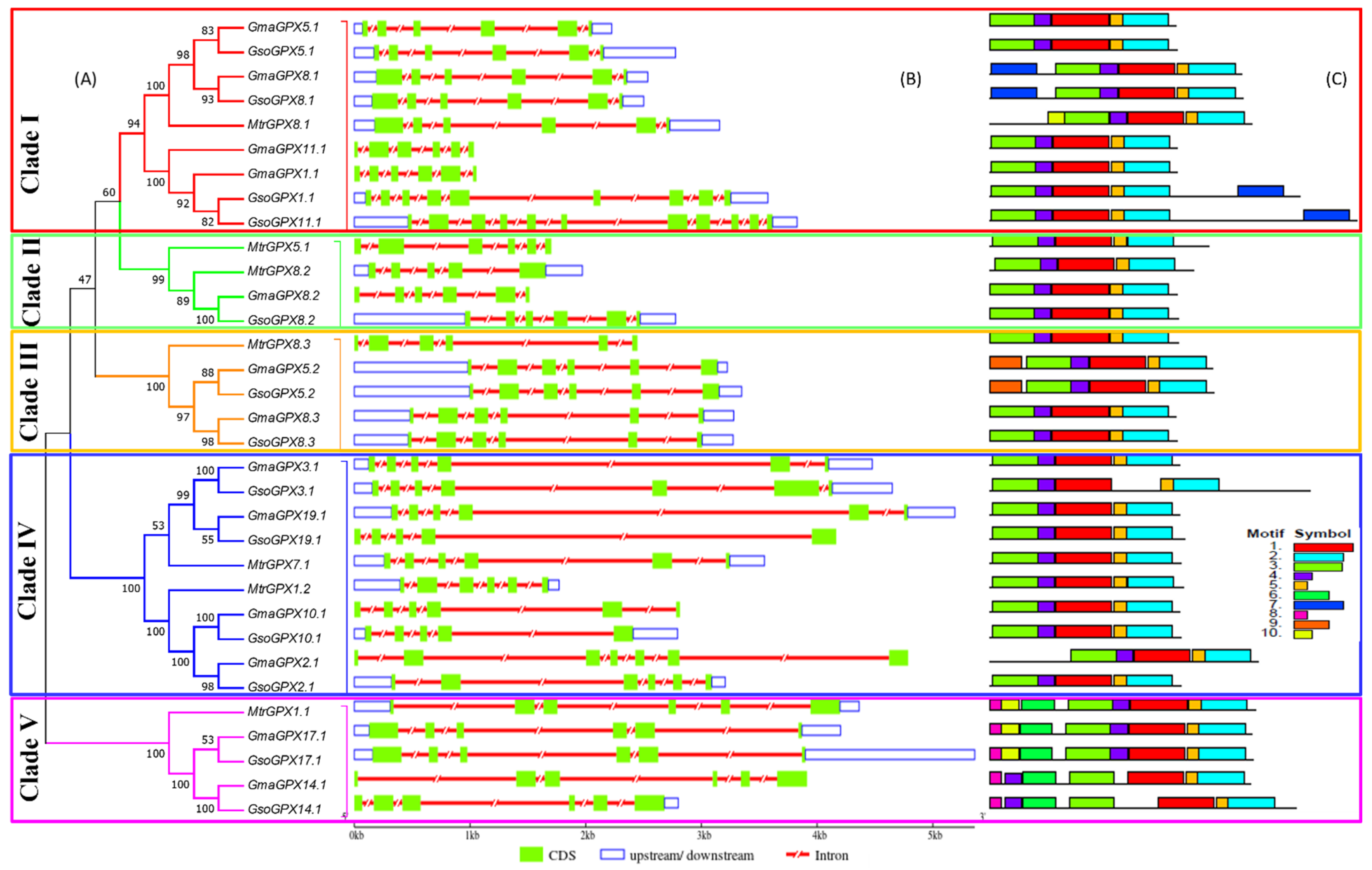
3.3. Exon–Intron Structures Analysis of SOD and GPX Genes in G. max, G. soja and Medicago truncatula
3.4. Conserved Motifs of SOD and GPX Genes in G. max, G. soja and Medicago truncatula
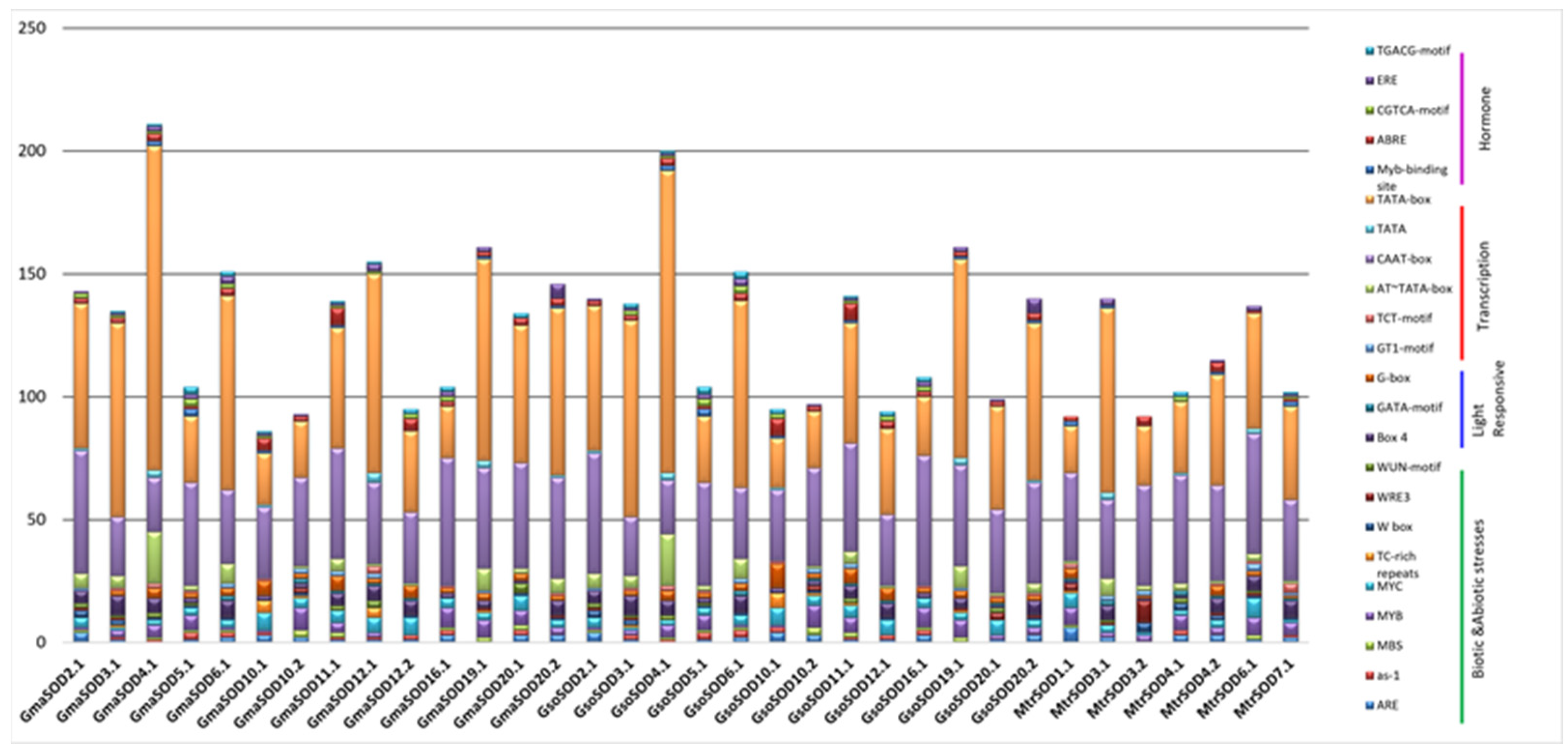
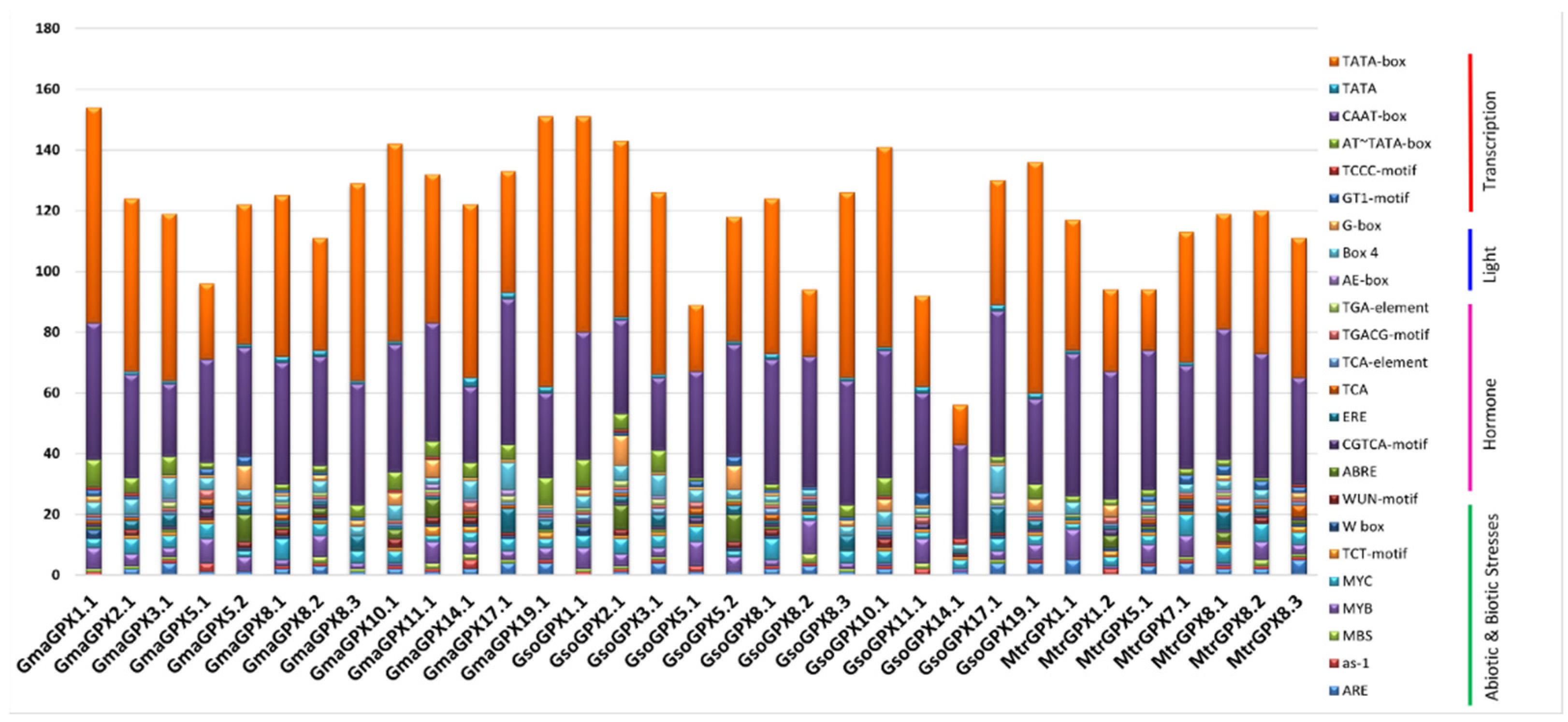
3.5. Promoter Analysis
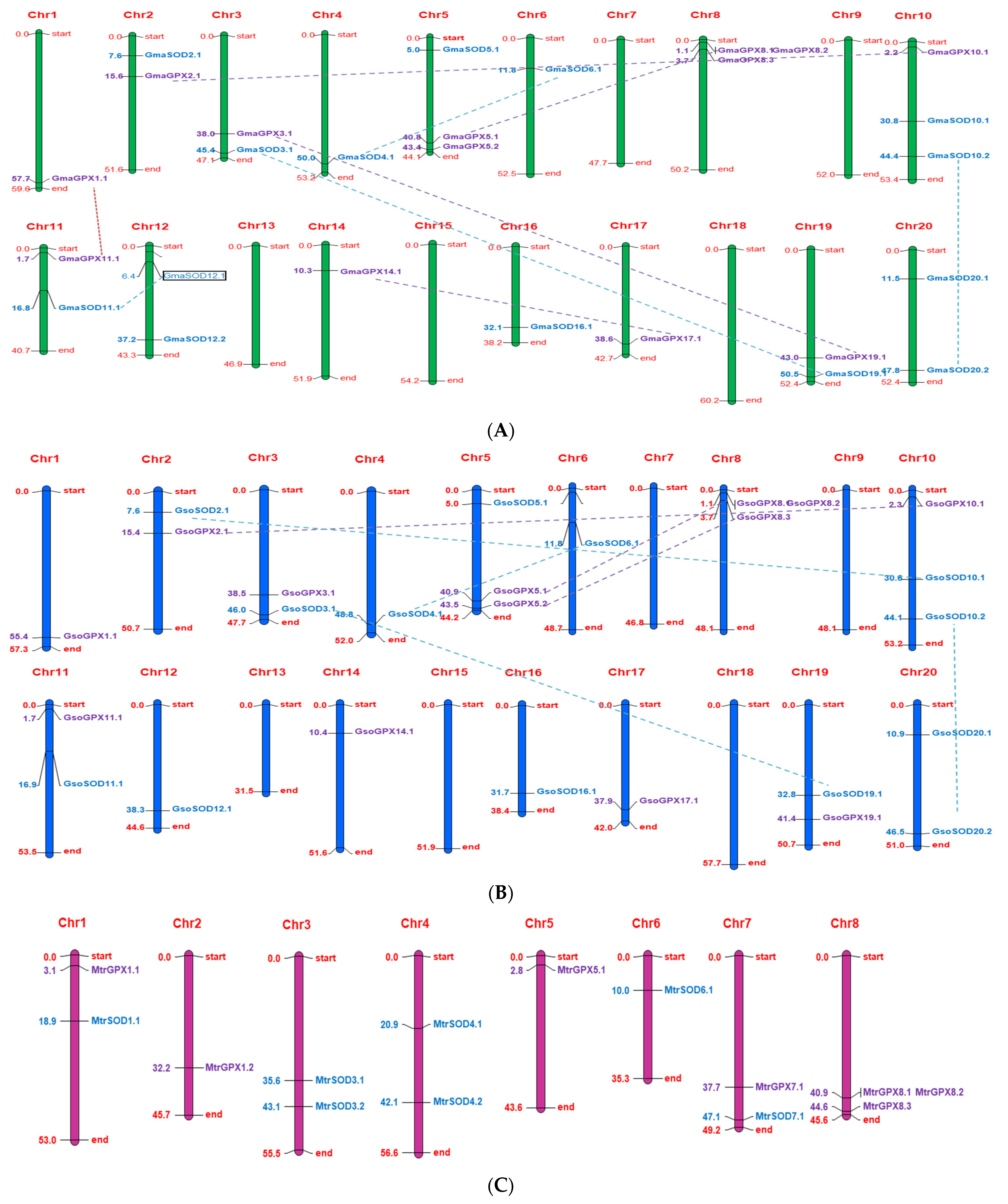
3.6. Chromosome Localization
3.7. Gene Duplication
3.8. Divergence Rate of SOD Genes in G. max and G. soja
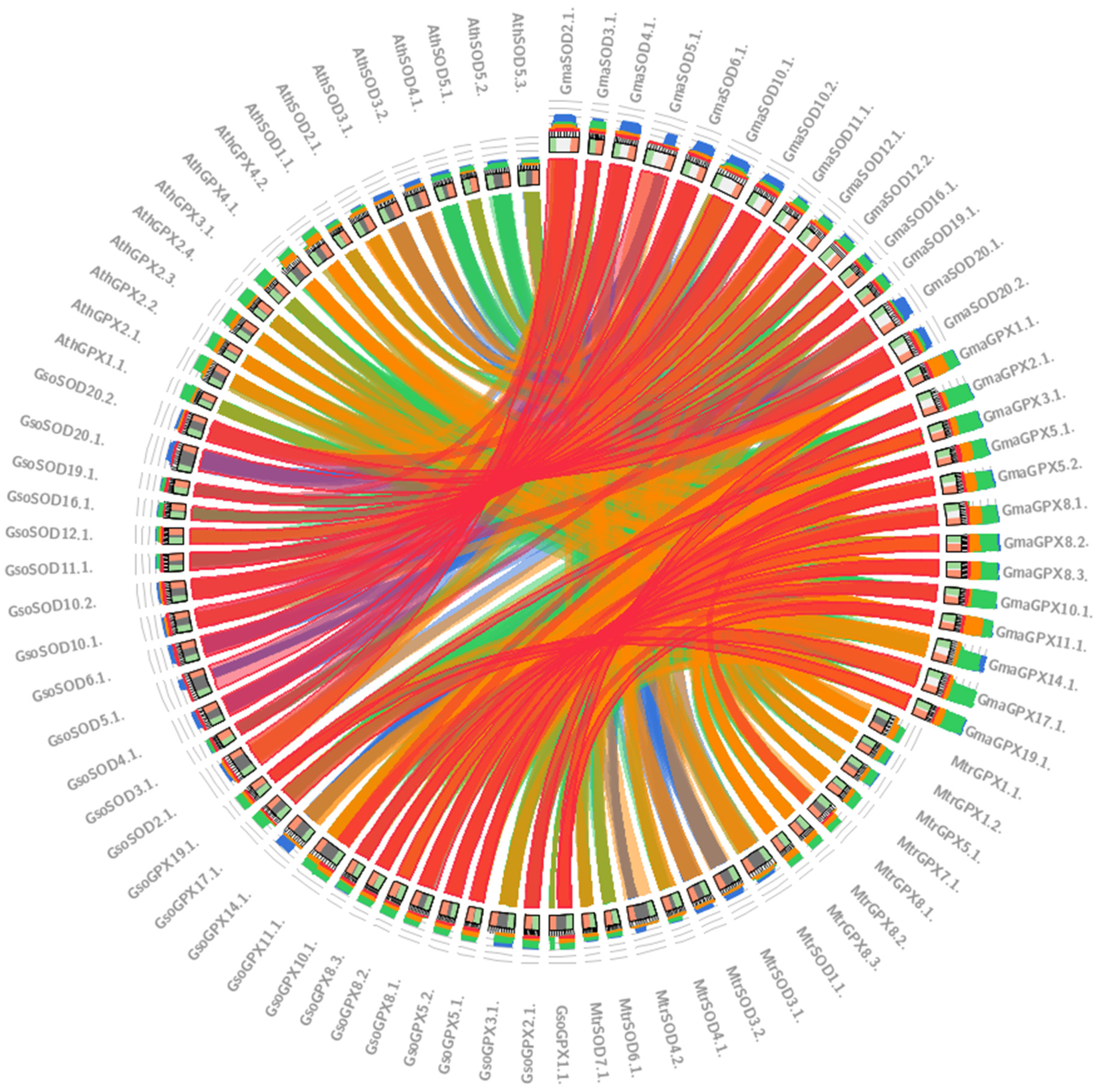
3.9. Relationships between GmaSODs with GsoSODs, MtrSODs and AthSODs
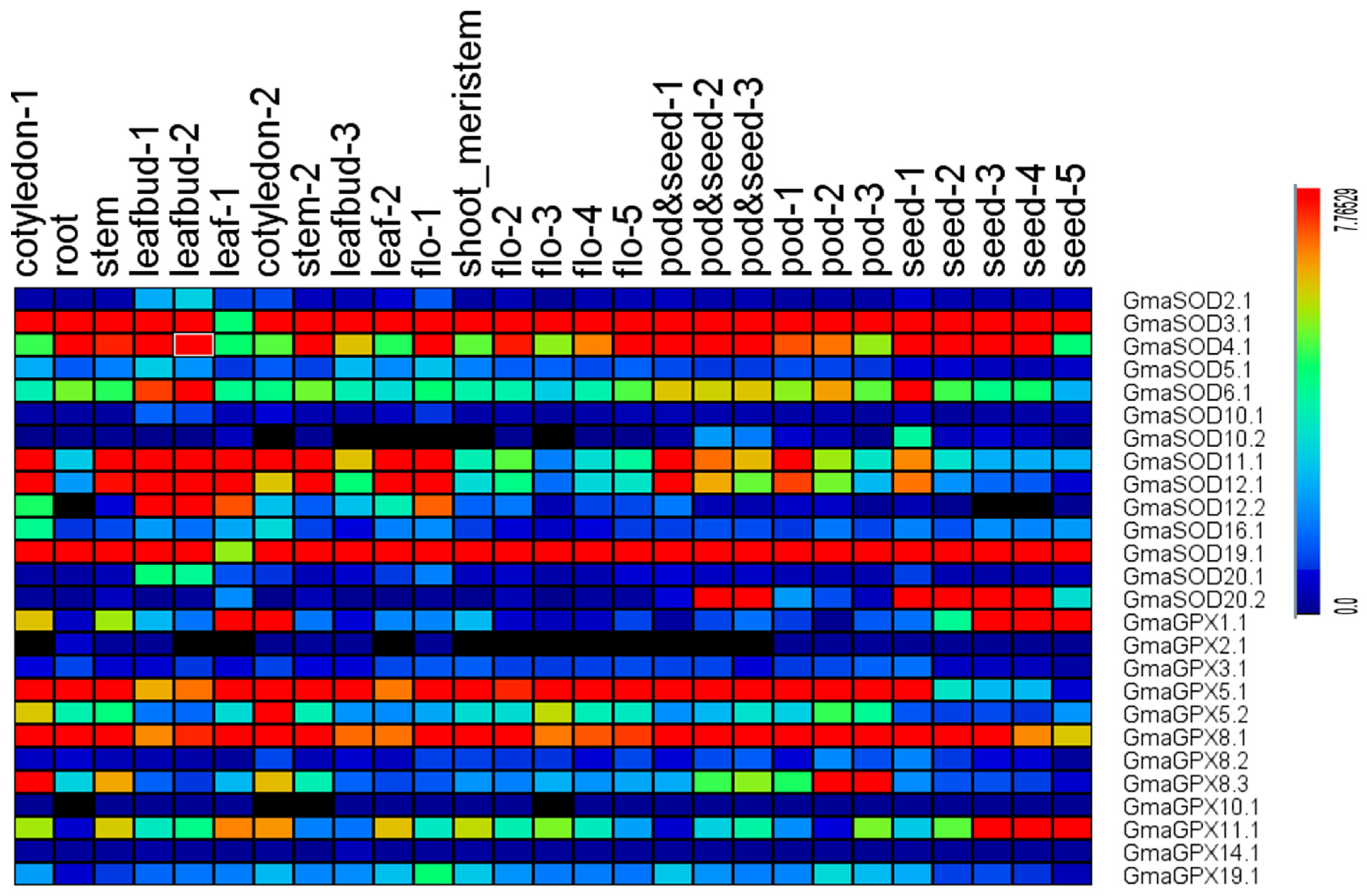
3.10. Expression Profiling of SOD and GPX Genes in Different Soybean (Glycine max L.) Tissues
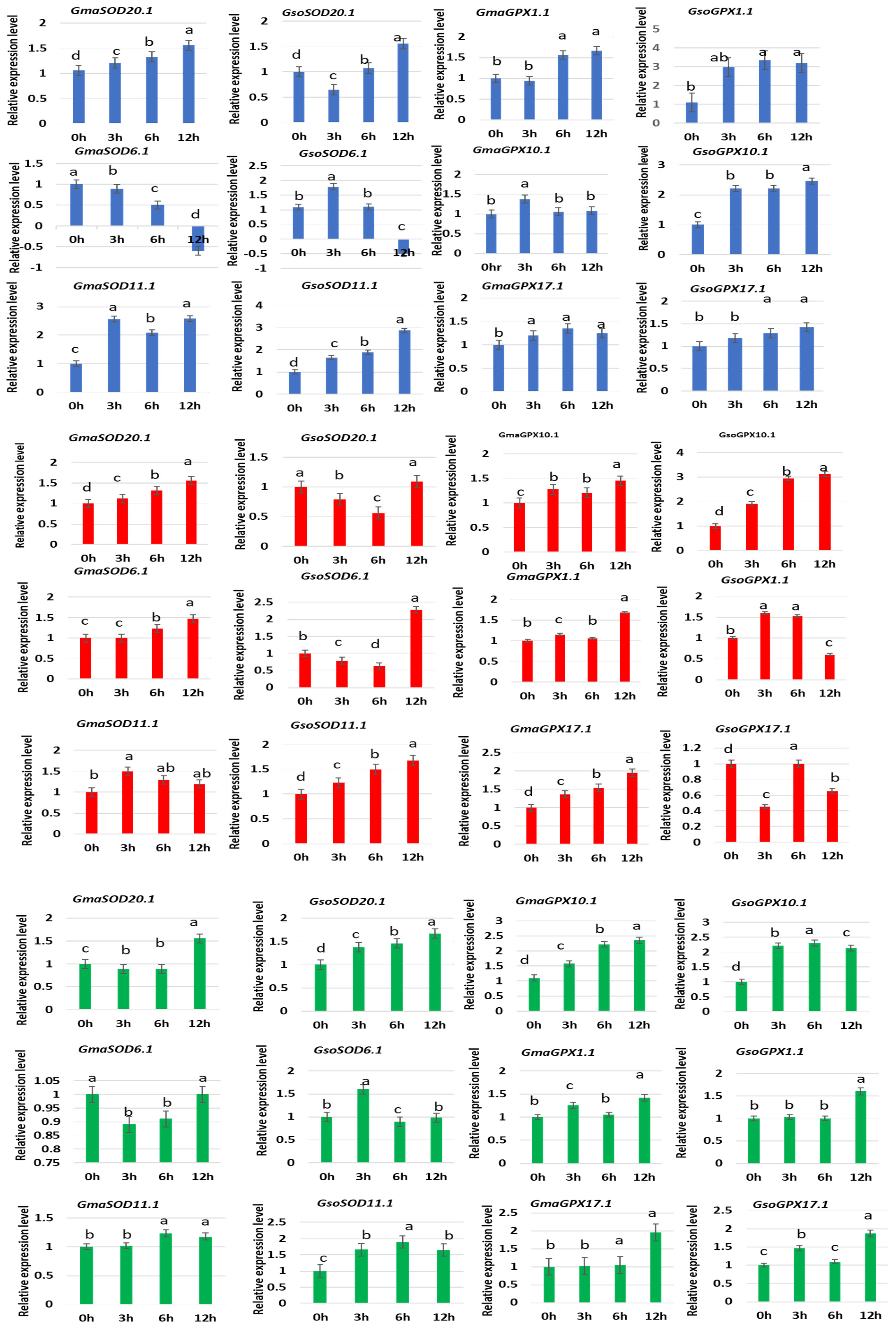
3.11. Response of GsoSOD and GsoGPX Genes to Different Abiotic Stresses
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Bhat, J.A.; Shivaraj, S.; Singh, P.; Navadagi, D.B.; Tripathi, D.K.; Dash, P.K.; Solanke, A.U.; Sonah, H.; Deshmukh, R. Role of silicon in mitigation of heavy metal stresses in crop plants. Plants 2019, 8, 71. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bhat, J.A.; Deshmukh, R.; Zhao, T.; Patil, G.; Deokar, A.; Shinde, S.; Chaudhary, J. Harnessing High-throughput Phenotyping and Genotyping for Enhanced Drought Tolerance in Crop Plants. J. Biotechnol. 2020, 324, 248–260. [Google Scholar] [CrossRef] [PubMed]
- Das, K.; Roychoudhury, A. Reactive oxygen species (ROS) and response of antioxidants as ROS-scavengers during environmental stress in plants. Front. Environ. Sci. 2014, 2, 53. [Google Scholar] [CrossRef] [Green Version]
- Molassiotis, A.; Job, D.; Ziogas, V.; Tanou, G. Citrus plants: A model system for unlocking the secrets of NO and ROS-inspired priming against salinity and drought. Front. Plant Sci. 2016, 7, 229. [Google Scholar] [CrossRef] [Green Version]
- Foyer, C.H.; Noctor, G. Redox homeostasis and antioxidant signaling: A metabolic interface between stress perception and physiological responses. Plant Cell 2005, 17, 1866–1875. [Google Scholar] [CrossRef] [Green Version]
- Gupta, S.; Dong, Y.; Dijkwel, P.P.; Mueller-Roeber, B.; Gechev, T.S. Genome-Wide Analysis of ROS Antioxidant Genes in Resurrection Species Suggest an Involvement of Distinct ROS Detoxification Systems during Desiccation. Int. J. Mol. Sci. 2019, 20, 3101. [Google Scholar] [CrossRef] [Green Version]
- Wang, F.-Z.; Wang, Q.-B.; Kwon, S.-Y.; Kwak, S.-S.; Su, W.-A. Enhanced drought tolerance of transgenic rice plants expressing a pea manganese superoxide dismutase. J. Plant Physiol. 2005, 162, 465–472. [Google Scholar] [CrossRef]
- Jalali-e-Emam, S.M.S.; Alizadeh, B.; Zaefizadeh, M.; Zakarya, R.A.; Khayatnezhad, M. Superoxide dismutase (SOD) activity in NaCl stress in salt-sensitive and salt-tolerance genotypes of Colza (Brassica napus L.). Middle East J. Sci. Res. 2011, 7, 7–11. [Google Scholar]
- De Rossi, S.; Di Marco, G.; Bruno, L.; Gismondi, A.; Canini, A. Investigating the drought and salinity effect on the redox components of Sulla Coronaria (L.) Medik. Antioxidants 2021, 10, 1048. [Google Scholar] [CrossRef]
- Verma, D.; Lakhanpal, N.; Singh, K. Genome-wide identification and characterization of abiotic-stress responsive SOD (superoxide dismutase) gene family in Brassica juncea and B. rapa. BMC Genom. 2019, 20, 227. [Google Scholar] [CrossRef] [Green Version]
- Kliebenstein, D.J.; Monde, R.-A.; Last, R.L. Superoxide dismutase in Arabidopsis: An eclectic enzyme family with disparate regulation and protein localization. Plant Physiol. 1998, 118, 637–650. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, W.; Zhang, X.; Deng, F.; Yuan, R.; Shen, F. Genome-wide characterization and expression analyses of superoxide dismutase (SOD) genes in Gossypium hirsutum. BMC Genom. 2017, 18, 376. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tyagi, S.; Sharma, S.; Taneja, M.; Kumar, R.; Sembi, J.K.; Upadhyay, S.K. Superoxide dismutases in bread wheat (Triticum aestivum L.): Comprehensive characterization and expression analysis during development and, biotic and abiotic stresses. Agric. Gene 2017, 6, 1–13. [Google Scholar] [CrossRef]
- Bela, K.; Horváth, E.; Gallé, Á.; Szabados, L.; Tari, I.; Csiszár, J. Plant glutathione peroxidases: Emerging role of the antioxidant enzymes in plant development and stress responses. J. Plant Physiol. 2015, 176, 192–201. [Google Scholar] [CrossRef]
- Avsian-Kretchmer, O.; Gueta-Dahan, Y.; Lev-Yadun, S.; Gollop, R.; Ben-Hayyim, G. The salt-stress signal transduction pathway that activates the gpx1 promoter is mediated by intracellular H2O2, different from the pathway induced by extracellular H2O2. Plant Physiol. 2004, 135, 1685–1696. [Google Scholar] [CrossRef] [Green Version]
- Li, W.; Huai, X.; Li, P.; Raza, A.; Mubarik, M.S.; Habib, M.; Fiaz, S.; Zhang, B.; Pan, J.; Khan, R.S.A. Genome-wide characterization of glutathione peroxidase (GPX) gene family in rapeseed (brassica napus L.) revealed their role in multiple abiotic stress response and hormone signaling. Antioxidants 2021, 10, 1481. [Google Scholar] [CrossRef]
- Zhang, L.; Wu, M.; Teng, Y.; Jia, S.; Yu, D.; Wei, T.; Chen, C.; Song, W. Overexpression of the glutathione peroxidase 5 (RcGPX5) gene from rhodiola crenulata increases drought tolerance in Salvia miltiorrhiza. Front. Plant Sci. 2019, 9, 1950. [Google Scholar] [CrossRef]
- Diao, Y.; Xu, H.; Li, G.; Yu, A.; Yu, X.; Hu, W.; Zheng, X.; Li, S.; Wang, Y.; Hu, Z. Cloning a glutathione peroxidase gene from Nelumbo nucifera and enhanced salt tolerance by overexpressing in rice. Mol. Biol. Rep. 2014, 41, 4919–4927. [Google Scholar] [CrossRef]
- Matamoros, M.A.; Saiz, A.; Penuelas, M.; Bustos-Sanmamed, P.; Mulet, J.M.; Barja, M.V.; Rouhier, N.; Moore, M.; James, E.K.; Dietz, K.J.; et al. Function of glutathione peroxidases in legume root nodules. J. Exp. Bot. 2015, 66, 2979–2990. [Google Scholar] [CrossRef] [Green Version]
- Tokarz, B.; Wójtowicz, T.; Makowski, W.; Jędrzejczyk, R.J.; Tokarz, K.M. What is the Difference between the Response of Grass Pea (Lathyrus sativus L.) to Salinity and Drought Stress?—A Physiological Study. Agronomy 2020, 10, 833. [Google Scholar] [CrossRef]
- Chen, M.; Li, K.; Li, H.; Song, C.P.; Miao, Y. The Glutathione Peroxidase Gene Family in Gossypium hirsutum: Genome-Wide Identification, Classification, Gene Expression and Functional Analysis. Sci. Rep. 2017, 7, 44743. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, Y.; Hu, L.; Ye, S.; Jiang, L.; Liu, S. Genome-wide identification of glutathione peroxidase (GPX) gene family and their response to abiotic stress in cucumber. 3 Biotech 2018, 8, 159. [Google Scholar] [CrossRef] [PubMed]
- Dash, S.; Campbell, J.D.; Cannon, E.K.; Cleary, A.M.; Huang, W.; Kalberer, S.R.; Karingula, V.; Rice, A.G.; Singh, J.; Umale, P.E. Legume information system (LegumeInfo. org): A key component of a set of federated data resources for the legume family. Nucleic Acids Res. 2015, 44, D1181–D1188. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gepts, P.; Beavis, W.D.; Brummer, E.C.; Shoemaker, R.C.; Stalker, H.T.; Weeden, N.F.; Young, N.D. Legumes as a model plant family. Genomics for food and feed report of the Cross-Legume Advances Through Genomics Conference. Plant Physiol. 2005, 137, 1228–1235. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nawaz, M.A.; Yang, S.H.; Chung, G. Wild soybeans: An opportunistic resource for soybean improvement. In Rediscovery of Landraces as a Resource for the Future; Intech Open: London, UK, 2018. [Google Scholar]
- Ma, T.; Wang, J.; Zhou, G.; Yue, Z.; Hu, Q.; Chen, Y.; Liu, B.; Qiu, Q.; Wang, Z.; Zhang, J. Genomic insights into salt adaptation in a desert poplar. Nat. Commun. 2013, 4, 2797. [Google Scholar] [CrossRef] [PubMed]
- Ozyigit, I.I.; Filiz, E.; Vatansever, R.; Kurtoglu, K.Y.; Koc, I.; Ozturk, M.X.; Anjum, N.A. Identification and Comparative Analysis of H2O2-Scavenging Enzymes (Ascorbate Peroxidase and Glutathione Peroxidase) in Selected Plants Employing Bioinformatics Approaches. Front. Plant Sci. 2016, 7, 301. [Google Scholar] [CrossRef] [Green Version]
- Schultz, J.; Copley, R.R.; Doerks, T.; Ponting, C.P.; Bork, P. SMART: A web-based tool for the study of genetically mobile domains. Nucleic Acids Res. 2000, 28, 231–234. [Google Scholar] [CrossRef] [PubMed]
- Gasteiger, E.; Hoogland, C.; Gattiker, A.; Wilkins, M.R.; Appel, R.D.; Bairoch, A. Protein identification and analysis tools on the ExPASy server. In The Proteomics Protocols Handbook; Springer: Berlin/Heidelberg, Germany, 2005; pp. 571–607. [Google Scholar]
- Chi, S.-M.; Nam, D. WegoLoc: Accurate prediction of protein subcellular localization using weighted Gene Ontology terms. Bioinformatics 2012, 28, 1028–1030. [Google Scholar] [CrossRef]
- Yu, C.S.; Chen, Y.C.; Lu, C.H.; Hwang, J.K. Prediction of protein subcellular localization. Proteins Struct. Funct. Bioinform. 2006, 64, 643–651. [Google Scholar] [CrossRef]
- Sievers, F.; Higgins, D.G. Clustal Omega for making accurate alignments of many protein sequences. Protein Sci. 2018, 27, 135–145. [Google Scholar] [CrossRef] [Green Version]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular evolutionary genetics analysis version 7.0 for bigger datasets. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hu, B.; Jin, J.; Guo, A.-Y.; Zhang, H.; Luo, J.; Gao, G. GSDS 2.0: An upgraded gene feature visualization server. Bioinformatics 2014, 31, 1296–1297. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bailey, T.L.; Williams, N.; Misleh, C.; Li, W.W. MEME: Discovering and analyzing DNA and protein sequence motifs. Nucleic Acids Res. 2006, 34, W369–W373. [Google Scholar] [CrossRef] [PubMed]
- Lescot, M.; Déhais, P.; Thijs, G.; Marchal, K.; Moreau, Y.; Van de Peer, Y.; Rouzé, P.; Rombauts, S. PlantCARE, a database of plant cis-acting regulatory elements and a portal to tools for in silico analysis of promoter sequences. Nucleic Acids Res. 2002, 30, 325–327. [Google Scholar] [CrossRef] [PubMed]
- Voorrips, R. MapChart: Software for the graphical presentation of linkage maps and QTLs. J. Hered. 2002, 93, 77–78. [Google Scholar] [CrossRef] [Green Version]
- Mifsud, W.; Bateman, A. Membrane-bound progesterone receptors contain a cytochrome b 5-like ligand-binding domain. Genome Biol. 2002, 3, research0068. [Google Scholar] [CrossRef] [PubMed]
- Darzentas, N. Circoletto: Visualizing sequence similarity with Circos. Bioinformatics 2010, 26, 2620–2621. [Google Scholar] [CrossRef]
- Korber, B. Computational analysis of HIV molecular sequences. In HIV Signature and Sequence Variation Analysis; Kluwer Academic Publishers: Dordrecht, The Netherlands, 2000; p. 55. [Google Scholar]
- Lynch, M.; Conery, J.S. The evolutionary fate and consequences of duplicate genes. Science 2000, 290, 1151–1155. [Google Scholar] [CrossRef] [Green Version]
- Shen, Y.; Du, H.; Liu, Y.; Ni, L.; Wang, Z.; Liang, C.; Tian, Z. Update soybean Zhonghuang 13 genome to a golden reference. Sci. China Life Sci. 2019, 62, 1257–1260. [Google Scholar] [CrossRef] [PubMed]
- Hoagland, D.R.; Arnon, D.I. The water-culture method for growing plants without soil. Circular. Calif. Agric. Exp. Stn. 1950, 347, 32. [Google Scholar]
- Li, Y.; Chen, Q.; Nan, H.; Li, X.; Lu, S.; Zhao, X.; Liu, B.; Guo, C.; Kong, F.; Cao, D. Overexpression of GmFDL19 enhances tolerance to drought and salt stresses in soybean. PLoS ONE 2017, 12, e0179554. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCT method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Zhou, C.; Zhu, C.; Fu, H.; Li, X.; Chen, L.; Lin, Y.; Lai, Z.; Guo, Y. Genome-wide investigation of superoxide dismutase (SOD) gene family and their regulatory miRNAs reveal the involvement in abiotic stress and hormone response in tea plant (Camellia sinensis). PLoS ONE 2019, 14, e0223609. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.; Xia, M.; Chen, J.; Deng, F.; Yuan, R.; Zhang, X.; Shen, F. Genome-wide analysis of superoxide dismutase gene family in Gossypium raimondii and G. arboreum. Plant Gene 2016, 6, 18–29. [Google Scholar] [CrossRef] [Green Version]
- Hu, X.; Hao, C.; Cheng, Z.M.; Zhong, Y. Genome-Wide Identification, Characterization, and Expression Analysis of the Grapevine Superoxide Dismutase (SOD) Family. Int. J. Genom. 2019, 2019, 7350414. [Google Scholar] [CrossRef]
- Mok, D.; Peloquin, S. Breeding value of 2n pollen (diplandroids) in tetraploid x diploid crosses in potatoes. Theor. Appl. Genet. 1975, 46, 307–314. [Google Scholar] [CrossRef]
- Jiang, W.; Yang, L.; He, Y.; Zhang, H.; Li, W.; Chen, H.; Ma, D.; Yin, J. Genome-wide identification and transcriptional expression analysis of superoxide dismutase (SOD) family in wheat (Triticum aestivum). PeerJ 2019, 7, e8062. [Google Scholar] [CrossRef] [Green Version]
- Kondrashov, F.A. Gene duplication as a mechanism of genomic adaptation to a changing environment. Proc. R. Soc. B Biol. Sci. 2012, 279, 5048–5057. [Google Scholar] [CrossRef] [Green Version]
- Bhat, J.A.; Shivaraj, S.; Ali, S.; Mir, Z.A.; Islam, A.; Deshmukh, R. Genomic resources and omics-assisted breeding approaches for pulse crop improvement. In Pulse Improvement; Springer: Cham, Switzerland, 2018; pp. 13–55. [Google Scholar]
- Magadum, S.; Banerjee, U.; Murugan, P.; Gangapur, D.; Ravikesavan, R. Gene duplication as a major force in evolution. J. Genet. 2013, 92, 155–161. [Google Scholar] [CrossRef]
- Wang, Y.; Wang, Q.; Zhao, Y.; Han, G.; Zhu, S. Systematic analysis of maize class III peroxidase gene family reveals a conserved subfamily involved in abiotic stress response. Gene 2015, 566, 95–108. [Google Scholar] [CrossRef]
- Xu, Y.; Magwanga, R.O.; Cai, X.; Zhou, Z.; Wang, X.; Wang, Y.; Zhang, Z.; Jin, D.; Guo, X.; Wei, Y.; et al. Deep Transcriptome Analysis Reveals Reactive Oxygen Species (ROS) Network Evolution, Response to Abiotic Stress, and Regulation of Fiber Development in Cotton. Int. J. Mol. Sci. 2019, 20, 1863. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Milla, M.A.R.; Maurer, A.; Huete, A.R.; Gustafson, J.P. Glutathione peroxidase genes in Arabidopsis are ubiquitous and regulated by abiotic stresses through diverse signaling pathways. Plant J. 2003, 36, 602–615. [Google Scholar] [CrossRef] [PubMed]
- Abdel-Wahab, A.; Shabeb, M.; Younis, M. Studies on the effect of salinity, drought stress and soil type on nodule activities of Lablab purpureus (L.) sweet (Kashrangeeg). J. Arid. Environ. 2002, 51, 587–602. [Google Scholar] [CrossRef]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sr. No. | Crop Plant | Species Studied | Abbreviation of Spp. Name | Link | Link Last Accessed Date |
|---|---|---|---|---|---|
| 1 | Cultivated Soybean | Glycine max | G. max | https://soybase.org/ | 12 October 2021 |
| 2 | Wild Soybean | Glycine soja | G. soja | https://soybase.org/ | 12 October 2021 |
| 3 | Barrel clover | Medicago truncatula | M. truncatula | http://www.medicagogenome.org/download | 8 April 2021 |
| 4 | Chickpea | Ciser arientinum | C. arientinum | https://legumeinfo.org/ | 22 August 2021 |
| 5 | Pigeonpea | Cajanus cajan | C. cajan | https://legumeinfo.org/ | 22 August 2021 |
| 6 | Wild peanut | Arachis duranensis | duranis | https://legumeinfo.org/ | 22 August 2021 |
| 7 | Pea | Pisum sativum | P. sativum | https://legumeinfo.org/ | 22 August 2021 |
| 8 | Cowpea | Vigna. unguiculata | V. unguiculata | https://legumeinfo.org/ | 22 August 2021 |
| 9 | Mungbean | Vigna radiate | V. radiata | https://legumeinfo.org/ | 22 August 2021 |
| 10 | Birdsfoot trefoil | Lotus japonicus | L. japonicus | https://legumeinfo.org/ | 22 August 2021 |
| 11 | Narrowleaf lupin | Lupinus angustifolius | L angustifolius | https://legumeinfo.org/ | 22 August 2021 |
| 12 | Adzuki bean | Vigna angularis | Vigna angularis | https://legumeinfo.org/ | 22 August 2021 |
| 13 | Common bean | Phaseolus vulgaris | P. vulgaris | https://legumeinfo.org/ | 22 August 2021 |
| 14 | Arabidopsis | Arabidopsis thaliana | A. thaliana | https://www.arabidopsis.org/ | 2 May 2021 |
| Duplicated Pair | Ka | Ks | Ka/Ks | Duplicate Type | Time (Mya *) | Purifying Selection | Type |
|---|---|---|---|---|---|---|---|
| GsoSOD3.1 GsoSOD19.1 | 0.067 | 0.051 | 1.298 | Segmental | 4.18 | Yes | Positive selection |
| GsoSOD4.1 GsoSOD6.1 | 0.022 | 0.034 | 0.656 | Segmental | 2.79 | Yes | Negative or purifying selection |
| GsoSOD10.2 GsoSOD20.2 | 0.027 | 0.085 | 0.311 | Segmental | 6.97 | Yes | Negative or purifying selection |
| GsoSOD2.1 GsoSOD10.1 | 0.048 | 0.062 | 0.773 | Segmental | 5.08 | Yes | Negative or purifying selection |
| GmaSOD11.1 GmaSOD12.1 | 0.034 | 0.015 | 2.182 | Segmental | 1.23 | No | Positive selection |
| GmaSOD10. GmaSOD20.2 | 0.036 | 0.054 | 0.666 | Segmental | 4.43 | Yes | Negative or purifying selection |
| GmaSOD3.1 GmaSOD19.1 | 0.066 | 0.037 | 1.811 | Segmental | 3.03 | No | Positive selection |
| GmaSOD4.1 GmaSOD6.1 | 0.025 | 0.031 | 0.807 | Segmental | 2.54 | Yes | Negative or purifying selection |
| GsoGPX5.2 GsoGPX8.3 | 0.018 | 0.090 | 0.197 | Segmental | 7.38 | Yes | Negative or purifying selection |
| GsoGPX5.1 GsoGPX8.1 | 0.005 | 0.075 | 0.069 | Segmental | 6.15 | Yes | Negative or purifying selection |
| GsoGPX2.1 GsoGPX10.1 | 0.017 | 0.136 | 0.128 | Segmental | 11.15 | Yes | Negative or purifying selection |
| GmaGPX8.1 GmaGPX5.1 | 0.021 | 0.020 | 1.051 | Segmental | 1.64 | No | Positive selection |
| GmaGPX14.1 GmaGPX17.1 | 0.059 | 0.062 | 0.941 | Segmental | 5.08 | Yes | Negative or purifying selection |
| GmaGPX5.2 GmaGPX8.3 | 0.033 | 0.000 | 0.000 | Segmental | 0.00 | Yes | Negative or purifying selection |
| GmaGPX2.1 GmaGPX10.1 | 0.050 | 0.019 | 2.637 | Segmental | 1.56 | No | Positive selection |
| GmaGPX11.1 GmaGPX1.1 | 0.013 | 0.010 | 1.306 | Segmental | 0.82 | No | Positive selection |
| GmaGPX3.1 GmaGPX19.1 | 0.061 | 0.063 | 0.970 | Segmental | 5.16 | No | Neutral selection |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Aleem, M.; Aleem, S.; Sharif, I.; Wu, Z.; Aleem, M.; Tahir, A.; Atif, R.M.; Cheema, H.M.N.; Shakeel, A.; Lei, S.; et al. Characterization of SOD and GPX Gene Families in the Soybeans in Response to Drought and Salinity Stresses. Antioxidants 2022, 11, 460. https://doi.org/10.3390/antiox11030460
Aleem M, Aleem S, Sharif I, Wu Z, Aleem M, Tahir A, Atif RM, Cheema HMN, Shakeel A, Lei S, et al. Characterization of SOD and GPX Gene Families in the Soybeans in Response to Drought and Salinity Stresses. Antioxidants. 2022; 11(3):460. https://doi.org/10.3390/antiox11030460
Chicago/Turabian StyleAleem, Muqadas, Saba Aleem, Iram Sharif, Zhiyi Wu, Maida Aleem, Ammara Tahir, Rana Muhammad Atif, Hafiza Masooma Naseer Cheema, Amir Shakeel, Sun Lei, and et al. 2022. "Characterization of SOD and GPX Gene Families in the Soybeans in Response to Drought and Salinity Stresses" Antioxidants 11, no. 3: 460. https://doi.org/10.3390/antiox11030460
APA StyleAleem, M., Aleem, S., Sharif, I., Wu, Z., Aleem, M., Tahir, A., Atif, R. M., Cheema, H. M. N., Shakeel, A., Lei, S., Yu, D., Wang, H., Kaushik, P., Alyemeni, M. N., Bhat, J. A., & Ahmad, P. (2022). Characterization of SOD and GPX Gene Families in the Soybeans in Response to Drought and Salinity Stresses. Antioxidants, 11(3), 460. https://doi.org/10.3390/antiox11030460