
The Relationship between Cancer and Paraoxonase 1

 , , and
, , and
Abstract
:1. Introduction
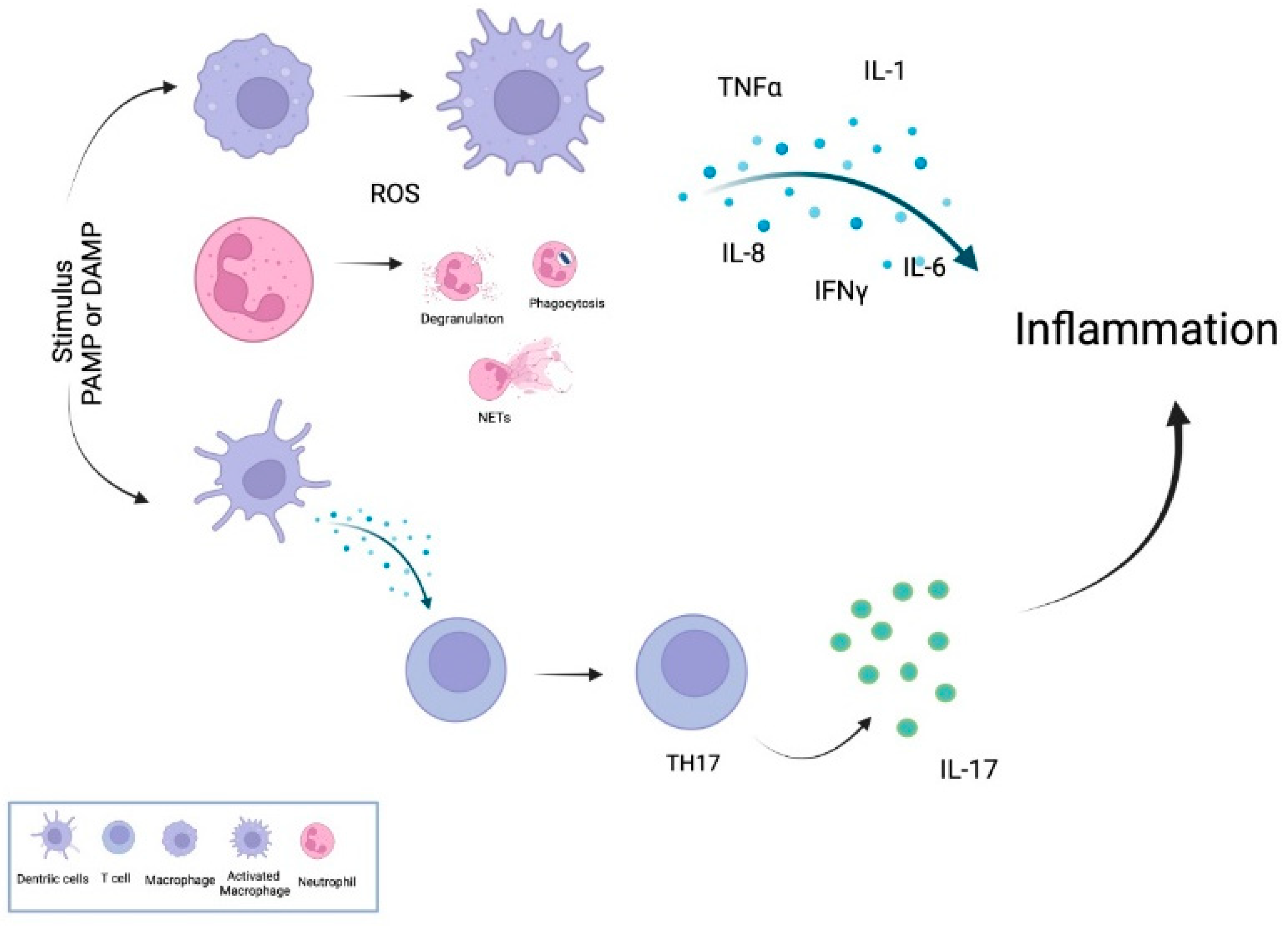
2. Inflammatory Mechanism
3. Genes Involved in Inflammatory Response
3.1. NF-κB Transcription
3.2. Signal Transducers and Activators of Transcription
3.3. Activator Protein 1
3.4. Hypoxia-Inducible Transcription Factor
3.5. Cyclooxygenases
3.6. RNA-Based Processes
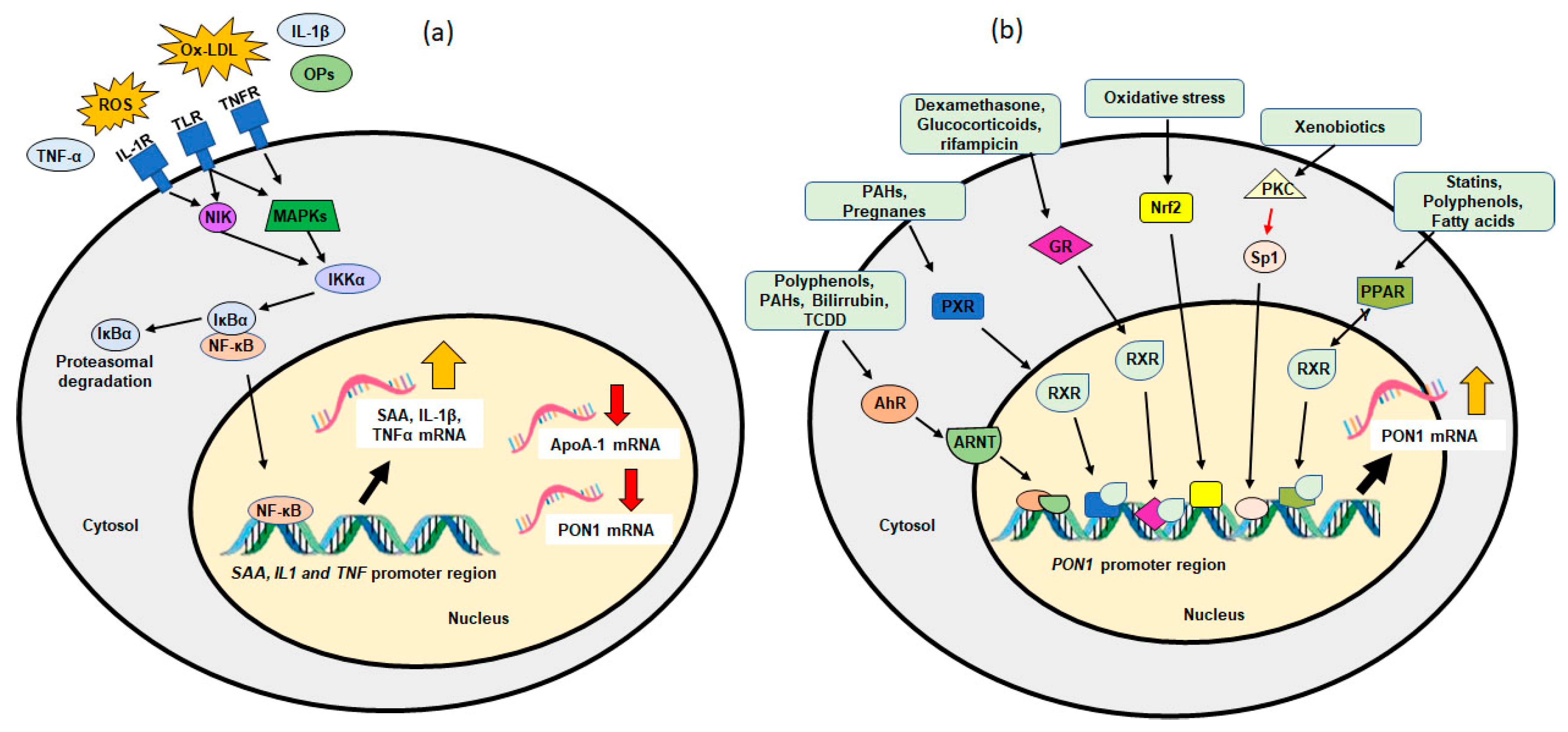
4. Proinflammatory Cytokines and PON1
5. Structure of the PON1 Gene and Polymorphism
5.1. PON1 Gene Structure
5.2. Single-Nucleotide Polymorphisms of the PON1 Gene
5.2.1. Polymorphism in the Promoter Region
5.2.2. Polymorphism in the Coding Region

{kind=link}
{kind=link}
| Variation | Allele Change | Residue Change | Rs Number | Location | Ref. |
|---|---|---|---|---|---|
| L55M | TTG ⇒ ATG | L [Leu] ⇒ M [Met] | rs854560 | Codon | [203] |
| Q192R | CAA ⇒ CGA | Q [Gln] ⇒ R [Arg] | rs662 | Codon | [204] |
| S23A | TCT ⇒ GCT | S [Ser] ⇒ A [Ala] | rs146211440 | Codon | [205] |
| A201V | GCG ⇒ GTG | A [Ala] ⇒ V [Val] | rs80019660 | Codon | [205] |
| P40L | CCT ⇒ CTT | P [Pro] ⇒ L [Leu] | rs141665531 | Codon | [205] |
| V109I | GTA ⇒ ATA | V [Val] ⇒ I [Ile] | rs61736513 | Codon | [205] |
| C-108T | NA | NA | rs705379 | Promoter | [206] |
| C-107T | NA | NA | Genbank, acc. number AF051133 | [168] | |
| C-126G | NA | NA | rs705380 | Promoter | [168] |
| G-162A | NA | NA | rs705381 | Promoter | [207] |
| C-761T | NA | NA | rs3735590 | Promoter | [208] |
| A-824G | NA | NA | Genbank, acc. number AF051133 | Promoter | [168] |
| G-832A | NA | NA | rs854571 | Promoter | [207] |
| G -909C | NA | NA | rs854572 | Promoter | [170,206] |
| A -1074G | NA | NA | rs705379 | Promoter | [173,209] |
| G -1266A | NA | NA | rs76283227 | Promoter | [206] |
| C -1434G | NA | NA | rs705382 | Promoter | [206] |
| A -1741G | NA | NA | rs757158 | Promoter | [206] |
6. Transcriptional Regulation of PON1
6.1. Endogenous Factors
6.1.1. PON1 and Aging
6.1.2. PON1 and Polymorphisms
6.1.3. PON1 and Disease
6.2. Exogenous Factors
6.2.1. Dietary Factors and PON1
6.2.2. Drugs and PON1
6.2.3. Environmental Pollutants and PON1
6.3. Epigenetics
| Subjects DNA Methylation | n | Analytical | Main Findings | Ref. |
|---|---|---|---|---|
| Obese adults with metabolic syndrome | 47 | Infinium Human Methylation 450 K BeadChip | A significant inverse correlation was found between the PON1 methylation in the CpG 1 (Chr position = 7:94,953,956), CpG 2 (Chr position = 7:94,954,059), CpG 3 (Chr position = 7:94,954,144) and CpG 4 (Chr position = 7:94,954,202) sites with the enzymatic AREase activity. | [297] |
| Age [mean ± SD] = 47 ± 10 years. | Negative correlations were found between the selected antioxidants (vitamin C, total tocopherols, and lycopene) with the percentage of methylation of the different PON1 gene CpG sites. | |||
| Patients with parenchymal ischemic stroke. | Pyrosequencing | No interaction was observed between the body composition and stroke diagnosis criteria for either of the analyzed CpG sites of the PON1 gene. No relevant changes were observed in the PON1 total methylation patterns considering stroke or obesity conditions. | [298] | |
| Age [mean ± SD] = 70 ± 12 years. | The CpGs at +15 and +241 bp in the PON1 promoter were related to weight, waist circumference, and energy intake in the group of patients without stoke, and an interaction was observed between the energy intake and total PON1 promoter methylation in the prediction of stroke condition (p = 0.017). | |||
| Children of a farmworker community (pesticide exposure). | 449 | Infinium Human Methylation 450 K BeadChip | Among sites in methylation block 1 (CpG sites 5, 8, 11, and 13), a separation by genotype was observed, providing evidence of allele-specific methylation. | [295] |
| Newborns and 9-year-old children | Strongly positive associations were found between PON1 −108 T alleles and methylation levels, particularly those in methylation block 1. Methylation at individual Block 1 CpG sites was significantly associated with AREase activity. | |||
| Newborns with prenatal mercury (Hg) exposure. | 321 | Infinium Human Methylation 450 K BeadChip | A DMR covering 9 CpG sites of Chr 7 in the PON1 gene was hypomethylated by prenatal Hg exposure among boys. In early childhood, a doubling in prenatal Hg concentration was associated with a 4.6% decrease in methylation levels of the DMR in PON1. Two CpG sites (cg07404485 and cg05342682) in the PON1 DMR located in the body of the gene and in the north shore region of a CpG island had the strongest association with expression evaluated in cord blood samples. | [300] |
| Children in early childhood (2.9–4.9 years) and mid-childhood (6.7–10.5 years). | ||||
| Children born by female greenhouse workers (pesticide exposure). | 48 | Infinium Human Methylation 450 K BeadChip array and pyrosequencing | Considering the effect modification by PON1 Q192R genotype, 767 significantly DMPs were identified, of which 128 were hypermethylated and 639 were hypomethylated. 5002 significant DMRs were identified, of which 2264 were hypermethylated and 2738 hypomethylated in the exposed PON1 192R carrier group compared to the other groups. The pyrosequencing methylation values were significantly more highly methylated in exposed children compared with the unexposed group carrying the PON1 192R-allele for most CpG sites. | [301] |
| Age: 6–11 years | ||||
| Patients with coronary artery disease. | 484 | Pyrosequencing | The PON1 −162 A>G genotype may significantly influence the methylation level at PON1 CpG site −162. Five CpG sites (positions −184, −170, −163, −161, and −142) exhibited hypomethylation in association with the occurrence of bleeding. Multivariate logistic regression analysis showed that methylation at PON1 site −161 and the use of angiotensin-converting enzyme inhibitors were associated with a decreased risk of bleeding events. | [299] |
| Age: 26–80 years | ||||
| Patients with overweight or obesity. | 790 | Pyrosequencing | Similar patterns between ten of the distinct CpGs for genetics, expression and activity across the 11 CpG sites were evaluated. Methylation levels varied between 6% and 60%; lower methylation levels were observed at CpG site −108 (10.4%) compared with the average promoter methylation (CpGmean = 27.9%). A significant association between average promoter hypermethylation and reduced expression on PON1. | [302] |
| Age: 18–74 years | ||||
| Histone modification | ||||
| In vivo study | 10 | ChIP analysis | At the promoter and coding regions, H3Ac was not significantly different between control and high fat offspring in males but slightly decreased in females. | [303] |
| Rats with two dietary treatments: control and high fat groups. | At the promoter, H4Ac and H3K4Me2 were significantly higher in both male and female high fat offspring compared with the control group. | |||
| Modifications in miRNA expression | ||||
| Patients with ischemic stroke. Age [mean ± SD] = 62.23 ± 11.72 years. | 2228 | MicroRNA expression profiling microarrays | miR-616 that binds to PON1 displayed increased expression in vascular smooth muscle cells treated with oxidized low-density lipoprotein and lipopolysaccharide. They found that miR-616 negatively regulates the expression of PON1. | [208] |
| Patients with coronary artery disease. Age: 24–79 years | 111 | Pathway-focused Human CVD miScript miRNA PCR array | Bivariate parametric correlation analysis showed that the serum PON1 activity correlated negatively with miR-486, miR-92a, and miR-122. The logistic regression model including miR-92a and miR-486 serum levels with adjustment for age, gender, serum lipids and apolipoproteins levels and PON1 activity as covariates resulted in a significant designation of vulnerable coronary artery disease patients with an accuracy of 84%. | [304] |
| Patients diagnosed with chronic obstructive pulmonary disease. | 292 | Quantitative real-time polymerase chain reaction | miR-616 regulates the PON1 expression in primary hepatocytes genotyped as CC. | [305] |
| Age [mean ± SD] = 60.22 ± 8.24 years. | miR-616 down-regulated the expression of PON1 and inhibited the PON1 activity in primary hepatocytes. | |||
| Patients with calcific aortic valve stenosis | 459 | Quantitative real-time polymerase chain reaction | The loci rs3735590 of PON1 3′UTR contains the binding site of miR-616. As a negative regulator of PON1, upregulation of miR-616 inhibited the expression of this gene, by binding to the 3′UTR of the mRNA. | [306] |
| Age [mean ± SD] = 72.2 ± 9.6 years. |
From Epigenetic Regulation of DNA to Cancer
| Subjects PON1 | n | Epigenetic Modification | Analytical Method | Main Findings | Ref. |
|---|---|---|---|---|---|
| Patients diagnosed with colorectal cancer (CRC). | 30 | Histone modification | Treatment with the lysine developer and analysis using a microplate reader |
| [313] |
| Patients with renal cell carcinoma (RCC). | 15 | DNA methylation | Methylation-specific polymerase chain reaction |
| [314] |
7. PON1 and Cancer
8. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Ďuračková, Z. Some Current Insights into Oxidative Stress. Physiol. Res. 2010, 59, 459–469. [Google Scholar] [CrossRef] [PubMed]
- Fang, J.; Seki, T.; Maeda, H. Therapeutic strategies by modulating oxygen stress in cancer and inflammation. Adv. Drug Deliv. Rev. 2009, 61, 290–302. [Google Scholar] [CrossRef] [PubMed]
- Khandrika, L.; Kumar, B.; Koul, S.; Maroni, P.; Koul, H.K. Oxidative stress in prostate cancer. Cancer Lett. 2009, 282, 125–136. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Visconti, R.; Grieco, D. New insights on oxidative stress in cancer. Curr. Opin. Drug Discov. Dev. 2009, 12, 240–245. [Google Scholar]
- Schraufstätter, I.; Hyslop, P.A.; Jackson, J.H.; Cochrane, C.G. Oxidant-induced DNA damage of target cells. J. Clin. Investig. 1988, 82, 1040–1050. [Google Scholar] [CrossRef] [Green Version]
- Hussain, S.P.; Hofseth, L.J.; Harris, C.C. Radical causes of cancer. Nat. Rev. Cancer 2003, 3, 276–285. [Google Scholar] [CrossRef]
- Klaunig, J.E.; Xu, Y.; Isenberg, J.S.; Bachowski, S.; Kolaja, K.L.; Jiang, J.; Stevenson, D.E.; Walborg, E.F., Jr. The role of oxidative stress in chemical carcinogenesis. Environ. Health Perspect. 1998, 106, 289–295. [Google Scholar] [CrossRef]
- Rendra, E.; Riabov, V.; Mossel, D.M.; Sevastyanova, T.; Harmsen, M.C.; Kzhyshkowska, J. Reactive oxygen species (ROS) in macrophage activation and function in diabetes. Immunobiology 2018, 224, 242–253. [Google Scholar] [CrossRef]
- Kattoor, A.J.; Pothineni, N.V.K.; Palagiri, D.; Mehta, J.L. Oxidative Stress in Atherosclerosis. Curr. Atheroscler. Rep. 2017, 19, 42. [Google Scholar] [CrossRef]
- Hussain, T.; Tan, B.; Yin, Y.; Blachier, F.; Tossou, M.C.; Rahu, N. Oxidative Stress and Inflammation: What Polyphenols Can Do for Us? Oxid. Med. Cell. Longev. 2016, 2016, 7432797. [Google Scholar] [CrossRef] [Green Version]
- Klaunig, J.E. Oxidative Stress and Cancer. Curr. Pharm. Des. 2019, 24, 4771–4778. [Google Scholar] [CrossRef] [PubMed]
- Sosa, V.; Moliné, T.; Somoza, R.; Paciucci, R.; Kondoh, H.; Lleonart, M.E. Oxidative stress and cancer: An overview. Ageing Res. Rev. 2013, 12, 376–390. [Google Scholar] [CrossRef] [PubMed]
- Trush, M.A.; Kensler, T.W. An overview of the relationship between oxidative stress and chemical carcinogenesis. Free Radic. Biol. Med. 1991, 10, 201–209. [Google Scholar] [CrossRef]
- Cerutti, P.A. Prooxidant States and Tumor Promotion. Science 1985, 227, 375–381. [Google Scholar] [CrossRef]
- Jackson, A.L.; Loeb, L.A. The contribution of endogenous sources of DNA damage to the multiple mutations in cancer. Mutat. Res. 2001, 477, 7–21. [Google Scholar] [CrossRef]
- Peter, S. Reactive oxygen species in tumor progression. Front. Biosci. 2005, 10, 1881–1896. [Google Scholar] [CrossRef] [Green Version]
- Ames, B.N.; Gold, L.S. Animal cancer tests and cancer prevention. J. Natl. Cancer Inst. Monogr. 1992, 12, 125–132. [Google Scholar]
- Guyton, K.Z.; Kensler, T.W. Oxidative mechanisms in carcinogenesis. Br. Med Bull. 1993, 49, 523–544. [Google Scholar] [CrossRef]
- Coussens, L.M.; Werb, Z. Inflammation and cancer. Nature 2002, 420, 860–867. [Google Scholar] [CrossRef]
- Valko, M.; Rhodes, C.J.; Moncol, J.; Izakovic, M.; Mazur, M. Free radicals, metals and antioxidants in oxidative stress-induced cancer. Chem. Biol. Interact. 2006, 160, 1–40. [Google Scholar] [CrossRef]
- Gupta, R.K.; Patel, A.K.; Shah, N.; Choudhary, A.K.; Jha, U.K.; Yadav, U.C.; Gupta, P.K.; Pakuwal, U. Oxidative Stress and Antioxidants in Disease and Cancer: A Review. Asian Pac. J. Cancer Prev. 2014, 15, 4405–4409. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pacher, P.; Beckman, J.S.; Liaudet, L. Nitric Oxide and Peroxynitrite in Health and Disease. Physiol. Rev. 2007, 87, 315–424. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Halliwell, B. Biochemistry of oxidative stress. Biochem. Soc. Trans. 2007, 35, 1147–1150. [Google Scholar] [CrossRef] [PubMed]
- Mackness, M.I.; Arrol, S.; Durrington, P.N. Paraoxonase prevents accumulation of lipoperoxides in low-density lipoprotein. FEBS Lett. 1991, 286, 152–154. [Google Scholar] [CrossRef] [Green Version]
- Deakin, S.P.; Bioletto, S.; Bochaton-Piallat, M.-L.; James, R.W. HDL-associated paraoxonase-1 can redistribute to cell membranes and influence sensitivity to oxidative stress. Free Radic. Biol. Med. 2011, 50, 102–109. [Google Scholar] [CrossRef] [Green Version]
- Aviram, M.; Rosenblat, M. Paraoxonases 1, 2, and 3, oxidative stress, and macrophage foam cell formation during atherosclerosis development. Free Radic. Biol. Med. 2004, 37, 1304–1316. [Google Scholar] [CrossRef]
- Camps, J.; Castañé, H.; Rodríguez-Tomàs, E.; Baiges-Gaya, G.; Hernández-Aguilera, A.; Arenas, M.; Iftimie, S.; Joven, J. On the Role of Paraoxonase-1 and Chemokine Ligand 2 (C-C motif) in Metabolic Alterations Linked to Inflammation and Disease. A 2021 Update. Biomolecules 2021, 11, 971. [Google Scholar] [CrossRef]
- Mackness, M.I.; Arrol, S.; Abbott, C.; Durrington, P.N. Protection of Low-Density Lipoprotein against Oxidative Modi-fication by High-Density Lipoprotein Associated Paraoxonase. Atherosclerosis 1993, 104, 12–21. [Google Scholar] [CrossRef]
- Ponce-Ruiz, N.; Murillo-González, F.E.; Rojas-García, A.E.; Mackness, M.; Bernal-Hernández, Y.Y.; Barrón-Vivanco, B.S.; González-Arias, C.A.; Medina-Díaz, I.M. Transcriptional regulation of human Paraoxonase 1 by nuclear receptors. Chem. Interact. 2017, 268, 77–84. [Google Scholar] [CrossRef]
- Shih, D.M.; Gu, L.; Xia, Y.-R.; Navab, M.; Li, W.-F.; Hama, S.; Castellani, L.W.; Furlong, C.E.; Costa, L.G.; Fogelman, A.M.; et al. Mice lacking serum paraoxonase are susceptible to organophosphate toxicity and atherosclerosis. Nature 1998, 394, 284–287. [Google Scholar] [CrossRef]
- Antognelli, C.; Del Buono, C.; Ludovini, V.; Gori, S.; Talesa, V.N.; Crinò, L.; Barberini, F.; Rulli, A. CYP17, GSTP1, PON1 and GLO1gene polymorphisms as risk factors for breast cancer: An Italian case-control study. BMC Cancer 2009, 9, 115. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Karaman, E.; Uzun, H.; Papila, I.; Balci, H.; Ozdilek, A.; Genc, H.; Yanardag, H.; Papila, C. Serum Paraoxonase Activity and Oxidative DNA Damage in Patients with Laryngeal Squamous Cell Carcinoma. J. Craniofac. Surg. 2010, 21, 1745–1749. [Google Scholar] [CrossRef] [PubMed]
- Ivanišević, J.; Kotur-Stevuljević, J.; Stefanović, A.; Miljković, M.; Jelić-Ivanović, Z.; Pejović, B.; Peco-Antić, A. Association of paraoxonase 1 and oxidative stress with acute kidney injury in premature asphyxiated neonates. Chem. Interact. 2017, 272, 47–52. [Google Scholar] [CrossRef] [PubMed]
- Foley, J.F. Focus Issue: Inflammatory mechanisms. Sci. Signal. 2015, 8, eg2. [Google Scholar] [CrossRef]
- Kuprash, D.V.; Nedospasov, S.A. Molecular and cellular mechanisms of inflammation. Biochemistry 2016, 81, 1237–1239. [Google Scholar] [CrossRef]
- Baizabal-Aguirre, V.M.; Rosales, C.; Lopez-Macias, C.I.R.; Gomez, M.I. Control and Resolution Mechanisms of the Inflammatory Response. Mediat. Inflamm. 2014, 2014, 387567. [Google Scholar] [CrossRef]
- Strowig, T.; Henao-Mejia, J.; Elinav, E.; Flavell, R. Inflammasomes in health and disease. Nature 2012, 481, 278–286. [Google Scholar] [CrossRef]
- Gallo, J.; Raska, M.; Kriegova, E.; Goodman, S.B. Inflammation and its resolution and the musculoskeletal system. J. Orthop. Transl. 2017, 10, 52–67. [Google Scholar] [CrossRef]
- Albrengues, J.; Shields, M.A.; Ng, D.; Park, C.G.; Ambrico, A.; Poindexter, M.E.; Upadhyay, P.; Uyeminami, D.L.; Pommier, A.; Küttner, V.; et al. Neutrophil extracellular traps produced during inflammation awaken dormant cancer cells in mice. Science 2018, 361, 4227. [Google Scholar] [CrossRef] [Green Version]
- Min, J.; Yang, D.; Kim, M.; Haam, K.; Yoo, A.; Choi, J.-H.; Schraml, B.; Kim, Y.S.; Kim, D.; Kang, S.-J. Inflammation induces two types of inflammatory dendritic cells in inflamed lymph nodes. Exp. Mol. Med. 2018, 50, e458. [Google Scholar] [CrossRef] [Green Version]
- Korn, T.; Oukka, M.; Kuchroo, V.; Bettelli, E. Th17 cells: Effector T cells with inflammatory properties. Semin. Immunol. 2007, 19, 362–371. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kolaczkowska, E.; Kubes, P. Neutrophil recruitment and function in health and inflammation. Nat. Rev. Immunol. 2013, 13, 159–175. [Google Scholar] [CrossRef] [PubMed]
- Nadeem, A.; Ahmad, S.F.; Attia, S.M.; Al-Ayadhi, L.Y.; Bakheet, S.A.; Al-Harbi, N.O. Oxidative and inflammatory mediators are upregulated in neutrophils of autistic children: Role of IL-17A receptor signaling. Prog. Neuro-Psychopharmacol. Biol. Psychiatry 2018, 90, 204–211. [Google Scholar] [CrossRef]
- Marichal, T.; Boyman, O.; Reber, L.L. Editorial: Role of Neutrophils in Inflammatory Diseases. Front. Immunol. 2020, 11, 627939. [Google Scholar] [CrossRef] [PubMed]
- Chiang, C.-C.; Cheng, W.-J.; Korinek, M.; Lin, C.-Y.; Hwang, T.-L. Neutrophils in Psoriasis. Front. Immunol. 2019, 10, 2376. [Google Scholar] [CrossRef]
- Zhang, D.; Chen, G.; Manwani, D.; Mortha, A.; Xu, C.; Faith, J.J.; Burk, R.D.; Kunisaki, Y.; Jang, J.-E.; Scheiermann, C.; et al. Neutrophil ageing is regulated by the microbiome. Nature 2015, 525, 528–532. [Google Scholar] [CrossRef] [PubMed]
- Vogel, D.Y.; Glim, J.E.; Stavenuiter, A.W.; Breur, M.; Heijnen, P.; Amor, S.; Dijkstra, C.D.; Beelen, R.H. Human macrophage polarization in vitro: Maturation and activation methods compared. Immunobiology 2014, 219, 695–703. [Google Scholar] [CrossRef]
- Kigerl, K.A.; Gensel, J.C.; Ankeny, D.P.; Alexander, J.K.; Donnelly, D.J.; Popovich, P.G. Identification of Two Distinct Macrophage Subsets with Divergent Effects Causing either Neurotoxicity or Regeneration in the Injured Mouse Spinal Cord. J. Neurosci. 2009, 29, 13435–13444. [Google Scholar] [CrossRef] [Green Version]
- Zhu, G.; Cheng, Z.; Lin, C.; Hoffman, R.M.; Huang, Y.; Singh, S.R.; Zheng, W.; Yang, S.; Ye, J. MyD88 Regulates LPS-induced NF-ĸB/MAPK Cytokines and Promotes Inflammation and Malignancy in Colorectal Cancer Cells. Cancer Genom. Proteom. 2019, 16, 409–419. [Google Scholar] [CrossRef]
- Tang, J.; Diao, P.; Shu, X.; Li, L.; Xiong, L. Quercetin and Quercitrin Attenuates the Inflammatory Response and Oxidative Stress in LPS-Induced RAW264.7 Cells: In Vitro Assessment and a Theoretical Model. BioMed Res. Int. 2019, 2019, 7039802. [Google Scholar] [CrossRef] [Green Version]
- Nathan, C. Points of control in inflammation. Nature 2002, 420, 846–852. [Google Scholar] [CrossRef] [PubMed]
- Conti, P.; Carinci, F.; Caraffa, A.; Ronconi, G.; Lessiani, G.; Theoharides, T.C. Link between mast cells and bacteria: Antimicrobial defense, function and regulation by cytokines. Med. Hypotheses 2017, 106, 10–14. [Google Scholar] [CrossRef] [PubMed]
- Wiegman, C.H.; Michaeloudes, C.; Haji, G.; Narang, P.; Clarke, C.J.; Russell, K.E.; Bao, W.; Pavlidis, S.; Barnes, P.J.; Kanerva, J.; et al. Oxidative stress–induced mitochondrial dysfunction drives inflammation and airway smooth muscle remodeling in patients with chronic obstructive pulmonary disease. J. Allergy Clin. Immunol. 2015, 136, 769–780. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chung, C.; Seo, W.; Silwal, P.; Jo, E.-K. Crosstalks between inflammasome and autophagy in cancer. J. Hematol. Oncol. 2020, 13, 100. [Google Scholar] [CrossRef]
- Poli, G.; Fabi, C.; Bellet, M.M.; Costantini, C.; Nunziangeli, L.; Romani, L.; Brancorsini, S. Epigenetic Mechanisms of Inflammasome Regulation. Int. J. Mol. Sci. 2020, 21, 5758. [Google Scholar] [CrossRef]
- Dai, Y.; Zhang, J.; Xiang, J.; Li, Y.; Wu, D.; Xu, J. Calcitriol inhibits ROS-NLRP3-IL-1β signaling axis via activation of Nrf2-antioxidant signaling in hyperosmotic stress stimulated human corneal epithelial cells. Redox Biol. 2018, 21, 101093. [Google Scholar] [CrossRef]
- Khandia, R.; Munjal, A. Interplay between inflammation and cancer. Adv. Protein Chem. Struct. Biol. 2019, 119, 199–245. [Google Scholar] [CrossRef]
- Jia, Z.; Babu, P.V.A.; Chen, W.; Sun, X. Natural Products Targeting on Oxidative Stress and Inflammation: Mechanisms, Therapies, and Safety Assessment. Oxidative Med. Cell. Longev. 2018, 2018, 6576093. [Google Scholar] [CrossRef] [Green Version]
- Bronner, D.N.; Abuaita, B.H.; Chen, X.; Fitzgerald, K.; Nuñez, G.; He, Y.; Yin, X.-M.; O’Riordan, M.X. Endoplasmic Reticulum Stress Activates the Inflammasome via NLRP3- and Caspase-2-Driven Mitochondrial Damage. Immunity 2015, 43, 451–462. [Google Scholar] [CrossRef] [Green Version]
- Keestra-Gounder, A.M.; Byndloss, M.; Seyffert, N.; Young, B.M.; Chávez-Arroyo, A.; Tsai, A.Y.; Cevallos, S.A.; Winter, M.G.; Pham, O.H.; Tiffany, C.R.; et al. NOD1 and NOD2 signalling links ER stress with inflammation. Nature 2016, 532, 394–397. [Google Scholar] [CrossRef] [Green Version]
- Lloberas, J.; Muñoz, J.P.; Hernández-Álvarez, M.I.; Cardona, P.-J.; Zorzano, A.; Celada, A. Macrophage mitochondrial MFN2 (mitofusin 2) links immune stress and immune response through reactive oxygen species (ROS) production. Autophagy 2020, 16, 2307–2309. [Google Scholar] [CrossRef] [PubMed]
- Brucklacher-Waldert, V.; Ferreira, C.; Stebegg, M.; Fesneau, O.; Innocentin, S.; Marie, J.; Veldhoen, M. Cellular Stress in the Context of an Inflammatory Environment Supports TGF-β-Independent T Helper-17 Differentiation. Cell Rep. 2017, 19, 2357–2370. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sadeghi, A.; Shabani, M.; Alizadeh, S.; Meshkani, R. Interplay between oxidative stress and autophagy function and its role in inflammatory cytokine expression induced by palmitate in skeletal muscle cells. Cytokine 2019, 125, 154835. [Google Scholar] [CrossRef] [PubMed]
- Carstensen, L.S.; Lie-Andersen, O.; Obers, A.; Crowther, M.D.; Svane, I.M.; Hansen, M. Long-Term Exposure to Inflammation Induces Differential Cytokine Patterns and Apoptosis in Dendritic Cells. Front. Immunol. 2019, 10, 2702. [Google Scholar] [CrossRef] [Green Version]
- Nie, F.; Ding, F.; Chen, B.; Huang, S.; Liu, Q.; Xu, C. Dendritic cells aggregate inflammation in experimental osteoarthritis through a toll-like receptor (TLR)-dependent machinery response to challenges. Life Sci. 2019, 238, 116920. [Google Scholar] [CrossRef]
- Raphael, I.; Nalawade, S.; Eagar, T.N.; Forsthuber, T.G. T cell subsets and their signature cytokines in autoimmune and inflammatory diseases. Cytokine 2014, 74, 5–17. [Google Scholar] [CrossRef] [Green Version]
- Basu, R.; Hatton, R.; Weaver, C. The Th17 family: Flexibility follows function. Immunol. Rev. 2013, 252, 89–103. [Google Scholar] [CrossRef] [Green Version]
- Ramesh, R.; Kozhaya, L.; McKevitt, K.; Djuretic, I.M.; Carlson, T.J.; Quintero, M.A.; McCauley, J.L.; Abreu, M.T.; Unutmaz, D.; Sundrud, M.S. Pro-inflammatory human Th17 cells selectively express P-glycoprotein and are refractory to glucocorticoids. J. Exp. Med. 2014, 211, 89–104. [Google Scholar] [CrossRef] [Green Version]
- Merino, K.M.; Jazwinski, S.M.; Rout, N. Th17-type immunity and inflammation of aging. Aging 2021, 13, 13378–13379. [Google Scholar] [CrossRef]
- Lee, B.; Gibbs, P. Inflammation, Biomarkers and Immuno-Oncology Pathways in Pancreatic Cancer. J. Pers. Med. 2019, 9, 20. [Google Scholar] [CrossRef] [Green Version]
- Liao, C.; Rosas, M.; Davies, L.C.; Giles, P.J.; Tyrrell, V.J.; O’Donnell, V.B.; Topley, N.; Humphreys, I.R.; Fraser, D.J.; Jones, S.A.; et al. IL-10 differentially controls the infiltration of inflammatory macrophages and antigen-presenting cells during inflammation. Eur. J. Immunol. 2016, 46, 2222–2232. [Google Scholar] [CrossRef]
- Ma, N.; Zhang, Y.; Liu, Q.; Wang, Z.; Liu, X.; Zhu, G.; Yu, D.; Han, G.; Chen, G.; Hou, C.; et al. B cell activating factor (BAFF) selects IL-10(−) B cells over IL-10(+) B cells during inflammatory responses. Mol. Immunol. 2017, 85, 18–26. [Google Scholar] [CrossRef]
- Hetta, H.F.; Mwafey, I.M.; Batiha, G.E.-S.; AlOmar, S.Y.; Mohamed, N.A.; Ibrahim, M.A.; Elkady, A.; Meshaal, A.K.; Alrefai, H.; Khodeer, D.M.; et al. CD19+ CD24hi CD38hi Regulatory B Cells and Memory B Cells in Periodontitis: Association with Pro-Inflammatory and Anti-Inflammatory Cytokines. Vaccines 2020, 8, 340. [Google Scholar] [CrossRef]
- Yang, W.; Tao, Y.; Wu, Y.; Zhao, X.; Ye, W.; Zhao, D.; Fu, L.; Tian, C.; Yang, J.; He, F.; et al. Neutrophils promote the development of reparative macrophages mediated by ROS to orchestrate liver repair. Nat. Commun. 2019, 10, 1076. [Google Scholar] [CrossRef] [Green Version]
- Medeiros, N.I.; Mattos, R.T.; Menezes, C.A.; Fares, R.C.G.; Talvani, A.; Dutra, W.O.; Rios-Santos, F.; Correa-Oliveira, R.; Gomes, J.A.S. IL-10 and TGF-β unbalanced levels in neutrophils contribute to increase inflammatory cytokine expression in childhood obesity. Eur. J. Nutr. 2017, 57, 2421–2430. [Google Scholar] [CrossRef]
- El-Kenawi, A.; Ruffell, B. Inflammation, ROS, and Mutagenesis. Cancer Cell 2017, 32, 727–729. [Google Scholar] [CrossRef] [Green Version]
- Jeong, Y.J.; Oh, H.K.; Park, S.H.; Bong, J.G. Association between inflammation and cancer stem cell phenotype in breast cancer. Oncol. Lett. 2017, 15, 2380–2386. [Google Scholar] [CrossRef]
- Himbert, C.; Ose, J.; Lin, T.; Warby, C.A.; Gigic, B.; Steindorf, K.; Schrotz-King, P.; Abbenhardt-Martin, C.; Zielske, L.; Boehm, J.; et al. Inflammation- and angiogenesis-related biomarkers are correlated with cancer-related fatigue in colorectal cancer patients: Results from the ColoCare Study. Eur. J. Cancer Care 2019, 28, e13055. [Google Scholar] [CrossRef]
- He, L.; Jhong, J.-H.; Chen, Q.; Huang, K.-Y.; Strittmatter, K.; Kreuzer, J.; DeRan, M.; Wu, X.; Lee, T.-Y.; Slavov, N.; et al. Global characterization of macrophage polarization mechanisms and identification of M2-type polarization inhibitors. Cell Rep. 2021, 37, 109955. [Google Scholar] [CrossRef]
- Bilska, M.; Pawłowska, A.; Zakrzewska, E.; Chudzik, A.; Suszczyk, D.; Gogacz, M.; Wertel, I. Th17 Cells and IL-17 As Novel Immune Targets in Ovarian Cancer Therapy. J. Oncol. 2020, 2020, 8797683. [Google Scholar] [CrossRef] [Green Version]
- Salazar, Y.; Zheng, X.; Brunn, D.; Raifer, H.; Picard, F.S.R.; Zhang, Y.; Winter, H.; Guenther, S.; Weigert, A.; Weigmann, B.; et al. Microenvironmental Th9 and Th17 lymphocytes induce metastatic spreading in lung cancer. J. Clin. Investig. 2020, 130, 3560–3575. [Google Scholar] [CrossRef] [Green Version]
- Bryant, G.; Wang, L.; Mulholland, D.J. Overcoming Oncogenic Mediated Tumor Immunity in Prostate Cancer. Int. J. Mol. Sci. 2017, 18, 1542. [Google Scholar] [CrossRef]
- Solís-Martínez, R.; Marentes, M.E.C.; Hernandez-Flores, G.; Ortiz-Lazareno, P.; Mandujano-Álvarez, G.; Cruz-Gálvez, C.; Sierra-Díaz, E.; Rodríguez-Padilla, C.; Jave-Suárez, L.; Aguilar-Lemarroy, A.; et al. Regulation of immunophenotype modulation of monocytes-macrophages from M1 into M2 by prostate cancer cell-culture supernatant via transcription factor STAT3. Immunol. Lett. 2018, 196, 140–148. [Google Scholar] [CrossRef]
- Song, W.; Mazzieri, R.; Yang, T.; Gobe, G.C. Translational Significance for Tumor Metastasis of Tumor-Associated Macrophages and Epithelial–Mesenchymal Transition. Front. Immunol. 2017, 8, 1106. [Google Scholar] [CrossRef] [Green Version]
- Escamilla-Tilch, M.; Filio-Rodríguez, G.; García-Rocha, R.; Mancilla-Herrera, I.; Mitchison, N.A.; Pacheco, J.A.R.; Sánchez-García, F.J.; Sandoval-Borrego, D.; Vázquez-Sánchez, E.A. The interplay between pathogen-associated and danger-associated molecular patterns: An inflammatory code in cancer? Immunol. Cell Biol. 2013, 91, 601–610. [Google Scholar] [CrossRef]
- Hussain, S.P.; Harris, C.C. Inflammation and cancer: An ancient link with novel potentials. Int. J. Cancer 2007, 121, 2373–2380. [Google Scholar] [CrossRef]
- Fan, Y.; Mao, R.; Yang, J. NF-κB and STAT3 signaling pathways collaboratively link inflammation to cancer. Protein Cell 2013, 4, 176–185. [Google Scholar] [CrossRef] [Green Version]
- Saha, S.; Buttari, B.; Panieri, E.; Profumo, E.; Saso, L. An Overview of Nrf2 Signaling Pathway and Its Role in Inflammation. Molecules 2020, 25, 5474. [Google Scholar] [CrossRef]
- Anant, S.; Sureban, S.M. Cyclooxygenase-2 Gene Expression. In Apoptosis, Cell Signaling, and Human Diseases; Srivastava, R., Ed.; Humana Press: Totowa, NJ, USA, 2006. [Google Scholar] [CrossRef]
- Cinelli, M.A.; Do, H.T.; Miley, G.P.; Silverman, R.B. Inducible nitric oxide synthase: Regulation, structure, and inhibition. Med. Res. Rev. 2019, 40, 158–189. [Google Scholar] [CrossRef]
- Yu, H.; Lin, L.; Zhang, Z.; Zhang, H.; Hu, H. Targeting NF-κB pathway for the therapy of diseases: Mechanism and clinical study. Signal Transduct. Target. Ther. 2020, 5, 209. [Google Scholar] [CrossRef]
- Hacker, H.; Karin, M. Regulation and Function of IKK and IKK-Related Kinases. Sci. STKE 2006, 357, re13. [Google Scholar] [CrossRef]
- Hayden, M.S.; Ghosh, S. Shared Principles in NF-κB Signaling. Cell 2008, 132, 344–362. [Google Scholar] [CrossRef] [Green Version]
- Wertz, I.E.; Dixit, V.M. Signaling to NF- B: Regulation by Ubiquitination. Cold Spring Harb. Perspect. Biol. 2009, 2, a003350. [Google Scholar] [CrossRef] [Green Version]
- Chen, L.-F.; Greene, W.C. Shaping the nuclear action of NF-κB. Nat. Rev. Mol. Cell Biol. 2004, 5, 392–401. [Google Scholar] [CrossRef]
- Hoffmann, A.; Natoli, G.; Ghosh, G. Transcriptional regulation via the NF-κB signaling module. Oncogene 2006, 25, 6706–6716. [Google Scholar] [CrossRef] [Green Version]
- Vallabhapurapu, S.; Karin, M. Regulation and Function of NF-κB Transcription Factors in the Immune System. Annu. Rev. Immunol. 2009, 27, 693–733. [Google Scholar] [CrossRef]
- Kawai, T.; Akira, S. The role of pattern-recognition receptors in innate immunity: Update on Toll-like receptors. Nat. Immunol. 2010, 11, 373–384. [Google Scholar] [CrossRef]
- Kumar, H.; Kawai, T.; Akira, S. Toll-like receptors and innate immunity. Biochem. Biophys. Res. Commun. 2009, 388, 621–625. [Google Scholar] [CrossRef]
- Takeuchi, O.; Akira, S. Pattern Recognition Receptors and Inflammation. Cell 2010, 140, 805–820. [Google Scholar] [CrossRef] [Green Version]
- Gong, T.; Liu, L.; Jiang, W.; Zhou, R. DAMP-sensing receptors in sterile inflammation and inflammatory diseases. Nat. Rev. Immunol. 2019, 20, 95–112. [Google Scholar] [CrossRef]
- Bonizzi, G.; Bebien, M.; Otero, D.C.; Johnson-Vroom, K.E.; Cao, Y.; Vu, D.; Jegga, A.; Aronow, B.J.; Ghosh, G.; Rickert, R.C.; et al. Activation of IKKα target genes depends on recognition of specific κB binding sites by RelB:p52 dimers. EMBO J. 2004, 23, 4202–4210. [Google Scholar] [CrossRef] [Green Version]
- Barnes, P.; Adcock, I. Transcription factors and asthma. Eur. Respir. J. 1998, 12, 221–234. [Google Scholar] [CrossRef] [Green Version]
- Chen, T.; Zhang, X.; Zhu, G.; Liu, H.; Chen, J.; Wang, Y.; He, X. Quercetin inhibits TNF-α induced HUVECs apoptosis and inflammation via downregulating NF-kB and AP-1 signaling pathway in vitro. Medicine 2020, 99, e22241. [Google Scholar] [CrossRef] [PubMed]
- Palazon, A.; Goldrath, A.W.; Nizet, V.; Johnson, R.S. HIF Transcription Factors, Inflammation, and Immunity. Immunity 2014, 41, 518–528. [Google Scholar] [CrossRef] [Green Version]
- Watts, E.R.; Walmsley, S.R. Inflammation and Hypoxia: HIF and PHD Isoform Selectivity. Trends Mol. Med. 2019, 25, 33–46. [Google Scholar] [CrossRef] [Green Version]
- Meijide, J.A.G.; Gómez-Reino Carnota, J.J. Fisiopatología de la ciclooxigenasa-1 y ciclooxigenasa-2. Rev. Esp. Reumatol. 2000, 27, 33–35. [Google Scholar]
- Simon, L.S. Role and regulation of cyclooxygenase-2 during inflammation. Am. J. Med. 1999, 106, 37S–42S. [Google Scholar] [CrossRef]
- Hla, T.; Neilson, K. Human cyclooxygenase-2 cDNA. Proc. Natl. Acad. Sci. USA 1992, 89, 7384–7388. [Google Scholar] [CrossRef] [Green Version]
- Martens, C.R.; Bansal, S.S.; Accornero, F. Cardiovascular inflammation: RNA takes the lead. J. Mol. Cell. Cardiol. 2019, 129, 247–256. [Google Scholar] [CrossRef]
- Ferrucci, L.; Fabbri, E. Inflammageing: Chronic inflammation in ageing, cardiovascular disease, and frailty. Nat. Rev. Cardiol. 2018, 15, 505–522. [Google Scholar] [CrossRef]
- Laaksonen, D.E.; Niskanen, L.; Punnonen, K.; Nyyssönen, K.; Tuomainen, T.-P.; Salonen, R.; Rauramaa, R.; Salonen, J.T. Sex hormones, inflammation and the metabolic syndrome: A population-based study. Eur. J. Endocrinol. 2003, 149, 601–608. [Google Scholar] [CrossRef] [PubMed]
- Festa, A.; D’Agostino, J.R.; Howard, G.; Mykkanen, L.; Tracy, R.P.; Haffner, S.M. Chronic Subclinical Inflammation as Part of the Insulin Resistance Syndrome: The Insulin Resistance Atherosclerosis Study (IRAS). Circulation 2000, 102, 42–47. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rodríguez-Hernández, H.; Simental-Mendía, L.E.; Rodríguez-Ramírez, G.; Reyes-Romero, M.A. Obesity and Inflammation: Epidemiology, Risk Factors, and Markers of Inflammation. Int. J. Endocrinol. 2013, 2013, 678159. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Idris-Khodja, N.; Mian, M.O.R.; Paradis, P.; Schiffrin, E.L. Dual opposing roles of adaptive immunity in hypertension. Eur. Heart J. 2014, 35, 1238–1244. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Solak, Y.; Afsar, B.; Vaziri, N.D.; Aslan, G.; Yalcin, C.E.; Covic, A.; Kanbay, M. Hypertension as an autoimmune and inflammatory disease. Hypertens. Res. 2016, 39, 567–573. [Google Scholar] [CrossRef]
- Annagür, A.; Örs, R.; Altunhan, H.; Kurban, S.; Ertuğrul, S.; Konak, M.; Uygun, S.S.; Pekcan, S.; Erbay, E.; Mehmetoğlu, I. Total antioxidant and total oxidant states, and serum paraoxonase-1 in neonatal sepsis. Pediatr. Int. 2014, 57, 608–613. [Google Scholar] [CrossRef] [PubMed]
- Valente, A.J.; Irimpen, A.M.; Siebenlist, U.; Chandrasekar, B. OxLDL induces endothelial dysfunction and death via TRAF3IP2: Inhibition by HDL3 and AMPK activators. Free Radic. Biol. Med. 2014, 70, 117–128. [Google Scholar] [CrossRef] [Green Version]
- Valacchi, G.; Virgili, F.; Cervellati, C.; Pecorelli, A. OxInflammation: From Subclinical Condition to Pathological Biomarker. Front. Physiol. 2018, 9, 858. [Google Scholar] [CrossRef] [Green Version]
- Madamanchi, N.R.; Vendrov, A.; Runge, M.S. Oxidative Stress and Vascular Disease. Arteroscler. Thromb. Vasc. Biol. 2005, 25, 29–38. [Google Scholar] [CrossRef] [Green Version]
- Cervellati, C.; Vigna, G.B.; Trentini, A.; Sanz, J.M.; Zimetti, F.; Dalla Nora, E.; Morieri, M.L.; Zuliani, G.; Passaro, A. Paraoxonase-1 activities in individuals with different HDL circulating levels: Implication in reverse cholesterol transport and early vascular damage. Atherosclerosis 2019, 285, 64–70. [Google Scholar] [CrossRef]
- Mackness, B.; Hine, D.; Liu, Y.; Mastorikou, M.; Mackness, M. Paraoxonase-1 inhibits oxidised LDL-induced MCP-1 production by endothelial cells. Biochem. Biophys. Res. Commun. 2004, 318, 680–683. [Google Scholar] [CrossRef] [PubMed]
- Aharoni, S.; Aviram, M.; Fuhrman, B. Paraoxonase 1 (PON1) reduces macrophage inflammatory responses. Atherosclerosis 2013, 228, 353–361. [Google Scholar] [CrossRef] [PubMed]
- Marek, G.; Ściskalska, M.; Grzebieniak, Z.; Milnerowicz, H. Decreases in Paraoxonase-1 Activities Promote a Pro-inflammatory Effect of Lipids Peroxidation Products in Non-smoking and Smoking Patients with Acute Pancreatitis. Int. J. Med. Sci. 2018, 15, 1619–1630. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aldonza, M.B.D.; Son, Y.S.; Sung, H.-J.; Ahn, J.M.; Choi, Y.-J.; Kim, Y.-I.; Cho, S.; Cho, J.-Y. Paraoxonase-1 (PON1) induces metastatic potential and apoptosis escape via its antioxidative function in lung cancer cells. Oncotarget 2017, 8, 42817–42835. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kumon, Y.; Nakauchi, Y.; Suehiro, T.; Shiinoki, T.; Tanimoto, N.; Inoue, M.; Nakamura, T.; Hashimoto, K.; Sipe, J.D. Proinflammatory Cytokines but Not Acute Phase Serum Amyloid A Or C-Reactive Protein, Downregulate Paraoxonase 1 (Pon1) Expression by Hepg2 Cells. Amyloid 2002, 9, 160–164. [Google Scholar] [CrossRef]
- Kumon, Y.; Suehiro, T.; Ikeda, Y.; Hashimoto, K. Human paraoxonase-1 gene expression by HepG2 cells is downregulated by interleukin-1β and tumor necrosis factor-α, but is upregulated by interleukin-6. Life Sci. 2003, 73, 2807–2815. [Google Scholar] [CrossRef]
- Han, C.Y.; Chiba, T.; Campbell, J.S.; Fausto, N.; Chaisson, M.; Orasanu, G.; Plutzky, J.; Chait, A. Reciprocal and Coordinate Regulation of Serum Amyloid A Versus Apolipoprotein A-I and Paraoxonase-1 by Inflammation in Murine Hepatocytes. Arter. Thromb. Vasc. Biol. 2006, 26, 1806–1813. [Google Scholar] [CrossRef] [Green Version]
- Gouédard, C.; Koum-Besson, N.; Barouki, R.; Morel, Y. Opposite regulation of the human paraoxonase-1 gene PON-1 by fenofibrate and statins. Mol. Pharmacol. 2003, 63, 945–956. [Google Scholar] [CrossRef]
- Cheng, C.-C.; Hsueh, C.-M.; Chen, C.-Y.; Chen, T.-H.; Hsu, S.-L. Interleukin-6 upregulates paraoxonase 1 gene expression via an AKT/NF-κB-dependent pathway. Biochem. Biophys. Res. Commun. 2013, 437, 55–61. [Google Scholar] [CrossRef]
- Kotani, K.; Yamada, T.; Gugliucci, A. Paired Measurements of Paraoxonase 1 and Serum Amyloid A as Useful Disease Markers. BioMed Res. Int. 2013, 2013, 481437. [Google Scholar] [CrossRef] [Green Version]
- Gugliucci, A.; Kotani, K.; Kimura, S. Paraoxonase 1 in Chronic Kidney Failure. J. Lipids 2012, 2012, 726048. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kerekes, G.; Szekanecz, Z.; Dér, H.; Sándor, Z.; Lakos, G.; Muszbek, L.; Csipö, I.; Sipka, S.; Seres, I.; Paragh, G.; et al. Endothelial dysfunction and atherosclerosis in rheumatoid arthritis: A multiparametric analysis using imaging techniques and laboratory markers of inflammation and autoimmunity. J. Rheumatol. 2008, 35, 398–406. [Google Scholar] [PubMed]
- Szczeklik, K.; Mach, T.; Cibor, D.; Owczarek, D.; Sapa, J.; Papież, M.; Pytko-Polończyk, J.; Krzyściak, W. Correlation of Paraoxonase-1 with the Severity of Crohn’s Disease. Molecules 2018, 23, 2603. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Durrington, P.N.; Mackness, B.; Mackness, M.I. Paraoxonase and Atherosclerosis. Arter. Thromb. Vasc. Biol. 2001, 21, 473–480. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mackness, M.; Mackness, B. Human paraoxonase-1 (PON1): Gene structure and expression, promiscuous activities and multiple physiological roles. Gene 2015, 567, 12–21. [Google Scholar] [CrossRef] [Green Version]
- Primo-Parmo, S.L.; Sorenson, R.C.; Teiber, J.; La Du, B.N. The Human Serum Paraoxonase/Arylesterase Gene (PON1) Is One Member of a Multigene Family. Genomics 1996, 33, 498–507. [Google Scholar] [CrossRef] [PubMed]
- Furlong, C.E.; Marsillach, J.; Jarvik, G.P.; Costa, L.G. Paraoxonases-1, -2 and -3: What are their functions? Chem-Biol. Interact. 2016, 259, 51–62. [Google Scholar] [CrossRef] [Green Version]
- Marsillach, J.; Camps, J.; Beltrán-Debón, R.; Rull, A.; Aragonès, G.; Maestre-Martínez, C.; Sabench, F.; Hernandez, L.; Castillo, D.D.; Joven, J.; et al. Immunohistochemical analysis of paraoxonases-1 and 3 in human atheromatous plaques. Eur. J. Clin. Investig. 2010, 41, 308–314. [Google Scholar] [CrossRef]
- Rodríguez-Sanabria, F.; Rull, A.; Beltrán-Debón, R.; Aragonès, G.; Camps, J.; Mackness, B.; Mackness, M.; Joven, J. Tissue distribution and expression of paraoxonases and chemokines in mouse: The ubiquitous and joint localisation suggest a systemic and coordinated role. Histochem. J. 2010, 41, 379–386. [Google Scholar] [CrossRef]
- Draganov, D.I.; Teiber, J.F.; Speelman, A.; Osawa, Y.; Sunahara, R.; La Du, B.N. Human paraoxonases (PON1, PON2, and PON3) are lactonases with overlapping and distinct substrate specificities. J. Lipid Res. 2005, 46, 1239–1247. [Google Scholar] [CrossRef] [Green Version]
- Androutsopoulos, V.; Kanavouras, K.; Tsatsakis, A. Role of paraoxonase 1 (PON1) in organophosphate metabolism: Implications in neurodegenerative diseases. Toxicol. Appl. Pharmacol. 2011, 256, 418–424. [Google Scholar] [CrossRef] [PubMed]
- Costa, L.G.; Giordano, G.; Cole, T.B.; Marsillach, J.; Furlong, C.E. Paraoxonase 1 (PON1) as a genetic determinant of susceptibility to organophosphate toxicity. Toxicology 2012, 307, 115–122. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mahrooz, A.; Mackness, M.; Bagheri, A.; Ghaffari-Cherati, M.; Masoumi, P. The epigenetic regulation of paraoxonase 1 (PON1) as an important enzyme in HDL function: The missing link between environmental and genetic regulation. Clin. Biochem. 2019, 73, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Camps, J.; Marsillach, J.; Joven, J. The paraoxonases: Role in human diseases and methodological difficulties in measurement. Crit. Rev. Clin. Lab. Sci. 2009, 46, 83–106. [Google Scholar] [CrossRef]
- Dardiotis, E.; Siokas, V.; Sokratous, M.; Tsouris, Z.; Michalopoulou, A.; Andravizou, A.; Dastamani, M.; Ralli, S.; Vinceti, M.; Tsatsakis, A.; et al. Genetic polymorphisms in amyotrophic lateral sclerosis: Evidence for implication in detoxification pathways of environmental toxicants. Environ. Int. 2018, 116, 122–135. [Google Scholar] [CrossRef] [Green Version]
- Murillo-González, F.; Ponce-Ruiz, N.; Rojas-García, A.; Rothenberg, S.; Bernal-Hernández, Y.; Cerda-Flores, R.; Mackness, M.; Barrón-Vivanco, B.; González-Arias, C.; Ponce-Gallegos, J.; et al. PON1 lactonase activity and its association with cardiovascular disease. Clin. Chim. Acta 2019, 500, 47–53. [Google Scholar] [CrossRef]
- Biggadike, K.; Angell, R.M.; Burgess, C.M.; Farrell, R.M.; Hancock, A.P.; Harker, A.J.; Irving, W.R.; Ioannou, C.; Procopiou, P.A.; Shaw, R.E.; et al. Selective Plasma Hydrolysis of Glucocorticoid γ-Lactones and Cyclic Carbonates by the Enzyme Paraoxonase: An Ideal Plasma Inactivation Mechanism. J. Med. Chem. 1999, 43, 19–21. [Google Scholar] [CrossRef]
- Teiber, J.F.; Draganov, D.I.; Du, B.N. Lactonase and lactonizing activities of human serum paraoxonase (PON1) and rabbit serum PON3. Biochem. Pharmacol. 2003, 66, 887–896. [Google Scholar] [CrossRef]
- Khersonsky, O.; Tawfik, D.S. Structure—Reactivity Studies of Serum Paraoxonase PON1 Suggest that Its Native Activity Is Lactonase. Biochemistry 2005, 44, 6371–6382. [Google Scholar] [CrossRef]
- Aviram, M.; Hardak, E.; Vaya, J.; Mahmood, S.; Milo, S.; Hoffman, A.; Billicke, S.; Draganov, D.; Rosenblat, M. Human Serum Paraoxonases (PON1) Q and R Selectively Decrease Lipid Peroxides in Human Coronary and Carotid Atherosclerotic Lesions: PON1 Esterase and Peroxidase-like Activities. Circulation 2000, 101, 2510–2517. [Google Scholar] [CrossRef] [Green Version]
- Sentí, M.; Tomás, M.; Fitó, M.; Weinbrenner, T.; Covas, M.-I.; Sala, J.; Masiá, R.; Marrugat, J. Antioxidant Paraoxonase 1 Activity in the Metabolic Syndrome. J. Clin. Endocrinol. Metab. 2003, 88, 5422–5426. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Camps, J.; García-Heredia, A.; Rull, A.; Alonso-Villaverde, C.; Aragonès, G.; Beltrán-Debón, R.; Rodríguez-Gallego, E.; Joven, J. PPARs in Regulation of Paraoxonases: Control of Oxidative Stress and Inflammation Pathways. PPAR Res. 2012, 2012, 616371. [Google Scholar] [CrossRef] [PubMed]
- Ozer, E.; Pezzulo, A.; Shih, D.M.; Chun, C.; Furlong, C.; Lusis, A.J.; Greenberg, E.; Zabner, J. Human and murine paraoxonase 1 are host modulators of Pseudomonas aeruginosa quorum-sensing. FEMS Microbiol. Lett. 2005, 253, 29–37. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tsatsakis, A.; Zafiropoulos, A.; Tzatzarakis, M.; Tzanakakis, G.; Kafatos, A. Relation of PON1 and CYP1A1 genetic polymorphisms to clinical findings in a cross-sectional study of a Greek rural population professionally exposed to pesticides. Toxicol. Lett. 2008, 186, 66–72. [Google Scholar] [CrossRef] [PubMed]
- Camps, J.; Pujol, I.; Ballester, F.; Joven, J.; Simó, J.M. Paraoxonases as Potential Antibiofilm Agents: Their Relationship with Quorum-Sensing Signals in Gram-Negative Bacteria. Antimicrob. Agents Chemother. 2011, 55, 1325–1331. [Google Scholar] [CrossRef] [Green Version]
- Elkiran, E.T.; Mar, N.; Aygen, B.; Gursu, F.; Karaoglu, A.; Koca, S. Serum paraoxonase and arylesterase activities in patients with lung cancer in a Turkish population. BMC Cancer 2007, 7, 48. [Google Scholar] [CrossRef] [Green Version]
- Faridvand, Y.; Oskuyi, A.E.; Khadem-Ansari, M.-H. Serum 8-isoprostane levels and paraoxonase 1 activity in patients with stage I multiple myeloma. Redox Rep. 2016, 21, 204–208. [Google Scholar] [CrossRef] [Green Version]
- Korkmaz, H.; Tabur, S.; Özkaya, M.; Aksoy, N.; Yildiz, H.; Akarsu, E. Paraoxonase and arylesterase activities in patients with papillary thyroid cancer. Scand. J. Clin. Lab. Investig. 2015, 75, 259–264. [Google Scholar] [CrossRef]
- Ng, C.J.; Shih, D.M.; Hama, S.Y.; Villa, N.; Navab, M.; Reddy, S.T. The paraoxonase gene family and atherosclerosis. Free Radic. Biol. Med. 2005, 38, 153–163. [Google Scholar] [CrossRef]
- Clendenning, J.B.; Humbert, R.; Green, E.D.; Wood, C.; Traver, D.; Furlong, C.E. Structural Organization of the HumanPON1Gene. Genomics 1996, 35, 586–589. [Google Scholar] [CrossRef]
- Deakin, S.; Leviev, I.; Guernier, S.; James, R.W. Simvastatin Modulates Expression of the PON1 Gene and Increases Serum Paraoxonase: A role for sterol regulatory element-binding protein-2. Arter. Thromb. Vasc. Biol. 2003, 23, 2083–2089. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mackness, B.; Durrington, P.N.; Mackness, M.I. Human Serum Paraoxonase. Gen. Pharmacol. Vasc. Syst. 1998, 31, 329–336. [Google Scholar] [CrossRef]
- Davidson, W.S.; Silva, R.G.D.; Chantepie, S.; Lagor, W.; Chapman, M.J.; Kontush, A. Proteomic Analysis of Defined HDL Subpopulations Reveals Particle-Specific Protein Clusters: Relevance to Antioxidative Function. Arter. Thromb. Vasc. Biol. 2009, 29, 870–876. [Google Scholar] [CrossRef] [PubMed]
- Kuo, C.L.; la Du, B.N. Calcium binding by human and rabbit serum paraoxonases. Structural stability and enzymatic activity. Drug Metab. Dispos. 1998, 26, 653–660. [Google Scholar] [PubMed]
- Harel, M.; Aharoni, A.; Gaidukov, L.; Brumshtein, B.; Khersonsky, O.; Meged, R.; Dvir, H.; Ravelli, R.; McCarthy, A.; Toker, L.; et al. Structure and evolution of the serum paraoxonase family of detoxifying and anti-atherosclerotic enzymes. Nat. Struct. Mol. Biol. 2004, 11, 412–419. [Google Scholar] [CrossRef] [PubMed]
- Leviev, I.; Negro, F.; James, R.W. Two Alleles of the Human Paraoxonase Gene Produce Different Amounts of mRNA: An explanation for differences in serum concentrations of paraoxonase associated with the (Leu-Met54) polymorphism. Arter. Thromb. Vasc. Biol. 1997, 17, 2935–2939. [Google Scholar] [CrossRef]
- Leviev, I.; James, R.W. Promoter Polymorphisms of Human Paraoxonase PON1 Gene and Serum Paraoxonase Activities and Concentrations. Arter. Thromb. Vasc. Biol. 2000, 20, 516–521. [Google Scholar] [CrossRef] [Green Version]
- Suehiro, T.; Nakamura, T.; Inoue, M.; Shiinoki, T.; Ikeda, Y.; Kumon, Y.; Shindo, M.; Tanaka, H.; Hashimoto, K. A polymorphism upstream from the human paraoxonase (PON1) gene and its association with PON1 expression. Atherosclerosis 2000, 150, 295–298. [Google Scholar] [CrossRef]
- Brophy, V.H.; Hastings, M.D.; Clendenning, J.B.; Richter, R.J.; Jarvik, G.P.; Furlong, C.E. Polymorphisms in the human paraoxonase (PON1) promoter. Pharmacogenetics 2001, 11, 77–84. [Google Scholar] [CrossRef]
- Osaki, F.; Ikeda, Y.; Suehiro, T.; Ota, K.; Tsuzura, S.; Arii, K.; Kumon, Y.; Hashimoto, K. Roles of Sp1 and protein kinase C in regulation of human serum paraoxonase 1 (PON1) gene transcription in HepG2 cells. Atherosclerosis 2004, 176, 279–287. [Google Scholar] [CrossRef]
- Draganov, D.I.; La Du, B.N. Pharmacogenetics of paraoxonases: A brief review. Naunyn-Schmiedebergs Arch. Exp. Pathol. Pharmakol. 2004, 369, 78–88. [Google Scholar] [CrossRef] [PubMed]
- Voetsch, B.; Benke, K.S.; Panhuysen, C.I.; Damasceno, B.P.; Loscalzo, J. The Combined Effect of Paraoxonase Promoter and Coding Region Polymorphisms on the Risk of Arterial Ischemic Stroke among Young Adults. Arch. Neurol. 2004, 61, 351–356. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Furlong, C.E.; Richter, R.J.; Chapline, C.; Crabb, J.W. Purification of rabbit and human serum paraoxonase. Biochemistry 1991, 30, 10133–10140. [Google Scholar] [CrossRef] [PubMed]
- Hassett, C.; Richter, R.J.; Humbert, R.; Chapline, C.; Crabb, J.W.; Omiecinski, C.J.; Furlong, C.E. Characterization of cDNA clones encoding rabbit and human serum paraoxonase: The mature protein retains its signal sequence. Biochemistry 1991, 30, 10141–10149. [Google Scholar] [CrossRef]
- Costa, L.G.; Cole, T.B.; Vitalone, A.; Furlong, C.E. Measurement of paraoxonase (PON1) status as a potential biomarker of susceptibility to organophosphate toxicity. Clin. Chim. Acta 2005, 352, 37–47. [Google Scholar] [CrossRef]
- Richter, R.J.; Jarvik, G.P.; Furlong, C.E. Paraoxonase 1 Status as a Risk Factor for Disease or Exposure. Adv. Exp. Med. Biol. 2009, 660, 29–35. [Google Scholar] [CrossRef] [Green Version]
- Gupta, N.; Singh, S.; Maturu, V.N.; Sharma, Y.P.; Gill, K.D. Paraoxonase 1 (PON1) Polymorphisms, Haplotypes and Activity in Predicting CAD Risk in North-West Indian Punjabis. PLoS ONE 2011, 6, e17805. [Google Scholar] [CrossRef] [Green Version]
- Tajbakhsh, A.; Rezaee, M.; Rivandi, M.; Forouzanfar, F.; Afzaljavan, F.; Pasdar, A. Paraoxonase 1 (PON1) and stroke; the dilemma of genetic variation. Clin. Biochem. 2017, 50, 1298–1305. [Google Scholar] [CrossRef]
- Brophy, V.H.; Jampsa, R.L.; Clendenning, J.B.; McKinstry, L.A.; Jarvik, G.P.; Furlong, C.E. Effects of 5′ Regulatory-Region Polymorphisms on Paraoxonase-Gene (PON1) Expression. Am. J. Hum. Genet. 2001, 68, 1428–1436. [Google Scholar] [CrossRef] [Green Version]
- Chen, J.; Chan, W.; Wallenstein, S.; Berkowitz, G.; Wetmur, J.G. Haplotype-Phenotype Relationships of Paraoxonase-1. Cancer Epidemiol. Biomark. Prev. 2005, 14, 731–734. [Google Scholar] [CrossRef] [Green Version]
- McDaniel, C.Y.; Dail, M.B.; Wills, R.W.; Chambers, H.W.; Chambers, J.E. Paraoxonase 1 Polymorphisms Within a Mississippi USA Population as Possible Biomarkers of Enzyme Activities Associated with Disease Susceptibility. Biochem. Genet. 2014, 52, 509–523. [Google Scholar] [CrossRef] [PubMed]
- Phuntuwate, W.; Suthisisang, C.; Koanantakul, B.; Mackness, M.I.; Mackness, B. Paraoxonase 1 status in the Thai population. J. Hum. Genet. 2005, 50, 293–300. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wysocka, A.; Zwolak, A. The Relevance of Noncoding DNA Variationsof Paraoxonase Gene Cluster in Atherosclerosis-Related Diseases. Int. J. Mol. Sci. 2021, 22, 2137. [Google Scholar] [CrossRef] [PubMed]
- Furlong, C.; Suzuki, S.; Stevens, R.; Marsillach, J.; Richter, R.; Jarvik, G.; Checkoway, H.; Samii, A.; Costa, L.; Griffith, A.; et al. Human PON1, a biomarker of risk of disease and exposure. Chem.-Biol. Interact. 2010, 187, 355–361. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rojasgarcia, A.E.; Solisheredia, M.J.; Pinaguzman, B.; Vega, L.; Lopezcarrillo, L.; Quintanillavega, B. Genetic polymorphisms and activity of PON1 in a Mexican population. Toxicol. Appl. Pharmacol. 2005, 205, 282–289. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Kumar, M.; Chan, W.; Berkowitz, G.; Wetmur, J.G. Increased influence of genetic variation on PON1 activity in neonates: An Independent Risk Factor for Nonfatal Arterial Ischemic Stroke among Young Adults. Environ. Health Perspect. 2003, 111, 1403–1409. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Voetsch, B.; Benke, K.S.; Damasceno, B.P.; Siqueira, L.H.; Loscalzo, J. Paraoxonase 192 Gln→Arg Polymorphism. Stroke 2002, 33, 1459–1464. [Google Scholar] [CrossRef] [Green Version]
- Sen-Banerjee, S.; Siles, X.; Campos, H. Tobacco Smoking Modifies Association between Gln-Arg192 Polymorphism of Human Paraoxonase Gene and Risk of Myocardial Infarction. Arter. Thromb. Vasc. Biol. 2000, 20, 2120–2126. [Google Scholar] [CrossRef]
- Bernal-Hernández, Y.Y.; Medina-Díaz, I.M.; Barrón-Vivanco, B.S.; Robledo-Marenco, M.D.L.; Girón-Pérez, M.I.; Pérez-Herrera, N.E.; Quintanilla-Vega, B.; Cerda-Flores, R.; Rojas-García, A.E. Paraoxonase 1 and Its Relationship with Pesticide Biomarkers in Indigenous Mexican Farmworkers. J. Occup. Environ. Med. 2014, 56, 281–290. [Google Scholar] [CrossRef]
- Allebrandt, K.V.; Souza, R.L.; Chautard-Freire-Maia, E.A. Variability of the Paraoxonase Gene (PON1) in Euro- and Afro-Brazilians. Toxicol. Appl. Pharmacol. 2002, 180, 151–156. [Google Scholar] [CrossRef]
- Pérez-Herrera, N.; May-Pech, C.; Hernández-Ochoa, I.; Castro-Mañé, J.; Rojas-García, E.; Borja-Aburto, V.H.; Castillo-Burguete, T.; Quintanilla-Vega, B. PON1Q192R polymorphism is associated with lipid profile in Mexican men with Mayan ascendancy. Exp. Mol. Pathol. 2008, 85, 129–134. [Google Scholar] [CrossRef] [PubMed]
- Holland, N.; Furlong, C.; Bastaki, M.; Richter, R.; Bradman, A.; Huen, K.; Beckman, K.; Eskenazi, B. Paraoxonase Polymorphisms, Haplotypes, and Enzyme Activity in Latino Mothersand Newborns. Environ. Health Perspect. 2006, 114, 985–991. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hernández, A.F.; Mackness, B.; Rodrigo, L.; López, O.; Pla, A.; Gil, F.; Durrington, P.N.; Pena, G.; Parrón, T.; Serrano, J.L.; et al. Paraoxonase activity and genetic polymorphisms in greenhouse workers with long term pesticide exposure. Hum. Exp. Toxicol. 2003, 22, 565–574. [Google Scholar] [CrossRef]
- Agachan, B.; Yilmaz, H.; Karaali, Z.; Isbir, T. Paraoxonase 55 and 192 polymorphism and its relationship to serum paraoxonase activity and serum lipids in Turkish patients with non-insulin dependent diabetes mellitus. Cell Biochem. Funct. 2004, 22, 163–168. [Google Scholar] [CrossRef] [PubMed]
- Leviev, I.; Deakin, S.; James, R.W. Decreased stability of the M54 isoform of paraoxonase as a contributory factor to variations in human serum paraoxonase concentrations. J. Lipid Res. 2001, 42, 528–535. [Google Scholar] [CrossRef]
- Davies, H.G.; Richter, R.J.; Keifer, M.; Broomfield, C.A.; Sowalla, J.; Furlong, C.E. The effect of the human serum paraoxonase polymorphism is reversed with diazoxon, soman and sarin. Nat. Genet. 1996, 14, 334–336. [Google Scholar] [CrossRef] [PubMed]
- Li, W.-F.; Costa, L.G.; Richter, R.J.; Hagen, T.; Shih, D.M.; Tward, A.; Lusis, A.J.; Furlong, C.E. Catalytic efficiency determines the in-vivo efficacy of PON1 for detoxifying organophosphorus compounds. Pharmacogenetics 2000, 10, 767–779. [Google Scholar] [CrossRef]
- Richter, R.J.; Furlong, C.E. Determination of paraoxonase (PON1) status requires more than genotyping. Pharmacogenetics 1999, 9, 745–753. [Google Scholar] [CrossRef]
- Billecke, S.; Draganov, D.; Counsell, R.; Stetson, P.; Watson, C.; Hsu, C.; La Du, B.N. Human serum paraoxonase (PON1) isozymes Q and R hydrolyze lactones and cyclic carbonate esters. Drug Metab. Dispos. 2000, 28, 1335–1342. [Google Scholar]
- Furlong, C.E.; Holland, N.; Richter, R.J.; Bradman, A.; Ho, A.; Eskenazi, B. PON1 status of farmworker mothers and children as a predictor of organophosphate sensitivity. Pharmacogenet. Genom. 2006, 16, 183–190. [Google Scholar] [CrossRef]
- Richter, R.J.; Jarvik, G.P.; Furlong, C.E. Determination of Paraoxonase 1 Status without the Use of Toxic Organophosphate Substrates. Circ. Cardiovasc. Genet. 2008, 1, 147–152. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bounafaa, A.; Berrougui, H.; Ghalim, N.; Nasser, B.; Bagri, A.; Moujahid, A.; Ikhlef, S.; Camponova, P.; Yamoul, N.; Simo, O.K.; et al. Association between Paraoxonase 1 (PON1) Polymorphisms and the Risk of Acute Coronary Syndrome in a North African Population. PLoS ONE 2015, 10, e0133719. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Banerjee, I. Relationship between Paraoxonase 1 (PON1) gene polymorphisms and susceptibility of stroke: A meta-analysis. Eur. J. Epidemiol. 2010, 25, 449–458. [Google Scholar] [CrossRef] [Green Version]
- Kim, D.S.; Crosslin, D.R.; Auer, P.L.; Suzuki, S.M.; Marsillach, J.; Burt, A.A.; Gordon, A.; Meschia, J.F.; Nalls, M.A.; Worrall, B.B.; et al. Rare coding variation in paraoxonase-1 is associated with ischemic stroke in the NHLBI Exome Sequencing Project. J. Lipid Res. 2014, 55, 1173–1178. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, N.S.; Kang, B.K.; Cha, M.H.; Oh, S.-M.; Ko, M.M.; Bang, O.-S. Association between PON1 5′-regulatory region polymorphisms, PON1 activity and ischemic stroke. Clin. Biochem. 2009, 42, 857–863. [Google Scholar] [CrossRef] [PubMed]
- Zhang, G.; Li, W.; Li, Z.; Lv, H.; Ren, Y.; Ma, R.; Li, X.; Kang, X.; Shi, Y.; Sun, Y. Association between paraoxonase gene and stroke in the Han Chinese population. BMC Med Genet. 2013, 14, 16. [Google Scholar] [CrossRef] [Green Version]
- Liu, M.-E.; Liao, Y.-C.; Lin, R.-T.; Wang, Y.-S.; Hsi, E.; Lin, H.-F.; Chen, K.-C.; Juo, S.-H.H. A functional polymorphism of PON1 interferes with microRNA binding to increase the risk of ischemic stroke and carotid atherosclerosis. Atherosclerosis 2013, 228, 161–167. [Google Scholar] [CrossRef]
- Hofer, S.E.; Bennetts, B.; Chan, A.K.; Holloway, B.; Karschimkus, C.; Jenkins, A.; Silink, M.; Donaghue, K.C. Association between PON 1 polymorphisms, PON activity and diabetes complications. J. Diabetes its Complicat. 2006, 20, 322–328. [Google Scholar] [CrossRef]
- Aviram, M.; Billecke, S.; Sorenson, R.; Bisgaier, C.; Newton, R.; Rosenblat, M.; Erogul, J.; Hsu, C.; Dunlop, C.; La Du, B. Paraoxonase Active Site Required for Protection Against LDL Oxidation Involves Its Free Sulfhydryl Group and Is Different from That Required for Its Arylesterase/Paraoxonase Activities: Selective Action of Human Paraoxonase Allozymes Q and R. Arter. Thromb. Vasc. Biol. 1998, 18, 1617–1624. [Google Scholar] [CrossRef] [Green Version]
- Luo, Z.; Li, S.; Muhammad, I.; Karim; Song, Y. Associations of the PON1 rs854560 polymorphism with plasma lipid levels: A meta-analysis. Lipids Health Dis. 2018, 17, 274. [Google Scholar] [CrossRef] [Green Version]
- Antikainen, M.; Murtomäki, S.; Syvänne, M.; Pahlman, R.; Tahvanainen, E.; Jauhiainen, M.; Frick, M.H.; Ehnholm, C. The Gln-Arg191 polymorphism of the human paraoxonase gene (HUMPONA) is not associated with the risk of coronary artery disease in Finns. J. Clin. Investig. 1996, 98, 883–885. [Google Scholar] [CrossRef] [PubMed]
- Arca, M.; Ombres, D.; Montali, A.; Campagna, F.; Mangieri, E.; Tanzilli, G.; Campa, P.P.; Ricci, G.; Verna, R.; Pannitteri, G. PON1 L55M polymorphism is not a predictor of coronary atherosclerosis either alone or in combination with Q192R polymorphism in an Italian population. Eur. J. Clin. Investig. 2002, 32, 9–15. [Google Scholar] [CrossRef] [PubMed]
- Costa, L.G.; Giordano, G.; Furlong, C.E. Pharmacological and dietary modulators of paraoxonase 1 (PON1) activity and expression: The hunt goes on. Biochem. Pharmacol. 2011, 81, 337–344. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Précourt, L.-P.; Amre, D.; Denis, M.-C.; Lavoie, J.-C.; Delvin, E.; Seidman, E.; Levy, E. The three-gene paraoxonase family: Physiologic roles, actions and regulation. Atherosclerosis 2011, 214, 20–36. [Google Scholar] [CrossRef]
- Schrader, C.; Rimbach, G. Determinants of Paraoxonase 1 Status: Genes, Drugs and Nutrition. Curr. Med. Chem. 2011, 18, 5624–5643. [Google Scholar] [CrossRef] [PubMed]
- Arii, K.; Suehiro, T.; Ikeda, Y.; Kumon, Y.; Inoue, M.; Inada, S.; Takata, H.; Ishibashi, A.; Hashimoto, K.; Terada, Y. Role of protein kinase C in pitavastatin-induced human paraoxonase I expression in Huh7 cells. Metabolism 2010, 59, 1287–1293. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Garige, M.; Gong, M.; Varatharajalu, R.; Lakshman, R. Quercetin up-regulates paraoxonase 1 gene expression via sterol regulatory element binding protein 2 that translocates from the endoplasmic reticulum to the nucleus where it specifically interacts with sterol responsive element–like sequence in paraoxonase 1 promoter in HuH7 liver cells. Metabolism 2010, 59, 1372–1378. [Google Scholar] [CrossRef] [Green Version]
- Cheng, C.-C.; Hsueh, C.-M.; Liang, K.-W.; Ting, C.-T.; Wen, C.-L.; Hsu, S.-L. Role of JNK and c-Jun signaling pathway in regulation of human serum paraoxonase 1 gene transcription by berberine in human HepG2 cells. Eur. J. Pharmacol. 2011, 650, 519–525. [Google Scholar] [CrossRef]
- Ponce-Ruiz, N.; Rojas-García, A.; Barrón-Vivanco, B.; Elizondo, G.; Bernal-Hernández, Y.; Mejía-García, A.; Medina-Díaz, I. Transcriptional regulation of human paraoxonase 1 by PXR and GR in human hepatoma cells. Toxicol. Vitr. 2015, 30, 348–354. [Google Scholar] [CrossRef]
- Chernyavskiy, I.; Veeranki, S.; Sen, U.; Tyagi, S.C. Atherogenesis: Hyperhomocysteinemia interactions with LDL, macrophage function, paraoxonase 1, and exercise. Ann. N. Y. Acad. Sci. 2016, 1363, 138–154. [Google Scholar] [CrossRef] [Green Version]
- Seres, I.; Paragh, G.; Deschene, E.; Fulop, T.; Khalil, A. Study of factors influencing the decreased HDL associated PON1 activity with aging. Exp. Gerontol. 2003, 39, 59–66. [Google Scholar] [CrossRef] [PubMed]
- Jaouad, L.; de Guise, C.; Berrougui, H.; Cloutier, M.; Isabelle, M.; Fulop, T.; Payette, H.; Khalil, A. Age-related decrease in high-density lipoproteins antioxidant activity is due to an alteration in the PON1′s free sulfhydyl groups. Atherosclerosis 2006, 185, 191–200. [Google Scholar] [CrossRef] [PubMed]
- Kumar, D.; Rizvi, S.I. Age-Dependent Paraoxonase 1 (PON1) Activity and LDL Oxidation in Wistar Rats during Their Entire Lifespan. Sci. World J. 2014, 2014, 538049. [Google Scholar] [CrossRef] [PubMed]
- Kumar, D.; Rizvi, S.I. Plasma paraoxonase 1 arylesterase activity in d-galactose-induced aged rat model: Correlation with LDL oxidation and redox status. Aging Clin. Exp. Res. 2013, 26, 261–267. [Google Scholar] [CrossRef] [PubMed]
- Cole, T.B.; Jampsa, R.L.; Walter, B.J.; Arndt, T.L.; Richter, R.J.; Shih, D.M.; Tward, A.; Lusis, A.J.; Jack, R.M.; Costa, L.G.; et al. Expression of human paraoxonase (PON1) during development. Pharmacogenetics 2003, 13, 357–364. [Google Scholar] [CrossRef]
- Lee, Y.S.; Park, C.O.; Noh, J.Y.; Jin, S.; Lee, N.R.; Noh, S.; Lee, J.H.; Lee, K.H. Knockdown of paraoxonase 1 expression influences the ageing of human dermal microvascular endothelial cells. Exp. Dermatol. 2012, 21, 682–687. [Google Scholar] [CrossRef]
- Garin, M.C.; James, R.W.; Dussoix, P.; Blanché, H.; Passa, P.; Froguel, P.; Ruiz, J. Paraoxonase polymorphism Met-Leu54 is associated with modified serum concentrations of the enzyme. A possible link between the paraoxonase gene and increased risk of cardiovascular disease in diabetes. J. Clin. Investig. 1997, 99, 62–66. [Google Scholar] [CrossRef] [Green Version]
- Demirdöğen, B.C.; Türkanoğlu, A.; Bek, S.; Sanisoğlu, Y.; Demirkaya, Ş.; Vural, O.; Arınç, E.; Adalı, O. Paraoxonase/arylesterase ratio, PON1 192Q/R polymorphism and PON1 status are associated with increased risk of ischemic stroke. Clin. Biochem. 2008, 41, 1–9. [Google Scholar] [CrossRef]
- Kim, D.S.; Burt, A.A.; Ranchalis, J.E.; Richter, R.J.; Marshall, J.K.; Eintracht, J.F.; Rosenthal, E.A.; Furlong, C.E.; Jarvik, G.P. Additional Common Polymorphisms in thePONGene Cluster Predict PON1 Activity but Not Vascular Disease. J. Lipids 2012, 2012, 476316. [Google Scholar] [CrossRef] [Green Version]
- Shenhar-Tsarfaty, S.; Waiskopf, N.; Ofek, K.; Shopin, L.; Usher, S.; Berliner, S.; Shapira, I.; Bornstein, N.M.; Ritov, Y.; Soreq, H.; et al. Atherosclerosis and arteriosclerosis parameters in stroke patients associate with paraoxonase polymorphism and esterase activities. Eur. J. Neurol. 2013, 20, 891–898. [Google Scholar] [CrossRef]
- Najafi, M.; Gohari, L.H.; Firoozrai, M. Paraoxonase 1 gene promoter polymorphisms are associated with the extent of stenosis in coronary arteries. Thromb. Res. 2009, 123, 503–510. [Google Scholar] [CrossRef] [PubMed]
- Mahrooz, A.; Gohari, G.; Hashemi-Soteh, S.M.; Zargari, M.; Musavi, H.; Abedini, M.; Alizadeh, A. R-carrying genotypes of serum paraoxonase (PON1) 192 polymorphism and higher activity ratio are related to susceptibility against ischemic stroke. Mol. Biol. Rep. 2012, 39, 11177–11185. [Google Scholar] [CrossRef] [PubMed]
- Shokri, Y.; Variji, A.; Nosrati, M.; Khonakdar-Tarsi, A.; Kianmehr, A.; Kashi, Z.; Bahar, A.; Bagheri, A.; Mahrooz, A. Importance of paraoxonase 1 (PON1) as an antioxidant and antiatherogenic enzyme in the cardiovascular complications of type 2 diabetes: Genotypic and phenotypic evaluation. Diabetes Res. Clin. Pract. 2020, 161, 108067. [Google Scholar] [CrossRef] [PubMed]
- Mackness, B.; Durrington, P.; McElduff, P.; Yarnell, J.; Azam, N.; Watt, M.; Mackness, M. Low Paraoxonase Activity Predicts Coronary Events in the Caerphilly Prospective Study. Circulation 2003, 107, 2775–2779. [Google Scholar] [CrossRef] [Green Version]
- Gamboa, R.; Regalado, J.C.; Huesca-Gómez, C.; Posadas-Romero, C.; Verdejo Paris, J.; Vargas-Alarcón, G.; Pérez-Méndez, O. Low paraoxonasa and arylesterase plasma activities in Mexican patients with coronary artery disease. Arch. Cardiol. Mex. 2008, 78, 360–368. [Google Scholar]
- Tang, W.W.; Hartiala, J.; Fan, Y.; Wu, Y.; Stewart, A.F.; Erdmann, J.; Kathiresan, S.; Roberts, R.; McPherson, R.; Allayee, H.; et al. Clinical and Genetic Association of Serum Paraoxonase and Arylesterase Activities with Cardiovascular Risk. Arter. Thromb. Vasc. Biol. 2012, 32, 2803–2812. [Google Scholar] [CrossRef] [Green Version]
- Ponce-Ruiz, N.; Murillo-González, F.; Rojas-García, A.; Barrón-Vivanco, B.; Bernal-Hernández, Y.; González-Arias, C.; Ortega-Cervantes, L.; Ponce-Gallegos, J.; López-Guarnido, O.; Medina-Díaz, I. PON1 status and homocysteine levels as potential biomarkers for cardiovascular disease. Exp. Gerontol. 2020, 140, 111062. [Google Scholar] [CrossRef]
- Shen, Y.; Ding, F.H.; Sun, J.T.; Pu, L.J.; Zhang, R.Y.; Zhang, Q.; Chen, Q.J.; Shen, W.F.; Lu, L. Association of elevated apoA-I glycation and reduced HDL-associated paraoxonase1, 3 activity, and their interaction with angiographic severity of coronary artery disease in patients with type 2 diabetes mellitus. Cardiovasc. Diabetol. 2015, 14, 52. [Google Scholar] [CrossRef] [Green Version]
- Kasprzak, M.P.; Iskra, M.; Majewski, W.; Budzyń-Napierała, M.; Gryszczyńska, B.; Strzyżewski, K.; Kasprzak, J. PON1 status evaluation in patients with chronic arterial occlusion of lower limbs due to atherosclerosis obliterans. Arch. Med. Sci. 2014, 6, 1101–1108. [Google Scholar] [CrossRef]
- Sentürk, T.; Sarandöl, E.; Güllülü, S.; Erdinç, S.; Ozdemir, B.; Baran, I.; Aydinlar, A. Association between paraoxonase 1 activity and severity of coronary artery disease in patients with acute coronary syndromes. Acta Cardiol. 2008, 63, 361–367. [Google Scholar] [CrossRef]
- Ikeda, Y.; Suehiro, T.; Itahara, T.; Inui, Y.; Chikazawa, H.; Inoue, M.; Arii, K.; Hashimoto, K. Human serum paraoxonase concentration predicts cardiovascular mortality in hemodialysis patients. Clin. Nephrol. 2007, 67, 358–365. [Google Scholar] [CrossRef] [PubMed]
- Bhattacharyya, T.; Nicholls, S.J.; Topol, E.J.; Zhang, R.; Yang, X.; Schmitt, D.; Fu, X.; Shao, M.; Brennan, D.M.; Ellis, S.G.; et al. Relationship of Paraoxonase 1 (PON1) Gene Polymorphisms and Functional Activity with Systemic Oxidative Stress and Cardiovascular Risk. JAMA 2008, 299, 1265–1276. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- van Himbergen, T.M.; van der Schouw, Y.T.; Voorbij, H.A.; van Tits, L.J.; Stalenhoef, A.F.; Peeters, P.H.; Roest, M. Paraoxonase (PON1) and the risk for coronary heart disease and myocardial infarction in a general population of Dutch women. Atherosclerosis 2008, 199, 408–414. [Google Scholar] [CrossRef] [PubMed]
- Ikeda, Y.; Inoue, M.; Suehiro, T.; Arii, K.; Kumon, Y.; Hashimoto, K. Low human paraoxonase predicts cardiovascular events in Japanese patients with type 2 diabetes. Geol. Rundsch. 2008, 46, 239–242. [Google Scholar] [CrossRef]
- Rosenblat, M.; Karry, R.; Aviram, M. Paraoxonase 1 (PON1) is a more potent antioxidant and stimulant of macrophage cholesterol efflux, when present in HDL than in lipoprotein-deficient serum: Relevance to diabetes. Atherosclerosis 2006, 187, 74.e1–74.e10. [Google Scholar] [CrossRef]
- Koren-Gluzer, M.; Aviram, M.; Meilin, E.; Hayek, T. The antioxidant HDL-associated paraoxonase-1 (PON1) attenuates diabetes development and stimulates β-cell insulin release. Atherosclerosis 2011, 219, 510–518. [Google Scholar] [CrossRef]
- Hedrick, C.C.; Thorpe, S.R.; Fu, M.-X.; Harper, C.M.; Yoo, J.; Kim, S.-M.; Wong, H.; Peters, A.L. Glycation impairs high-density lipoprotein function. Diabetologia 2000, 43, 312–320. [Google Scholar] [CrossRef] [Green Version]
- Kalmár, T.; Seres, I.; Balogh, Z.; Káplár, M.; Winkler, G.; Paragh, G. Correlation between the activities of lipoprotein lipase and paraoxonase in type 2 diabetes mellitus. Diabetes Metab. 2005, 31, 574–580. [Google Scholar] [CrossRef]
- Prakash, M.; Kedage, V.; Muttigi, M.S.; Shetty, M.S.; Suvarna, R.; Rao, S.S.; Joshi, C. Serum paraoxonase 1 activity status in patients with liver disorders. Saudi J. Gastroenterol. 2010, 16, 79–83. [Google Scholar] [CrossRef]
- Kotani, K.; Watanabe, J.; Miura, K.; Gugliucci, A. Paraoxonase 1 and Non-Alcoholic Fatty Liver Disease: A Meta-Analysis. Molecules 2021, 26, 2323. [Google Scholar] [CrossRef]
- Ferré, N.; Camps, J.; Prats, E.; Vilella, E.; Paul, A.; Figuera, L.; Joven, J. Serum paraoxonase activity: A new additional test for the improved evaluation of chronic liver damage. Clin. Chem. 2002, 48, 261–268. [Google Scholar] [CrossRef] [PubMed]
- Ferré, N.; Marsillach, J.; Camps, J.; Mackness, B.; Mackness, M.; Riu, F.; Coll, B.; Tous, M.; Joven, J. Paraoxonase-1 is associated with oxidative stress, fibrosis and FAS expression in chronic liver diseases. J. Hepatol. 2006, 45, 51–59. [Google Scholar] [CrossRef] [PubMed]
- Wang, B.; Yang, R.-N.; Zhu, Y.-R.; Xing, J.-C.; Lou, X.-W.; He, Y.-J.; Ding, Q.-L.; Zhang, M.-Y.; Qiu, H. Involvement of xanthine oxidase and paraoxonase 1 in the process of oxidative stress in nonalcoholic fatty liver disease. Mol. Med. Rep. 2016, 15, 387–395. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Goswami, B.; Tayal, D.; Gupta, N.; Mallika, V. Paraoxonase: A multifaceted biomolecule. Clin. Chim. Acta 2009, 410, 1–12. [Google Scholar] [CrossRef]
- Saeidi, M.; Shakeri, R.; Marjani, A.; Khajeniazi, S. Alzheimer’s Disease and Paraoxonase 1 (PON1) Gene Polymorphisms. Open Biochem. J. 2017, 11, 47–55. [Google Scholar] [CrossRef] [Green Version]
- Pola, R.; Gaetani, E.; Flex, A.; Gerardino, L.; Aloi, F.; Flore, R.; Serricchio, M.; Pola, P.; Bernabei, R. Lack of association between Alzheimer’s disease and Gln-Arg 192 Q/R polymorphism of the PON-1 gene in an Italian population. Dement. Geriatr. Cogn. Disord. 2003, 15, 88–91. [Google Scholar] [CrossRef]
- Shi, J.J.; Zhang, S.Z.; Ma, C.; Tang, M.N.; Liu, X.H.; Wang, Y.C.; Han, H.Y.; Guo, Y.B.; Feng, R.M.; Miao, G.D. Gln192 Arg polymorphism of the paraoxonase-1 gene is not associated with Alzheimer’s disease in Chinese. J. First Mil. Med. Univ. 2004, 24, 371–374. [Google Scholar]
- Romani, A.; Trentini, A.; Van Der Flier, W.M.; Bellini, T.; Zuliani, G.; Cervellati, C.; Teunissen, C.E. Arylesterase Activity of Paraoxonase-1 in Serum and Cerebrospinal Fluid of Patients with Alzheimer’s Disease and Vascular Dementia. Antioxidants 2020, 9, 456. [Google Scholar] [CrossRef]
- Trentini, A.; Bellini, T.; Bonaccorsi, G.; Cavicchio, C.; Hanau, S.; Passaro, A.; Cervellati, C. Sex difference: An important issue to consider in epidemiological and clinical studies dealing with serum paraoxonase-1. J. Clin. Biochem. Nutr. 2019, 64, 250–256. [Google Scholar] [CrossRef] [Green Version]
- Kumru, S.; Aydin, S.; Aras, A.; Gursu, M.; Gulcu, F. Effects of Surgical Menopause and Estrogen Replacement Therapy on Serum Paraoxonase Activity and Plasma Malondialdehyde Concentration. Gynecol. Obstet. Investig. 2005, 59, 108–112. [Google Scholar] [CrossRef]
- Sepahvand, F.; Shafiei, M.; Ghaffari, S.M.; Rahimi-Moghaddam, P.; Mahmoudian, M. Paraoxonase Phenotype Distribution in a Healthy Iranian Population. Basic Clin. Pharmacol. Toxicol. 2007, 101, 104–107. [Google Scholar] [CrossRef] [PubMed]
- Mascarenhas-Melo, F.; Marado, D.; Palavra, F.; Sereno, J.; Coelho, Á.; Pinto, R.; Teixeira-Lemos, E.; Teixeira, F.; Reis, F. Diabetes abrogates sex differences and aggravates cardiometabolic risk in postmenopausal women. Cardiovasc. Diabetol. 2013, 12, 61. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bin Ali, A.; Zhang, Q.; Lim, Y.K.; Fang, D.; Retnam, L.; Lim, S.-K. Expression of major HDL-associated antioxidant PON-1 is gender dependent and regulated during inflammation. Free Radic. Biol. Med. 2003, 34, 824–829. [Google Scholar] [CrossRef]
- Thomàs-Moyà, E.; Gianotti, M.; Lladó, I.; Proenza, A.M. Effects of caloric restriction and gender on rat serum paraoxonase 1 activity. J. Nutr. Biochem. 2006, 17, 197–203. [Google Scholar] [CrossRef]
- Fuhrman, B. Regulation of Hepatic Paraoxonase-1 Expression. J. Lipids 2012, 2012, 684010. [Google Scholar] [CrossRef] [Green Version]
- Deakin, S.P.; James, R.W. Genetic and environmental factors modulating serum concentrations and activities of the antioxidant enzyme paraoxonase-1. Clin. Sci. 2004, 107, 435–447. [Google Scholar] [CrossRef] [Green Version]
- Navarro-García, F.; Ponce-Ruíz, N.; Rojas-García, A.E.; Ávila-Villarreal, G.; Herrera-Moreno, J.F.; Barrón-Vivanco, B.S.; Bernal-Hernández, Y.Y.; González-Arias, C.A.; Medina-Díaz, I.M. The Role of Nutritional Habits and Moderate Red Wine Consumption in PON1 Status in Healthy Population. Appl. Sci. 2021, 11, 9503. [Google Scholar] [CrossRef]
- Gouédard, C.; Barouki, R.; Morel, Y. Dietary Polyphenols Increase Paraoxonase 1 Gene Expression by an Aryl Hydrocarbon Receptor-Dependent Mechanism. Mol. Cell. Biol. 2004, 24, 5209–5222. [Google Scholar] [CrossRef] [Green Version]
- Shen, H.; Robertson, L.W.; Ludewig, G. Regulation of paraoxonase 1 (PON1) in PCB 126-exposed male Sprague Dawley rats. Toxicol. Lett. 2012, 209, 291–298. [Google Scholar] [CrossRef] [Green Version]
- Shen, H.; Li, M.; Wang, B.; Lai, I.K.; Robertson, L.W.; Ludewig, G. Dietary antioxidants (selenium and N-acetylcysteine) modulate paraoxonase 1 (PON1) in PCB 126-exposed rats. Environ. Sci. Pollut. Res. 2013, 21, 6384–6399. [Google Scholar] [CrossRef] [Green Version]
- Shen, H.; Robertson, L.W.; Ludewig, G. Regulatory effects of dioxin-like and non-dioxin-like PCBs and other AhR ligands on the antioxidant enzymes paraoxonase 1/2/3. Environ. Sci. Pollut. Res. Int. 2015, 23, 2108–2118. [Google Scholar] [CrossRef] [PubMed]
- Guyot, E.; Coumoul, X.; Chassé, J.-F.; Khallouki, F.; Savouret, J.F.; Poirot, M.; Barouki, R. Identification of a new stilbene-derived inducer of paraoxonase 1 and ligand of the Aryl hydrocarbon Receptor. Biochem. Pharmacol. 2012, 83, 627–632. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lou-Bonafonte, J.M.; Gabás-Rivera, C.; Navarro, M.A.; Osada, J. PON1 and Mediterranean Diet. Nutrients 2015, 7, 4068–4092. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tomás, M.; Sentí, M.; García-Faria, F.; Vila, J.; Torrents, A.; Covas, M.; Marrugat, J. Effect of Simvastatin Therapy on Paraoxonase Activity and Related Lipoproteins in Familial Hypercholesterolemic Patients. Arter. Thromb. Vasc. Biol. 2000, 20, 2113–2119. [Google Scholar] [CrossRef] [Green Version]
- Harangi, M.; Varga, Z.; Emri, G.; Seres, I.; Szilvássy, Z.; Paragh, G.; Remenyik, E. Atorvastatin effect on high-density lipoprotein-associated paraoxonase activity and oxidative DNA damage. Eur. J. Clin. Pharmacol. 2004, 60, 685–691. [Google Scholar] [CrossRef]
- Kassai, A.; Illyés, L.; Mirdamadi, H.Z.; Seres, I.; Kalmár, T.; Audikovszky, M.; Paragh, G. The effect of atorvastatin therapy on lecithin:cholesterol acyltransferase, cholesteryl ester transfer protein and the antioxidant paraoxonase. Clin. Biochem. 2007, 40, 1–5. [Google Scholar] [CrossRef]
- Mirdamadi, H.Z.; Sztanek, F.; Derdak, Z.; Seres, I.; Harangi, M.; Paragh, G. The human paraoxonase-1 phenotype modifies the effect of statins on paraoxonase activity and lipid parameters. Br. J. Clin. Pharmacol. 2008, 66, 366–374. [Google Scholar] [CrossRef] [Green Version]
- Shen, Y.; Wu, H.; Wang, C.; Shao, H.; Huang, H.; Jing, H.; Li, D. Simvastatin attenuates cardiopulmonary bypass-induced myocardial inflammatory injury in rats by activating peroxisome proliferator-activated receptor γ. Eur. J. Pharmacol. 2010, 649, 255–262. [Google Scholar] [CrossRef]
- Khateeb, J.; Kiyan, Y.; Aviram, M.; Tkachuk, S.; Dumler, I.; Fuhrman, B. Urokinase-Type Plasminogen Activator Downregulates Paraoxonase 1 Expression in Hepatocytes by Stimulating Peroxisome Proliferator–Activated Receptor-γ Nuclear Export. Arter. Thromb. Vasc. Biol. 2012, 32, 449–458. [Google Scholar] [CrossRef] [Green Version]
- Dail, M.B.; Pickin, R.; Crow, J.A.; Chambers, J.E. The Effect ofIn VitroDieldrin Exposure on the Rat Paraoxonase 1 (Pon1) Promoter. J. Biochem. Mol. Toxicol. 2014, 28, 224–231. [Google Scholar] [CrossRef]
- Coumoul, X.; Diry, M.; Barouki, R. PXR-dependent induction of human CYP3A4 gene expression by organochlorine pesticides. Biochem. Pharmacol. 2002, 64, 1513–1519. [Google Scholar] [CrossRef]
- Guo, R.; Chen, L.; Lian, J.; Wu, B.; Hong, G.; Ge, Y.; Liu, Y.; Qiu, Q.; Lu, Z. The protective effect of regulation of paraoxonase 1 gene on liver oxidative stress injury induced by dichlorvos poisoning in mice. Zhonghua Wei Zhong Bing Ji Jiu Yi Xue 2015, 27, 285–290. [Google Scholar] [CrossRef] [PubMed]
- Wagner, A.E.; Boesch-Saadatmandi, C.; Breckwoldt, D.; Schrader, C.; Schmelzer, C.; Döring, F.; Hashida, K.; Hori, O.; Matsugo, S.; Rimbach, G. Ascorbic acid partly antagonizes resveratrol mediated heme oxygenase-1 but not paraoxonase-1 induction in cultured hepatocytes—Role of the redox-regulated transcription factor Nrf2. BMC Complement. Altern. Med. 2011, 11, 1. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Esatbeyoglu, T.; Ulbrich, K.; Rehberg, C.; Rohn, S.; Rimbach, G. Thermal stability, antioxidant, and anti-inflammatory activity of curcumin and its degradation product 4-vinyl guaiacol. Food Funct. 2015, 6, 887–893. [Google Scholar] [CrossRef] [Green Version]
- Fortunato, J.J.; Feier, G.; Vitali, A.M.; Petronilho, F.C.; Dal-Pizzol, F.; Quevedo, J. Malathion-induced Oxidative Stress in Rat Brain Regions. Neurochem. Res. 2006, 31, 671–678. [Google Scholar] [CrossRef] [PubMed]
- Karami-Mohajeri, S.; Abdollahi, M. Toxic influence of organophosphate, carbamate, and organochlorine pesticides on cellular metabolism of lipids, proteins, and carbohydrates: A systematic review. Hum. Exp. Toxicol. 2010, 30, 1119–1140. [Google Scholar] [CrossRef]
- Lee, C.-H.; Kamijima, M.; Kim, H.; Shibata, E.; Ueyama, J.; Suzuki, T.; Takagi, K.; Saito, I.; Gotoh, M.; Hibi, H.; et al. 8-Hydroxydeoxyguanosine levels in human leukocyte and urine according to exposure to organophosphorus pesticides and paraoxonase 1 genotype. Int. Arch. Occup. Environ. Health 2006, 80, 217–227. [Google Scholar] [CrossRef]
- Brinholi, F.F.; Noto, C.; Maes, M.; Bonifácio, K.L.; Brietzke, E.; Ota, V.K.; Gadelha, A.; Cordeiro, Q.; Belangero, S.; Bressan, R.; et al. Lowered paraoxonase 1 (PON1) activity is associated with increased cytokine levels in drug naïve first episode psychosis. Schizophr. Res. 2015, 166, 225–230. [Google Scholar] [CrossRef]
- Li, Y. Modern epigenetics methods in biological research. Methods 2020, 187, 104–113. [Google Scholar] [CrossRef]
- Giorda, R. Chapter 1—Principles of epigenetics and DNA methylation. In Developmental Human Behavioral Epigenetics, 1st ed.; Provenci, L., Montirosso, R., Provenzi, L., Montirosso, R., Eds.; Academic press: Cambridge MA, USA, 2020; Volume 23, pp. 3–26. [Google Scholar]
- Al Aboud, N.M.; Tupper, C.; Jialal, I. Genetics, Epigenetic Mechanism; StatPearls: Treasure Island, FL, USA, 2021; Volume 1. [Google Scholar]
- Yen, I.H.; Syme, S.L. The Social Environment and Health: A Discussion of the Epidemiologic Literature. Annu. Rev. Public Health 1999, 20, 287–308. [Google Scholar] [CrossRef]
- Von dem Knesebeck, O. Concepts of social epidemiology in health services research. BMC Health Serv. Res. 2015, 15, 357. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huen, K.; Yousefi, P.; Street, K.; Eskenazi, B.; Holland, N. PON1 as a model for integration of genetic, epigenetic, and expression data on candidate susceptibility genes. Environ. Epigenetics 2015, 1, dvv003. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Holland, N.; Lizarraga, D.; Huen, K. Recent progress in the genetics and epigenetics of paraoxonase: Why it is relevant to children’s environmental health. Curr. Opin. Pediatr. 2015, 27, 240–247. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De La Iglesia, R.; Mansego, M.L.; Sánchez-Muniz, F.J.; Zulet, M.A.; Martinez, J.A. Arylesterase activity is associated with antioxidant intake and paraoxonase-1 (PON1) gene methylation in metabolic syndrome patients following an energy restricted diet. EXCLI J. 2014, 13, 416–426. [Google Scholar] [PubMed]
- Gomez-Uriz, A.M.; Goyenechea, E.; Campion, J.; De Arce, A.; Martinez, M.T.; Puchau, B.; Milagro, F.I.; Abete, I.; Martínez, J.A.; Lopez De Munain, A. Epigenetic patterns of two gene promoters (TNF-α and PON) in stroke considering obesity condition and dietary intake. J. Physiol. Biochem. 2014, 70, 603–614. [Google Scholar] [CrossRef]
- Lei, H.-P.; Yu, X.-Y.; Wu, H.; Kang, Y.-H.; Zhong, W.-P.; Cai, L.-Y.; Zhang, M.-Z.; Chen, J.-Y.; Mai, L.-P.; Ding, Q.-S.; et al. Effects of PON1 Gene Promoter DNA Methylation and Genetic Variations on the Clinical Outcomes of Dual Antiplatelet Therapy for Patients Undergoing Percutaneous Coronary Intervention. Clin. Pharmacokinet. 2017, 57, 817–829. [Google Scholar] [CrossRef]
- Cardenas, A.; Rifas-Shiman, S.L.; Agha, G.; Hivert, M.-F.; Litonjua, A.A.; DeMeo, D.L.; Lin, X.; Amarasiriwardena, C.J.; Oken, E.; Gillman, M.W.; et al. Persistent DNA methylation changes associated with prenatal mercury exposure and cognitive performance during childhood. Sci. Rep. 2017, 7, 288. [Google Scholar] [CrossRef]
- Declerck, K.; Remy, S.; Wohlfahrt-Veje, C.; Main, K.M.; Van Camp, G.; Schoeters, G.; Berghe, W.V.; Andersen, H.R. Interaction between prenatal pesticide exposure and a common polymorphism in the PON1 gene on DNA methylation in genes associated with cardio-metabolic disease risk—An exploratory study. Clin. Epigenetics 2017, 9, 35. [Google Scholar] [CrossRef] [Green Version]
- Diels, S.; Cuypers, B.; Tvarijonaviciute, A.; Derudas, B.; Van Dijck, E.; Verrijken, A.; Van Gaal, L.F.; Laukens, K.; Lefebvre, P.; Ceron, J.J.; et al. A targeted multi-omics approach reveals paraoxonase-1 as a determinant of obesity-associated fatty liver disease. Clin. Epigenetics 2021, 13, 158. [Google Scholar] [CrossRef]
- Strakovsky, R.S.; Zhang, X.; Zhou, D.; Pan, Y.-X. The regulation of hepatic Pon1 by a maternal high-fat diet is gender specific and may occur through promoter histone modifications in neonatal rats. J. Nutr. Biochem. 2014, 25, 170–176. [Google Scholar] [CrossRef]
- Niculescu, L.S.; Simionescu, N.; Sanda, G.M.; Carnuta, M.G.; Stancu, C.S.; Popescu, A.C.; Popescu, M.; Vlad, A.; Dimulescu, D.R.; Simionescu, M.; et al. MiR-486 and miR-92a Identified in Circulating HDL Discriminate between Stable and Vulnerable Coronary Artery Disease Patients. PLoS ONE 2015, 10, e0140958. [Google Scholar] [CrossRef] [PubMed]
- Lv, M.; Sun, D.; Chen, L. Identification of Prognostic Value of Rs3735590 Polymorphism in 3′-Untranslated Region (3′-UTR) of Paraoxonase 1 (PON-1) in Chronic Obstructive Pulmonary Disease Patients who Received Coronary Artery Bypass Grafting (CABG). Cell. Physiol. Biochem. 2018, 47, 1809–1818. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Chen, S.; Zhu, M.; Zhang, W.; Zhang, H.; Li, H.; Zou, C. Functional SNP in the 3′UTR of PON1 is Associated with the Risk of Calcific Aortic Valve Stenosis via MiR-616. Cell. Physiol. Biochem. 2018, 45, 1390–1398. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mahrooz, A.; Mackness, M. Epigenetics of paraoxonases. Curr. Opin. Lipidol. 2020, 31, 200–205. [Google Scholar] [CrossRef] [PubMed]
- Balci, H.; Genc, H.; Papila, C.; Can, G.; Papila, B.; Yanardag, H.; Uzun, H. Serum Lipid Hydroperoxide Levels and Paraoxonase Activity in Patients with Lung, Breast, and Colorectal Cancer. J. Clin. Lab. Anal. 2012, 26, 155–160. [Google Scholar] [CrossRef]
- Saadat, M. Paraoxonase 1 genetic polymorphisms and susceptibility to breast cancer: A meta-analysis. Cancer Epidemiol. 2012, 36, e101–e103. [Google Scholar] [CrossRef]
- Eroglu, M.; Yilmaz, N.; Yalcinkaya, S.; Ay, N.; Aydin, O.; Sezer, C. Enhanced HDL-cholesterol-associated anti-oxidant PON-1 activity in prostate cancer patients. Kaohsiung J. Med Sci. 2013, 29, 368–373. [Google Scholar] [CrossRef] [Green Version]
- Ahmed, N.S.; Shafik, N.; Elraheem, O.A.; Abou-Elnoeman, S.-E.A. Association of Paraoxonase-1(Q192R and L55M) Gene Polymorphisms and Activity with Colorectal Cancer and Effect of Surgical Intervention. Asian Pac. J. Cancer Prev. 2015, 16, 803–809. [Google Scholar] [CrossRef] [Green Version]
- Bacchetti, T.; Ferretti, G.; Sahebkar, A. The role of paraoxonase in cancer. Semin. Cancer Biol. 2019, 56, 72–86. [Google Scholar] [CrossRef]
- Kamel, E.-T.A.; Hussein, Z.E.-D.; Mahmoud, F.A.A.; Ibrahim, M.G.; Abd El-Fatah, A.E.S.E. The interplay between paraoxonase-1 and epigenetic changes in colorectal carcinoma. Afr. J. Biochem. Res. 2018, 12, 63–72. [Google Scholar] [CrossRef] [Green Version]
- Li, X.; Yu, Q. PON1 hypermethylation is associated with progression of renal cell carcinoma. J. Cell. Mol. Med. 2019, 23, 6646–6657. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Utangac, M.M.; Yeni, E.; Savas, M.; Altunkol, A.; Ciftci, H.; Gumus, K.; Demir, M. Paraoxonase and arylesterase activity in bladder cancer. Turk. J. Urol. 2017, 43, 147–151. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Akçay, M.N.; Yilmaz, I.; Polat, M.F.; Akçay, G. Serum paraoxonase levels in gastric cancer. Hepatogastroenterology 2003, 50 (Suppl. S2), cclxxiii–cclxxv. [Google Scholar]
- Bulbuller, N.; Eren, E.; Ellidag, H.Y.; Oner, O.Z.; Sezer, C.; Aydin, O.; Yılmaz, N. Diagnostic value of thiols, paraoxonase 1, arylesterase and oxidative balance in colorectal cancer in human. Neoplasma 2013, 60, 419–424. [Google Scholar] [CrossRef] [Green Version]
- Samra, Z.Q.; Pervaiz, S.; Shaheen, S.; Dar, N.; Athar, M.A. Determination of oxygen derived free radicals producer (xanthine oxidase) and scavenger (paraoxonase1) enzymes and lipid parameters in different cancer patients. Clin. Lab. 2011, 57, 741–747. [Google Scholar]
- Kafadar, A.M.; Ergen, A.; Zeybek, U.; Agachan, B.; Kuday, C.; Isbir, T. Paraoxonase 192 gene polymorphism and serum paraoxonase activity in high grade gliomas and meningiomas. Cell Biochem. Funct. 2005, 24, 455–460. [Google Scholar] [CrossRef]
- Ellidag, H.Y.; Eren, E.; Aydin, O.; Yıldırım, M.; Sezer, C.; Yilmaz, N. Multiple Myeloma: Relationship to Antioxidant Esterases. Med. Princ. Pract. 2014, 23, 18–23. [Google Scholar] [CrossRef]
- Huang, C.; Wang, Y.; Liu, S.; Ding, G.; Liu, W.; Zhou, J.; Kuang, M.; Ji, Y.; Kondo, T.; Fan, J. Quantitative Proteomic Analysis Identified Paraoxonase 1 as a Novel Serum Biomarker for Microvascular Invasion in Hepatocellular Carcinoma. J. Proteome Res. 2013, 12, 1838–1846. [Google Scholar] [CrossRef]
- Zhang, M.; Xiong, H.; Fang, L.; Lu, W.; Wu, X.; Huang, Z.-S.; Wang, Y.-Q.; Cai, Z.-M.; Wu, S. Paraoxonase 1 (PON1) Q192R Gene Polymorphism and Cancer Risk: A Meta-Analysis Based on 30 Publications. Asian Pac. J. Cancer Prev. 2015, 16, 4457–4463. [Google Scholar] [CrossRef] [Green Version]
- Pan, X.; Huang, L.; Li, M.; Mo, D.; Liang, Y.; Liu, Z.; Huang, Z.; Huang, L.; Liu, J.; Zhu, B. The Association between PON1 (Q192R and L55M) Gene Polymorphisms and Risk of Cancer: A Meta-Analysis Based on 43 Studies. BioMed Res. Int. 2019, 2019, 1–14. [Google Scholar] [CrossRef]
- Liu, P.; Wang, Q.; Cui, Y.; Wang, J. A meta-analysis of the relationship between paraoxonase 1 polymorphisms and cancer. Free Radic. Res. 2019, 53, 1045–1050. [Google Scholar] [CrossRef] [PubMed]
- Stevens, V.L.; Rodriguez, C.; Pavluck, A.L.; Thun, M.J.; Calle, E.E. Association of Polymorphisms in the Paraoxonase 1 Gene with Breast Cancer Incidence in the CPS-II Nutrition Cohort. Cancer Epidemiol. Biomark. Prev. 2006, 15, 1226–1228. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hussein, Y.M.; Gharib, A.; Etewa, R.L.; Elsawy, W. Association of L55M and Q192R polymorphisms in paraoxonase 1 (PON1) gene with breast cancer risk and their clinical significance. Mol. Cell. Biochem. 2011, 351, 117–123. [Google Scholar] [CrossRef] [PubMed]
- Naidu, R.; Har, Y.C.; Taib, N.A.M. Genetic Polymorphisms of Paraoxonase 1 (PON1) Gene: Association between L55M or Q192R with Breast Cancer Risk and Clinico-Pathological Parameters. Pathol. Oncol. Res. 2010, 16, 533–540. [Google Scholar] [CrossRef]
- Conesa-Zamora, P.; Ruiz-Cosano, J.; Torres-Moreno, D.; Español, I.; Gutiérrez-Meca, M.D.; Trujillo-Santos, J.; Pérez-Ceballos, E.; González-Conejero, R.; Corral, J.; Vicente, V.; et al. Polymorphisms in xenobiotic metabolizing genes (EPHX1, NQO1 and PON1) in lymphoma susceptibility: A case control study. BMC Cancer 2013, 13, 228. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, P.; Zhao, L.; Zou, P.; Lu, A.; Liu, N.; Yan, W.; Kang, C.; Fu, Z.; You, Y.; Jiang, T. Genetic oxidative stress variants and glioma risk in a Chinese population: A hospital-based case–control study. BMC Cancer 2012, 12, 617. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- de Aguiar Gonçalves, B.A.; Brazilian Collaborative Study Group of Infant Acute Leukemia; Vasconcelos, G.M.; Thuler, L.C.S.; Andrade, C.; Faro, A.; De-Pombo-Oliveira, M.S. NQO1 rs1800566 (C609T), PON1 rs662 (Q192R), and PON1 rs854560 (L55M) polymorphisms segregate the risk of childhood acute leukemias according to age range distribution. Cancer Causes Control 2012, 23, 1811–1819. [Google Scholar] [CrossRef]
- Kokouva, M.; Koureas, M.; Dardiotis, E.; Almpanidou, P.; Kalogeraki, A.; Kyriakou, D.; Hadjigeorgiou, G.M.; Hadjichristodoulou, C. Relationship between the paraoxonase 1 (PON1) M55L and Q192R polymorphisms and lymphohaematopoietic cancers in a Greek agricultural population. Toxicology 2013, 307, 12–16. [Google Scholar] [CrossRef]
- Aksoy-Sagirli, P.; Cakmakoglu, B.; Isbir, T.; Kaytan-Saglam, E.; Kizir, A.; Topuz, E.; Berkkan, H. Paraoxonase-1 192/55 Polymorphisms and the Risk of Lung Cancer in a Turkish Population. Anticancer Res. 2011, 31, 2225–2230. [Google Scholar]
- Lurie, G.; Wilkens, L.R.; Thompson, P.J.; McDuffie, K.E.; Carney, M.E.; Terada, K.Y.; Goodman, M.T. Genetic Polymorphisms in theParaoxonase 1Gene and Risk of Ovarian Epithelial Carcinoma. Cancer Epidemiol. Biomark. Prev. 2008, 17, 2070–2077. [Google Scholar] [CrossRef] [Green Version]
- Ergen, A.; Kılıcoglu, O.; Ozger, H.; Agachan, B.; Isbir, T.; Kilicoglu, O.; Cakmakoglu, B. Paraoxonase 1 192 and 55 polymorphisms in osteosarcoma. Mol. Biol. Rep. 2010, 38, 4181–4184. [Google Scholar] [CrossRef] [PubMed]
- Martínez, C.; Molina, J.A.; Alonso-Navarro, H.; Jiménez-Jiménez, F.J.; Agúndez, J.A.; García-Martín, E. Two common nonsynonymous paraoxonase 1 (PON1) gene polymorphisms and brain astrocytoma and meningioma. BMC Neurol. 2010, 10, 71. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oztürk, O.; Kağnici, O.F.; Oztürk, T.; Durak, H.; Tüzüner, B.M.; Kisakesen, H.I.; Cakalir, C.; Isbir, T. 192R allele of paraoxanase 1 (PON1) gene as a new marker for susceptibility to bladder cancer. Cell Biochem. Funct. 2009, 29, 4041–4046. [Google Scholar]
- Kerridge, I.; Lincz, L.; Scorgie, F.; Hickey, D.; Granter, N.; Spencer, A. Association between xenobiotic gene polymorphisms and non-Hodgkin’s lymphoma risk. Br. J. Haematol. 2002, 118, 477–481. [Google Scholar] [CrossRef]
- Arpaci, A.; Görmüs, U.; Dalan, B.; Berkman, S.; Isbir, T. Investigation of PON1 192 and PON1 55 polymorphisms in ovarian cancer patients in Turkish population. In Vivo 2009, 23, 421–424. [Google Scholar]
- Nielsen, S.S.; Mueller, B.A.; De Roos, A.J.; Viernes, H.-M.A.; Farin, F.M.; Checkoway, H. Risk of Brain Tumors in Children and Susceptibility to Organophosphorus Insecticides: The Potential Role of Paraoxonase (PON1). Environ. Health Perspect. 2005, 113, 909–913. [Google Scholar] [CrossRef] [Green Version]


| Cancer Type | Type of Study Control vs. Case | PON1 Activity | Reference |
|---|---|---|---|
| Bladder cancer | Control (n = 57) | PONase (U/L):137.6 ± 53.3 | [315] |
| AREase (U/L): 168.82 ± 37.4 | |||
| Case (n = 56) | PONase (U/L):103.35 ± 41.1 | ||
| AREase (U/L): 131.8 ± 39.9 | |||
| Colorectal cancer | Control (n = 25) | PONase (U/mL):124.8 ± 21.7 | [308] |
| AREase (U/mL): 98.5 ± 18.8 | |||
| Case (n = 40) | PONase (U/mL):65.9 ± 22.5 | ||
| AREase (U/mL): 57.5 ± 13.34 | |||
| Control (n = 39) | PONase (U/L):230.5 ± 97.9 | [316] | |
| AREase (k/mL): 230.7 ± 53.8 | |||
| Case (n = 40) | PONase (U/L):128.2 ± 63.2 | ||
| AREase (k/mL): 150.6 ± 49.2 | |||
| Control (n = 80) | PONase (U/mL):394.1 ± 83.4 | [317] | |
| AREase (U/mL): 228.4 ± 83.4 | |||
| Case (n = 50) | PONase (U/mL): 272.6 ± 118.8 | ||
| AREase (U/mL): 197.3 ± 72.1 | |||
| Breast cancer | Control (n = 25) | PONase (U/mL):124.8 ± 21.7 | [308] |
| AREase (U/mL): 98.5 ± 18.8 | |||
| Case (n = 25) | PONase (U/mL):42.9 ± 7.9 | ||
| AREase (U/mL): 54.5 ± 11.5 | |||
| Control (n = 50) | PONase (U/mL):95.0 ± 30.3 | [318] | |
| AREase (U/mL): 52.1 ± 11.9 | |||
| Case (n = 50) | PONase (U/mL):65.0 ± 28.2 | ||
| AREase (U/mL): 29.0 ± 03.02 | |||
| Prostate cancer | Control (n = 20) | PONase (U/mL):92.1 ± 36.4 | [310] |
| AREase (U/mL): 51.1 ± 13.9 | |||
| Case (n = 25) | PONase (U/mL):67.2 ± 6.2 | ||
| AREase(U/mL): 31.0 ± 14.6 | |||
| Control (n = 40) | PONase (U/L): 76.5 ± 46.5 | [318] | |
| AREase (kU/L): 135.5 ± 65.7 | |||
| Case (n = 23) | PONase (U/L):103.8 ± 64.7 | ||
| AREase (kU/L): 136.7 ± 59.9 | |||
| Lung cancer | Control (n = 25) | PONase (U/mL):124.8 ± 21.7 | [157] |
| AREase (U/mL): 98.5 ± 18.8 | |||
| Case (n = 45) | PONase (U/mL):67.6 ± 22.0 | ||
| AREase (U/mL): 65.6 ± 27.1 | |||
| Control (n = 39) | PONase (U/mL): 395.8 ± 116.6 | [308] | |
| AREase (U/mL): 167.7 ± 45 | |||
| Case (n = 39) | PONase (U/mL):252.7 ± 104.4 | ||
| AREase (U/mL): 137.4 ± 57.6 | |||
| Control (n = 12) | PONase (U/mL): 94.6 ± 24.2 | [318] | |
| AREase (U/mL): 54.7 ± 05.3 | |||
| Case (n = 12) | PONase (U/mL):70.2 ± 12.4 | ||
| AREase (U/mL): 36.4 ± 02.3 | |||
| Non-Hodgkin lymphoma | Control (n = 10) | PONase (U/mL): 89.3 ± 32.4 | [318] |
| AREase (U/mL): 51.1 ± 12.7 | |||
| Case (n = 10) | PONase (U/mL):62.2 ± 0.7 | ||
| AREase (U/mL): 37.0 ± 0.6 | |||
| Central Nervous System Cancer | Control (n = 50) | PONase (U/L): 218.8 ± 144.8 | [319] |
| Case (n = 42) | PONase (U/L):73.2 ± 62.0 | ||
| Control (n = 25) | PONase (U/L): 131.1 ± 14.0 | ||
| Case (n = 25) | PONase (U/L):70.6 ± 17.0 |
| SNP | Genotype | Cancer Type | Population | Reference |
|---|---|---|---|---|
| L55M | 55M | Breast cancer | Caucasian | [325] |
| L55M Q192R | 55M 192QQ | Breast cancer | Caucasian Italian | [31] |
| L55M | 55M | Breast cancer | Egyptian | [326] |
| L55M | 55M | Breast cancer | Malaysian | [327] |
| Q192R | 192QQ | Lymphoma | Spain | [328] |
| Q192R | 192QQ | Glioma | Chinese | [329] |
| L55M | 55M | Leukemia | Brazilian | [330] |
| Q192R | 192QQ | Lymphohaematopoietic cancer | Greeks | [331] |
| Q192R | 192QQ | Lung cancer | Turkish | [332] |
| L55M Q192R | 55M 192QQ | Ovarian cancer | Hawaii | [333] |
| Q192R | 192QQ | Osteosarcoma | Caucasian Turkish | [334] |
| L55M Q192R | 55M 192QQ | Brain tumor | Spain | [335] |
| Q192R | 192RR | Bladder cancer | Turkish | [336] |
| Q192R | Hodgkin’s lymphoma | Caucasian | [337] | |
| L55M | 55M | Breast cancer | Caucasian Italian | [31] |
| Q192R | RR | Brain tumor | Turkish | [319] |
| Q192R | 192RR | Ovarian cancer | Turkish | [338] |
| C108T | Brain tumor | Mixed | [339] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Medina-Díaz, I.M.; Ponce-Ruíz, N.; Rojas-García, A.E.; Zambrano-Zargoza, J.F.; Bernal-Hernández, Y.Y.; González-Arias, C.A.; Barrón-Vivanco, B.S.; Herrera-Moreno, J.F. The Relationship between Cancer and Paraoxonase 1. Antioxidants 2022, 11, 697. https://doi.org/10.3390/antiox11040697
Medina-Díaz IM, Ponce-Ruíz N, Rojas-García AE, Zambrano-Zargoza JF, Bernal-Hernández YY, González-Arias CA, Barrón-Vivanco BS, Herrera-Moreno JF. The Relationship between Cancer and Paraoxonase 1. Antioxidants. 2022; 11(4):697. https://doi.org/10.3390/antiox11040697
Chicago/Turabian StyleMedina-Díaz, Irma Martha, Néstor Ponce-Ruíz, Aurora Elizabeth Rojas-García, José Francisco Zambrano-Zargoza, Yael Y. Bernal-Hernández, Cyndia Azucena González-Arias, Briscia S. Barrón-Vivanco, and José Francisco Herrera-Moreno. 2022. "The Relationship between Cancer and Paraoxonase 1" Antioxidants 11, no. 4: 697. https://doi.org/10.3390/antiox11040697
APA StyleMedina-Díaz, I. M., Ponce-Ruíz, N., Rojas-García, A. E., Zambrano-Zargoza, J. F., Bernal-Hernández, Y. Y., González-Arias, C. A., Barrón-Vivanco, B. S., & Herrera-Moreno, J. F. (2022). The Relationship between Cancer and Paraoxonase 1. Antioxidants, 11(4), 697. https://doi.org/10.3390/antiox11040697