
A Whiff of Sulfur: One Wind a Day Keeps the Doctor Away

 ,
,  ,
,  and
and 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Inorganic VSCs
2.1. Hydrogen Sulfide (H2S)
2.2. Sulfur Dioxide (SO2)
3. Volatile Natural Products
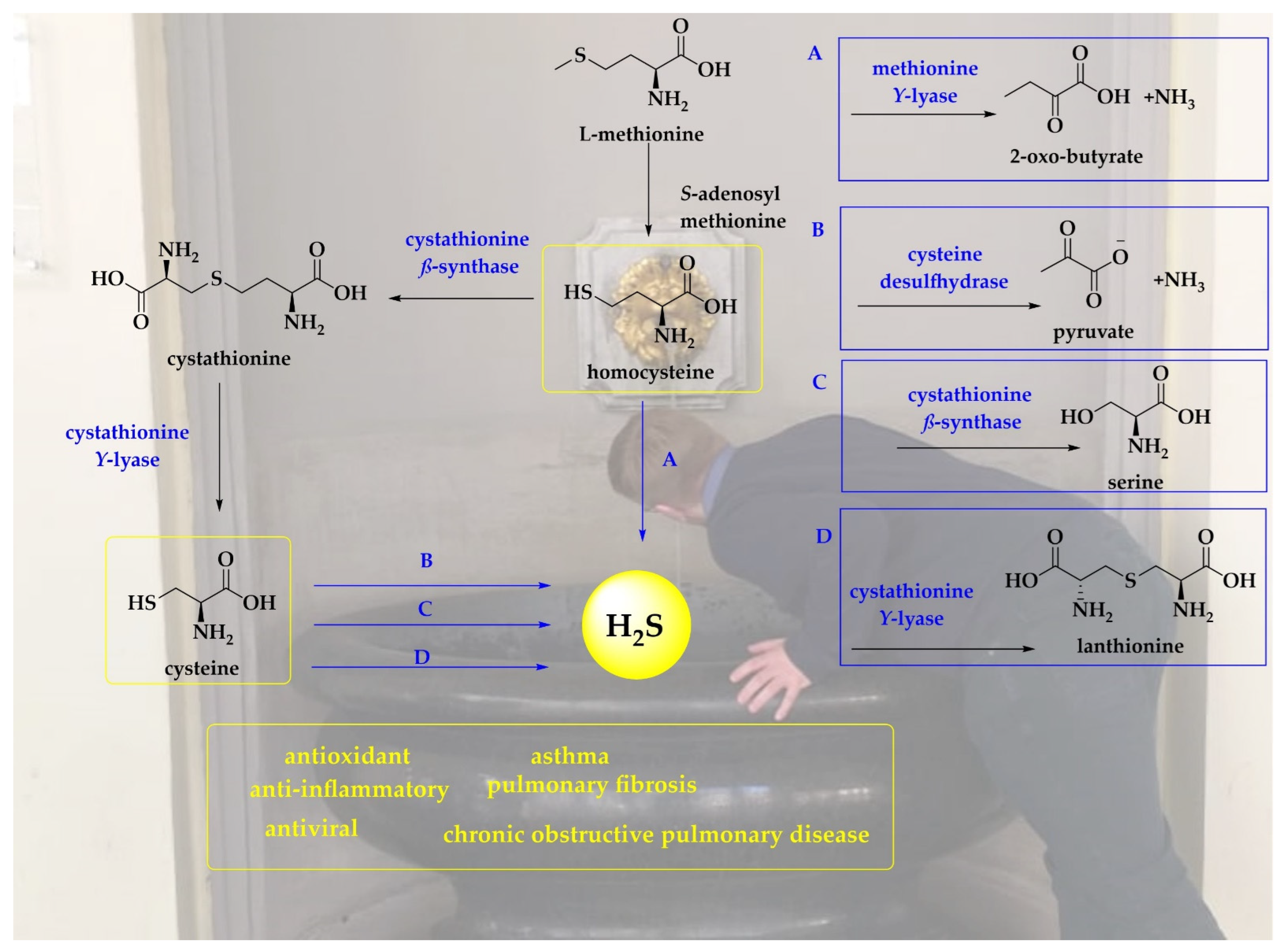
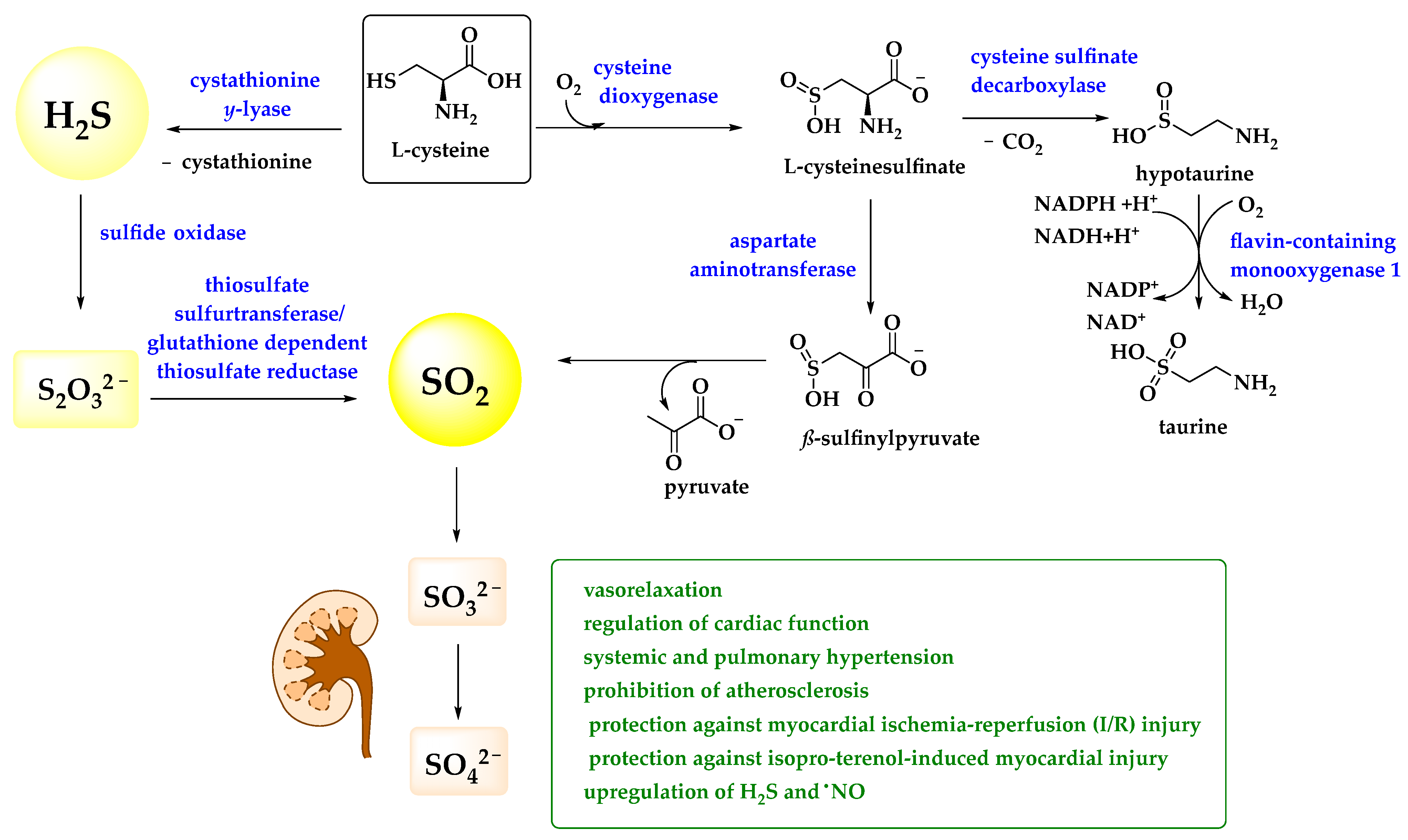
3.1. Metabolites of H2S
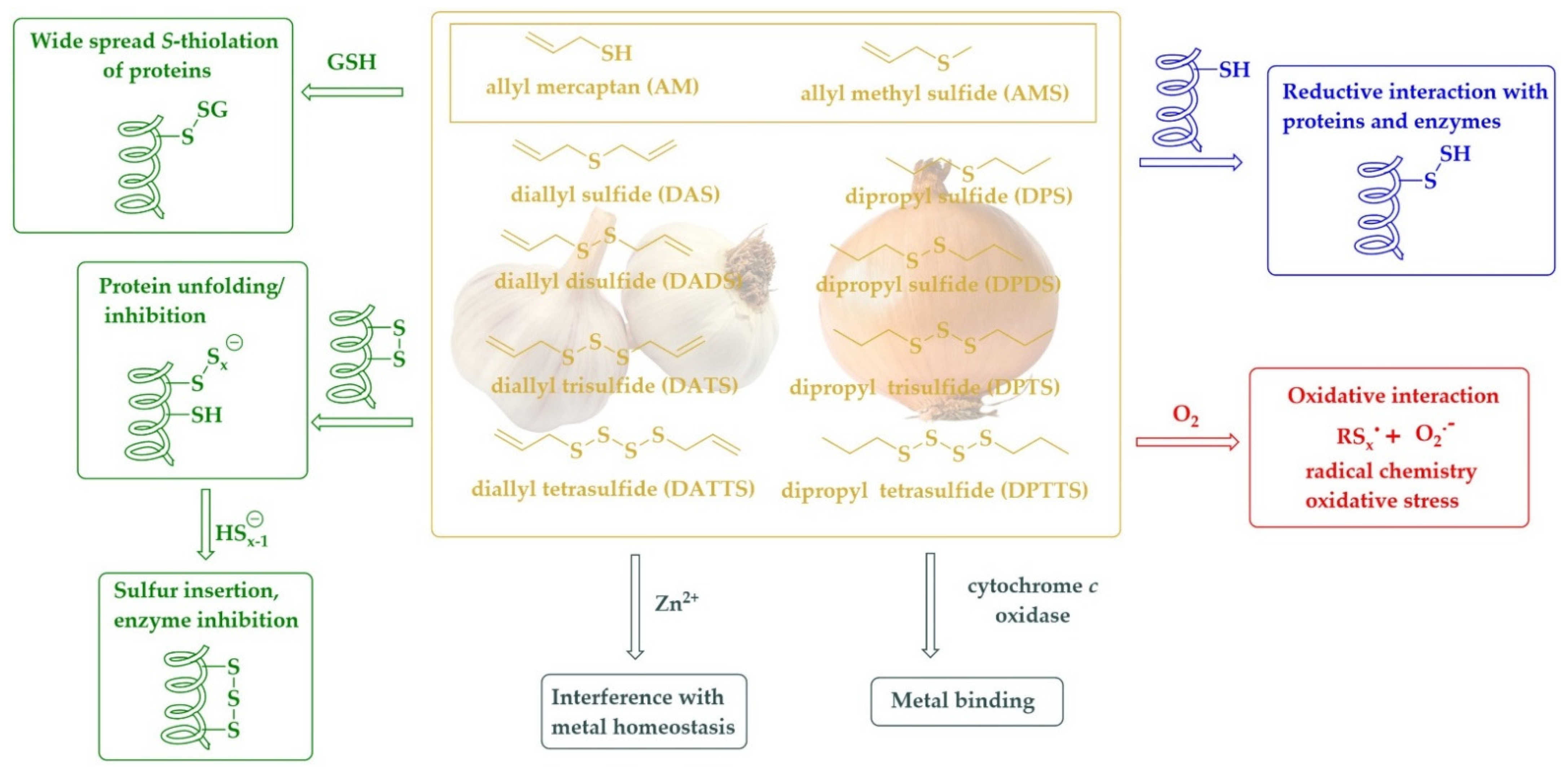
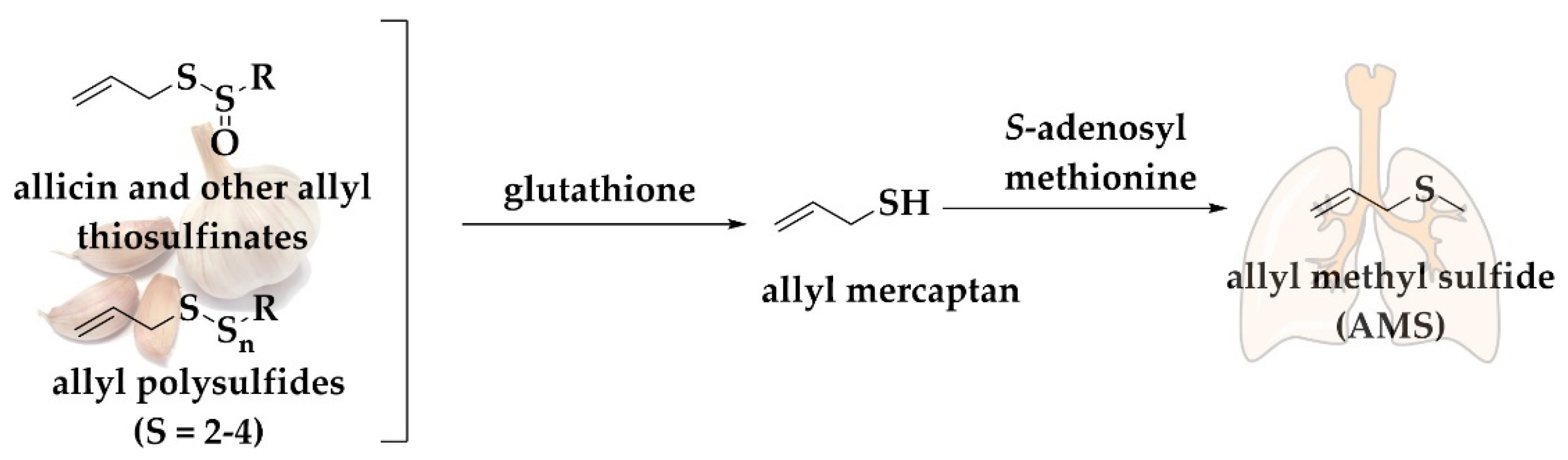
3.2. Simple Alkyl Sulfides
3.3. Metabolites
3.4. The Durian Fruit—A Treasure Chest of Public Annoyance and Volatile Sulfur Compounds
4. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Milito, A.; Brancaccio, M.; D’Argenio, G.; Castellano, I. Natural Sulfur-Containing Compounds: An Alternative Therapeutic Strategy against Liver Fibrosis. Cells 2019, 8, 1356. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Miękus, N.; Marszałek, K.; Podlacha, M.; Iqbal, A.; Puchalski, C.; Świergiel, A.H. Health Benefits of Plant-Derived Sulfur Compounds, Glucosinolates, and Organosulfur Compounds. Molecules 2020, 25, 3804. [Google Scholar] [CrossRef] [PubMed]
- Kalaras, M.D.; Richie, J.P.; Calcagnotto, A.; Beelman, R.B. Mushrooms: A Rich Source of the Antioxidants Ergothioneine and Glutathione. Food. Chem. 2017, 233, 429–433. [Google Scholar] [CrossRef]
- Ruhee, R.T.; Roberts, L.A.; Ma, S.; Suzuki, K. Organosulfur Compounds: A Review of Their Anti-Inflammatory Effects in Human Health. Front. Nutr. 2020, 7, 64. [Google Scholar] [CrossRef] [PubMed]
- Doleman, J.F.; Grisar, K.; Van Liedekerke, L.; Saha, S.; Roe, M.; Tapp, H.S.; Mithen, R.F. The Contribution of Alliaceous and Cruciferous Vegetables to Dietary Sulphur Intake. Food. Chem. 2017, 234, 38–45. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.-L.; Lin, S.-Y.; Du, H.-T.; Qin, L.; Lei, L.-M.; Chen, D. An Insight by Molecular Sensory Science Approaches to Contributions and Variations of the Key Odorants in Shiitake Mushrooms. Foods 2021, 10, 622. [Google Scholar] [CrossRef]
- Cao, X.; Cao, L.; Zhang, W.; Lu, R.; Bian, J.-S.; Nie, X. Therapeutic Potential of Sulfur-Containing Natural Products in Inflammatory Diseases. Pharmacol. Ther. 2020, 216, 107687. [Google Scholar] [CrossRef]
- Borlinghaus, J.; Foerster (née Reiter), J.; Kappler, U.; Antelmann, H.; Noll, U.; Gruhlke, M.C.H.; Slusarenko, A.J. Allicin, the Odor of Freshly Crushed Garlic: A Review of Recent Progress in Understanding Allicin’s Effects on Cells. Molecules 2021, 26, 1505. [Google Scholar] [CrossRef]
- Gruhlke, M.C.H.; Nicco, C.; Batteux, F.; Slusarenko, A.J. The Effects of Allicin, a Reactive Sulfur Species from Garlic, on a Selection of Mammalian Cell Lines. Antioxidants 2016, 6, 1. [Google Scholar] [CrossRef] [Green Version]
- Gruhlke, M.C.H.; Antelmann, H.; Bernhardt, J.; Kloubert, V.; Rink, L.; Slusarenko, A.J. The Human Allicin-Proteome: S-Thioallylation of Proteins by the Garlic Defence Substance Allicin and Its Biological Effects. Free Radic. Biol. Med. 2019, 131, 144–153. [Google Scholar] [CrossRef]
- Leontiev, R.; Hohaus, N.; Jacob, C.; Gruhlke, M.C.H.; Slusarenko, A.J. A Comparison of the Antibacterial and Antifungal Activities of Thiosulfinate Analogues of Allicin. Sci. Rep. 2018, 8, 6763. [Google Scholar] [CrossRef] [PubMed]
- Bhatwalkar, S.B.; Shukla, P.; Srivastava, R.K.; Mondal, R.; Anupam, R. Validation of Environmental Disinfection Efficiency of Traditional Ayurvedic Fumigation Practices. J. Ayurveda Integr. Med. 2019, 10, 203–206. [Google Scholar] [CrossRef] [PubMed]
- Anwar, A.; Gould, E.; Tinson, R.; Groom, M.; Hamilton, C.J. Think Yellow and Keep Green—Role of Sulfanes from Garlic in Agriculture. Antioxidants 2016, 6, 3. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yu, Y.; Hoffhines, A.J.; Moore, K.L.; Leary, J.A. Determination of the Sites of Tyrosine O-Sulfation in Peptides and Proteins. Nat. Methods 2007, 4, 583–588. [Google Scholar] [CrossRef]
- Nagy, P.; Beal, J.L.; Ashby, M.T. Thiocyanate Is an Efficient Endogenous Scavenger of the Phagocytic Killing Agent Hypobromous Acid. Chem. Res. Toxicol. 2006, 19, 587–593. [Google Scholar] [CrossRef]
- Bonifay, V.; Barrett, T.J.; Pattison, D.I.; Davies, M.J.; Hawkins, C.L.; Ashby, M.T. Tryptophan Oxidation in Proteins Exposed to Thiocyanate-Derived Oxidants. Arch. Biochem. Biophys. 2014, 564, 1–11. [Google Scholar] [CrossRef]
- Raciti, L.; Calabrò, R.S. Can Volcanic Trace Elements Facilitate Covid-19 Diffusion? A Hypothesis Stemming from the Mount Etna Area, Sicily. Med. Hypotheses 2020, 144, 110058. [Google Scholar] [CrossRef]
- Karagülle, M.Z.; Karagülle, M. Effects of Drinking Natural Hydrogen Sulfide (H2S) Waters: A Systematic Review of in Vivo Animal Studies. Int. J. Biometeorol. 2020, 64, 1011–1022. [Google Scholar] [CrossRef]
- Doujaiji, B.; Al-Tawfiq, J.A. Hydrogen Sulfide Exposure in an Adult Male. Ann. Saudi Med. 2010, 30, 76–80. [Google Scholar] [CrossRef]
- Bad Nenndorfer Schwefelquelle-Eine Der Stärksten in Europa Bad Nenndorf. Available online: https://www.badnenndorf.de/kur-and-gesundheit/unsere-heilmittel-moor-schwefel-sole/schwefel/ (accessed on 16 March 2022).
- Sokrateva, T.D.; Roussev, B.H.; Nashar, M.A.; Kiselova-Kaneva, Y.D.; Mihaylova, G.M.; Todorova, M.N.; Pasheva, M.G.; Tasinov, O.B.; Nazifova-Tasinova, N.F.; Vankova, D.G.; et al. Effects of Sulphur-Containing Mineral Water Intake on Oxidative Status and Markers for Inflammation in Healthy Subjects. Arch. Physiol. Biochem. 2021, 127, 327–336. [Google Scholar] [CrossRef]
- Physicochemical Analysis of Varna Basin Mineral Water-ProQuest. Available online: https://www.proquest.com/openview/4b6fab94d5ade3ac7a79409f75fcde9d/1?pq-origsite=gscholar&cbl=1536338 (accessed on 16 March 2022).
- Li, X.; O’Moore, L.; Song, Y.; Bond, P.L.; Yuan, Z.; Wilkie, S.; Hanzic, L.; Jiang, G. The Rapid Chemically Induced Corrosion of Concrete Sewers at High H2S Concentration. Water Res. 2019, 162, 95–104. [Google Scholar] [CrossRef] [PubMed]
- Braccia, D.J.; Jiang, X.; Pop, M.; Hall, A.B. The Capacity to Produce Hydrogen Sulfide (H2S) via Cysteine Degradation Is Ubiquitous in the Human Gut Microbiome. Front. Microbiol. 2021, 12, 705583. [Google Scholar] [CrossRef] [PubMed]
- Dordević, D.; Jančíková, S.; Vítězová, M.; Kushkevych, I. Hydrogen Sulfide Toxicity in the Gut Environment: Meta-Analysis of Sulfate-Reducing and Lactic Acid Bacteria in Inflammatory Processes. J. Adv. Res. 2021, 27, 55–69. [Google Scholar] [CrossRef] [PubMed]
- Kolluru, G.K.; Shen, X.; Bir, S.C.; Kevil, C.G. Hydrogen Sulfide Chemical Biology: Pathophysiological Roles and Detection. Nitric Oxide 2013, 35, 5–20. [Google Scholar] [CrossRef] [Green Version]
- Cao, X.; Ding, L.; Xie, Z.; Yang, Y.; Whiteman, M.; Moore, P.K.; Bian, J.-S. A Review of Hydrogen Sulfide Synthesis, Metabolism, and Measurement: Is Modulation of Hydrogen Sulfide a Novel Therapeutic for Cancer? Antioxid. Redox Sign. 2019, 31, 1–38. [Google Scholar] [CrossRef]
- Tomasova, L.; Konopelski, P.; Ufnal, M. Gut Bacteria and Hydrogen Sulfide: The New Old Players in Circulatory System Homeostasis. Molecules 2016, 21, 1558. [Google Scholar] [CrossRef] [PubMed]
- Suarez, F.; Furne, J.; Springfield, J.; Levitt, M. Insights into Human Colonic Physiology Obtained from the Study of Flatus Composition. Am. J. Physiol. Gastroint Liver Physiol. 1997, 272, G1028–G1033. [Google Scholar] [CrossRef]
- Furne, J.; Saeed, A.; Levitt, M.D. Whole Tissue Hydrogen Sulfide Concentrations Are Orders of Magnitude Lower than Presently Accepted Values. Am. J. Physiol. Reg. Integ. Comp. Physiol. 2008, 295, R1479–R1485. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Buret, A.G.; Allain, T.; Motta, J.-P.; Wallace, J.L. Effects of Hydrogen Sulfide on the Microbiome: From Toxicity to Therapy. Antioxid. Redox Sign. 2021, 36, 211–219. [Google Scholar] [CrossRef]
- Wallace, J.L.; Wang, R. Hydrogen Sulfide-Based Therapeutics: Exploiting a Unique but Ubiquitous Gasotransmitter. Nat. Rev. Drug Discov. 2015, 14, 329–345. [Google Scholar] [CrossRef]
- Blachier, F.; Beaumont, M.; Kim, E. Cysteine-Derived Hydrogen Sulfide and Gut Health: A Matter of Endogenous or Bacterial Origin. Curr. Opin. Clin. Nutr. Metab. Care 2019, 22, 68–75. [Google Scholar] [CrossRef] [PubMed]
- Searcy, D.G.; Lee, S.H. Sulfur Reduction by Human Erythrocytes. J. Exp. Zool. 1998, 282, 310–322, Wiley Online Library. Available online: https://onlinelibrary.wiley.com/doi/10.1002/(SICI)1097-010X(19981015)282:3%3C310::AID-JEZ4%3E3.0.CO;2-P (accessed on 21 March 2022). [CrossRef]
- Benavides, G.A.; Squadrito, G.L.; Mills, R.W.; Patel, H.D.; Isbell, T.S.; Patel, R.P.; Darley-Usmar, V.M.; Doeller, J.E.; Kraus, D.W. Hydrogen Sulfide Mediates the Vasoactivity of Garlic. Proc. Nat. Acad. Sci. USA 2007, 104, 17977–17982. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- di Masi, A.; Ascenzi, P. H2S: A “Double Face” Molecule in Health and Disease. BioFactors 2013, 39, 186–196. [Google Scholar] [CrossRef]
- Dóka, É.; Pader, I.; Bíró, A.; Johansson, K.; Cheng, Q.; Ballagó, K.; Prigge, J.R.; Pastor-Flores, D.; Dick, T.P.; Schmidt, E.E.; et al. A Novel Persulfide Detection Method Reveals Protein Persulfide- and Polysulfide-Reducing Functions of Thioredoxin and Glutathione Systems. Sci. Adv. 2016, 2, e1500968. [Google Scholar] [CrossRef] [Green Version]
- Yang, J.; Minkler, P.; Grove, D.; Wang, R.; Willard, B.; Dweik, R.; Hine, C. Non-Enzymatic Hydrogen Sulfide Production from Cysteine in Blood Is Catalyzed by Iron and Vitamin B6. Commun. Biol. 2019, 2, 1–14. [Google Scholar] [CrossRef] [Green Version]
- Singh, S.; Banerjee, R. PLP-Dependent H2S Biogenesis. Biochim. Biophys. Acta (BBA) Proteins Proteom. 2011, 1814, 1518–1527. [Google Scholar] [CrossRef] [Green Version]
- Li, P.; Liu, H.; Shi, X.; Prokosch, V. Hydrogen Sulfide: Novel Endogenous and Exogenous Modulator of Oxidative Stress in Retinal Degeneration Diseases. Molecules 2021, 26, 2411. [Google Scholar] [CrossRef]
- Instagram, E.F.-W.-P. 52062 A.D.F. Elisenbrunnen. Available online: https://www.aachen-tourismus.de/en/discover/sights/details/elisenbrunnen/ (accessed on 4 April 2022).
- Li, Q.; Lancaster, J.R. Chemical Foundations of Hydrogen Sulfide Biology. Nitric Oxide 2013, 35, 21–24. [Google Scholar] [CrossRef] [Green Version]
- Hyšpler, R.; Tichá, A.; Indrová, M.; Zadák, Z.; Hyšplerová, L.; Gasparič, J.; Churáček, J. A Simple, Optimized Method for the Determination of Sulphide in Whole Blood by GC–MS as a Marker of Bowel Fermentation Processes. J. Chromatog. B 2002, 770, 255–259. [Google Scholar] [CrossRef]
- Ghasemi, A.; Zahedi Asl, S.; Mehrabi, Y.; Saadat, N.; Azizi, F. Serum Nitric Oxide Metabolite Levels in a General Healthy Population: Relation to Sex and Age. Life Sci. 2008, 83, 326–331. [Google Scholar] [CrossRef]
- Wang, R. Physiological Implications of Hydrogen Sulfide: A Whiff Exploration That Blossomed. Physiol. Rev. 2012, 92, 791–896. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Khattak, S.; Zhang, Q.-Q.; Sarfraz, M.; Muhammad, P.; Ngowi, E.E.; Khan, N.H.; Rauf, S.; Wang, Y.-Z.; Qi, H.-W.; Wang, D.; et al. The Role of Hydrogen Sulfide in Respiratory Diseases. Biomolecules 2021, 11, 682. [Google Scholar] [CrossRef] [PubMed]
- Suzuki, Y.; Saito, J.; Munakata, M.; Shibata, Y. Hydrogen Sulfide as a Novel Biomarker of Asthma and Chronic Obstructive Pulmonary Disease. Allergol. Int. 2021, 70, 181–189. [Google Scholar] [CrossRef]
- Bourgonje, A.R.; Offringa, A.K.; van Eijk, L.E.; Abdulle, A.E.; Hillebrands, J.-L.; van der Voort, P.H.J.; van Goor, H.; van Hezik, E.J. N-Acetylcysteine and Hydrogen Sulfide in Coronavirus Disease 2019. Antioxid. Redox Signal. 2021, 35, 1207–1225. [Google Scholar] [CrossRef]
- Citi, V.; Martelli, A.; Brancaleone, V.; Brogi, S.; Gojon, G.; Montanaro, R.; Morales, G.; Testai, L.; Calderone, V. Anti-Inflammatory and Antiviral Roles of Hydrogen Sulfide: Rationale for Considering H2S Donors in COVID-19 Therapy. Br. J. Pharmacol. 2020, 177, 4931–4941. [Google Scholar] [CrossRef]
- Yang, G. H2S as a Potential Defense against COVID-19? Am. J. Physiol. Cell Physiol. 2020, 319, C244–C249. [Google Scholar] [CrossRef] [PubMed]
- Dai, J.; Teng, X.; Jin, S.; Wu, Y. The Antiviral Roles of Hydrogen Sulfide by Blocking the Interaction between SARS-CoV-2 and Its Potential Cell Surface Receptors. Oxid. Med. Cell Longev. 2021, 2021, e7866992. [Google Scholar] [CrossRef]
- Xiao, Q.; Xiong, L.; Tang, J.; Li, L.; Li, L. Hydrogen Sulfide in Skin Diseases: A Novel Mediator and Therapeutic Target. Oxid. Med. Cell Longev. 2021, 2021, e6652086. [Google Scholar] [CrossRef]
- Beinert, H.; Holm, R.H.; Münck, E. Iron-Sulfur Clusters: Nature’s Modular, Multipurpose Structures. Science 1997, 277, 653–659. [Google Scholar] [CrossRef]
- Coavoy-Sánchez, S.A.; Costa, S.K.P.; Muscará, M.N. Hydrogen Sulfide and Dermatological Diseases. Br. J. Pharmacol. 2020, 177, 857–865. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Grennfelt, P.; Engleryd, A.; Forsius, M.; Hov, Ø.; Rodhe, H.; Cowling, E. Acid Rain and Air Pollution: 50 Years of Progress in Environmental Science and Policy. Ambio 2020, 49, 849–864. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Duan, S.-M.; Xu, J.; Bai, Y.-J.; Ding, Y.; Kong, M.; Liu, H.-H.; Li, X.-Y.; Zhang, Q.-S.; Chen, H.-B.; Liu, L.-F.; et al. Sulfur Dioxide Residue in Sulfur-Fumigated Edible Herbs: The Fewer, the Safer? Food Chem. 2016, 192, 119–124. [Google Scholar] [CrossRef] [PubMed]
- Arapitsas, P.; Guella, G.; Mattivi, F. The Impact of SO2 on Wine Flavanols and Indoles in Relation to Wine Style and Age. Sci. Rep. 2018, 8, 858. [Google Scholar] [CrossRef] [Green Version]
- Du, S.; Jin, H.; Bu, D.; Zhao, X.; Geng, B.; Tang, C.; Du, J. Endogenously Generated Sulfur Dioxide and Its Vasorelaxant Effect in Rats. Acta Pharmacol. Sin. 2008, 29, 923–930. [Google Scholar] [CrossRef] [Green Version]
- Ji, A.J.; Savon, S.R.; Jacobsen, D.W. Determination of Total Serum Sulfite by HPLC with Fluorescence Detection. Clin. Chem. 1995, 41, 897–903. [Google Scholar] [CrossRef]
- Wang, W.; Wang, B. SO2 Donors and Prodrugs, and Their Possible Applications: A Review. Front. Chem. 2018, 6, 559. [Google Scholar] [CrossRef] [Green Version]
- Zhang, L.; Jin, H.; Song, Y.; Chen, S.Y.; Wang, Y.; Sun, Y.; Tang, C.; Du, J.; Huang, Y. Endogenous Sulfur Dioxide Is a Novel Inhibitor of Hypoxia-Induced Mast Cell Degranulation. J. Adv. Res. 2021, 29, 55–65. [Google Scholar] [CrossRef]
- Huang, Y.; Tang, C.; Du, J.; Jin, H. Endogenous Sulfur Dioxide: A New Member of Gasotransmitter Family in the Cardiovascular System. Oxid. Med. Cell Longev. 2016, 2016, 8961951. [Google Scholar] [CrossRef] [Green Version]
- Li, Y.; Feng, Y.; Ye, X.; Peng, H.; Du, J.; Yao, X.; Huang, Y.; Jin, H.; Du, J. Endogenous SO2 Controls Cell Apoptosis: The State-of-the-Art. Front. Cell Dev. Biol. 2021, 9, 729728. [Google Scholar] [CrossRef]
- Veeravalli, S.; Phillips, I.R.; Freire, R.T.; Varshavi, D.; Everett, J.R.; Shephard, E.A. Flavin-Containing Monooxygenase 1 Catalyzes the Production of Taurine from Hypotaurine. Drug Metab. Dispos. 2020, 48, 378–385. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, D.; Huang, Y.; Bu, D.; Liu, A.D.; Holmberg, L.; Jia, Y.; Tang, C.; Du, J.; Jin, H. Sulfur Dioxide Inhibits Vascular Smooth Muscle Cell Proliferation via Suppressing the Erk/MAP Kinase Pathway Mediated by CAMP/PKA Signaling. Cell Death Dis. 2014, 5, e1251. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, S.; Xu, Z.; Xia, J.; Qin, G.; Sang, N. Sulfur Dioxide Induces Apoptosis via Reactive Oxygen Species Generation in Rat Cardiomyocytes. Environ. Sci. Pollut. Res. Int. 2019, 26, 8758–8767. [Google Scholar] [CrossRef] [PubMed]
- Niu, M.; Han, Y.; Li, Q.; Zhang, J. Endogenous Sulfur Dioxide Regulates Hippocampal Neuron Apoptosis in Developing Epileptic Rats and Is Associated with the PERK Signaling Pathway. Neurosci. Lett. 2018, 665, 22–28. [Google Scholar] [CrossRef]
- Han, Y.; Yi, W.; Qin, J.; Zhao, Y.; Zhang, J.; Chang, X. Dose-Dependent Effect of Sulfur Dioxide on Brain Damage Induced by Recurrent Febrile Seizures in Rats. Neurosci. Lett. 2014, 563, 149–154. [Google Scholar] [CrossRef]
- Wenzel, A.; Grimm, C.; Samardzija, M.; Remé, C.E. Molecular Mechanisms of Light-Induced Photoreceptor Apoptosis and Neuroprotection for Retinal Degeneration. Progr. Retin. Eye Res. 2005, 24, 275–306. [Google Scholar] [CrossRef]
- Ma, H.; Huang, X.; Liu, Y.; Fan, Y. Sulfur Dioxide Attenuates LPS-Induced Acute Lung Injury via Enhancing Polymorphonuclear Neutrophil Apoptosis. Acta Pharmacol. Sin. 2012, 33, 983–990. [Google Scholar] [CrossRef] [Green Version]
- Giles, G.I.; Nasim, M.J.; Ali, W.; Jacob, C. The Reactive Sulfur Species Concept: 15 Years On. Antioxidants 2017, 6, 38. [Google Scholar] [CrossRef] [Green Version]
- Toohey, J.I. Sulfur Signaling: Is the Agent Sulfide or Sulfane? Anal. Biochem. 2011, 413, 1–7. [Google Scholar] [CrossRef]
- Nagy, P.; Winterbourn, C.C. Rapid Reaction of Hydrogen Sulfide with the Neutrophil Oxidant Hypochlorous Acid to Generate Polysulfides. Available online: https://pubs.acs.org/doi/full/10.1021/tx100266a (accessed on 9 May 2022).
- Kimura, H. Hydrogen Polysulfide (H2Sn) Signaling along with Hydrogen Sulfide (H2S) and Nitric Oxide (NO). J. Neural. Transm. 2016, 123, 1235–1245. [Google Scholar] [CrossRef]
- Greiner, R.; Pálinkás, Z.; Bäsell, K.; Becher, D.; Antelmann, H.; Nagy, P.; Dick, T.P. Polysulfides Link H2S to Protein Thiol Oxidation. Antioxid. Redox Signal. 2013, 19, 1749–1765. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Koike, S.; Ogasawara, Y.; Shibuya, N.; Kimura, H.; Ishii, K. Polysulfide Exerts a Protective Effect against Cytotoxicity Caused by T-Buthylhydroperoxide through Nrf2 Signaling in Neuroblastoma Cells. FEBS Lett. 2013, 587, 3548–3555. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jarosz, A.P.; Wei, W.; Gauld, J.W.; Auld, J.; Özcan, F.; Aslan, M.; Mutus, B. Glyceraldehyde 3-Phosphate Dehydrogenase (GAPDH) Is Inactivated by S-Sulfuration In Vitro. Free Radic. Biol. Med. 2015, 89, 512–521. [Google Scholar] [CrossRef] [PubMed]
- Jackson, M.R.; Melideo, S.L.; Jorns, M.S. Human Sulfide:Quinone Oxidoreductase Catalyzes the First Step in Hydrogen Sulfide Metabolism and Produces a Sulfane Sulfur Metabolite. Biochemistry 2012, 51, 6804–6815. [Google Scholar] [CrossRef]
- Libiad, M.; Yadav, P.K.; Vitvitsky, V.; Martinov, M.; Banerjee, R. Organization of the Human Mitochondrial Hydrogen Sulfide Oxidation Pathway. J. Biol. Chem. 2014, 289, 30901–30910. [Google Scholar] [CrossRef] [Green Version]
- Yadav, P.K.; Martinov, M.; Vitvitsky, V.; Seravalli, J.; Wedmann, R.; Filipovic, M.R.; Banerjee, R. Biosynthesis and Reactivity of Cysteine Persulfides in Signaling. J. Am. Chem. Soc. 2016, 138, 289–299. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ono, K.; Akaike, T.; Sawa, T.; Kumagai, Y.; Wink, D.A.; Tantillo, D.J.; Hobbs, A.J.; Nagy, P.; Xian, M.; Lin, J.; et al. Redox Chemistry and Chemical Biology of H2S, Hydropersulfides, and Derived Species: Implications of Their Possible Biological Activity and Utility. Free Radic. Biol. Med. 2014, 77, 82–94. [Google Scholar] [CrossRef] [Green Version]
- Ida, T.; Sawa, T.; Ihara, H.; Tsuchiya, Y.; Watanabe, Y.; Kumagai, Y.; Suematsu, M.; Motohashi, H.; Fujii, S.; Matsunaga, T.; et al. Reactive Cysteine Persulfides and S-Polythiolation Regulate Oxidative Stress and Redox Signaling. Proc. Natl. Acad. Sci. USA 2014, 111, 7606–7611. [Google Scholar] [CrossRef] [Green Version]
- Paulsen, C.E.; Carroll, K.S. Cysteine-Mediated Redox Signaling: Chemistry, Biology, and Tools for Discovery. Chem. Rev. 2013, 113, 4633–4679. [Google Scholar] [CrossRef]
- Carballal, S.; Radi, R.; Kirk, M.C.; Barnes, S.; Freeman, B.A.; Alvarez, B. Sulfenic Acid Formation in Human Serum Albumin by Hydrogen Peroxide and Peroxynitrite. Biochemistry 2003, 42, 9906–9914. [Google Scholar] [CrossRef]
- Zhang, D.; Macinkovic, I.; Devarie-Baez, N.O.; Pan, J.; Park, C.-M.; Carroll, K.S.; Filipovic, M.R.; Xian, M. Detection of Protein S-Sulfhydration by a Tag-Switch Technique. Angew. Chem. Int. Ed. Engl. 2014, 53, 575–581. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Talipov, M.R.; Timerghazin, Q.K. Protein Control of S-Nitrosothiol Reactivity: Interplay of Antagonistic Resonance Structures. J. Phys. Chem. B 2013, 117, 1827–1837. [Google Scholar] [CrossRef] [PubMed]
- Miljkovic, J.L.; Kenkel, I.; Ivanović-Burmazović, I.; Filipovic, M.R. Generation of HNO and HSNO from Nitrite by Heme-Iron-Catalyzed Metabolism with H2S. Angew. Chem. Int. Ed. Engl. 2013, 52, 12061–12064. [Google Scholar] [CrossRef] [PubMed]
- Lajin, B.; Francesconi, K.A. The Hydrogen Sulfide Metabolite Trimethylsulfonium Is Found in Human Urine. Sci. Rep. 2016, 6, 27038. [Google Scholar] [CrossRef] [Green Version]
- Snel, J.; Burgering, M.; Smit, B.; Noordman, W.; Tangerman, A.; Winkel, E.G.; Kleerebezem, M. Volatile Sulphur Compounds in Morning Breath of Human Volunteers. Arch. Oral. Biol. 2011, 56, 29–34. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abdullah, M.A.; Alasqah, M.; Sanaa, M.S.; Gufran, K. The Relationship between Volatile Sulfur Compounds and the Severity of Chronic Periodontitis: A Cross-Sectional Study. J. Pharm. Bioallied Sci. 2020, 12, S268–S273. [Google Scholar] [CrossRef]
- Yoo, S.H.; Jung, H.S.; Sohn, W.S.; Kim, B.H.; Ku, B.H.; Kim, Y.S.; Park, S.W.; Hahm, K.-B. Volatile Sulfur Compounds as a Predictor for Esophagogastroduodenal Mucosal Injury. Gut Liver 2008, 2, 113–118. [Google Scholar] [CrossRef]
- Guan, X.; Wu, P.; Wang, S.; Zhang, J.; Shen, Z.; Luo, H.; Chen, H.; Long, L.; Chen, J.; Wang, F. Dimethyl Sulfide Protects against Oxidative Stress and Extends Lifespan via a Methionine Sulfoxide Reductase A-dependent Catalytic Mechanism. Aging Cell 2017, 16, 226–236. [Google Scholar] [CrossRef]
- Lim, J.C.; You, Z.; Kim, G.; Levine, R.L. Methionine Sulfoxide Reductase A Is a Stereospecific Methionine Oxidase. Proc. Nat. Acad. Sci. USA 2011, 108, 10472–10477. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Anwar, A.; Burkholz, T.; Scherer, C.; Abbas, M.; Lehr, C.-M.; Diederich, M.; Jacob, C. Naturally Occurring Reactive Sulfur Species, Their Activity against Caco-2 Cells, and Possible Modes of Biochemical Action. J. Sulfur. Chem. 2008, 29, 251–268. [Google Scholar] [CrossRef]
- Grman, M.; Nasim, M.J.; Leontiev, R.; Misak, A.; Jakusova, V.; Ondrias, K.; Jacob, C. Inorganic Reactive Sulfur-Nitrogen Species: Intricate Release Mechanisms or Cacophony in Yellow, Blue and Red? Antioxidants 2017, 6, 14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nasim, M.J.; Denezhkin, P.; Sarfraz, M.; Leontiev, R.; Ney, Y.; Kharma, A.; Griffin, S.; Masood, M.I.; Jacob, C. The Small Matter of a Red Ox, a Particularly Sensitive Pink Cat, and the Quest for the Yellow Stone of Wisdom. Curr. Pharmacol. Rep. 2018, 4, 380–396. [Google Scholar] [CrossRef]
- Allah, D.R.; Schwind, L.; Asali, I.A.; Nasim, J.; Jacob, C.; Götz, C.; Montenarh, M. A Scent of Therapy: Synthetic Polysulfanes with Improved Physico-Chemical Properties Induce Apoptosis in Human Cancer Cells. Int. J. Oncol. 2015, 47, 991–1000. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Czepukojc, B.; Leroch, M.; Salm, F.; Viswanathan, U.M.; Burkholz, T.; Hahn, M.; Jacob, C. Antifungal Activity of Tetrasulfanes against Botrytis Cinerea. Nat. Prod. Commun. 2013, 8, 1599–1603. [Google Scholar] [CrossRef] [Green Version]
- Marut, W.; Jamier, V.; Kavian, N.; Servettaz, A.; Winyard, P.G.; Eggleton, P.; Anwar, A.; Nicco, C.; Jacob, C.; Chéreau, C.; et al. The Natural Organosulfur Compound Dipropyltetrasulfide Prevents HOCl-Induced Systemic Sclerosis in the Mouse. Arthritis Res. Ther. 2013, 15, R167. [Google Scholar] [CrossRef] [Green Version]
- Tyagi, S.; Lee, K.-J.; Shukla, P.; Chae, J.-C. Dimethyl Disulfide Exerts Antifungal Activity against Sclerotinia Minor by Damaging Its Membrane and Induces Systemic Resistance in Host Plants. Sci. Rep. 2020, 10, 6547. [Google Scholar] [CrossRef] [Green Version]
- Dandurishvili, N.; Toklikishvili, N.; Ovadis, M.; Eliashvili, P.; Giorgobiani, N.; Keshelava, R.; Tediashvili, M.; Vainstein, A.; Khmel, I.; Szegedi, E.; et al. Broad-Range Antagonistic Rhizobacteria Pseudomonas Fluorescens and Serratia Plymuthica Suppress Agrobacterium Crown Gall Tumours on Tomato Plants. J. Appl. Microbiol. 2011, 110, 341–352. [Google Scholar] [CrossRef]
- Lammers, A.; Lalk, M.; Garbeva, P. Air Ambulance: Antimicrobial Power of Bacterial Volatiles. Antibiotics 2022, 11, 109. [Google Scholar] [CrossRef]
- Paladin®EC Soil Fumigant, Active Ingredient Dimethyl Disulfide (DMDS), Drip Application. Available online: https://www.arkema.com/global/en/products/product-finder/product-range/thiochemicals/dmds-for-agricultural-soil-fumig/ (accessed on 23 March 2022).
- Chernin, L.; Toklikishvili, N.; Ovadis, M.; Kim, S.; Ben-Ari, J.; Khmel, I.; Vainstein, A. Quorum-Sensing Quenching by Rhizobacterial Volatiles. Environ. Microbiol. Rep. 2011, 3, 698–704. [Google Scholar] [CrossRef]
- Tang, Y.; Li, F.; Gu, D.; Wang, W.; Huang, J.; Jiao, X. Antimicrobial Effect and the Mechanism of Diallyl Trisulfide against Campylobacter Jejuni. Antibiotics 2021, 10, 246. [Google Scholar] [CrossRef]
- Gong, X.; Su, X.; Liu, H. Diallyl Trisulfide, the Antifungal Component of Garlic Essential Oil and the Bioactivity of Its Nanoemulsions Formed by Spontaneous Emulsification. Molecules 2021, 26, 7186. [Google Scholar] [CrossRef] [PubMed]
- Xia, S.-L.; Ma, Z.-Y.; Wang, B.; Gao, F.; Yi, C.-G.; Zhou, X.-X.; Guo, S.-Y.; Zhou, L. In Vitro Anti-Synovial Sarcoma Effect of Diallyl Trisulfide and MRNA Profiling. Gene 2022, 816, 146172. [Google Scholar] [CrossRef] [PubMed]
- Rao, P.; Midde, N.M.; Miller, D.D.; Chauhan, S.; Kumar, A.; Kumar, S. Diallyl Sulfide: Potential Use in Novel Therapeutic Interventions in Alcohol, Drugs, and Disease Mediated Cellular Toxicity by Targeting Cytochrome P450 2E1. Curr. Drug Metab. 2015, 16, 486–503. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ecospray|Ecospray Is the Pioneer of Internationally Approved and Patented Crop Protection Products Formulated Using a 100% Safe, Natural and Sustainable Technology Derived from Garlic. Available online: https://ecospray.com/ (accessed on 23 March 2022).
- Zhou, W.; Zou, X.; Lu, Y.; Xia, L.; Huang, C.; Shen, C.; Chen, X.; Chu, Y. Characterization of Volatiles in Garlic and in Exhaled Breath after Garlic Ingestion by On-Line Atmospheric Pressure Photoionization Quadrupole Time-of-Flight Mass Spectrometry. Food Sci. Technol. Res. 2017, 23, 613–620. [Google Scholar] [CrossRef] [Green Version]
- Taucher, J.; Hansel, A.; Jordan, A.; Lindinger, W. Analysis of Compounds in Human Breath after Ingestion of Garlic Using Proton-Transfer-Reaction Mass Spectrometry. J. Agric. Food Chem. 1996, 44, 3778–3782. [Google Scholar] [CrossRef]
- Sato, S.; Sekine, Y.; Kakumu, Y.; Hiramoto, T. Measurement of Diallyl Disulfide and Allyl Methyl Sulfide Emanating from Human Skin Surface and Influence of Ingestion of Grilled Garlic. Sci. Rep. 2020, 10, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Laakso, I.; Seppänen-Laakso, T.; Hiltunen, R.; Müller, B.; Jansen, H.; Knobloch, K. Volatile Garlic Odor Components: Gas Phases and Adsorbed Exhaled Air Analysed by Headspace Gas Chromatography-Mass Spectrometry. Planta Med. 1989, 55, 257–261. [Google Scholar] [CrossRef]
- Lawson, L.D.; Wang, Z.J. Allicin and Allicin-Derived Garlic Compounds Increase Breath Acetone through Allyl Methyl Sulfide: Use in Measuring Allicin Bioavailability. J. Agric. Food Chem. 2005, 53, 1974–1983. [Google Scholar] [CrossRef]
- Lawson, L.D.; Hunsaker, S.M. Allicin Bioavailability and Bioequivalence from Garlic Supplements and Garlic Foods. Nutrients 2018, 10, 812. [Google Scholar] [CrossRef] [Green Version]
- Kuttan, G.; Kuttan, R. Effect of Diallyl Sulphide, Diallyl Disulphide, and Allyl Methyl Sulphide on the Inhibition of Lung Metastasis of B16F-10 Melanoma Cells in Mice. J. Clin. Biochem. Nutr. 1999, 27, 131–139. [Google Scholar] [CrossRef] [Green Version]
- Sinha, A.K.; Farooqui, S.A.; Sharma, A.; Mishra, A.; Verma, V. Reactivity of Allyl Methyl Sulphide, the in-Vitro Metabolite of Garlic, with Some Amino Acids and with Phospholipid Involved in Viral Infections. J. Biomol. Struct. Dyn. 2022, 40, 565–571. [Google Scholar] [CrossRef] [PubMed]
- Becker, P.M.; van Wikselaar, P.G.; Mul, M.F.; Pol, A.; Engel, B.; Wijdenes, J.W.; van der Peet-Schwering, C.M.C.; Wisselink, H.J.; Stockhofe-Zurwieden, N. Actinobacillus Pleuropneumoniae Is Impaired by the Garlic Volatile Allyl Methyl Sulfide (AMS) in Vitro and in-Feed Garlic Alleviates Pleuropneumonia in a Pig Model. Vet Microbiol. 2012, 154, 316–324. [Google Scholar] [CrossRef] [PubMed]
- Yoshioka, Y.; Matsumura, S.; Morimoto, M.; Takemoto, Y.; Kishi, C.; Moriyama, T.; Zaima, N. Inhibitory Activities of Sulfur Compounds in Garlic Essential Oil against Alzheimer’s Disease-Related Enzymes and Their Distribution in the Mouse Brain. J. Agric. Food Chem. 2021, 69, 10163–10173. [Google Scholar] [CrossRef] [PubMed]
- Lietzow, J. Biologically Active Compounds in Mustard Seeds: A Toxicological Perspective. Foods 2021, 10, 2089. [Google Scholar] [CrossRef] [PubMed]
- Rahman, M.; Khatun, A.; Liu, L.; Barkla, B.J. Brassicaceae Mustards: Traditional and Agronomic Uses in Australia and New Zealand. Molecules 2018, 23, 231. [Google Scholar] [CrossRef] [Green Version]
- Striegel, L.; Chebib, S.; Dumler, C.; Lu, Y.; Huang, D.; Rychlik, M. Durian Fruits Discovered as Superior Folate Sources. Front. Nutr. 2018, 5, 114. [Google Scholar] [CrossRef] [Green Version]
- Husin, N.A.; Rahman, S.; Karunakaran, R.; Bhore, S.J. A Review on the Nutritional, Medicinal, Molecular and Genome Attributes of Durian (Durio zibethinus L.), the King of Fruits in Malaysia. Bioinformation 2018, 14, 265–270. [Google Scholar] [CrossRef]
- Weenen, H.; Koolhaas, W.E.; Apriyantono, A. Sulfur-Containing Volatiles of Durian Fruits (Durio zibethinus Murr.). J. Agric. Food Chem. 1996, 44, 3291–3293. [Google Scholar] [CrossRef]
- Aziz, N.A.A.; Mhd Jalil, A.M. Bioactive Compounds, Nutritional Value, and Potential Health Benefits of Indigenous Durian (Durio zibethinus Murr.): A Review. Foods 2019, 8, 96. [Google Scholar] [CrossRef] [Green Version]





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tiganescu, E.; Lämmermann, M.A.; Ney, Y.; Abdin, A.Y.; Nasim, M.J.; Jacob, C. A Whiff of Sulfur: One Wind a Day Keeps the Doctor Away. Antioxidants 2022, 11, 1036. https://doi.org/10.3390/antiox11061036
Tiganescu E, Lämmermann MA, Ney Y, Abdin AY, Nasim MJ, Jacob C. A Whiff of Sulfur: One Wind a Day Keeps the Doctor Away. Antioxidants. 2022; 11(6):1036. https://doi.org/10.3390/antiox11061036
Chicago/Turabian StyleTiganescu, Eduard, Markus Alexander Lämmermann, Yannick Ney, Ahmad Yaman Abdin, Muhammad Jawad Nasim, and Claus Jacob. 2022. "A Whiff of Sulfur: One Wind a Day Keeps the Doctor Away" Antioxidants 11, no. 6: 1036. https://doi.org/10.3390/antiox11061036
APA StyleTiganescu, E., Lämmermann, M. A., Ney, Y., Abdin, A. Y., Nasim, M. J., & Jacob, C. (2022). A Whiff of Sulfur: One Wind a Day Keeps the Doctor Away. Antioxidants, 11(6), 1036. https://doi.org/10.3390/antiox11061036