
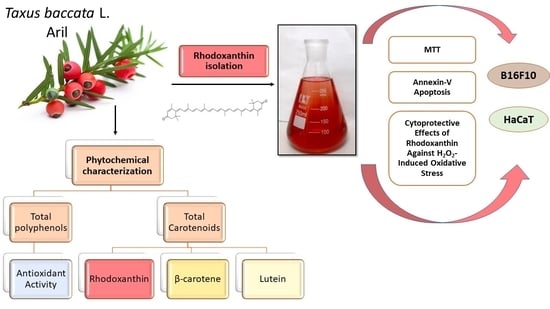
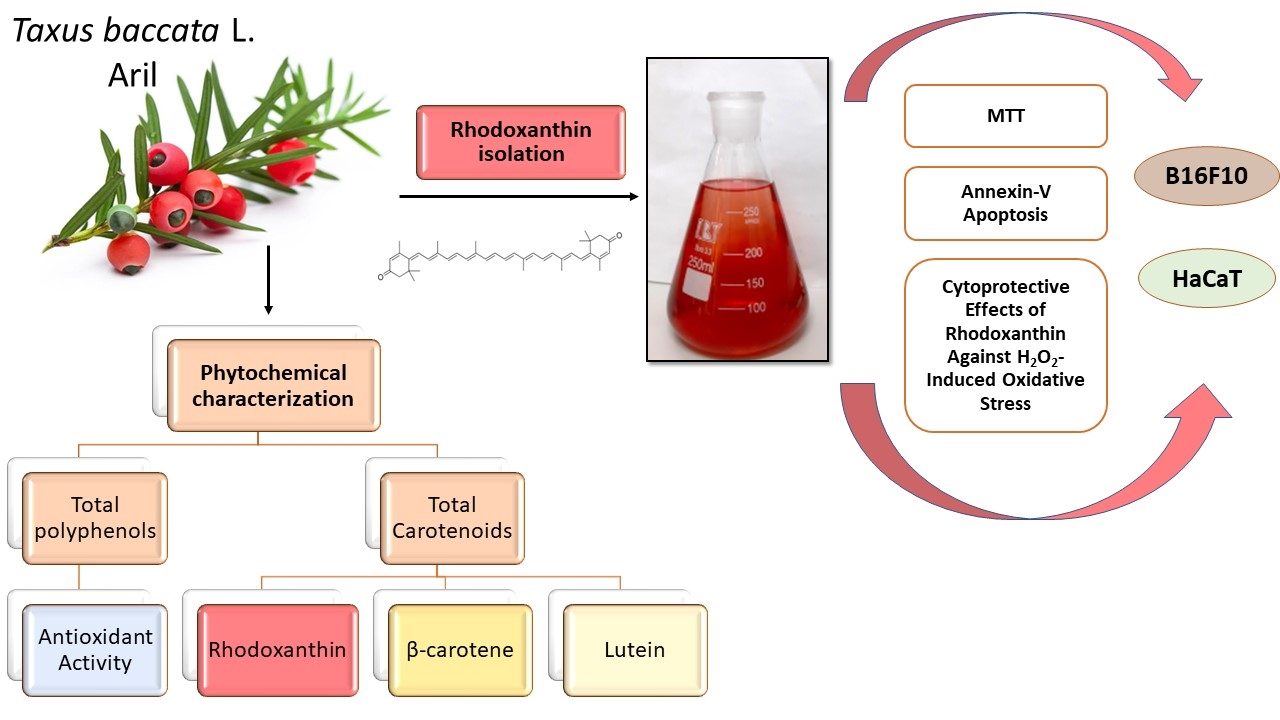
Phytochemical Characterization of Taxus baccata L. Aril with Emphasis on Evaluation of the Antiproliferative and Pro-Apoptotic Activity of Rhodoxanthin

 ,
, 
 ,
, 
 ,
, 

Abstract
:
1. Introduction
2. Materials and Methods
2.1. Plant Samples and Growing Conditions
2.2. Chemicals and Reagents
2.3. Determination of Carotenoid Composition
2.3.1. Preparation of Extract
2.3.2. Total Carotenoid Content
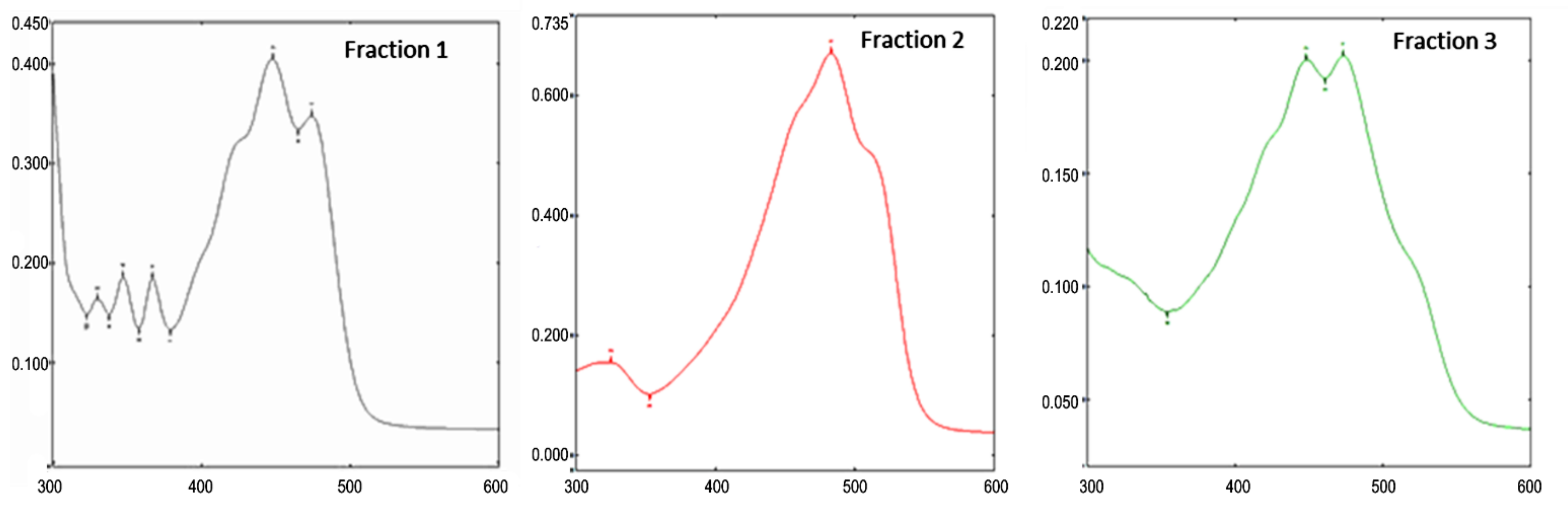
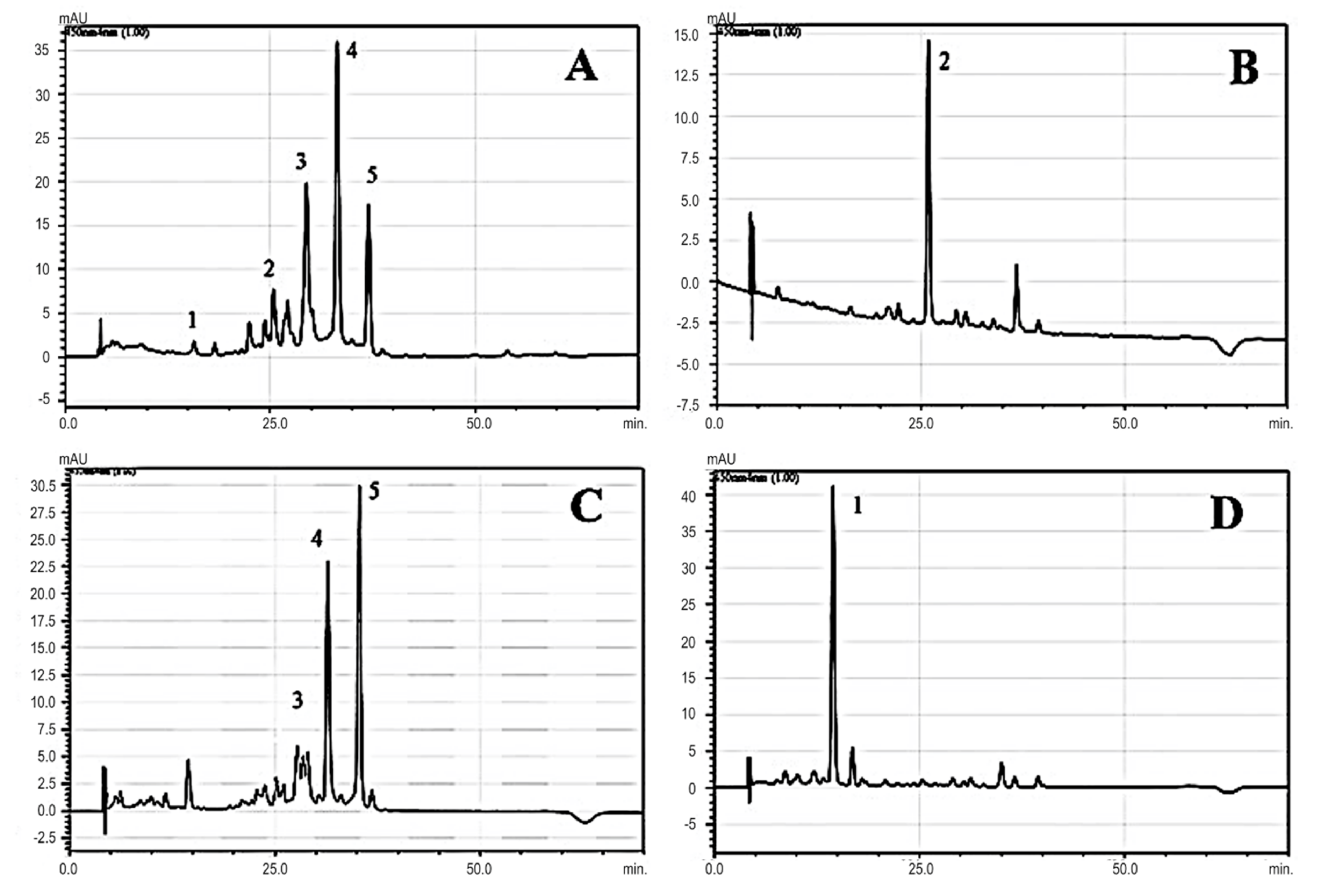
2.3.3. Carotenoids Analysis by Open-Column Chromatography
2.3.4. Carotenoids Analysis by RP–PAD–HPLC
2.4. Total and Individual Content of Phenolic Compounds
2.4.1. Preparation of Extract
2.4.2. Total Phenolic Content
2.4.3. Total Flavonoid Content
2.4.4. HPLC-DAD-ESI-MS Analysis
2.5. Antioxidant Activity
2.5.1. Determination of FRAP Scavenging Activity
2.5.2. Determination of DPPH Scavenging Activity
2.6. Cell Cultures and Cytotoxic Assay
2.6.1. Cell Lines and Cultures
2.6.2. Cell Viability Assay
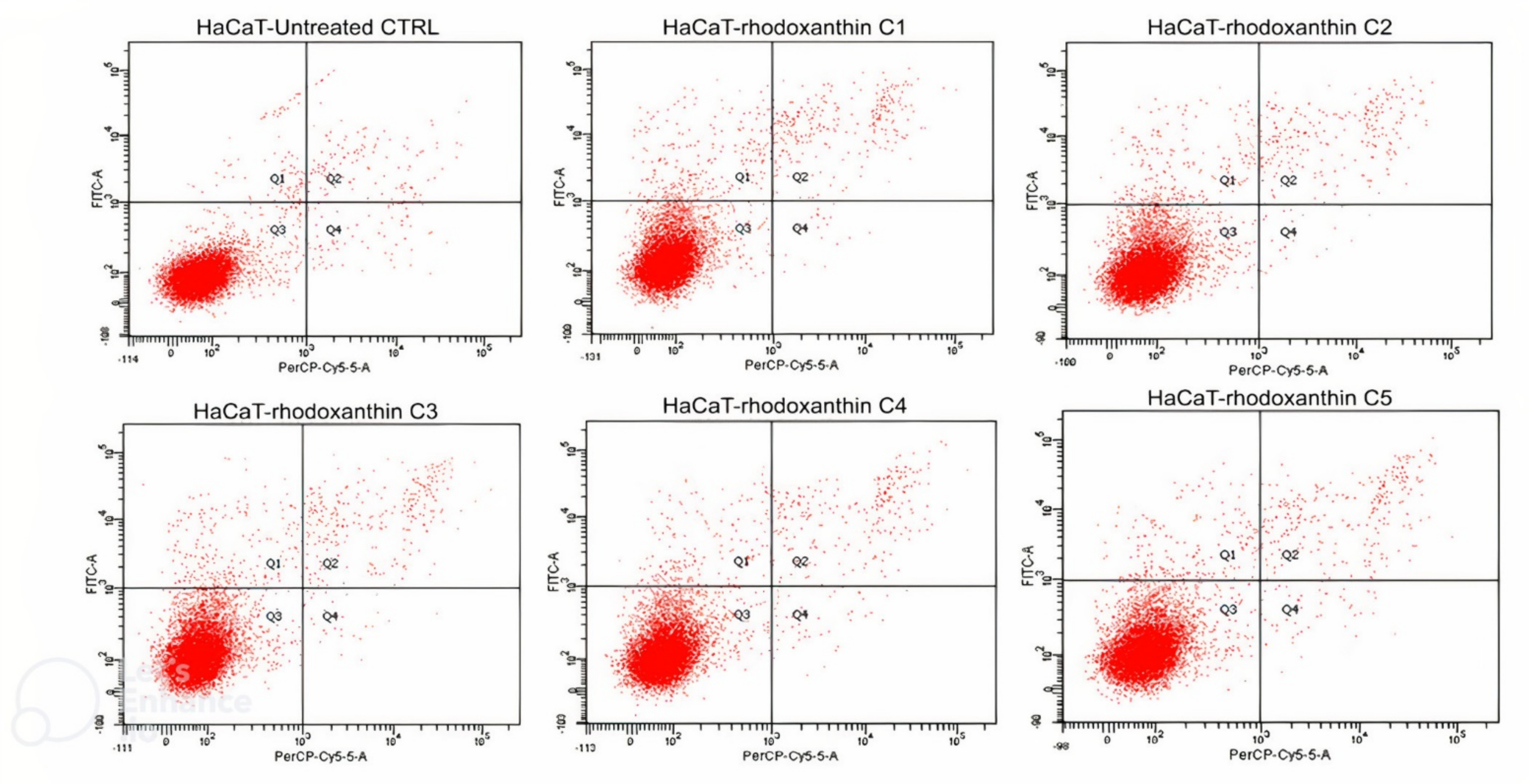
2.6.3. Apoptosis Assay
2.7. Statistical Analysis
3. Results and Discussions
3.1. Phytochemicals Content in Taxus baccata L. Aril
3.1.1. Carotenoids Content
3.1.2. Total Phenolic and Total Flavonoid Contents
3.2. Antioxidant Activity
3.3. Cell Cultures and Cytotoxic Assay
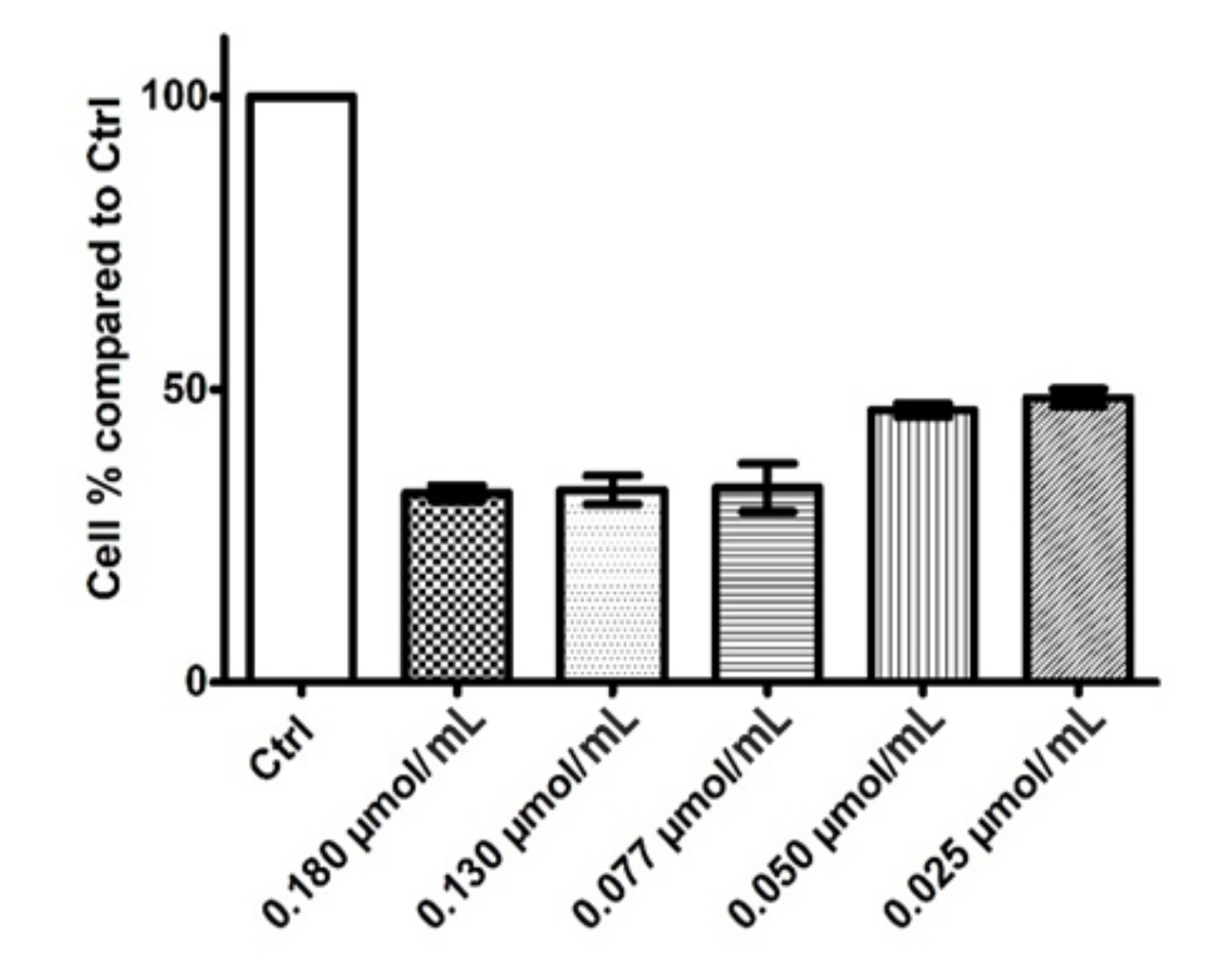
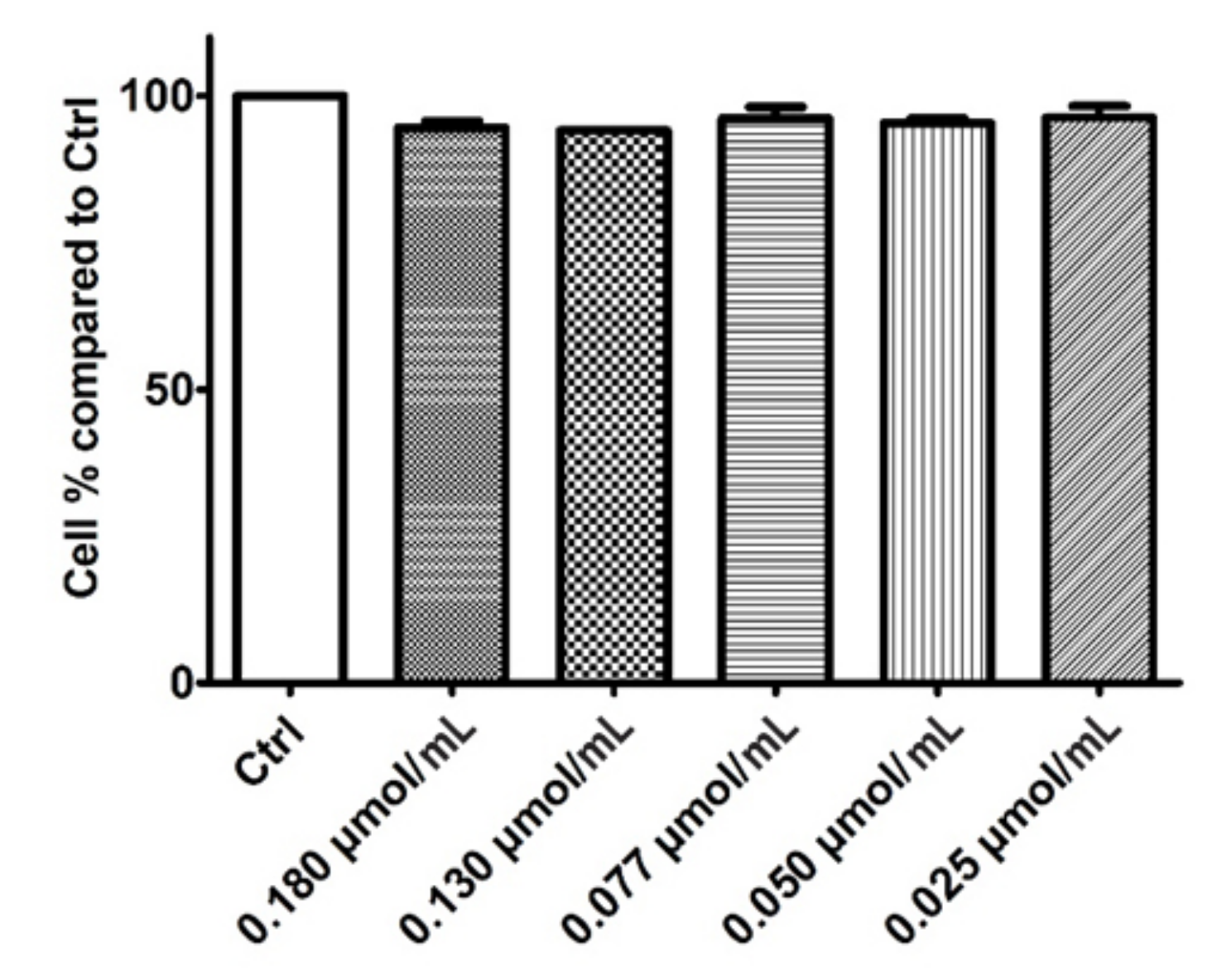
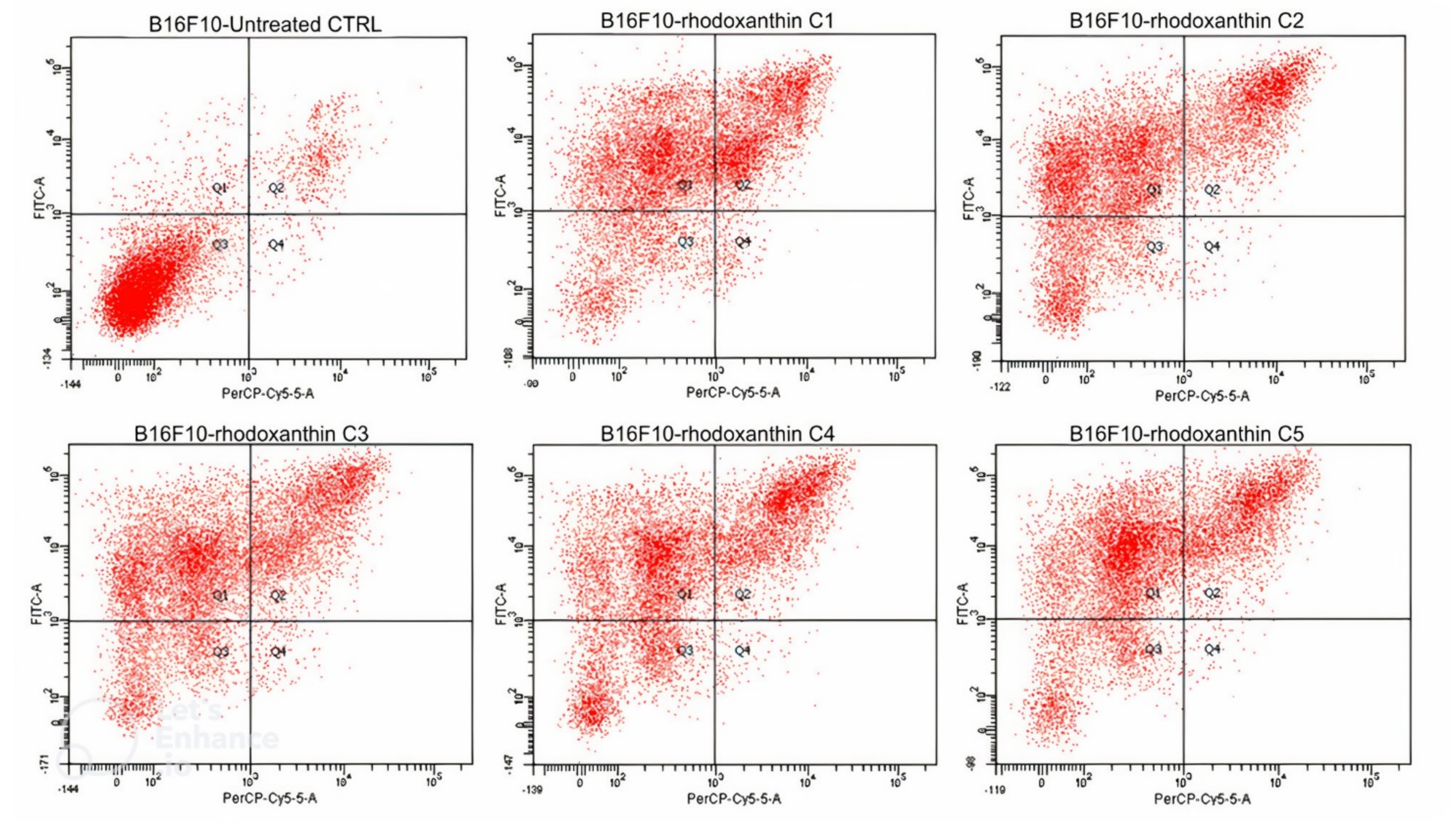
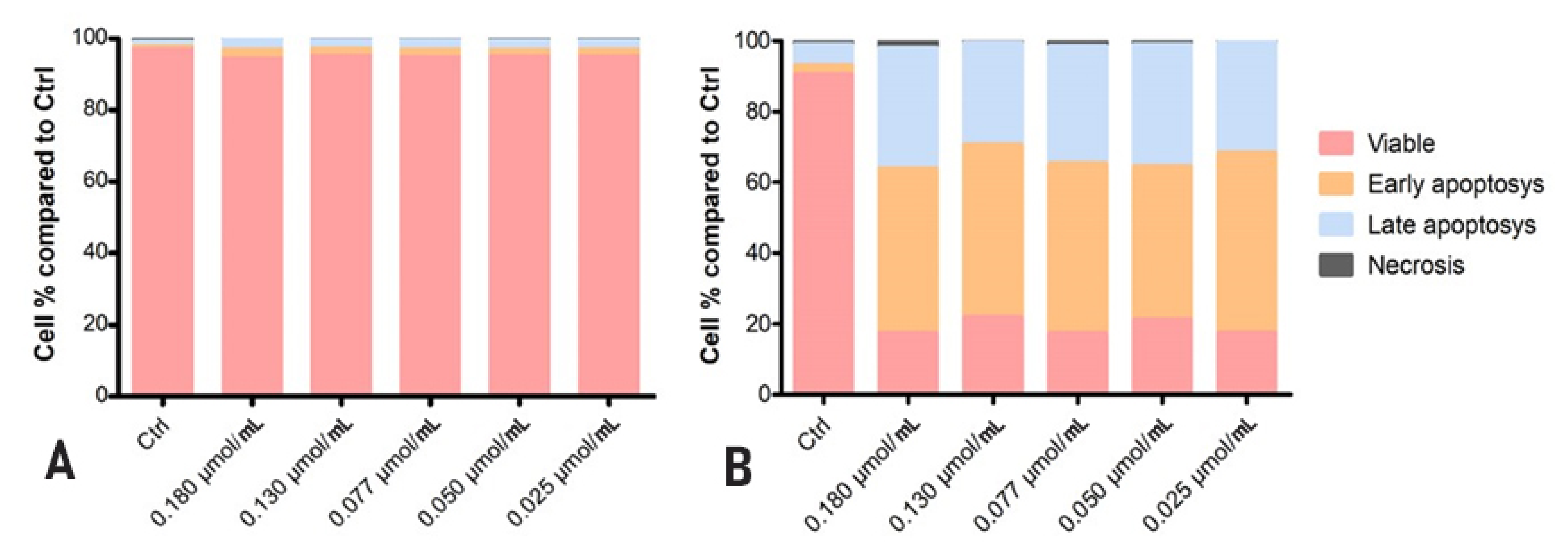
3.3.1. Cytotoxic Activity of Rhodoxanthin on B16F10 Metastatic Murine Melanoma and Human HaCaT Keratinocytes
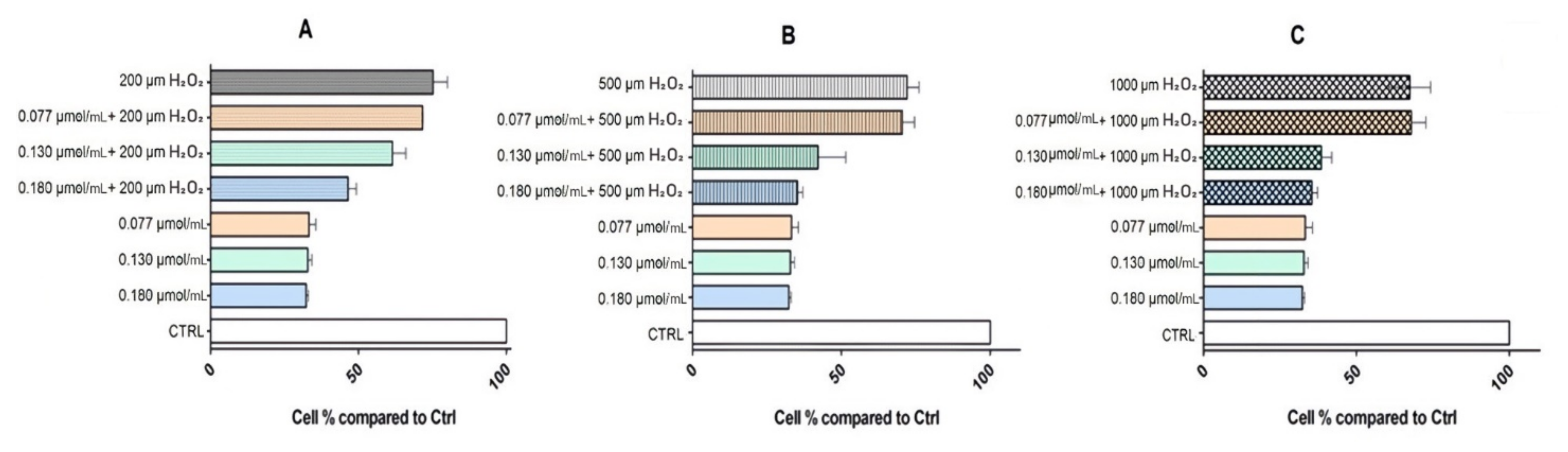
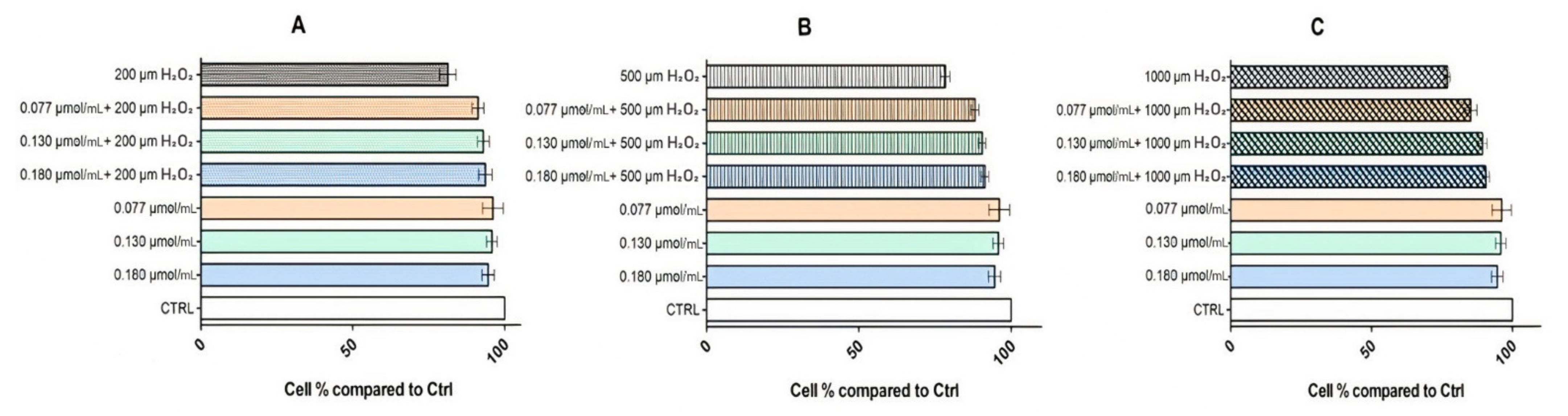
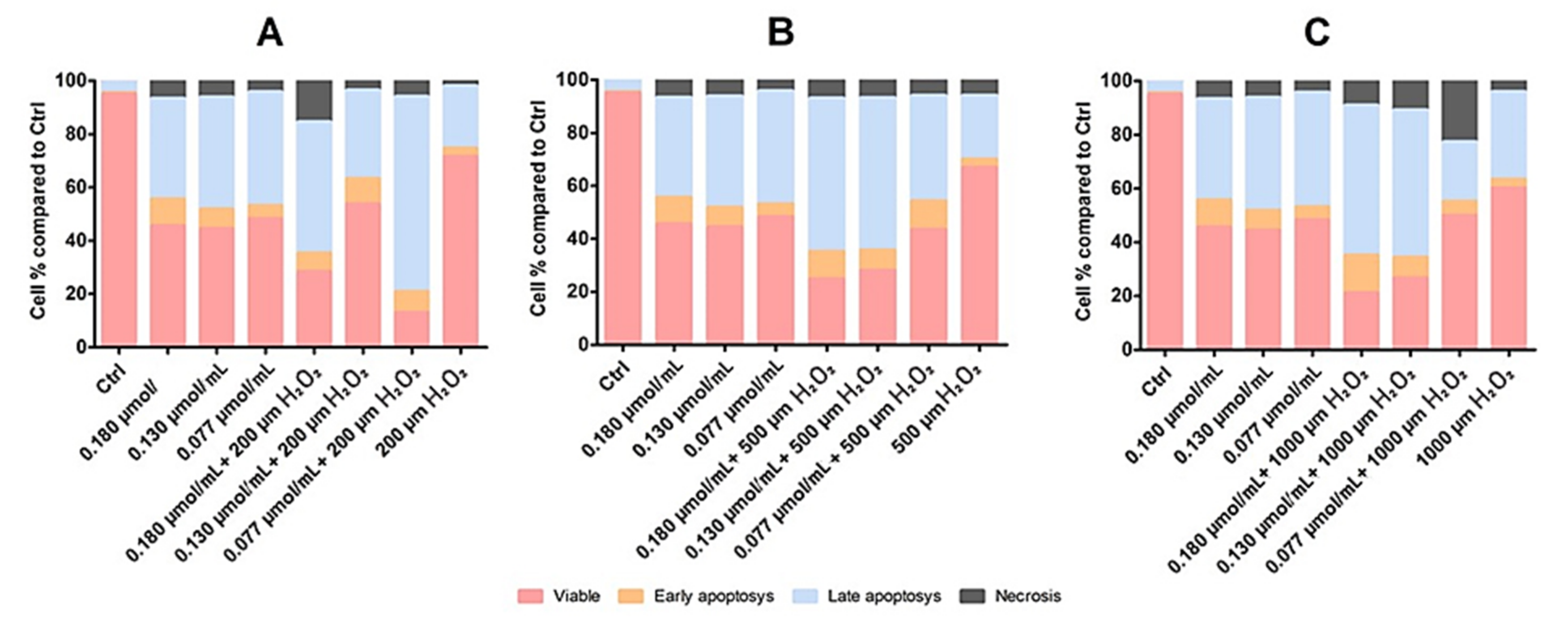
3.3.2. Cytoprotective Effects of Rhodoxanthin against Hydrogen Peroxide-Induced Oxidative Stress in Human Keratinocytes
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Thomas, P.A.; Polwart, A. Taxus baccata L. J. Ecol. 2003, 91, 489–524. [Google Scholar] [CrossRef]
- Andrei, S.; Pintea, A.; Bolos, F.; Bunea, A. Comparative studies regarding antioxidant actions of oxygenated normal and retro carotenoids in mouse liver and skin. Bull. USAMV-CN 2007, 64, 31–35. [Google Scholar]
- Stanković, D.M.; Cvetković, M.D.; Topuzović, V.B.; Mihailović, S.D.; Marković, S.D. Antioxidant and anticancer properties of leaves and seed cones from european yew (Taxus baccata L.). Arch. Biol. Sci. Belgrade 2015, 67, 525–534. [Google Scholar]
- Natasha, G.; Chan, M.; Gue, Y.X.; Gorog, D.A. Fatal heart block from intentional yew tree (Taxus baccata) ingestion: A case report. Eur. Heart J. Case Rep. 2020, 4, 1–4. [Google Scholar] [CrossRef] [Green Version]
- Zutter, A.; Hauri, K.; Evers, K.S.; Uhde, S.; Fassl, J.; Reuthebuch, O.T.; Berset, A.; Kühne, M.; Donner, B.C. Chaotic arrhythmia during successful resuscitation after ingestion of yew (Taxus baccata) needles. Pediatr. Emerg. Care 2017, 35, e223–e225. [Google Scholar] [CrossRef]
- Willaert, W.; Claessens, P.; Vankelecom, B.; Vanderheyden, M. Intoxication with Taxus baccata: Cardiac arrhythmias following yew leaves ingestion. Pacing Clin. Electrophysiol. 2002, 25 Pt 1, 511–512. [Google Scholar] [CrossRef]
- BBC. Ben Hines Died After Ingesting Yew Tree Poison. 2014. Available online: https://www.bbc.co.uk/news/uk-england-norfolk-26089688 (accessed on 13 December 2021).
- Malik, S.; Cusido, R.M.; Mirjalili, M.H.; Moyano, E.; Palazon, J.; Bonfill, M. Production of the anticancer drug taxol in Taxus baccata suspension cultures: A review. Process Biochem. 2011, 46, 23–34. [Google Scholar] [CrossRef]
- Grobosch, T.; Schwarze, B.; Stoecklein, D.; Binscheck, T. Fatal poisoning with Taxus baccata. Quantification of paclitaxel (taxol A), 10-Deacetyltaxol, baccatin III, 10-deacetylbaccatin III, cephalomannine (taxol B), and 3, 5-dimethoxyphenol in body fluids by liquid chromatography–tandem mass spectrometry. J. Anal. Toxicol. 2012, 36, 36–43. [Google Scholar] [CrossRef]
- Siegle, L.; Pietsch, J. Taxus ingredients in the red arils of Taxus baccata L. determined by HPLC-MS/MS. Phytochem. Anal. 2018, 29, 446–451. [Google Scholar] [CrossRef]
- Patel, P.K.; Patel, K.V.; Gandhi, T.R. Evaluation of effect of Taxus baccata leaves extract on bronchoconstriction and bronchial hyperreactivity in experimental animals. Glob. J. Pharmacol. 2009, 3, 141–148. [Google Scholar] [CrossRef] [Green Version]
- DeFilipps, R.A.; Krupnick, G.A. The medicinal plants of Myanmar. PhytoKeys 2018, 102, 23–24. [Google Scholar] [CrossRef]
- Sati, B.; Purohit, V.; Bhatt, S.; Andola, H.C. Isolation of bioactive compounds of Taxus baccata and Swertia chirata plants of Uttarakhand region by GC-MS. Int. J. Sci. Dev. Res. (IJSDR) 2021, 6, 108–110. [Google Scholar]
- Lanker, U.; Malik, A.R.; Gupta, N.K.; Butola, J. Natural regeneration status of the endangered medicinal plant, Taxus baccata Hook. F. syn. T. wallichiana, in northwest Himalaya. Int. J. Biodivers. Sci. Ecosyst. Serv. Manag. 2010, 6, 20–27. [Google Scholar] [CrossRef]
- Tabaszewska, M.; Rutkowska, J.; Skoczylas, Ł.; Słupski, J.; Antoniewska, A.; Smoleń, S.; Łukasiewicz, M.; Baranowski, D.; Duda, I.; Pietsch, J. Red Arils of Taxus baccata L.—A New Source of Valuable Fatty Acids and Nutrients. Molecules 2021, 26, 723. [Google Scholar] [CrossRef]
- Hudon, J.; Derbyshire, D.; Leckie, S.; Flinn, T. Diet-induced plumage erythrism in Baltimore Orioles as a result of the spread of introduced shrubs. Wilson J. Ornithol. 2013, 125, 88–96. [Google Scholar] [CrossRef]
- Hudon, J.; Driver, R.J.; Rice, N.H.; Lloyd-Evans, T.L.; Craves, J.A.; Shustack, D.P. Diet explains red fl ight feathers in Yellow-shafted Flickers in eastern North America. Auk Ornithol. Adv. 2017, 134, 22–33. [Google Scholar]
- Andrei, S.; Pintea, A.; Bunea, A. Studies regarding retro carotenoids content from Taxus baccata, 653–655. In Proceedings of the XL Croatian Symposium on Agriculture with International Participation, Opatija, Croatia, 15–18 February 2005. [Google Scholar]
- Kim, B.H.; Choi, M.S.; Lee, H.G.; Lee, S.H.; Noh, K.H.; Kwon, S.; Jeong, H.; Lee, E.H.; Yi, J.Y.; Park, J.; et al. Photoprotective potential of penta-o-galloyl-beta-dglucose by targeting NF-kappaB and MAPK signaling in UVB radiation-induced human dermal fibroblasts and mouse skin. Mol. Cells 2015, 38, 982–990. [Google Scholar]
- Britton, G.; Khachik, F. Carotenoids in Food. In Carotenoids: Nutrition and Health; Britton, G., Liaaen-Jensen, S., Pfander, H., Eds.; Birkhäuser Verlag: Basel, Switzerland; Boston, MA, USA; Berlin/Heidelberg, Germany, 2009; Volume 5, pp. 55–57. [Google Scholar]
- Pérez-Gálvez, A.; Viera, I.; Roca, M. Carotenoids and Chlorophylls as Antioxidants. Antioxidants 2020, 9, 505. [Google Scholar] [CrossRef]
- Available online: https://retirementrovers.com/retiring-in-cluj-napoca-romania/ (accessed on 22 May 2022).
- Available online: https://www.climatestotravel.com/climate/romania/cluj-napoca (accessed on 20 January 2022).
- Available online: https://en.wikipedia.org/wiki/Cluj-Napoca#Climate (accessed on 20 January 2022).
- Available online: https://www.timeanddate.com/weather/romania/cluj-napoca/climate (accessed on 20 January 2022).
- Available online: https://business-review.eu/featured/cluj-napoca-tops-the-list-of-cities-with-best-air-quality-in-europe-64994 (accessed on 20 January 2022).
- Britton, G. UV/vis Spectroscopy. In Carotenoids. Volume 1B: Spectroscopy; Britton, G., Liaaen-Jensen, S., Pfander, H., Eds.; Birkhäuser Verlag: Basel, Switzerland; Boston, MA, USA; Berlin/Heidelberg, Germany, 1995; pp. 13–62. [Google Scholar]
- Chex, R.; Schweiggert, F.; Wüstenberg, B.; Bonrath, W.; Schäfer, C.; Schweiggert, R. Kinetic and Thermodynamic Study of the Thermally Induced (E/Z)-Isomerization of the retro-Carotenoid Rhodoxanthin. J. Agric. Food Chem. 2020, 68, 5259–5269. [Google Scholar] [CrossRef]
- Criste, A.; Urcan, A.C.; Bunea, A.; Pripon Furtuna, F.R.; Olah, N.K.; Madden, R.H.; Corcionivoschi, N. Phytochemical composition and biological activity of berries and leaves from four Romanian sea buckthorn (Hippophae rhamnoides L.) varieties. Molecules 2020, 25, 1170. [Google Scholar] [CrossRef] [Green Version]
- Ștefănescu, B.-E.; Călinoiu, L.F.; Ranga, F.; Fetea, F.; Mocan, A.; Vodnar, D.C.; Crișan, G. Chemical Composition and Biological Activities of the North-West Romanian Wild Bilberry (Vaccinium myrtillus L.) and Lingonberry (Vaccinum vitis-idaea L.) Leaves. Antioxidants 2020, 9, 495. [Google Scholar] [CrossRef]
- Siddiqui, N.; Rauf, A.; Latif, A.; Mahmood, Z. Spectrophotometric determination of the total phenolic content, spectral and fluorescence study of the herbal Unani drug Gul-e-Zoofa (Nepeta bracteata Benth). J. Taibah Univ. Med. Sci. 2017, 12, 360–363. [Google Scholar] [CrossRef]
- Pop, R.M.; Puia, I.C.; Puia, A.; Chedea, V.S.; Leopold, N.; Bocsan, I.C.; Buzoianu, A.D. Characterization of Trametes versicolor: Medicinal Mushroom with Important Health Benefits. Not. Bot. Horti. Agrobot. 2018, 46, 343–349. [Google Scholar] [CrossRef] [Green Version]
- Vlase, L.; Mocan, A.; Hanganu, D.; Benedec, D.; Gheldiu, A.-M.; Crişan, G. Comparative study of polyphenolic content, antioxidant and antimicrobial activity of four Galium species (Rubiaceae). Dig. J. Nanomater. Biostruct. 2014, 9, 1085–1094. [Google Scholar]
- Ebrahimabadi, A.H.; Mazoochi, A.; Kashi, F.J.; Djafari-Bidgoli, Z.; Batooli, H. Essential oil composition and antioxidant and antimicrobial properties of the aerial parts of Salvia eremophila Boiss. from Iran. Food Chem. Toxicol. Int. J. Publ. Br. Ind. Biol. Res. Assoc. 2010, 48, 1371–1376. [Google Scholar]
- Benedec, D.; Hanganu, D.; Lorena, F.; Oniga, I.; Brindusa, T.; Olah, N.-K.; Gheldiu, A.-M.; Raita, O.; Vlase, L. Chemical, antioxidant and antibacterial studies of Romanian heracleum sphondylium. Farmacia 2017, 65, 252–256. [Google Scholar]
- Iacovită, C.; Florea, A.; Scoruş, L.; Pall, E.; Dudric, R.; Moldovan, A.I.; Ştiufiuc, R.; Tetean, R.; Lucaciu, C.M. Hyperthermia, cytotoxicity, and cellular uptake properties of manganese and zinc ferrite magnetic nanoparticles synthesized by a polyolmediated process. Nanomaterials 2019, 9, 1489. [Google Scholar] [CrossRef] [Green Version]
- Marian, E.; Duteanu, N.; Vicas, L.; Rusu, G.; Jurca, T.; Muresan, M.; Micle, O.; Hangan, A.C.; Stan, R.L.; Ionescu, C.; et al. Synthesis, characterization of inclusion compounds of amygdalin with β-cyclodextrin and sod-like activity and cytotoxicity on hela tumor cells. Arab. J. Chem. 2020, 13, 6828–6837. [Google Scholar] [CrossRef]
- Schex, R.; Lieb, V.M.; Schäfer, C.; Schweiggert, R.; Steingass, C.B. Carotenoid profiles of red-and yellow-colored arils of cultivars of Taxus baccata L. and Taxus× media Rehder. Phytochemistry 2021, 186, 112741. [Google Scholar] [CrossRef]
- LLaczkó-Zöld, E.; Komlósi, A.; Ülkei, T.; Fogarasi, E.; Croitoru, M.; Fülöp, I.; Domokos, E.; Ştefănescu, R.; Varga, E. Extractability of polyphenols from black currant, red currant and gooseberry and their antioxidant activity. Acta Biol. Hung. 2018, 69, 156–169. [Google Scholar] [CrossRef] [PubMed]
- Domínguez, R.; Zhang, L.; Rocchetti, G.; Lucini, L.; Pateiro, M.; Munekata, P.E.S.; Lorenzo, J.M. Elderberry (Sambucus nigra L.) as potential source of antioxidants. Characterization, optimization of extraction parameters and bioactive properties. Food Chem. 2020, 330, 127266. [Google Scholar] [CrossRef]
- Tóth, G.; Barabás, C.; Tóth, A.; Kéry, Á.; Béni, S.; Boldizsár, I.; Varga, E.; Noszál, B. Characterization of antioxidant phenolics in Syringa vulgaris L. flowers and fruits by HPLC-DAD-ESI-MS. Biomed. Chromatogr. 2016, 30, 923–932. [Google Scholar] [CrossRef]
- Varga, E.; Barabás, C.; Tóth, A.; Boldizsár, I.; Noszál, B.; Tóth, G. Phenolic composition, antioxidant and antinociceptive activities of Syringa vulgaris L. bark and leaf extracts. Nat. Prod. Res. 2019, 33, 1664–1669. [Google Scholar] [CrossRef]
- Shin, J.; Song, M.-H.; Oh, J.-W.; Keum, Y.-S.; Saini, R.K. Pro-oxidant Actions of Carotenoids in Triggering Apoptosis of Cancer Cells: A Review of Emerging Evidence. Antioxidants 2020, 9, 532. [Google Scholar] [CrossRef]
- Saini, R.K.; Keum, Y.-S.; Daglia, M.; Rengasamy, K.R. Dietary carotenoids in cancer chemoprevention and chemotherapy: A review of emerging evidence. Pharmacol. Res. 2020, 157, 104830. [Google Scholar] [CrossRef]
- Liu, C.; Guo, H.; DaSilva, N.A.; Li, D.; Zhang, K.; Wan, Y.; Gao, X.-H.; Chen, H.-D.; Seeram, N.P.; Ma, H. Pomegranate (Punica granatum) phenolics ameliorate hydrogen peroxide-induced oxidative stress and cytotoxicity in human keratinocytes. J. Funct. Foods 2019, 54, 559–567. [Google Scholar] [CrossRef]
- Andrei, S.; Pintea, A.; Crainic, D.; Rugina, D.; Bunea, A. In vitro studies regarding the antioxidant effect of rhodoxanthin and canthaxanthin. Cluj Vet. J. 2009, 16, 14–19. [Google Scholar]
- Mundi, N.; Jordan, K.; Doyle, P.; Moore, C. 33% hydrogen peroxide as a Neoadjuvant treatment in the surgical excision of non-melanoma skin cancers: A case series. J. Otolaryngol. Head Neck Surg. 2020, 49, 1–7. [Google Scholar] [CrossRef]
- Sishc, B.J.; Ding, L.; Nam, T.-K.; Heer, C.D.; Rodman, S.N.; Schoenfeld, J.D.; Fath, M.A.; Saha, D.; Pulliam, C.F.; Langen, B.; et al. Avasopasem manganese synergizes with hypofractionated radiation to ablate tumors through the generation of hydrogen peroxide. Sci. Transl. Med. 2021, 13, eabb3768. [Google Scholar] [CrossRef]
- Yang, G.Y.; Liao, J.; Kim, K.; Yurkow, E.J.; Yang, C.S. Inhibition of growth and induction of apoptosis in human cancer cell lines by tea polyphenols. Carcinogenesis 1998, 19, 611–616. [Google Scholar] [CrossRef] [Green Version]












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Analyzed Samples | Total Carotenoids mg/100 g FW | Rhodoxanthin mg/100 g FW | Total Polyphenols mg GAE/100 g FWF | Flavonoids mg QE/100 g FWF |
|---|---|---|---|---|
| Sample 1 | 3.384 | 2.536 | 146.69 | 43.95 |
| Sample 2 | 3.371 | 2.582 | 146.64 | 45.01 |
| Sample 3 | 3.379 | 2.575 | 143.8 | 45.26 |
| Average ± SD | 3.378 ± 0.0053 | 2.564 ± 1.0079 | 145.71 ± 22.3648 | 44.723 ± 15.4256 |
| Compound | Peak | RT (min) | Max Absorption (nm) |
|---|---|---|---|
| Lutein | 1 | 14.44 | 421,445,474 |
| Beta-Carotene | 2 | 25.41 | 425,450,477 |
| Rhodoxanthin isomer | 3 | 29.45 | 337,487 |
| Rhodoxanthin isomer | 4 | 33.21 | 337,487 |
| Rhodoxanthin isomer | 5 | 37.01 | 337,503 |
| Peak No. | Compound | Retention Time Rt (min) | UV λmax (nm) | [M + H]+ (m/z) | Subclass | Concentration (μg/g) |
|---|---|---|---|---|---|---|
| 1 | NI | 3.30 | 230 | 381, 219 | 42.2 | |
| 2 | p-Coumaric acid-glucoside | 6.12 | 320 | 327, 166 | Hydroxycinnamic acid | 1046.235 |
| 3 | Protocatechuic acid | 9.62 | 290 | 155 | Hydroxybenzoic acid | 149.49 |
| 4 | Hydroxy-caffeic acid | 12.20 | 322 | 197 | Hydroxycinnamic acid | 49.37 |
| 5 | Caffeic acid | 13.78 | 322 | 181 | Hydroxycinnamic acid | 95.76 |
| 6 | Catechin-glucoside | 16.45 | 280 | 453, 291 | Flavanol | 131.36 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dumitraş, D.-A.; Bunea, A.; Vodnar, D.C.; Hanganu, D.; Pall, E.; Cenariu, M.; Gal, A.F.; Andrei, S. Phytochemical Characterization of Taxus baccata L. Aril with Emphasis on Evaluation of the Antiproliferative and Pro-Apoptotic Activity of Rhodoxanthin. Antioxidants 2022, 11, 1039. https://doi.org/10.3390/antiox11061039
Dumitraş D-A, Bunea A, Vodnar DC, Hanganu D, Pall E, Cenariu M, Gal AF, Andrei S. Phytochemical Characterization of Taxus baccata L. Aril with Emphasis on Evaluation of the Antiproliferative and Pro-Apoptotic Activity of Rhodoxanthin. Antioxidants. 2022; 11(6):1039. https://doi.org/10.3390/antiox11061039
Chicago/Turabian StyleDumitraş, Daria-Antonia, Andrea Bunea, Dan Cristian Vodnar, Daniela Hanganu, Emoke Pall, Mihai Cenariu, Adrian Florin Gal, and Sanda Andrei. 2022. "Phytochemical Characterization of Taxus baccata L. Aril with Emphasis on Evaluation of the Antiproliferative and Pro-Apoptotic Activity of Rhodoxanthin" Antioxidants 11, no. 6: 1039. https://doi.org/10.3390/antiox11061039
APA StyleDumitraş, D. -A., Bunea, A., Vodnar, D. C., Hanganu, D., Pall, E., Cenariu, M., Gal, A. F., & Andrei, S. (2022). Phytochemical Characterization of Taxus baccata L. Aril with Emphasis on Evaluation of the Antiproliferative and Pro-Apoptotic Activity of Rhodoxanthin. Antioxidants, 11(6), 1039. https://doi.org/10.3390/antiox11061039