
Resveratrol Modulates the Redox Response and Bile Acid Metabolism to Maintain the Cholesterol Homeostasis in Fish Megalobrama amblycephala Offered a High-Carbohydrate Diet



Abstract
:1. Introduction
2. Materials and Methods
2.1. Ethics Statement
2.2. Diets, Fish, and Sampling
2.3. Measurements of Growth Performance and Tissue Metabolites Concentrations
2.4. Real-Time PCR (RT-PCR) and Western Blot
2.5. Statistical Analysis
3. Results
3.1. Feed Utilization and Growth Performance
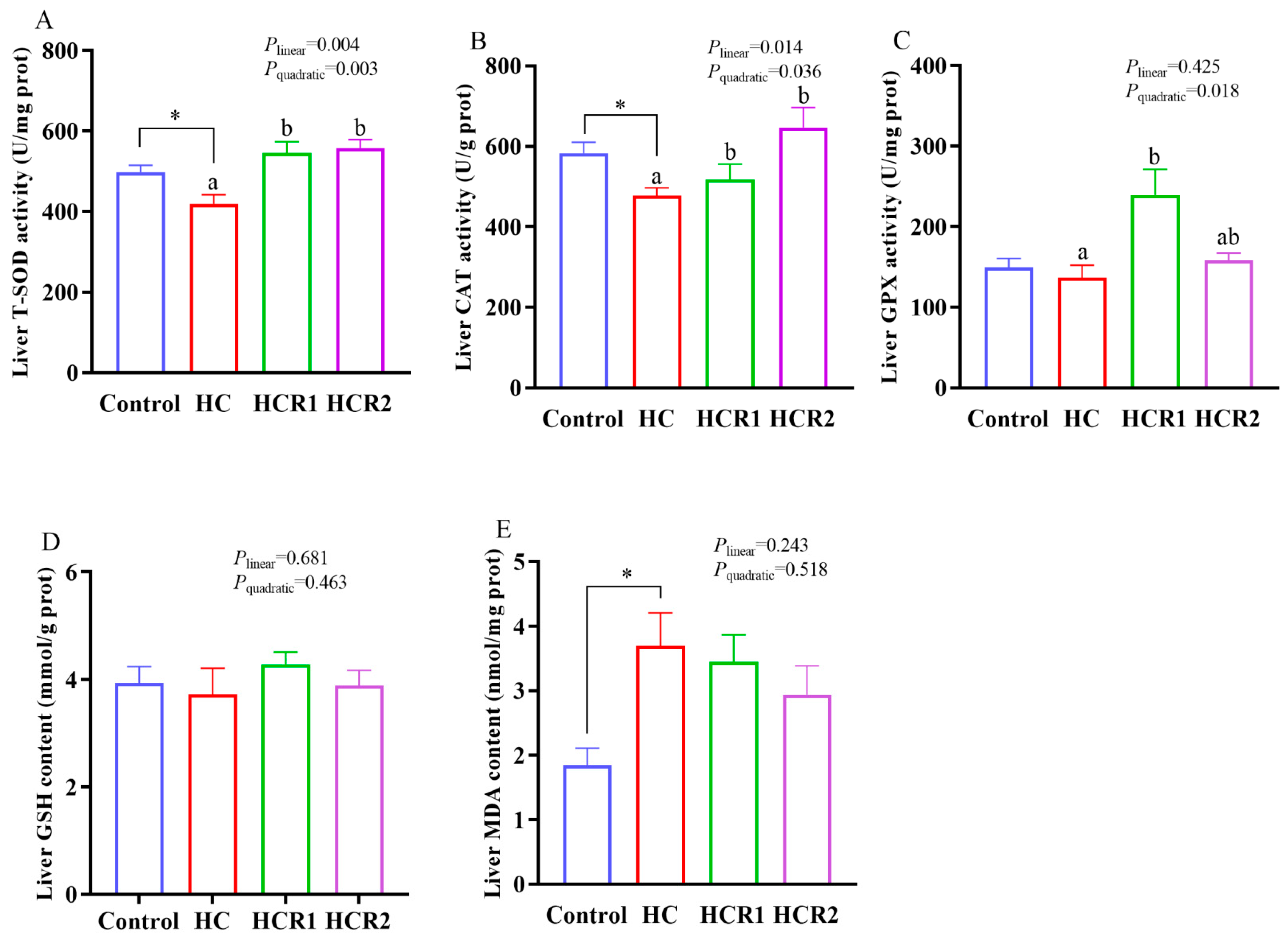
3.2. Hepatic Antioxidant Capability
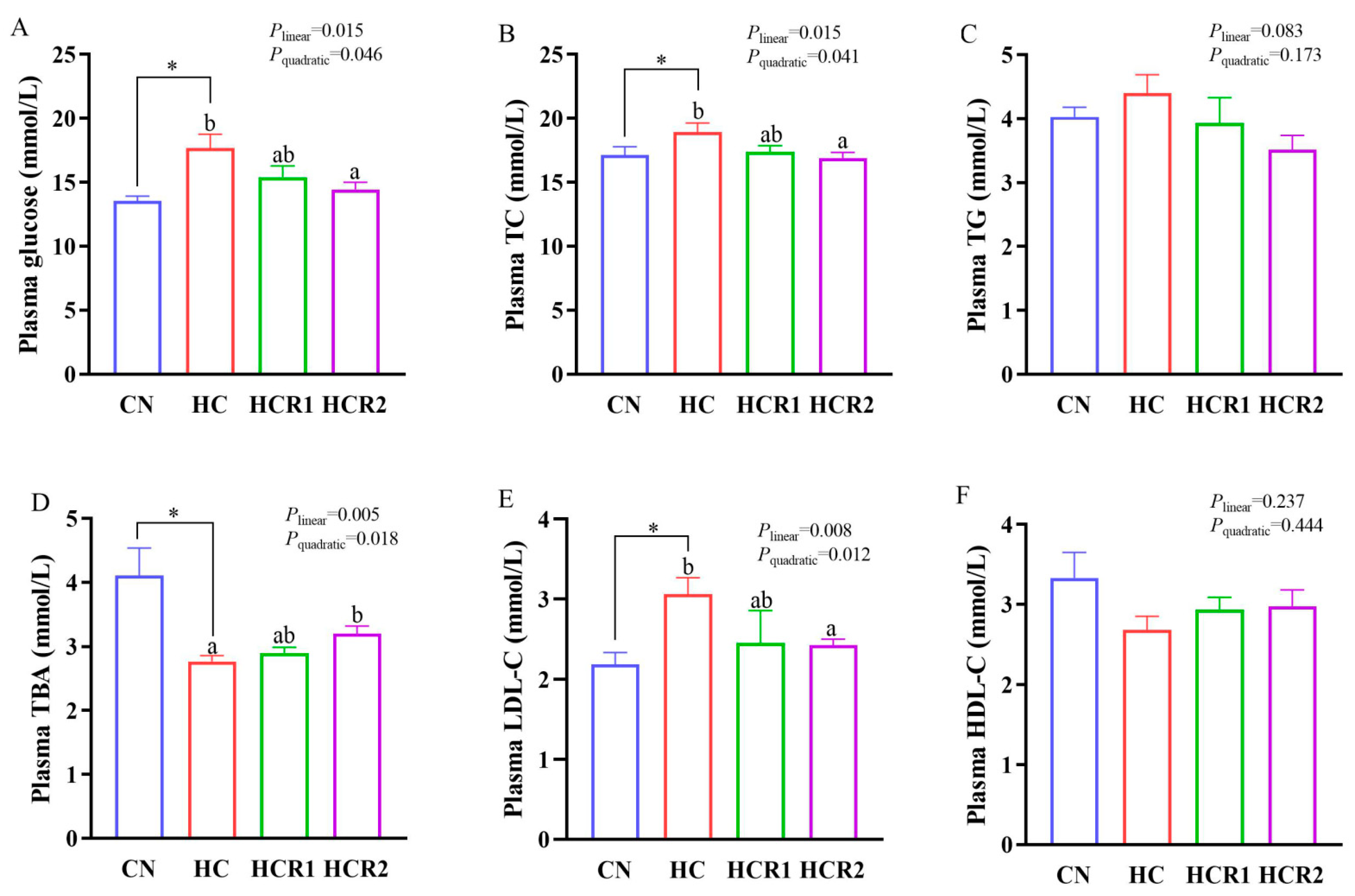
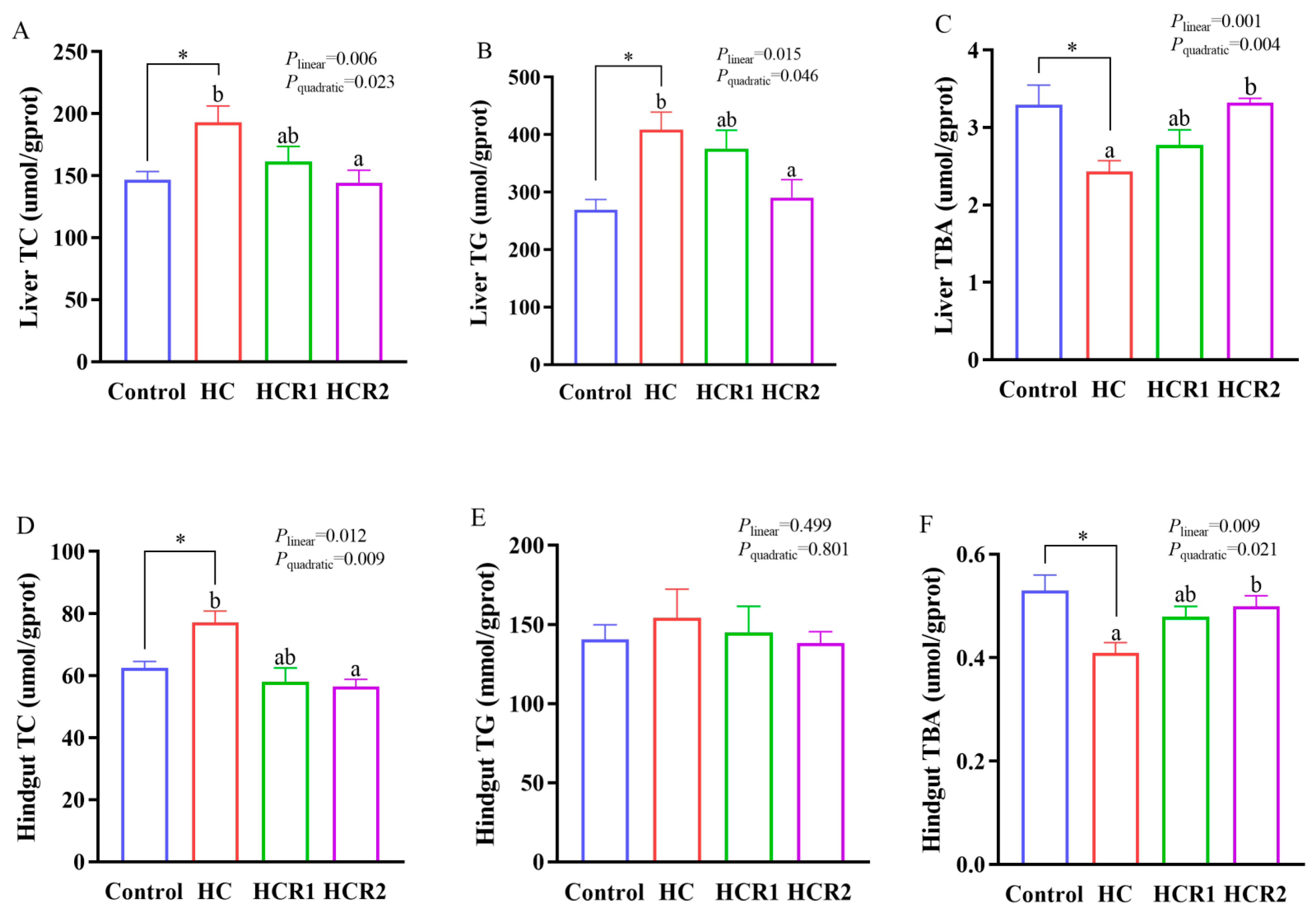
3.3. Tissue and Plasma Metabolites Concentrations
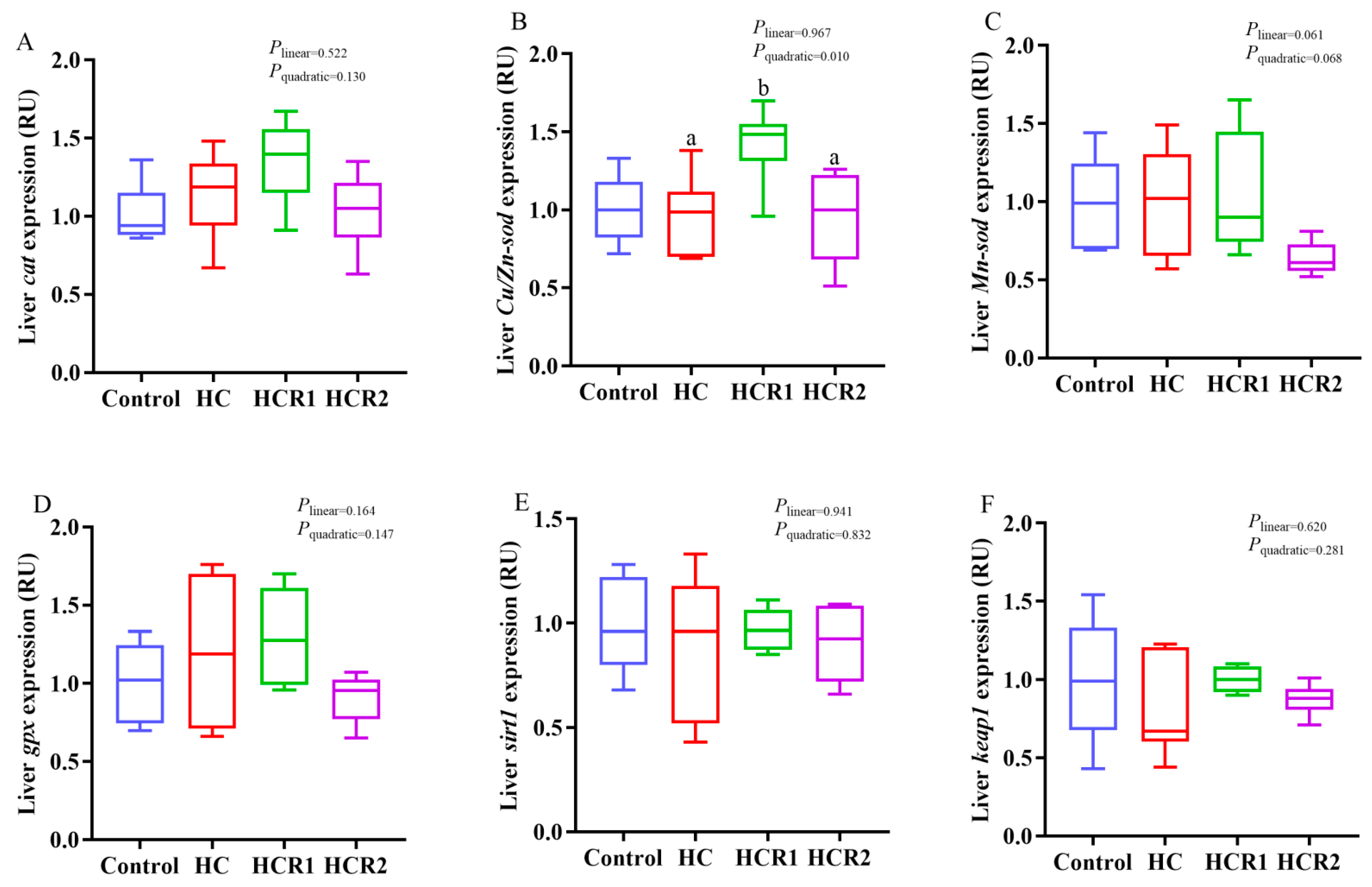
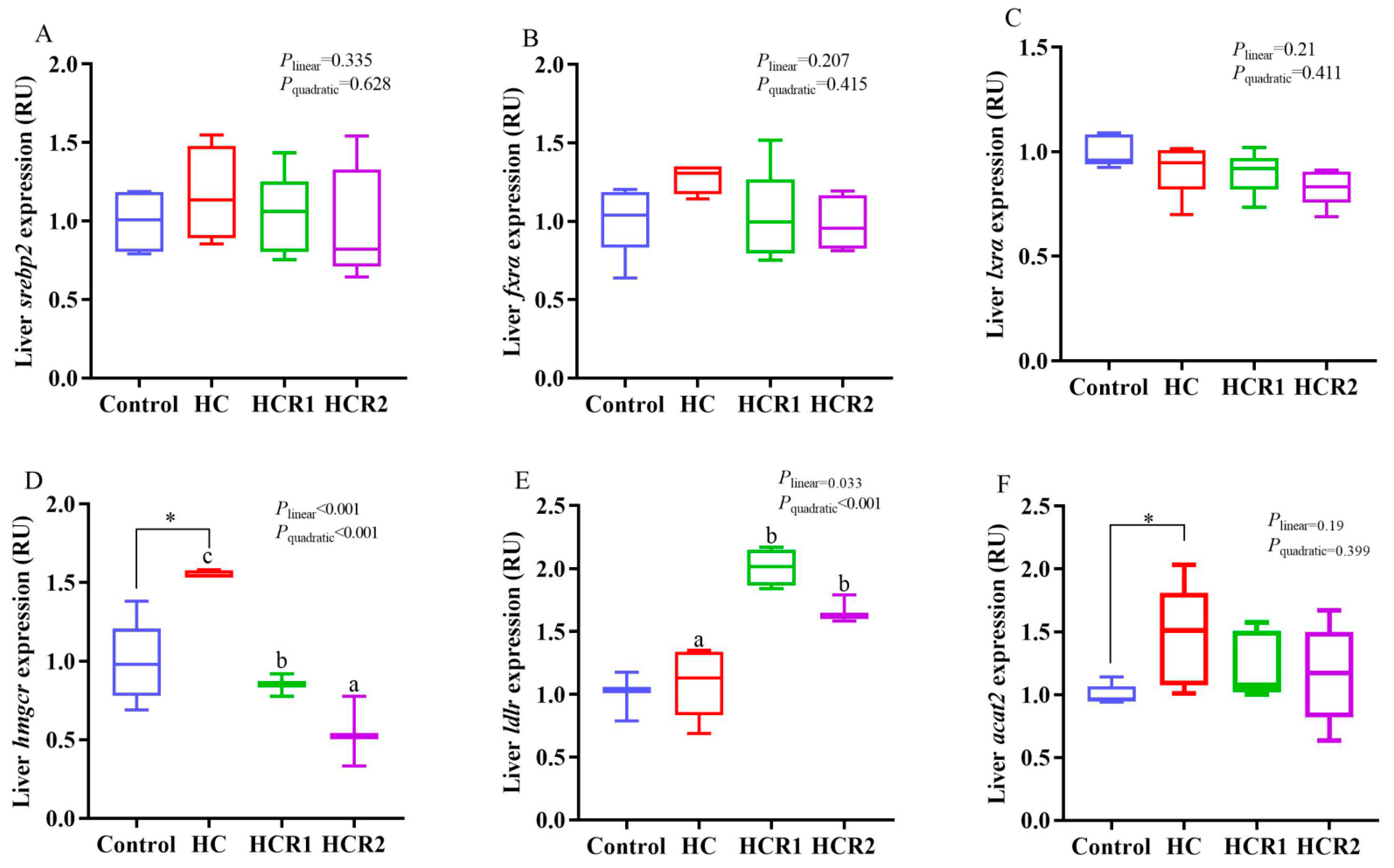
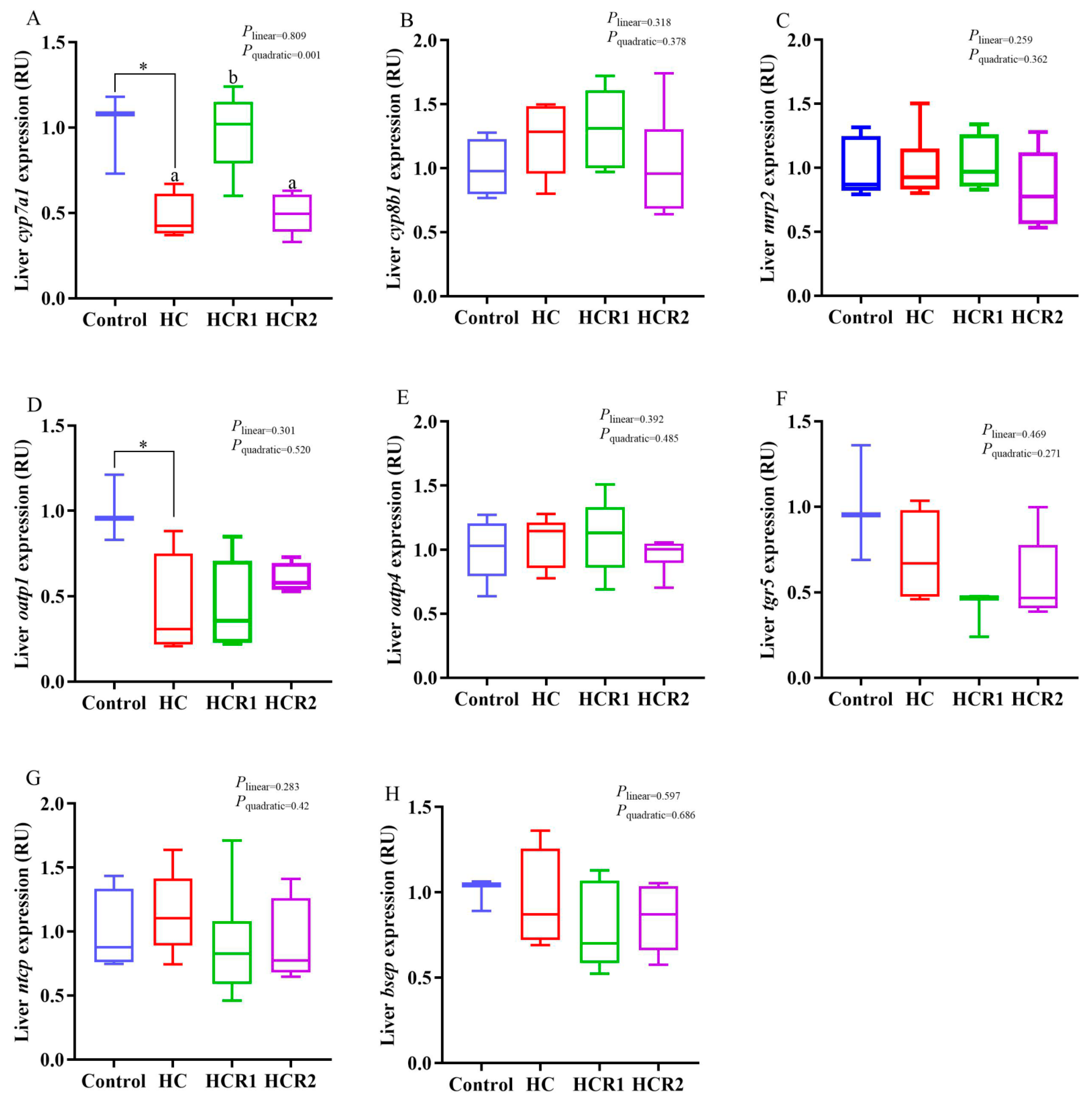
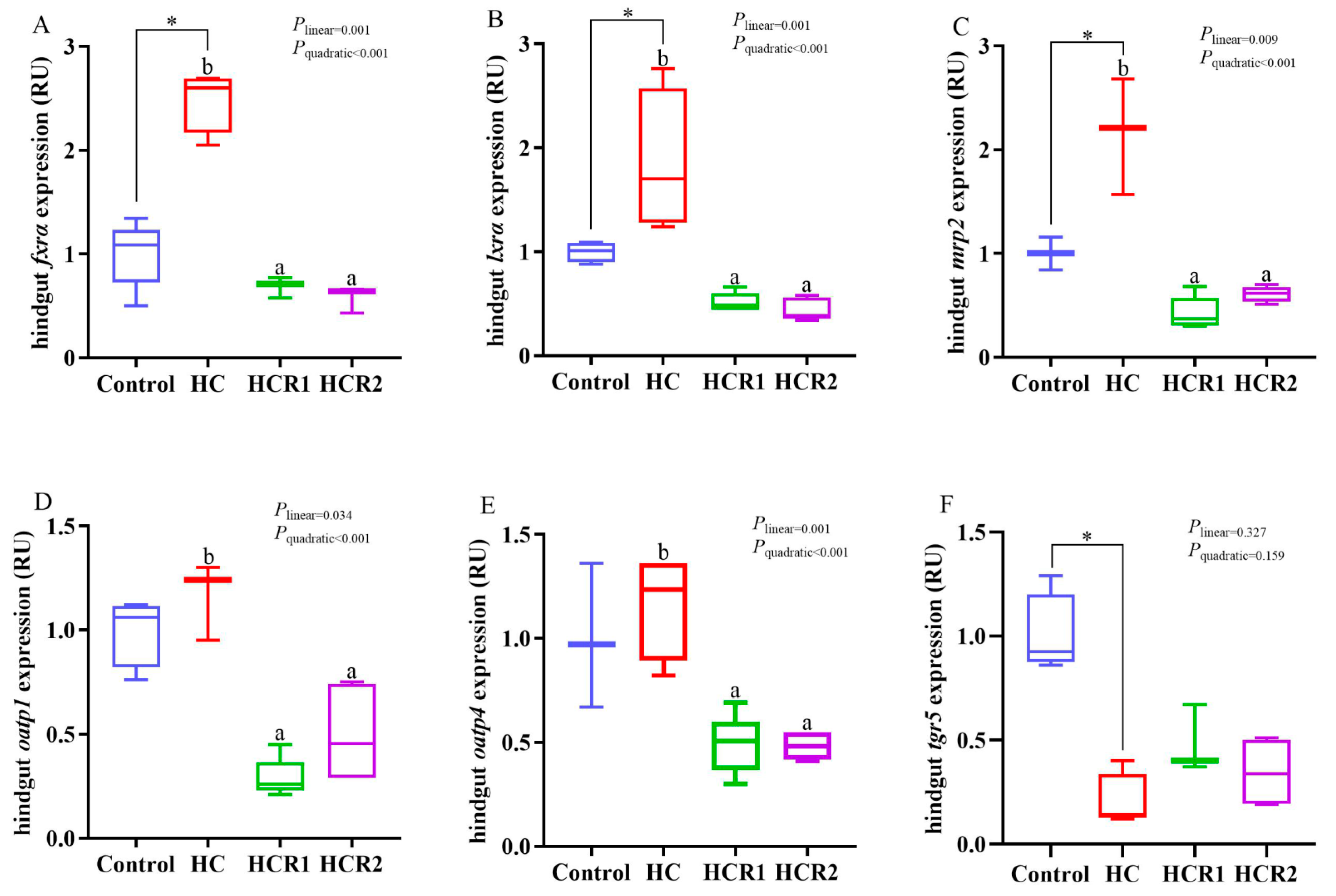
3.4. Transcriptions of the Genes Involved in Cholesterol and Bile Acid Metabolism and the Antioxidant Defense
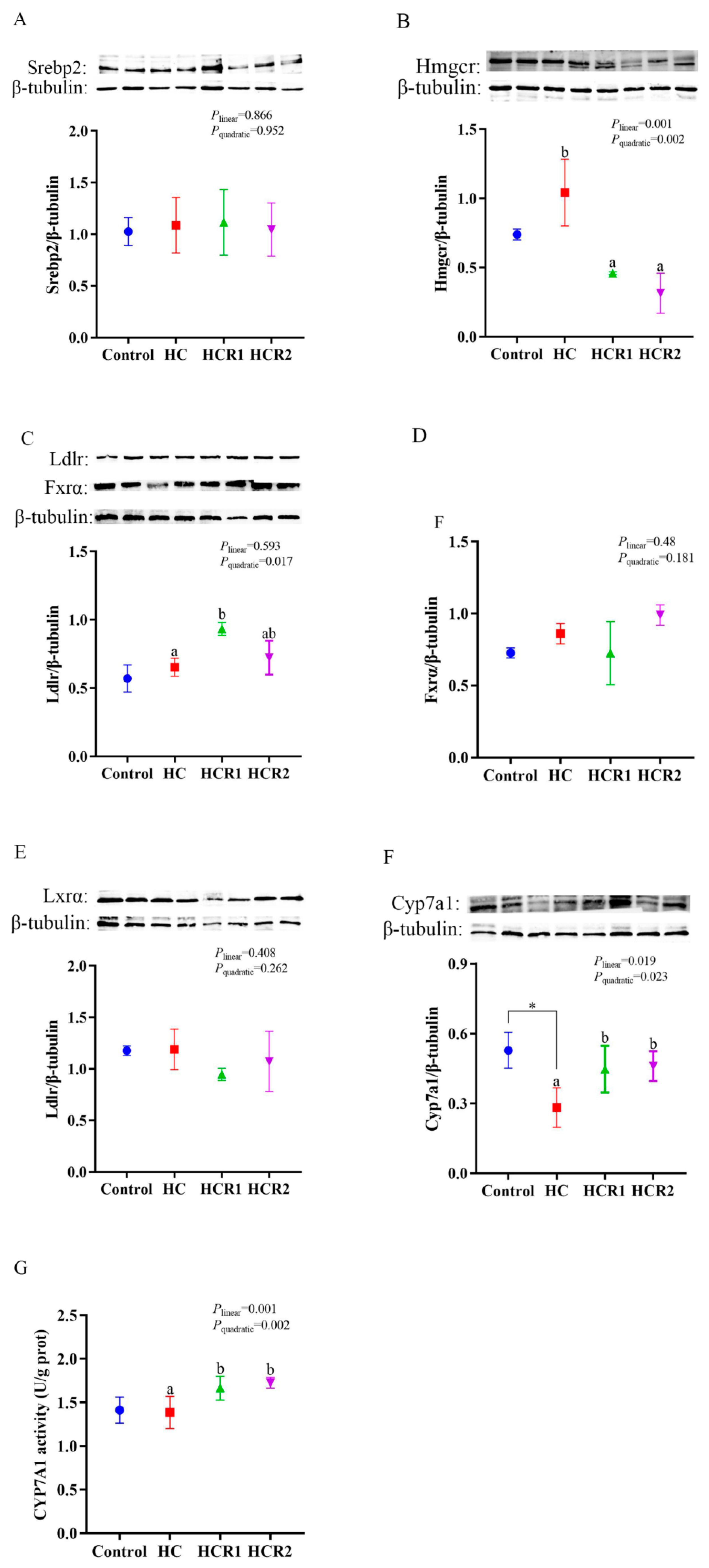
3.5. Hepatic Contents of the Proteins Involved in Cholesterol and Bile Acid Metabolism as Well as the Cyp7a1 Activity
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Li, X.F.; Liu, W.B.; Jiang, Y.Y.; Hao, Z.; Ge, X.P. Effects of dietary protein and lipid levels in practical diets on growth performance and body composition of blunt snout bream (Megalobrama amblycephala) fingerlings. Aquaculture 2010, 303, 65–70. [Google Scholar] [CrossRef]
- Wilson, R. Utilization of dietary carbohydrate by fish. Aquaculture 1994, 124, 67–80. [Google Scholar] [CrossRef]
- Shi, H.J.; Xu, C.; Liu, M.Y.; Wang, B.K.; Liu, W.B.; Chen, D.H.; Zhang, L.; Xu, C.Y.; Li, X.F. Resveratrol improves the energy sensing and glycolipid metabolism of blunt snout bream Megalobrama amblycephala fed high-carbohydrate diets by activating the AMPK–SIRT1–PGC-1α network. Front. Physiol. 2018, 9, 1258. [Google Scholar] [CrossRef] [Green Version]
- Hemre, G.I.; Mommsen, T.P.; Krogdahl, Å. Carbohydrates in fish nutrition: Effects on growth, glucose metabolism and hepatic enzymes. Aquac. Nutr. 2002, 8, 175–194. [Google Scholar] [CrossRef]
- Liu, Q.; Tan, Q.S.; Chen, X.X.; Du, Y.D.; Xia, J.; Yang, Q.; Ma, Y.X. Blood biochemical characteristics and tissue structure changes of grass carp with “liver and gallbladder syndrome”. Anhui Agric. Sci. 2009, 37, 6463–6465, 6467, (In Chinese with English abstract). [Google Scholar] [CrossRef]
- Goodwin, A.E.; Lochmann, R.T.; Tieman, D.M.; Mitchell, A.J. Massive hepatic necrosis and nodular regeneration in largemouth bass fed diets high in available carbohydrate. J. World Aquac. Soc. 2010, 33, 466–477. [Google Scholar] [CrossRef]
- Liang, G.; Wang, J.; Qi, W.; Miao, H.H.; Jian, C.; Qu, Y.X.; Li, B.L.; Song, B.L. The cholesterol absorption inhibitor ezetimibe acts by blocking the sterol-induced internalization of NPC1L1. Cell Metab. 2008, 7, 508–519. [Google Scholar] [CrossRef] [Green Version]
- Tomkin, G.H.; Owens, D. Obesity diabetes and the role of bile acids in metabolism. J. Transl. Intern. Med. 2016, 4, 73–80. [Google Scholar] [CrossRef] [Green Version]
- Li, B.T.; Sun, M.; Li, Y.F.; Wang, J.Q.; Luo, J. Disruption of the ERLIN–TM6SF2–APOB complex destabilizes APOB and contributes to non-alcoholic fatty liver disease. PLoS Genet. 2020, 16, e1008955. [Google Scholar] [CrossRef]
- Chu, B.B.; Liao, Y.C.; Qi, W.; Xie, C.; Du, X.; Wang, J.; Yang, H.; Miao, H.H.; Li, B.L.; Song, B.L. Cholesterol transport through lysosome-peroxisome membrane contacts. Cell 2015, 161, 291–306. [Google Scholar] [CrossRef]
- Luo, J.; Yang, H.; Song, B.L. Mechanisms and regulation of cholesterol homeostasis. Nat. Rev. Mol. Cell Biol. 2020, 21, 225–245. [Google Scholar] [CrossRef] [PubMed]
- Luo, J.; Jiang, L.; Yang, H.; Song, B.L. Routes and mechanisms of post-endosomal cholesterol trafficking: A story that never ends. Traffic 2017, 18, 209–217. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Goldstein, J.L.; Brown, M.S. The LDL receptor. Arterioscler. Thromb. Vasc. Biol. 2009, 29, 431–438. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chang, T.Y.; Li, B.L.; Chang, C.; Urano, Y. Acyl-coenzyme A:cholesterol acyltransferases. Am. J. Physiol. Endocrinol. Metab. 2009, 297, E1. [Google Scholar] [CrossRef] [Green Version]
- Phillips, M.C. Molecular mechanisms of cellular cholesterol efflux. J. Biol. Chem. 2014, 289, 24020–24029. [Google Scholar] [CrossRef] [Green Version]
- Lefebvre, P.; Cariou, B.; Lien, F.; Kuipers, F.; Staels, B. Role of bile acids and bile acid receptors in metabolic regulation. Physiol. Rev. 2009, 89, 147–191. [Google Scholar] [CrossRef] [Green Version]
- Su, G.N.; Sun, H.; Wu, G.; Hu, J. Intestinal and hepatic circulation of bile acids and the role of bile acid receptors in glucose and lipid metabolism. Anim. Nutr. 2019, 31, 88–98, (In Chinese with English abstract). [Google Scholar] [CrossRef]
- Ananthanarayanan, M.; Balasubramanian, N.; Makishima, M.; Mangelsdorf, D.J.; Suchy, F.J. Human bile salt export pump promoter is transactivated by the farnesoid X receptor/bile acid receptor. J. Biol. Chem. 2001, 276, 28857–28865. [Google Scholar] [CrossRef] [Green Version]
- Kast, H.R.; Goodwin, B.; Tarr, P.T.; Jones, S.A.; Anisfeld, A.M.; Stoltz, C.M.; Tontonoz, P.; Kliewer, S.; Willson, T.M.; Edwards, P.A. Regulation of multidrug resistance-associated protein 2 (ABCC2) by the nuclear receptors pregnane X receptor, farnesoid X-activated receptor, and constitutive androstane receptor. J. Biol. Chem. 2002, 277, 2908–2915. [Google Scholar] [CrossRef] [Green Version]
- Ridlon, J.M.; Harris, S.C.; Bhowmik, S.; Kang, D.J.; Hylemon, P.B. Consequences of bile salt biotransformations by intestinal bacteria. Gut Microbes 2016, 7, 22–39. [Google Scholar] [CrossRef]
- Chen, F.; Ma, L.; Dawson, P.A.; Sinal, C.J.; Sehayek, E.; Gonzalez, F.J.; Breslow, J.; Ananthanarayanan, M.; Shneider, B.L. Liver receptor homologue-1 mediates species- and cell line-specific bile acid-dependent negative feedback regulation of the apical sodium-dependent bile acid transporter. J. Biol. Chem. 2003, 278, 19909–19916. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zollner, G.; Wagner, M.; Fickert, P.; Geier, A.; Fuchsbichler, A.; Silbert, D.; Gumhold, J.; Zatloukal, K.; Kaser, A.; Tilg, H. Role of nuclear receptors and hepatocyte-enriched transcription factors for Ntcp repression in biliary obstruction in mouse liver. Am. J. Physiol. Gastrointest. Liver Physiol. 2005, 289, G798. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maeda, T.; Miyata, M.; Yotsumoto, T.; Kobayashi, D.; Nozawa, T.; Toyama, K.; Gonzalez, F.J.; Yamazoe, Y.; Tamai, I. Regulation of drug transporters by the farnesoid X receptor in mice. Mol. Pharm. 2004, 1, 281–289. [Google Scholar] [CrossRef] [PubMed]
- Kalaany, N.Y.; Mangelsdorf, D.J. LXRs and FXR: The yin and yang of cholesterol and fat metabolism. Annu. Rev. Physiol. 2006, 68, 159–191. [Google Scholar] [CrossRef] [PubMed]
- Baur, J.A.; Sinclair, D.A. Therapeutic potential of resveratrol: The in vivo evidence. Nat. Rev. Drug Discov. 2006, 5, 493–506. [Google Scholar] [CrossRef]
- Chen, M.L.; Yi, L.; Jin, X.; Liang, X.Y.; Zhou, Y.; Zhang, T.; Xie, Q.; Zhou, X.; Chang, H.; Fu, Y.J. Resveratrol attenuates vascular endothelial inflammation by inducing autophagy through the cAMP signaling pathway. Autophagy 2013, 9, 2033–2045. [Google Scholar] [CrossRef]
- Cottart, C.H.; Nivet-Antoine, V.; Beaudeux, J.L. Review of recent data on the metabolism, biological effects, and toxicity of resveratrol in humans. Mol. Nutr. Food Res. 2014, 58, 7–21. [Google Scholar] [CrossRef]
- Miura, D.; Miura, Y.; Yagasaki, K. Hypolipidemic action of dietary resveratrol, a phytoalexin in grapes and red wine, in hepatoma-bearing rats. Life Sci. 2003, 73, 1393–1400. [Google Scholar] [CrossRef]
- Chen, Q.; Wang, E.; Ma, L.; Zhai, P. Dietary resveratrol increases the expression of hepatic 7α-hydroxylase and ameliorates hypercholesterolemia in high-fat fed C57BL/6J mice. Lipids Health Dis. 2012, 11, 1–8. [Google Scholar] [CrossRef] [Green Version]
- Chothe, P.P.; Swaan, P.W. Resveratrol promotes degradation of the human bile acid transporter ASBT (SLC10A2). Biochem. J. 2014, 459, 301–312. [Google Scholar] [CrossRef]
- Torno, C.; Staats, S.; de Pascual-Teresa, S.; Rimbach, G.; Schulz, C. Fatty Acid Profile Is Modulated by Dietary Resveratrol in Rainbow Trout (Oncorhynchus mykiss). Mar. Drugs 2017, 15, 252. [Google Scholar] [CrossRef] [Green Version]
- Torno, C.; Staats, S.; Michl, S.C.; de Pascual-Teresa, S.; Izquierdo, M.; Rimbach, G.; Schulz, C. Fatty Acid Composition and Fatty Acid Associated Gene-Expression in Gilthead Sea Bream (Sparus aurata) Are Affected by Low-Fish Oil Diets, Dietary Resveratrol, and Holding Temperature. Mar. Drugs 2018, 16, 379. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, D.; Yan, Y.; Tian, H.; Jiang, G.; Li, X.; Liu, W. Resveratrol supplementation improves lipid and glucose metabolism in high-fat diet-fed blunt snout bream. Fish Physiol. Biochem. 2018, 44, 163–173. [Google Scholar] [CrossRef]
- Ran, G.; Ying, L.; Li, L.; Yan, Q.; Yi, W.; Ying, C.; Wu, H.; Ye, X. Resveratrol ameliorates diet-induced dysregulation of lipid metabolism in zebrafish (Danio rerio). PLoS ONE 2017, 12, e0180865. [Google Scholar] [CrossRef] [Green Version]
- Jia, E.; Yan, Y.; Zhou, M.; Li, X.; Jiang, G.; Liu, W.; Zhang, D. Combined effects of dietary quercetin and resveratrol on growth performance, antioxidant capability and innate immunity of blunt snout bream (Megalobrama amblycephala). Anim. Feed. Sci. Technol. 2019, 256, 114268. [Google Scholar] [CrossRef]
- Li, X.F.; Wang, Y.; Liu, W.B.; Jiang, G.Z.; Zhu, J. Effects of dietary carbohydrate/lipid ratios on growth performance, body composition and glucose metabolism of fingerling blunt snout bream Megalobrama amblycephala. Aquac. Nutr. 2013, 19, 701–708. [Google Scholar] [CrossRef]
- Tian, J.J.; Jin, Y.Q.; Yu, E.M.; Sun, J.H.; Xia, Y.; Zhang, K.; Li, Z.F.; Gong, W.B.; Wang, G.J.; Xie, J. Farnesoid X receptor is an effective target for modulating lipid accumulation in grass carp, Ctenopharyngodon idella. Aquaculture 2021, 534, 736248. [Google Scholar] [CrossRef]
- Guo, L.; Wang, L.; Liu, F.; Li, B.; Tang, Y.; Yu, S.; Zhang, D.; Huo, G. Effect of bile salt hydrolase-active Lactobacillus plantarum KLDS 1.0344 on cholesterol metabolism in rats fed a high-cholesterol diet. J. Funct. Foods 2019, 61, 103497. [Google Scholar] [CrossRef]
- Xu, H.; Zhang, Q.; Kim, S.K.; Liao, Z.; Wei, Y.; Sun, B.; Jia, L.; Chi, S.; Liang, M. Dietary taurine stimulates the hepatic biosynthesis of both bile acids and cholesterol in the marine teleost, tiger puffer (Takifugu rubripes). Br. J. Nutr. 2020, 123, 1345–1356. [Google Scholar] [CrossRef] [Green Version]
- Asadi, F.; Hallajian, A.; Asadian, P.; Shahriari, A.; Pourkabir, M. Serum lipid, free fatty acid, and proteins in juvenile sturgeons: Acipenser persicus and Acipenser stellatus. Comp. Clin. Pathol. 2009, 18, 287–289. [Google Scholar] [CrossRef]
- Yang, Q.; Liang, H.; Mokrani, A.; Ji, K.; Yu, H.; Ge, X.; Ren, M.; Xie, J.; Pan, L.; Sun, A. Dietary histidine affects intestinal antioxidant enzyme activities, antioxidant gene expressions and inflammatory factors in juvenile blunt snout bream (Megalobrama amblycephala). Aquac. Nutr. 2019, 25, 249–259. [Google Scholar] [CrossRef] [Green Version]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCT method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Wei, X.L.; Chen, L.P.; Chen, N.; Li, Y.H.; Wang, W.M.; Wang, H.L. Sequence analysis and expression differentiation of chemokine receptor CXCR4b among three populations of Megalobrama amblycephala. Dev. Comp. Immunol. 2013, 40, 195–201. [Google Scholar] [CrossRef]
- Zhou, C.; Liu, B.; Ge, X.; Xie, J.; Xu, P. Effect of dietary carbohydrate on the growth performance, immune response, hepatic antioxidant abilities and heat shock protein 70 expression of Wuchang bream, Megalobrama amblycephala. J. Appl. Ichthyol. 2013, 29, 1348–1356. [Google Scholar] [CrossRef]
- Xu, C.; Liu, W.B.; Shi, H.J.; Mi, H.F.; Li, X.F. Benfotiamine ameliorates high-carbohydrate diet-induced hepatic oxidative stress, inflammation and apoptosis in Megalobrama amblycephala. Aquac. Res. 2021, 52, 3174–3185. [Google Scholar] [CrossRef]
- Pang, J.; Xu, H.; Wang, X.; Chen, X.; Li, Q.; Liu, Q.; You, Y.; Zhang, H.; Xu, Z.; Zhao, Y. Resveratrol enhances trans-intestinal cholesterol excretion through selective activation of intestinal liver X receptor alpha. Biochem. Pharmacol. 2021, 186, 114481. [Google Scholar] [CrossRef]
- Zhang, Y.; Chen, M.L.; Zhou, Y.; Yi, L.; Gao, Y.X.; Ran, L.; Chen, S.H.; Zhang, T.; Zhou, X.; Zou, D. Resveratrol improves hepatic steatosis by inducing autophagy through the cAMP signaling pathway. Mol. Nutr. Food Res. 2015, 59, 1443–1457. [Google Scholar] [CrossRef] [PubMed]
- Yamasaki, S.; Tomihara, T.; Kimura, G.; Ueno, Y.; Ketema, R.M.; Sato, S.; Mukai, Y.; Sikder, T.; Kurasaki, M.; Hosokawa, T. Long-term effects of maternal resveratrol intake during lactation on cholesterol metabolism in male rat offspring. Int. J. Food Sci. Nutr. 2020, 71, 226–234. [Google Scholar] [CrossRef]
- Ou, J.; Tu, H.; Shan, B.; Luk, A.; DeBose-Boyd, R.A.; Bashmakov, Y.; Goldstein, J.L.; Brown, M.S. Unsaturated fatty acids inhibit transcription of the sterol regulatory element-binding protein-1c (SREBP-1c) gene by antagonizing ligand-dependent activation of the LXR. Proc. Natl. Acad. Sci. USA 2001, 98, 6027–6032. [Google Scholar] [CrossRef] [Green Version]
- Schultz, J.R.; Tu, H.; Luk, A.; Repa, J.J.; Medina, J.C.; Li, L.; Schwendner, S.; Wang, S.; Thoolen, M.; Mangelsdorf, D.J. Role of LXRs in control of lipogenesis. Genes Dev. 2000, 14, 2831–2838. [Google Scholar] [CrossRef]
- Gao, M.; Liu, D. Resveratrol suppresses T0901317-induced hepatic fat accumulation in mice. AAPS J. 2013, 15, 744–752. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dolezelova, E.; Prasnicka, A.; Cermanova, J.; Carazo, A.; Hyrsova, L.; Hroch, M.; Mokry, J.; Adamcova, M.; Mrkvicova, A.; Pavek, P. Resveratrol modifies biliary secretion of cholephilic compounds in sham-operated and cholestatic rats. World J. Gastroenterol. 2017, 23, 7678. [Google Scholar] [CrossRef] [PubMed]
- Chambers, K.F.; Day, P.E.; Aboufarrag, H.T.; Kroon, P.A. Polyphenol effects on cholesterol metabolism via bile acid biosynthesis, CYP7A1: A review. Nutrients 2019, 11, 2588. [Google Scholar] [CrossRef] [Green Version]
- Kamisako, T.; Ogawa, H.; Yamamoto, K. Effect of cholesterol, cholic acid and cholestyramine administration on the intestinal mRNA expressions related to cholesterol and bile acid metabolism in the rat. J. Gastroenterol. Hepatol. 2007, 22, 1832–1837. [Google Scholar] [CrossRef]
- Kemper, J.K.; Xiao, Z.; Ponugoti, B.; Miao, J.; Fang, S.; Kanamaluru, D.; Tsang, S.; Wu, S.Y.; Chiang, C.M.; Veenstra, T.D. FXR acetylation is normally dynamically regulated by p300 and SIRT1 but constitutively elevated in metabolic disease states. Cell Metab. 2009, 10, 392–404. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thomas, C.; Gioiello, A.; Noriega, L.; Strehle, A.; Oury, J.; Rizzo, G.; Macchiarulo, A.; Yamamoto, H.; Mataki, C.; Pruzanski, M. TGR5-mediated bile acid sensing controls glucose homeostasis. Cell Metab. 2009, 10, 167–177. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Ingredients (%) | Control | HC | HCR1 | HCR2 |
|---|---|---|---|---|
| Fish meal | 5.10 | 5.10 | 5.10 | 5.10 |
| Soybean meal | 29.59 | 29.59 | 29.59 | 29.59 |
| Rapeseed meal | 15.31 | 15.31 | 15.31 | 15.31 |
| Cottonseed meal | 15.31 | 15.31 | 15.31 | 15.31 |
| Fish oil | 2.04 | 2.04 | 2.04 | 2.04 |
| Soybean oil | 2.04 | 2.04 | 2.04 | 2.04 |
| Corn starch | 13.27 | 27.55 | 27.55 | 27.55 |
| Microcrystalline cellulose | 14.28 | 0.00 | 0.00 | 0.00 |
| Resveratrol | 0.00 | 0.00 | 0.04 | 0.08 |
| Calcium biphosphate | 2.04 | 2.04 | 2.04 | 2.04 |
| Premix 1 | 1.02 | 1.02 | 1.02 | 1.02 |
| Proximate composition (% air-dry basis) | ||||
| Moisture | 10.6 | 11.3 | 10.7 | 10.7 |
| Crude protein | 29.9 | 29.1 | 29.3 | 29.0 |
| Crude lipid | 5.3 | 5.2 | 5.4 | 5.5 |
| Ash | 6.6 | 6.5 | 6.4 | 6.6 |
| Crude fiber | 16.0 | 3.5 | 3.5 | 3.6 |
| Nitrogen-free extract 2 | 31.6 | 44.4 | 44.7 | 44.6 |
| Energy (MJ/kg) | 18.5 | 18.0 | 17.8 | 18.4 |
| Target Genes | Forward (5′-3′) | Reverse (5′-3′) | Accession Numbers |
|---|---|---|---|
| fxrα | CACCAAGAAACTGCCCAACG | CTGTGTCACTGAACGTCCCA | MZ506835 |
| lxrα | AACGTGCAGGATCACGAGTT | GCTTCATCAGCATACGGGGA | NC_063044.1 |
| cyp7a1 | ATATGATCAGGTGCCCTGCG | TCGTGCACAGCAAAGAAACG | XM_048164491.1 |
| cyp8b1 | AAACAGGACAGGGGGCAAAA | CCCTCGCGGATCTTGTACTC | NC_063044.1 |
| srebp2 | CTCAGCTTTCTGCCGGGTTA | TCACCGACCAATCACAGCTC | NC_063044.1 |
| hmgcr | CGCAGAAAAATGCTCACCCC | CATGGGGACCTGAAACTGCT | XM_048189198.1 |
| acat2 | CCCGAAGAAGGTTCGCCTTA | TCTTTCAGGCCCGTCACATC | XM_048206846.1 |
| ldlr | TCATTCCCCGCCTGAAGAAC | GGGTCGACCACTATAGCACG | XM_048158987.1 |
| cyp27a1 | AGGGTGACAGAGTCCCAACA | GCGGTCAGGTTTGAACTTTCG | NC_063044.1 |
| tgr5 | CCATCCTTTCCATTGCTGCG | CGCATTTCCAGTCTCCGTCT | NC_063044.1 |
| mrp2 | GTTCGATACCGGCCTGAGTT | CCAGTTCGACCAACAATGCC | NC_063044.1 |
| bsep | GGCCGTGAATCTACTAAGGTCA | GGCTCCTGAGACACAATTCCA | NC_063044.1 |
| ntcp | GATCCCAATACCGCAGGGAA | ATGCCCCTCCTGCTCTTAGT | NC_063044.1 |
| oatp1 | TCGGTCCAGTGTTTGGCTAC | TCCTGGGGTGATGCTAATGC | NC_063044.1 |
| oatp4 | GGGCAGCTACATGAAAAGCTC | ATCCAATCAGCGAGCTGGGAAT | NC_063044.1 |
| cat | CAGTGCTCCTGATACCCAGC | TTCTGACACAGACGCTCTCG | XM_048158628.1 |
| Cu/Zn-sod | AGTTGCCATGTGCACTTTTCT | AGGTGCTAGTCGAGTGTTAGG | KF479046.1 |
| Mn-sod | AGCTGCACCACAGCAAGCAC | TCCTCCACCATTCGGTGACA | KF195932.1 |
| gpx | GAACGCCCACCCTCTGTTTG | CGATGTCATTCCGGTTCACG | KF378713.1 |
| sirt1 | TCGGTTCATTCAGCAGCACA | ATGATGATCTGCCACAGCGT | MT518159.1 |
| keap1 | AATATCCGCCGGCTGTGTAG | TGAGTCCGAGGTGTTTCGTG | XM_048200093.1 |
| ef1α | CTTCTCAGGCTGACTGTGC | CCGCTAGCATTACCCTCC | X77689.1 |
| Parameters | Control | HC | HCR1 | HCR2 | Polynomial Contrasts | |
|---|---|---|---|---|---|---|
| Linear | Quadratic | |||||
| Initial weight (g) | 35.00 ± 0.15 | 34.97 ± 0.03 | 34.90 ± 0.17 | 35.03 ± 0.06 | ns | ns |
| Final weight (g) | 233.50 ± 28.50 | 217.17 ± 14.35 | 220.52 ± 20.52 | 204.40 ± 12.95 | ns | ns |
| WGR (%) 1 | 566.81 ± 86.07 | 521.13 ± 41.42 | 531.89 ± 58.73 | 483.42 ± 36.80 | ns | ns |
| SGR (%/day) 2 | 2.33 ± 0.16 | 2.25 ± 0.09 | 2.26 ± 0.12 | 2.17 ± 0.08 | ns | ns |
| Feed intake (g/fish) 3 | 298.40 ± 39.40 | 267.89 ± 10.60 | 281.51 ± 26.12 | 267.35 ± 11.13 | ns | ns |
| FCR 4 | 1.51 ± 0.02 | 1.48 ± 0.06 | 1.52 ± 0.05 | 1.59 ± 0.09 | ns | ns |
| PER 5 | 2.22 ± 0.03 | 2.34 ± 0.10 | 2.25 ± 0.07 | 2.19 ± 0.13 | ns | ns |
| HSI (%) 6 | 1.44 ± 0.07 | 1.79 ± 0.07 *b | 1.65 ± 0.05 ab | 1.54 ± 0.05 a | 0.04 | 0.017 |
| VSI (%) 7 | 9.58 ± 0.27 | 10.53 ± 0.31 * | 10.22 ± 0.44 | 10.22 ± 0.27 | ns | ns |
| IPF (%) 8 | 1.84 ± 0.15 | 2.68 ± 0.14 * | 2.86 ± 0.31 | 2.36 ± 0.20 | ns | ns |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ge, Y.; Zhang, L.; Chen, W.; Sun, M.; Liu, W.; Li, X. Resveratrol Modulates the Redox Response and Bile Acid Metabolism to Maintain the Cholesterol Homeostasis in Fish Megalobrama amblycephala Offered a High-Carbohydrate Diet. Antioxidants 2023, 12, 121. https://doi.org/10.3390/antiox12010121
Ge Y, Zhang L, Chen W, Sun M, Liu W, Li X. Resveratrol Modulates the Redox Response and Bile Acid Metabolism to Maintain the Cholesterol Homeostasis in Fish Megalobrama amblycephala Offered a High-Carbohydrate Diet. Antioxidants. 2023; 12(1):121. https://doi.org/10.3390/antiox12010121
Chicago/Turabian StyleGe, Yaping, Ling Zhang, Weiliang Chen, Miao Sun, Wenbin Liu, and Xiangfei Li. 2023. "Resveratrol Modulates the Redox Response and Bile Acid Metabolism to Maintain the Cholesterol Homeostasis in Fish Megalobrama amblycephala Offered a High-Carbohydrate Diet" Antioxidants 12, no. 1: 121. https://doi.org/10.3390/antiox12010121
APA StyleGe, Y., Zhang, L., Chen, W., Sun, M., Liu, W., & Li, X. (2023). Resveratrol Modulates the Redox Response and Bile Acid Metabolism to Maintain the Cholesterol Homeostasis in Fish Megalobrama amblycephala Offered a High-Carbohydrate Diet. Antioxidants, 12(1), 121. https://doi.org/10.3390/antiox12010121