
Genistein Promotes Anti-Heat Stress and Antioxidant Effects via the Coordinated Regulation of IIS, HSP, MAPK, DR, and Mitochondrial Pathways in Caenorhabditis elegans

Abstract
:1. Introduction
2. Materials and Methods
2.1. Materials
2.2. C. elegans Strains and Maintenance
2.3. Synchronization of C. elegans
2.4. Oxidative Stress and Heat Stress Test
2.5. Determination of Lipofuscin Accumulation in C. elegans
2.6. Determination of ROS in C. elegans
2.7. Survival Determination of Mutant Strains of EU1 [skn-1(zu67)] and MQ130[clk-1(qm30)] under Oxidative and Heat Stress Conditions
2.8. Observation of the Nuclear Translocation of daf-16::GFP
2.9. Quantification of hsp-16.2::GFP and skn-1::GFP Expression
2.10. Measurement of SOD Enzyme Activity
2.11. Extraction of C. elegans RNA
2.12. mRNA Analysis of Aging-Related Genes Using Quantitative Polymerase Chain Reaction (qPCR)
2.13. Data Treatment and Statistical Analyses
3. Results
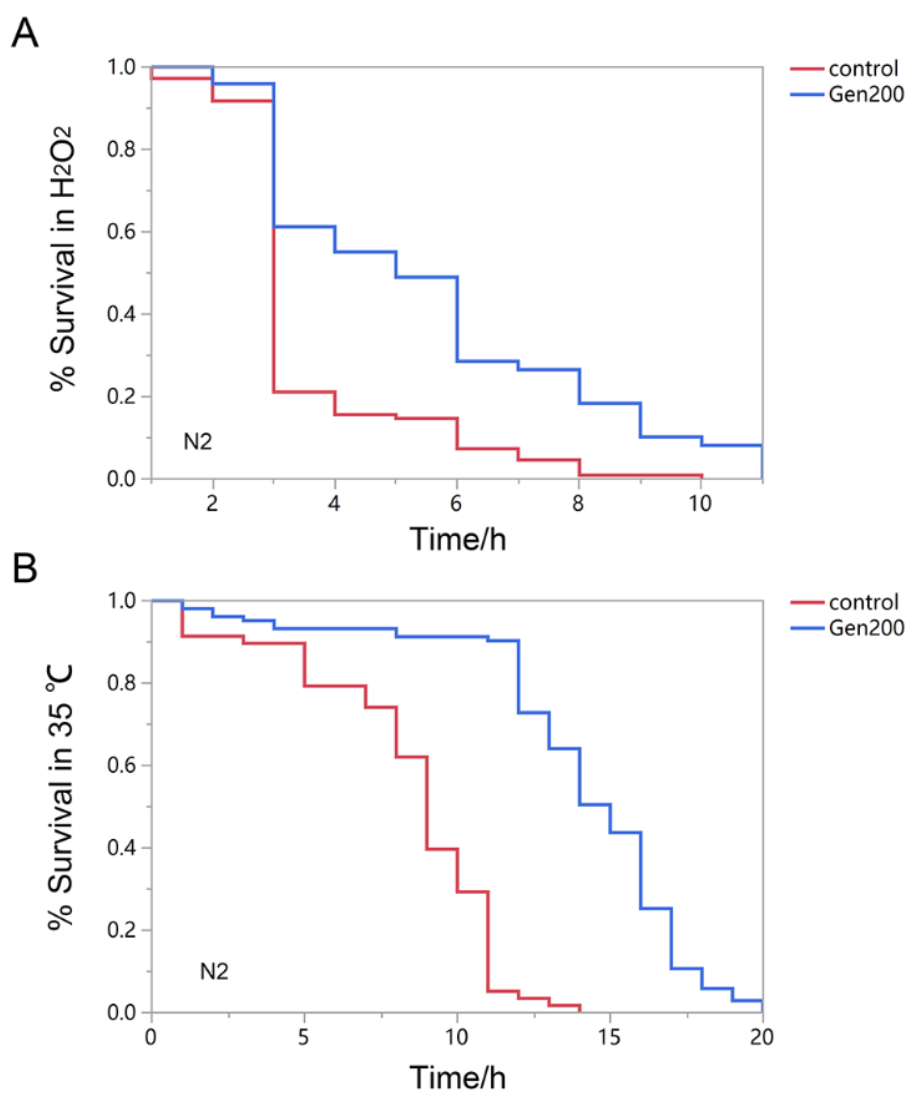
3.1. Genistein Improves Stress Resistance of Wild-Type Nematodes Exposed to Oxidative and Heat Stress
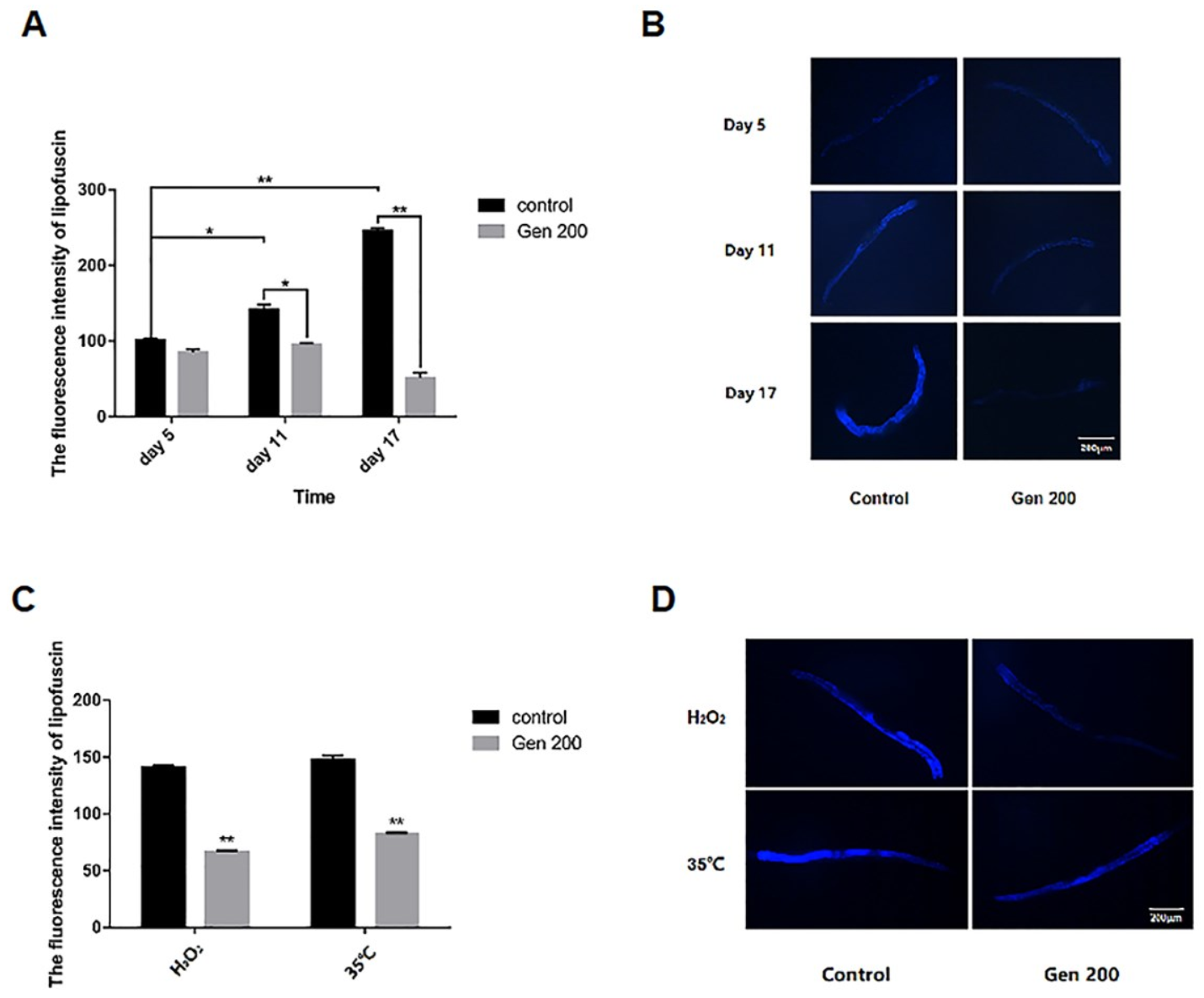
3.2. Genistein Lowered Lipofuscin Accumulation in C. elegans under Control, Heat and Oxidative Conditions
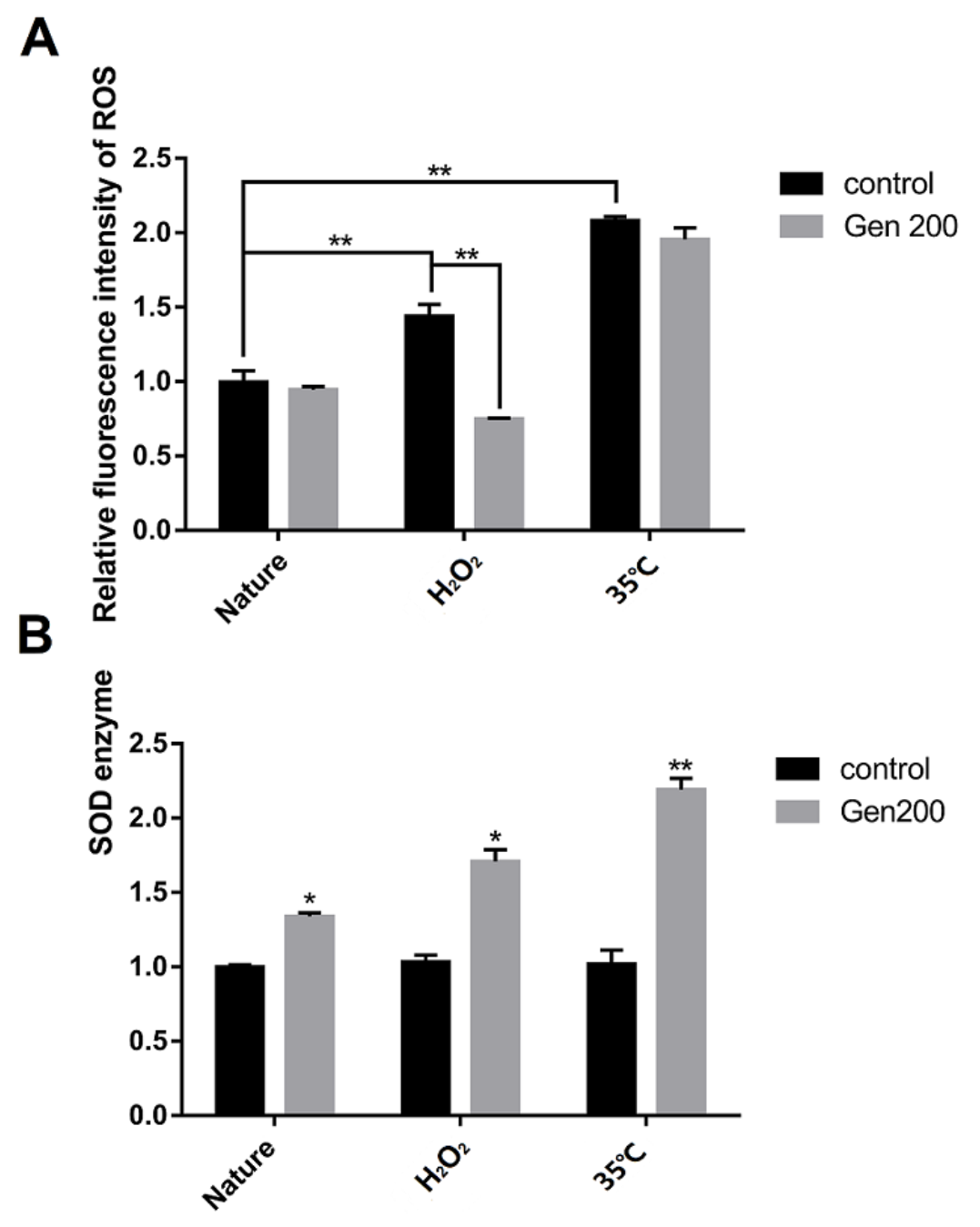
3.3. Genistein Lowered ROS Accumulation in H2O2-Treated Nematodes, and Increased the SOD Activity of Nematodes under Control, 35 °C and H2O2 Conditions
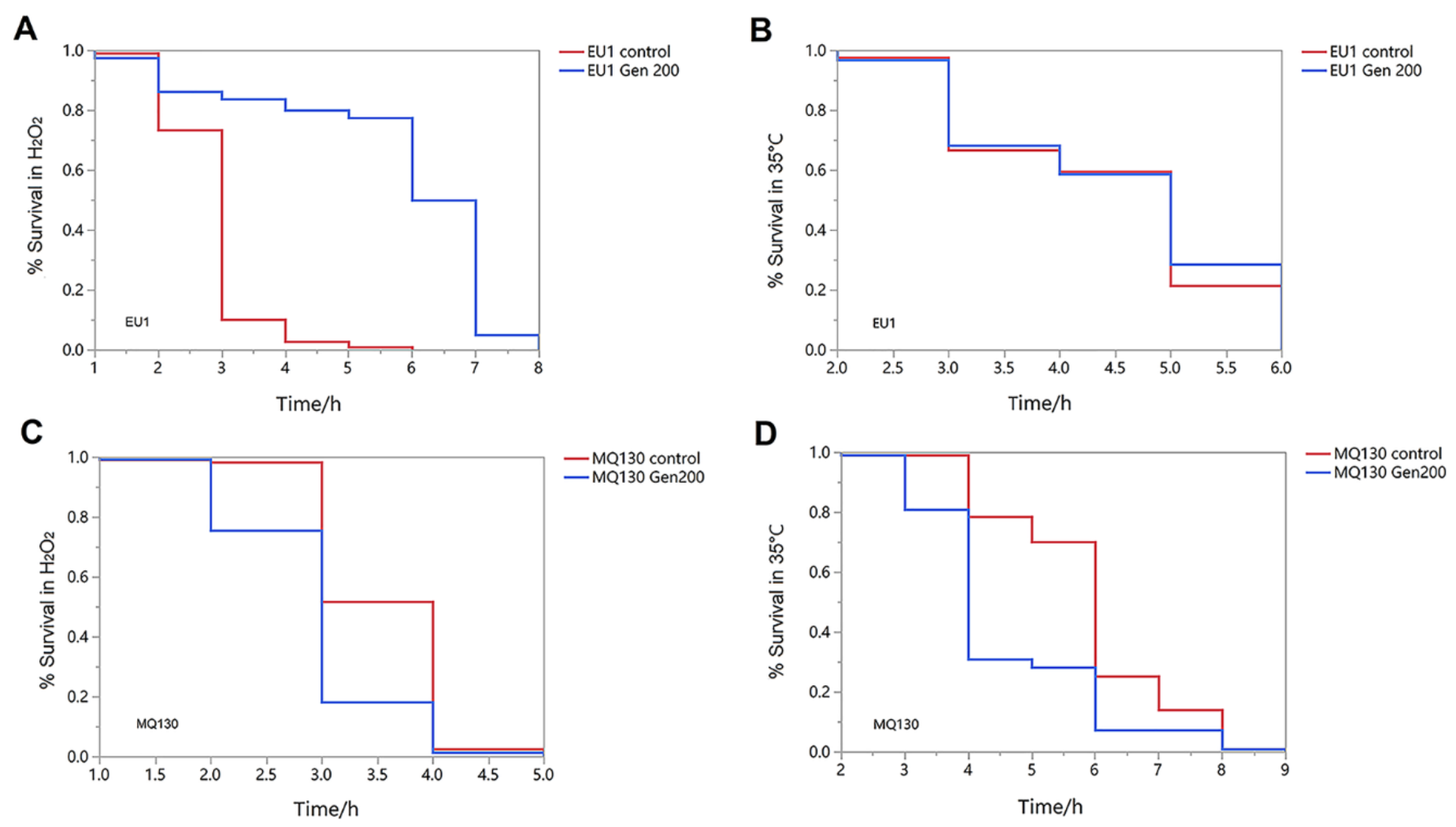
3.4. Genistein Partially Improved the Survival Rate of EU1 [skn-1(zu67)] Mutants, but It Did Not Significantly Change the Survival Rate of MQ130[clk-1(qm30)] Mutant under Heat and Oxidative Conditions
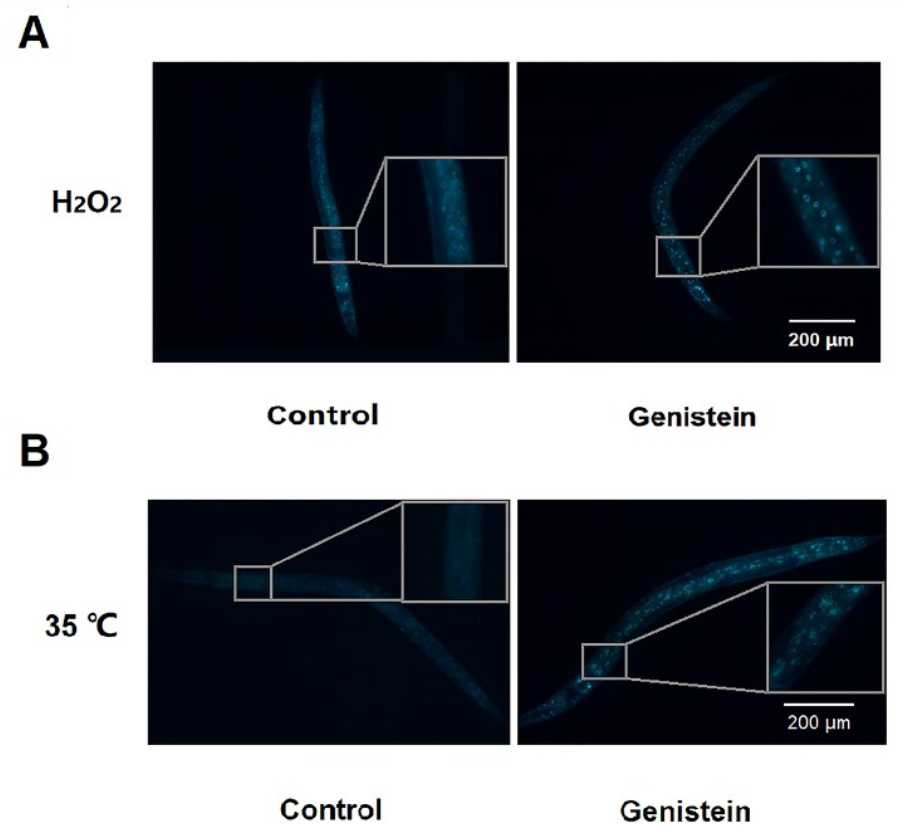
3.5. Genistein Increased the Nuclear Translocation of DAF-16 Transcription Factor under 35 °C and H2O2 Conditions
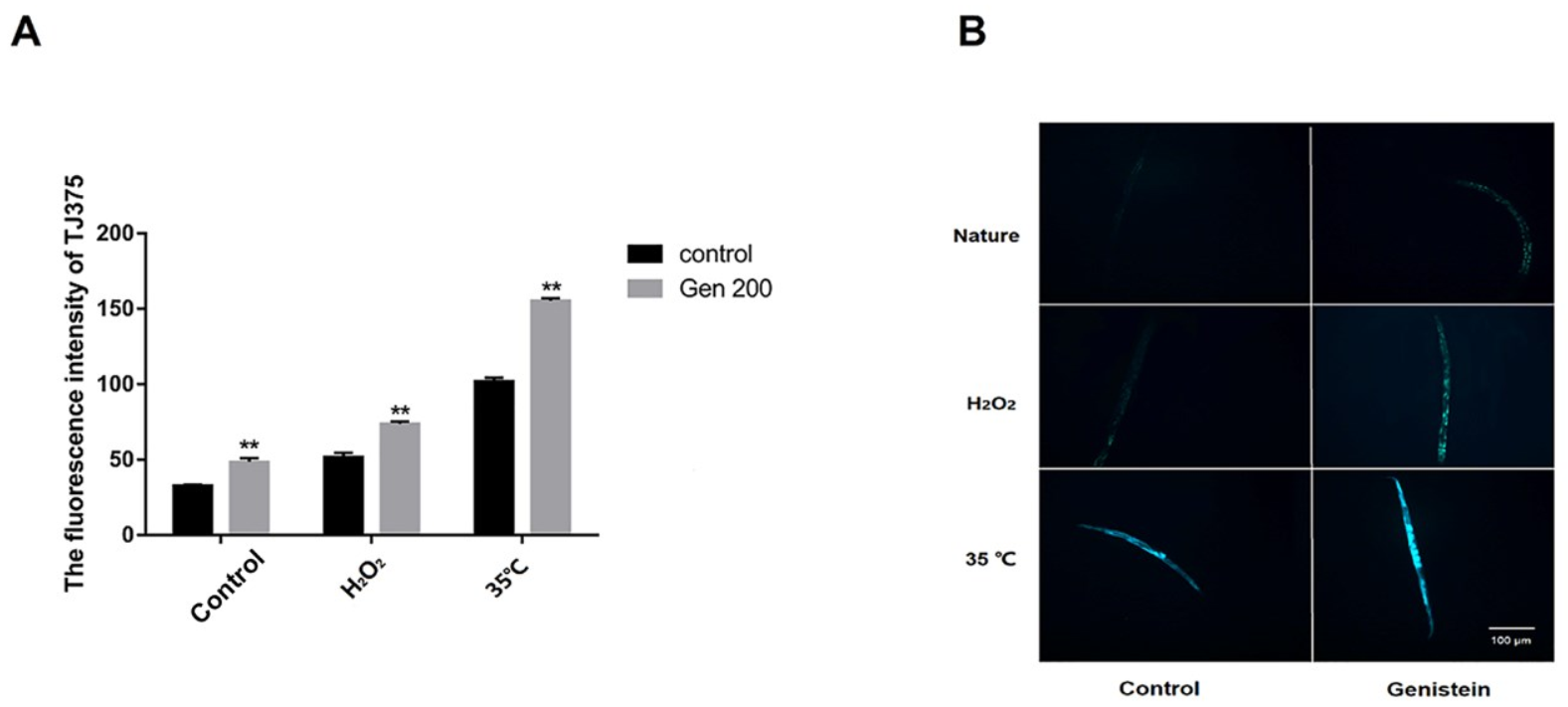
3.6. Genistein Enhanced the Accumulation of HSP-16.2 Protein in Nematodes under Control, 35 °C, and H2O2 Conditions
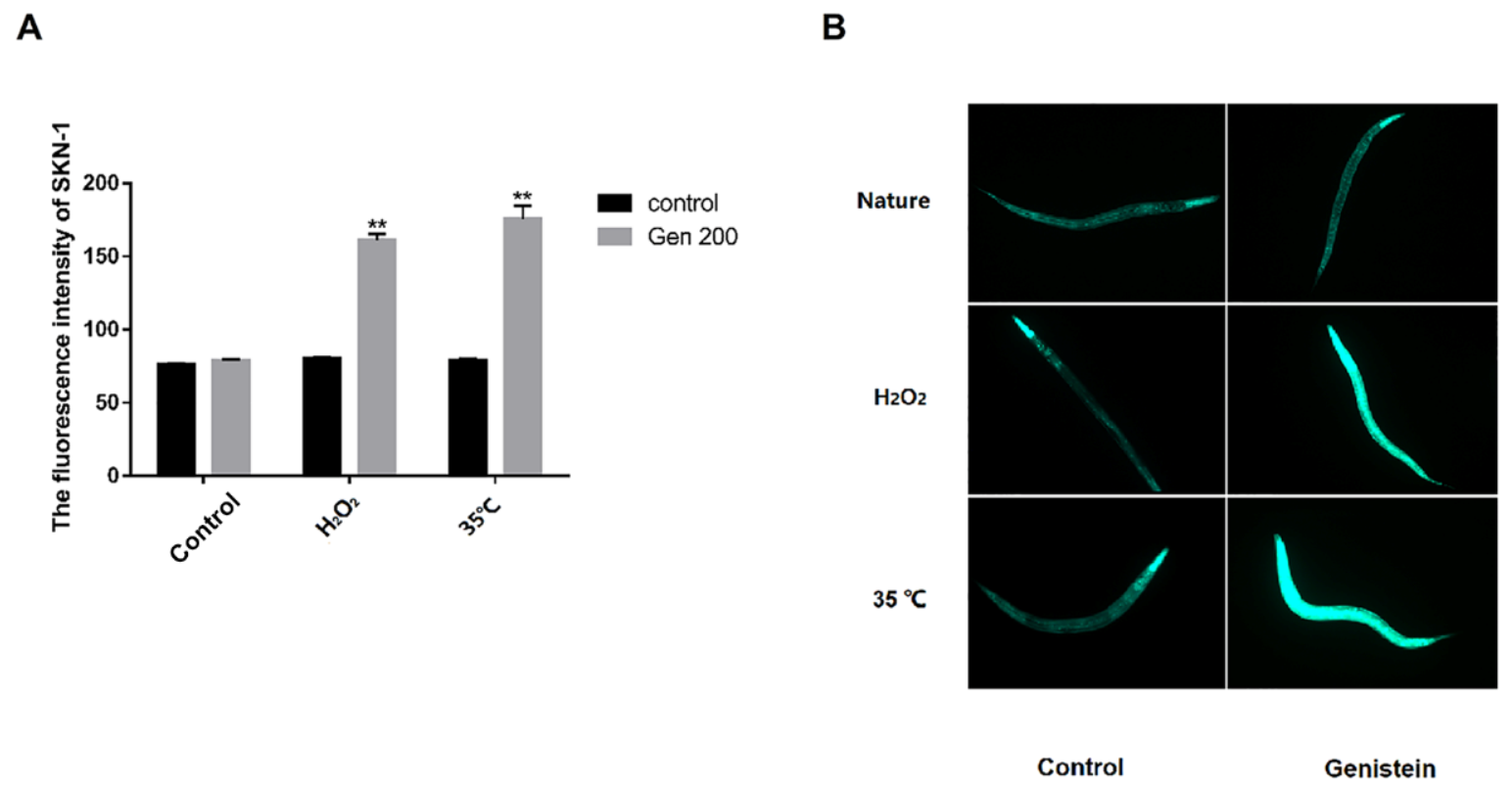
3.7. Genistein Enhanced the Accumulation of SKN-1 Protein in Nematodes under 35 °C and H2O2 Conditions
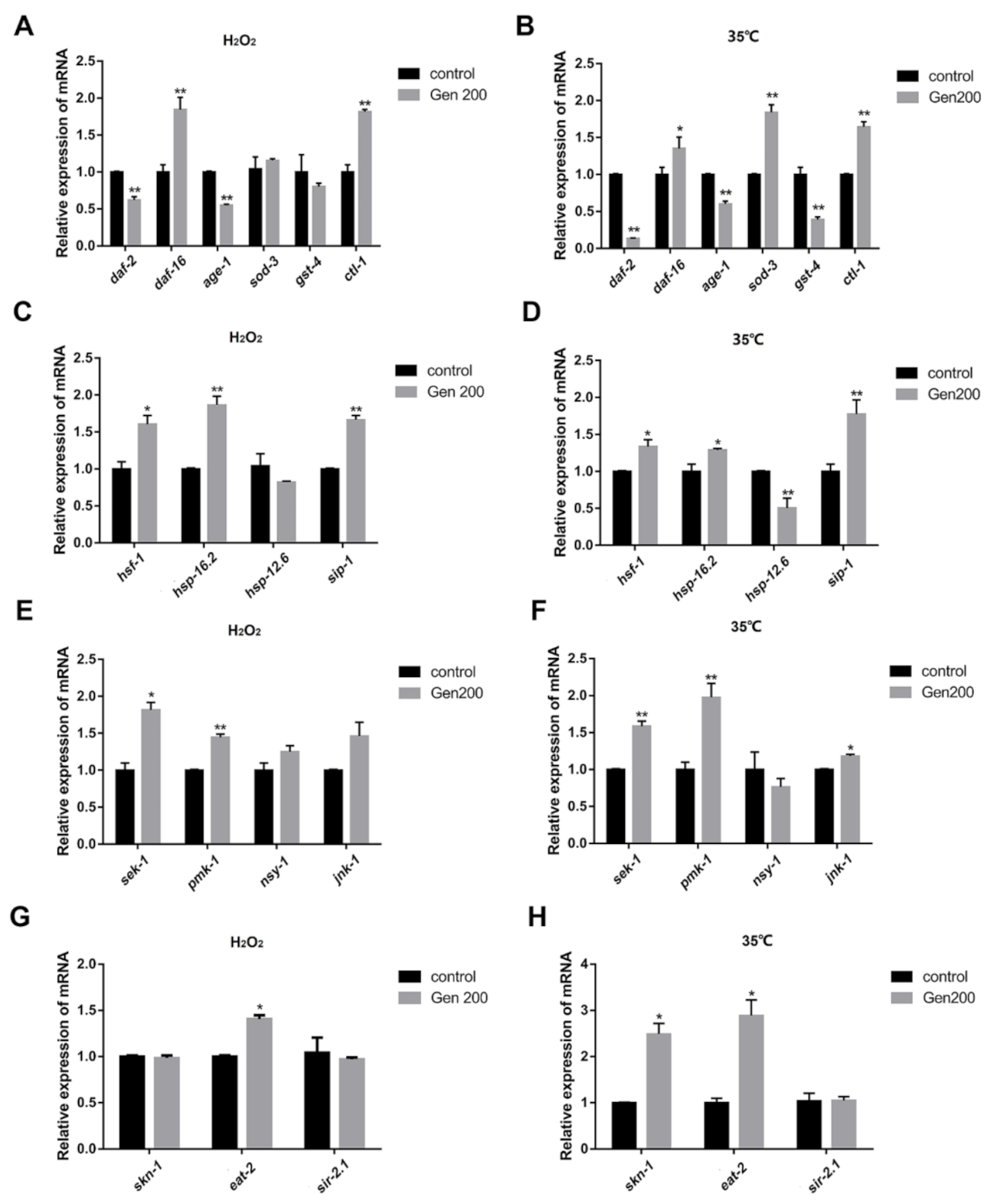
3.8. Genistein Regulated the Expression of Messenger RNA (mRNA) in C. elegans under 35 °C and H2O2 Conditions
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
References
- da Costa, J.P.; Vitorino, R.; Silva, G.M.; Vogel, C.; Duarte, A.C.; Rocha-Santos, T. A synopsis on aging-Theories, mechanisms and future prospects. Ageing Res. Rev. 2016, 29, 90–112. [Google Scholar] [CrossRef] [PubMed]
- Wickens, A.P. Ageing and the free radical theory. Respir. Physiol. 2001, 128, 379–391. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Davies, K.J.A.; Forman, H.J. Oxidative stress response and Nrf2 signaling in aging. Free Radic. Biol. Med. 2015, 88, 314–336. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Muñoz, M.J. Longevity and heat stress regulation in Caenorhabditis elegans. Mech. Ageing Dev. 2003, 124, 43–48. [Google Scholar] [CrossRef] [PubMed]
- Jin, K. New perspectives on healthy aging. Prog. Neurobiol. 2017, 157, 1. [Google Scholar] [CrossRef] [PubMed]
- Kim, B.; Lee, J.; Kim, Y.; Lee, S.V. Regulatory systems that mediate the effects of temperature on the lifespan of Caenorhabditis elegans. J. Neurogenet. 2020, 34, 518–526. [Google Scholar] [CrossRef]
- Senoner, T.; Dichtl, W. Oxidative Stress in Cardiovascular Diseases: Still a Therapeutic Target? Nutrients 2019, 11, 2090. [Google Scholar] [CrossRef] [Green Version]
- Schmitt-Schillig, S.; Schaffer, S.; Weber, C.C.; Eckert, G.P.; Müller, W.E. Flavonoids and the aging brain. J. Physiol. Pharmacol. 2005, 56 (Suppl. 1), 23–36. [Google Scholar]
- Mukherjee, P.K.; Banerjee, S.; Biswas, S.; Das, B.; Kar, A.; Katiyar, C.K. Withania somnifera (L.) Dunal—Modern perspectives of an ancient Rasayana from Ayurveda. J. Ethnopharmacol. 2021, 264, 113157. [Google Scholar] [CrossRef]
- Spagnuolo, C.; Russo, G.L.; Orhan, I.E.; Habtemariam, S.; Daglia, M.; Sureda, A.; Nabavi, S.F.; Devi, K.P.; Loizzo, M.R.; Tundis, R.; et al. Genistein and cancer: Current status, challenges, and future directions. Adv. Nutr. 2015, 6, 408–419. [Google Scholar] [CrossRef] [Green Version]
- Martiniakova, M.; Babikova, M.; Omelka, R. Pharmacological agents and natural compounds: Available treatments for osteoporosis. J. Physiol. Pharmacol. 2020, 71, 307–320. [Google Scholar]
- Křížová, L.; Dadáková, K.; Kašparovská, J.; Kašparovský, T. Isoflavones. Molecules 2019, 24, 1076. [Google Scholar] [CrossRef]
- Thangavel, P.; Puga-Olguín, A.; Rodríguez-Landa, J.F.; Zepeda, R.C. Genistein as Potential Therapeutic Candidate for Menopausal Symptoms and Other Related Diseases. Molecules 2019, 24, 3892. [Google Scholar] [CrossRef] [Green Version]
- Vona, R.; Gambardella, L.; Cittadini, C.; Straface, E.; Pietraforte, D. Biomarkers of Oxidative Stress in Metabolic Syndrome and Associated Diseases. Oxidative Med. Cell. Longev. 2019, 2019, 8267234. [Google Scholar] [CrossRef] [Green Version]
- Wang, L.; Waltenberger, B.; Pferschy-Wenzig, E.M.; Blunder, M.; Liu, X.; Malainer, C.; Blazevic, T.; Schwaiger, S.; Rollinger, J.M.; Heiss, E.H.; et al. Natural product agonists of peroxisome proliferator-activated receptor gamma (PPARγ): A review. Biochem. Pharmacol. 2014, 92, 73–89. [Google Scholar] [CrossRef] [Green Version]
- Murphy, C.T.; Hu, P.J. Insulin/insulin-like growth factor signaling in C. elegans. WormBook 2018, 1–43. [Google Scholar] [CrossRef] [Green Version]
- Zhang, S.; Li, F.; Zhou, T.; Wang, G.; Li, Z. Caenorhabditis elegans as a Useful Model for Studying Aging Mutations. Front. Endocrinol. 2020, 11, 554994. [Google Scholar] [CrossRef]
- Zečić, A.; Braeckman, B.P. DAF-16/FoxO in Caenorhabditis elegans and Its Role in Metabolic Remodeling. Cells 2020, 9, 109. [Google Scholar] [CrossRef] [Green Version]
- Seo, K.; Choi, E.; Lee, D.; Jeong, D.E.; Jang, S.K.; Lee, S.J. Heat shock factor 1 mediates the longevity conferred by inhibition of TOR and insulin/IGF-1 signaling pathways in C. elegans. Aging Cell 2013, 12, 1073–1081. [Google Scholar] [CrossRef]
- Yan, F.; Chen, Y.; Azat, R.; Zheng, X. Mulberry Anthocyanin Extract Ameliorates Oxidative Damage in HepG2 Cells and Prolongs the Lifespan of Caenorhabditis elegans through MAPK and Nrf2 Pathways. Oxidative Med. Cell. Longev. 2017, 2017, 7956158. [Google Scholar] [CrossRef] [Green Version]
- Lakowski, B.; Hekimi, S. The genetics of caloric restriction in Caenorhabditis elegans. Proc. Natl. Acad. Sci. USA 1998, 95, 13091–13096. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Monaghan, R.M.; Barnes, R.G.; Fisher, K.; Andreou, T.; Rooney, N.; Poulin, G.B.; Whitmarsh, A.J. A nuclear role for the respiratory enzyme CLK-1 in regulating mitochondrial stress responses and longevity. Nat. Cell Biol. 2015, 17, 782–792. [Google Scholar] [CrossRef] [PubMed]
- Qi, J.H.; Dong, F.X. The relevant targets of anti-oxidative stress: A review. J. Drug Target. 2021, 29, 677–686. [Google Scholar] [CrossRef] [PubMed]
- Zhou, W.; Lin, J.M.; Wang, S.L.; Lin, H.Z. Effect of Herba Scutellariae Barbatae flavonoids in delaying aging of Caenorhabditis elegans and human umbilical vein endothelial cells in vitro. Nan Fang Yi Ke Da Xue Xue Bao 2017, 37, 821–826. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Q.; Qu, Y.; Zhou, X.G.; Chen, J.N.; Luo, H.R.; Wu, G.S. A Dihydroflavonoid Naringin Extends the Lifespan of C. elegans and Delays the Progression of Aging-Related Diseases in PD/AD Models via DAF-16. Oxidative Med. Cell. Longev. 2020, 2020, 6069354. [Google Scholar] [CrossRef]
- Guo, H.; Cao, M.; Zou, S.; Ye, B.; Dong, Y. Cranberry Extract Standardized for Proanthocyanidins Alleviates β-Amyloid Peptide Toxicity by Improving Proteostasis Through HSF-1 in Caenorhabditis elegans Model of Alzheimer’s Disease. J. Gerontol. A Biol. Sci. Med. Sci. 2016, 71, 1564–1573. [Google Scholar] [CrossRef] [Green Version]
- Lee, E.B.; Ahn, D.; Kim, B.J.; Lee, S.Y.; Seo, H.W.; Cha, Y.S.; Jeon, H.; Eun, J.S.; Cha, D.S.; Kim, D.K. Genistein from Vigna angularis Extends Lifespan in Caenorhabditis elegans. Biomol. Ther. 2015, 23, 77–83. [Google Scholar] [CrossRef] [Green Version]
- Li, K.; Zhang, S.; Sun, Y.; Chen, Y.; Chen, W.; Ruan, W.; Liu, Y. Antiamyloid β toxicity effect of genistein via activation of DAF-16 and HSP-16.2 signal pathways in Caenorhabditis elegans. J. Biochem. Mol. Toxicol. 2022, 36, e23055. [Google Scholar] [CrossRef]
- Wu, Z.; Tan, B.; Liu, Y.; Dunn, J.; Martorell Guerola, P.; Tortajada, M.; Cao, Z.; Ji, P. Chemical Composition and Antioxidant Properties of Essential Oils from Peppermint, Native Spearmint and Scotch Spearmint. Molecules 2019, 24, 2825. [Google Scholar] [CrossRef] [Green Version]
- Doshi, S.; Braganza, V. Ameliorative effect of Argyreia boseana Sant. & Pat. on stress in C. elegans. J. Ayurveda Integr. Med. 2020, 11, 147–152. [Google Scholar]
- Phulara, S.C.; Pandey, S.; Jha, A.; Chauhan, P.S.; Gupta, P.; Shukla, V. Hemiterpene Compound, 3,3-Dimethylallyl Alcohol Promotes Longevity and Neuroprotection in Caenorhabditis Elegans. Geroscience 2021, 43, 791–807. [Google Scholar] [CrossRef]
- Zhang, H.; Yang, X.; Pang, X.; Zhao, Z.; Yu, H.; Zhou, H. Genistein protects against ox-LDL-induced senescence through enhancing SIRT1/LKB1/AMPK-mediated autophagy flux in HUVECs. Mol. Cell. Biochem. 2019, 455, 127–134. [Google Scholar] [CrossRef]
- Lee, K.Y.; Kim, J.R.; Choi, H.C. Genistein-induced LKB1-AMPK activation inhibits senescence of VSMC through autophagy induction. Vasc. Pharmacol. 2016, 81, 75–82. [Google Scholar] [CrossRef]
- Wang, H.; Liu, J.; Li, T.; Liu, R.H. Blueberry extract promotes longevity and stress tolerance via DAF-16 in Caenorhabditis elegans. Food Funct. 2018, 9, 5273–5282. [Google Scholar] [CrossRef]
- Song, B.; Zheng, B.; Li, T.; Liu, R.H. Raspberry extract promoted longevity and stress tolerance via the insulin/IGF signaling pathway and DAF-16 in Caenorhabditis elegans. Food Funct. 2020, 11, 3598–3609. [Google Scholar] [CrossRef]
- Vatner, S.F.; Zhang, J.; Oydanich, M.; Berkman, T.; Naftalovich, R.; Vatner, D.E. Healthful aging mediated by inhibition of oxidative stress. Ageing Res. Rev. 2020, 64, 101194. [Google Scholar] [CrossRef]
- Kittimongkolsuk, P.; Roxo, M.; Li, H.; Chuchawankul, S.; Wink, M.; Tencomnao, T. Extracts of the Tiger Milk Mushroom (Lignosus Rhinocerus) Enhance Stress Resistance and Extend Lifespan in Caenorhabditis Elegans Via the Daf-16/Foxo Signaling Pathway. Pharmaceuticals. 2021, 14, 93. [Google Scholar] [CrossRef]
- Lee, E.B.; Kim, J.H.; An, C.W.; Kim, Y.J.; Noh, Y.J.; Kim, S.J.; Kim, J.E.; Shrestha, A.C.; Ham, H.N.; Leem, J.Y.; et al. Longevity and Stress Resistant Property of 6-Gingerol from Zingiber officinale Roscoe in Caenorhabditis elegans. Biomol. Ther. 2018, 26, 568–575. [Google Scholar] [CrossRef]
- Pincus, Z.; Mazer, T.C.; Slack, F.J. Autofluorescence as a measure of senescence in C. elegans: Look to red, not blue or green. Aging 2016, 8, 889–898. [Google Scholar] [CrossRef] [Green Version]
- Yang, X.; Wang, H.; Li, T.; Chen, L.; Zheng, B.; Liu, R.H. Nobiletin Delays Aging and Enhances Stress Resistance of Caenorhabditis elegans. Int. J. Mol. Sci. 2020, 21, 341. [Google Scholar] [CrossRef] [Green Version]
- Grönke, S.; Clarke, D.F.; Broughton, S.; Andrews, T.D.; Partridge, L. Molecular evolution and functional characterization of Drosophila insulin-like peptides. PLoS Genet. 2010, 6, e1000857. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, L.; Liu, J.; Bu, L.L.; Liao, D.F.; Cheng, S.W.; Zheng, X.L. Curcumin Acetylsalicylate Extends the Lifespan of Caenorhabditis elegans. Molecules 2021, 26, 6609. [Google Scholar] [CrossRef] [PubMed]
- Vanfleteren, J.R.; Braeckman, B.P. Mechanisms of life span determination in Caenorhabditis elegans. Neurobiol. Aging 1999, 20, 487–502. [Google Scholar] [CrossRef] [PubMed]
- Lin, K.; Hsin, H.; Libina, N.; Kenyon, C. Regulation of the Caenorhabditis elegans longevity protein DAF-16 by insulin/IGF-1 and germline signaling. Nat. Genet. 2001, 28, 139–145. [Google Scholar] [CrossRef] [PubMed]
- Ke, T.; Antunes Soares, F.A.; Santamaría, A.; Bowman, A.B.; Skalny, A.V.; Aschner, M. N,N’ bis-(2-mercaptoethyl) isophthalamide induces developmental delay in Caenorhabditis elegans by promoting DAF-16 nuclear localization. Toxicol. Rep. 2020, 7, 930–937. [Google Scholar] [CrossRef] [PubMed]
- Baird, N.A.; Douglas, P.M.; Simic, M.S.; Grant, A.R.; Moresco, J.J.; Wolff, S.C.; Yates, J.R.; Manning, G.; Dillin, A. HSF-1–mediated cytoskeletal integrity determines thermotolerance and life span. Science 2014, 346, 360–363. [Google Scholar] [CrossRef] [Green Version]
- Jagdale, G.B.; Grewal, P.S.; Salminen, S.O. Both heat-shock and cold-shock influence trehalose metabolism in an entomopathogenic nematode. J. Parasitol. 2005, 91, 988–994. [Google Scholar] [CrossRef]
- Lin, C.; Chen, Y.; Lin, Y.; Wang, X.; Hu, L.; Cao, Y.; Chen, Y. Antistress and anti-aging activities of Caenorhabditis elegans were enhanced by Momordica saponin extract. Eur. J. Nutr. 2021, 60, 1819–1832. [Google Scholar] [CrossRef]
- Prithika, U.; Deepa, V.; Balamurugan, K. External induction of heat shock stimulates the immune response and longevity of Caenorhabditis elegans towards pathogen exposure. Innate Immun. 2016, 22, 466–478. [Google Scholar] [CrossRef] [Green Version]
- Kumsta, C.; Chang, J.T.; Schmalz, J.; Hansen, M. Hormetic heat stress and HSF-1 induce autophagy to improve survival and proteostasis in C. elegans. Nat. Commun. 2017, 8, 14337. [Google Scholar] [CrossRef] [Green Version]
- Sugawara, T.; Sakamoto, K. Killed Bifidobacterium longum enhanced stress tolerance and prolonged life span of Caenorhabditis elegans via DAF-16. Br. J. Nutr. 2018, 120, 872–880. [Google Scholar] [CrossRef] [Green Version]
- Smita, S.S.; Trivedi, S.; Pandey, T.; Trivedi, M.; Pandey, R. A Bioactive compound Shatavarin IV-mediated longevity as revealed by dietary restriction-induced autophagy in Caenorhabditis elegans. Biogerontology 2020, 21, 827–844. [Google Scholar] [CrossRef]
- Snutch, T.P.; Heschl, M.F.; Baillie, D.L. The Caenorhabditis elegans hsp70 gene family: A molecular genetic characterization. Gene 1988, 64, 241–255. [Google Scholar] [CrossRef]
- Hsu, A.L.; Murphy, C.T.; Kenyon, C. Regulation of aging and age-related disease by DAF-16 and heat-shock factor. Science 2003, 300, 1142–1145. [Google Scholar] [CrossRef] [Green Version]
- Linder, B.; Jin, Z.; Freedman, J.H.; Rubin, C.S. Molecular characterization of a novel, developmentally regulated small embryonic chaperone from Caenorhabditis elegans. J. Biol. Chem. 1996, 271, 30158–30166. [Google Scholar] [CrossRef] [Green Version]
- Walther, D.M.; Kasturi, P.; Zheng, M.; Pinkert, S.; Vecchi, G.; Ciryam, P.; Morimoto, R.I.; Dobson, C.M.; Vendruscolo, M.; Mann, M.; et al. Widespread Proteome Remodeling and Aggregation in Aging C. elegans. Cell 2017, 168, 944. [Google Scholar] [CrossRef] [Green Version]
- Deonarine, A.; Walker, M.W.G.; Westerheide, S.D. HSF-1 displays nuclear stress body formation in multiple tissues in Caenorhabditis elegans upon stress and following the transition to adulthood. Cell Stress Chaperones 2021, 26, 417–431. [Google Scholar] [CrossRef]
- Prasanth, M.I.; Gayathri, S.; Bhaskar, J.P.; Krishnan, V.; Balamurugan, K. Understanding the role of p38 and JNK mediated MAPK pathway in response to UV-A induced photoaging in Caenorhabditis elegans. J. Photochem. Photobiol. B 2020, 205, 111844. [Google Scholar] [CrossRef]
- Radi, E.; Formichi, P.; Battisti, C.; Federico, A. Apoptosis and oxidative stress in neurodegenerative diseases. J. Alzheimer’s Dis. 2014, 42 (Suppl. 3), S125–S152. [Google Scholar] [CrossRef] [Green Version]
- Davalli, P.; Mitic, T.; Caporali, A.; Lauriola, A.; D’Arca, D. ROS, Cell Senescence, and Novel Molecular Mechanisms in Aging and Age-Related Diseases. Oxidative Med. Cell. Longev. 2016, 2016, 3565127. [Google Scholar] [CrossRef] [Green Version]
- Kawasaki, M.; Hisamoto, N.; Iino, Y.; Yamamoto, M.; Ninomiya-Tsuji, J.; Matsumoto, K. A Caenorhabditis elegans JNK signal transduction pathway regulates coordinated movement via type-D GABAergic motor neurons. EMBO J. 1999, 18, 3604–3615. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sagasti, A.; Hisamoto, N.; Hyodo, J.; Tanaka-Hino, M.; Matsumoto, K.; Bargmann, C.I. The CaMKII UNC-43 activates the MAPKKK NSY-1 to execute a lateral signaling decision required for asymmetric olfactory neuron fates. Cell 2001, 105, 221–232. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, J.; Du, H.; Nie, Y.; Wang, Y.; Dai, H.; Wang, M.; Wang, D.; Xu, A. Mitochondria and MAPK cascades modulate endosulfan-induced germline apoptosis in Caenorhabditis elegans. Toxicol. Res. 2017, 6, 412–419. [Google Scholar] [CrossRef] [PubMed]
- Berman, K.; Mckay, J.; Avery, L.; Cobb, M. Isolation and characterization of pmk-(1-3): Three p38 homologs in Caenorhabditis elegans. Mol. Cell Biol. Res. Commun. 2001, 4, 337–344. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Blackwell, T.K.; Steinbaugh, M.J.; Hourihan, J.M.; Ewald, C.Y.; Isik, M. SKN-1/Nrf, stress responses, and aging in Caenorhabditis elegans. Free Radic Biol. Med. 2015, 88, 290–301. [Google Scholar] [CrossRef] [Green Version]
- Ichijo, H.; Nishida, E.; Irie, K.; ten Dijke, P.; Saitoh, M.; Moriguchi, T.; Takagi, M.; Matsumoto, K.; Miyazono, K.; Gotoh, Y. Induction of apoptosis by ASK1, a mammalian MAPKKK that activates SAPK/JNK and p38 signaling pathways. Science 1997, 275, 90–94. [Google Scholar] [CrossRef]
- Back, P.; Matthijssens, F.; Vlaeminck, C.; Braeckman, B.P.; Vanfleteren, J.R. Effects of sod gene overexpression and deletion mutation on the expression profiles of reporter genes of major detoxification pathways in Caenorhabditis elegans. Exp. Gerontol. 2010, 45, 603–610. [Google Scholar] [CrossRef]
- Wu, C.W.; Deonarine, A.; Przybysz, A.; Strange, K.; Choe, K.P. The Skp1 Homologs SKR-1/2 Are Required for the Caenorhabditis elegans SKN-1 Antioxidant/Detoxification Response Independently of p38 MAPK. PLoS Genet. 2016, 12, e1006361. [Google Scholar] [CrossRef] [Green Version]
- Buccitelli, C.; Selbach, M. mRNAs, proteins and the emerging principles of gene expression control. Nat. Rev. Genet. 2020, 21, 630–644. [Google Scholar] [CrossRef]
- Kobayashi, Y.; Furukawa-Hibi, Y.; Chen, C.; Horio, Y.; Isobe, K.; Ikeda, K.; Motoyama, N. SIRT1 is critical regulator of FOXO-mediated transcription in response to oxidative stress. Int. J. Mol. Med. 2005, 16, 237–243. [Google Scholar] [CrossRef]
- Viswanathan, M.; Kim, S.K.; Berdichevsky, A.; Guarente, L. A role for SIR-2.1 regulation of ER stress response genes in determining C. elegans life span. Dev. Cell 2005, 9, 605–615. [Google Scholar] [CrossRef] [Green Version]
- Berdichevsky, A.; Viswanathan, M.; Horvitz, H.R.; Guarente, L. C. elegans SIR-2.1 interacts with 14-3-3 proteins to activate DAF-16 and extend life span. Cell 2006, 125, 1165–1177. [Google Scholar] [CrossRef] [Green Version]
- Vajo, Z.; King, L.M.; Jonassen, T.; Wilkin, D.J.; Ho, N.; Munnich, A.; Clarke, C.F.; Francomano, C.A. Conservation of the Caenorhabditis elegans timing gene clk-1 from yeast to human: A gene required for ubiquinone biosynthesis with potential implications for aging. Mamm. Genome 1999, 10, 1000–1004. [Google Scholar] [CrossRef]
- Yanase, S.; Yasuda, K.; Ishii, N. Interaction between the ins/IGF-1 and p38 MAPK signaling pathways in molecular compensation of sod genes and modulation related to intracellular ROS levels in C. elegans. Biochem. Biophys. Rep. 2020, 23, 100796. [Google Scholar] [CrossRef]
- Lu, L.; Zhao, X.; Zhang, J.; Li, M.; Qi, Y.; Zhou, L. Calycosin promotes lifespan in Caenorhabditis elegans through insulin signaling pathway via daf-16, age-1 and daf-2. J. Biosci. Bioeng. 2017, 124, 1–7. [Google Scholar] [CrossRef]
- Tullet, J.M.; Hertweck, M.; An, J.H.; Baker, J.; Hwang, J.Y.; Liu, S.; Oliveira, R.P.; Baumeister, R.; Blackwell, T.K. Direct inhibition of the longevity-promoting factor SKN-1 by insulin-like signaling in C. elegans. Cell 2008, 132, 1025–1038. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Genes | Forward Primer | Reverse Primer |
|---|---|---|
| daf-16(NM_001381205.1) | AGGAGTCGAAGCCGATTAAGAC | GGTAGTGGCATTGGCTTGAAG |
| daf-2(NM_065249.7) | TACTTGAATCGGGCGTCGTT | GACGACTTCAACAACCGCTG |
| age-1(NM_064061.6) | CTCCTGAACCGACTGCCAAT | AAATGCGAGTTCGGAGAGCA |
| gst-4(NM_069447.8) | CCCATTTTACAAGTCGATGG | CTTCCTCTGCAGTTTTTCCA |
| sod-3(NM_078363.9) | TGGCTAAGGATGGTGGAGAA | GCCTTGAACCGCAATAGTGAT |
| ctl-1 (NM_064578.6) | CGATACCGTACTCGTGATGAT | CCAAACAGCCACCCAAATCA |
| skn-1(NM_171345.6) | CGTCCAACCAACCACATCATCTC | ATCTTCCAATTCGGCTTTT |
| sir-2.1(NM_001268555.5) | AAATCTTCCCAGGACAGTTCGTA | ATGGGCAACACGCATAGCA |
| eat-2(NM_064558.6) | ACCATGGGGAATTTGCAACG | GGATTTGCGTGAGGGGTATGA |
| hsp-16.2(NM_001392482.1) | CTGCAGAATCTCTCCATCTGAGTC | AGATTCGAAGCAACTGCACC |
| hsf-1(NM_060630.7) | ATGTACGGCTTCCGAAAGATGA | TCTTGCCGATTGCTTTCTCTTAA |
| hsp-12.6(NM_069267.4) | TGGAGTTGTCAATGTCCTCG | GACTTCAATCTCTTTTGGGAGG |
| sip-1(NM_066915.5) | CGAGCACGGGTTCAGCAAGAG | CAGCGTGTCCAGCAGAAGTGTG |
| jnk-1(NM_001026099.6) | TGGAACCAGCCAATTCCCAA | TCACAACACTCTGCTCGCAT |
| nsy-1(NM_001383826.1) | AGCGGCTCGATCAACAAGAA | CCCATTCCACCGATATGCGA |
| sek-1(NM_076921.8) | CACTGTTTGGCGACGATGAG | ATTCCGTCCACGTTGCTGAT |
| pmk-1(NM_068964.7) | CCAAAAATGACTCGCCGTGA | CTTTTGCAGTTGGACGACGA |
| Group | Number | Mean Lifespan (h) | Median Survival (Days) | Maximum Lifespan (Days) | p Value |
|---|---|---|---|---|---|
| Control(H2O2) | 109 | 3.541 ± 0.15 | 3 | 10 | _ |
| Gen (H2O2) | 102 | 5.531 ± 0.39 | 5 | 11 | <0.01 |
| Control (35 °C) | 103 | 8.362 ± 0.41 | 9 | 14 | _ |
| Gen (35 °C) | 107 | 14.02 ± 0.39 | 15 | 20 | <0.01 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, S.-Y.; Qin, Z.-C.; Sun, Y.-Y.; Chen, Y.-S.; Chen, W.-B.; Wang, H.-G.; An, D.; Sun, D.; Liu, Y.-Q. Genistein Promotes Anti-Heat Stress and Antioxidant Effects via the Coordinated Regulation of IIS, HSP, MAPK, DR, and Mitochondrial Pathways in Caenorhabditis elegans. Antioxidants 2023, 12, 125. https://doi.org/10.3390/antiox12010125
Zhang S-Y, Qin Z-C, Sun Y-Y, Chen Y-S, Chen W-B, Wang H-G, An D, Sun D, Liu Y-Q. Genistein Promotes Anti-Heat Stress and Antioxidant Effects via the Coordinated Regulation of IIS, HSP, MAPK, DR, and Mitochondrial Pathways in Caenorhabditis elegans. Antioxidants. 2023; 12(1):125. https://doi.org/10.3390/antiox12010125
Chicago/Turabian StyleZhang, Sai-Ya, Zi-Chen Qin, Yi-Yang Sun, Yu-Si Chen, Wen-Bo Chen, Hong-Gang Wang, Di An, Dan Sun, and Yan-Qiang Liu. 2023. "Genistein Promotes Anti-Heat Stress and Antioxidant Effects via the Coordinated Regulation of IIS, HSP, MAPK, DR, and Mitochondrial Pathways in Caenorhabditis elegans" Antioxidants 12, no. 1: 125. https://doi.org/10.3390/antiox12010125
APA StyleZhang, S. -Y., Qin, Z. -C., Sun, Y. -Y., Chen, Y. -S., Chen, W. -B., Wang, H. -G., An, D., Sun, D., & Liu, Y. -Q. (2023). Genistein Promotes Anti-Heat Stress and Antioxidant Effects via the Coordinated Regulation of IIS, HSP, MAPK, DR, and Mitochondrial Pathways in Caenorhabditis elegans. Antioxidants, 12(1), 125. https://doi.org/10.3390/antiox12010125