
Interrelation between miRNAs Expression Associated with Redox State Fluctuations, Immune and Inflammatory Response Activation, and Neonatal Outcomes in Complicated Pregnancy, Accompanied by Placental Insufficiency

 , , ,
, , ,
Abstract
:1. Introduction
2. Materials and Methods
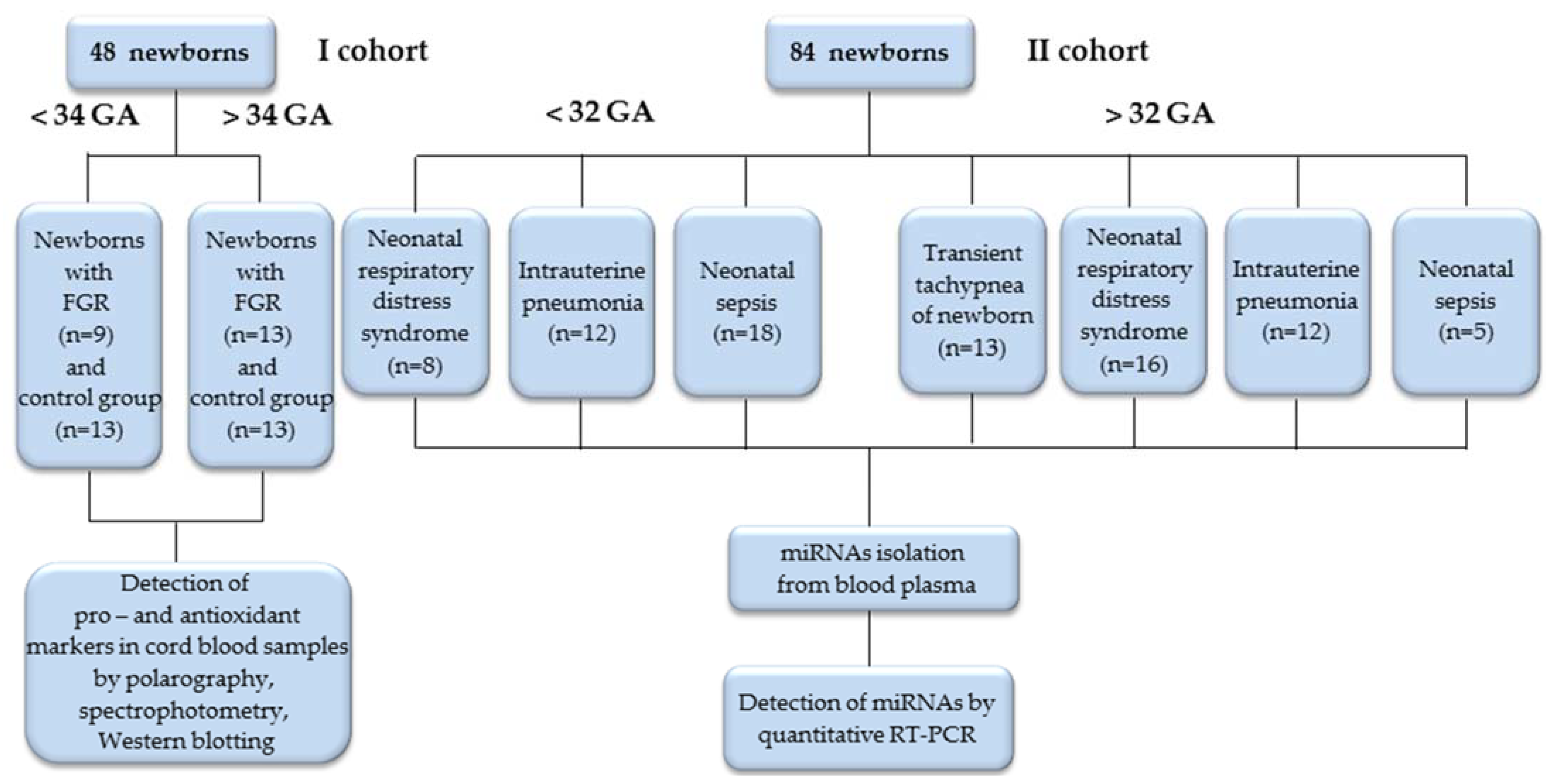
2.1. Study Design and Patient Cohort
2.2. Clinical State Assessment of Newborns with Adverse Neonatal Outcomes
2.3. Detection of Pro- and Anti-Oxidant Markers in Umbilical Cord Blood of Newborns with FGR
2.4. Blood Collection of Newborns, miRNAs Extraction from Blood Plasma and Real-Time Quantitative RT-PCR
2.5. Signaling Pathway Analysis
2.6. Statistical Analysis
3. Results
3.1. Evaluation of Pro- and Anti-Oxidant Markers in Umbilical Cord Blood of Newborns with FGR
3.2. Correlations of Clinical Assessments, Changes in the Expression of Markers of Pro- and Anti-Oxidant Systems in Newborns with FGR and ROC-Curves
3.3. Comparative Evaluation of miRNAs Expression in Newborn Blood Plasma
3.4. Correlation of Clinical Assessments in Newborns with Perinatal Outcomes and ROC-Curves
3.5. Involvement of miR-25-3p and miR-127-3p in Redox Regulation, Immune and Inflammatory Responses
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Ilekis, J.V.; Tsilou, E.; Fisher, S.; Abrahams, V.M.; Soares, M.J.; Cross, J.C.; Zamudio, S.; Illsley, N.P.; Myatt, L.; Colvis, C.; et al. Placental Origins of Adverse Pregnancy Outcomes: Potential Molecular Targets: An Executive Workshop Summary of the Eunice Kennedy Shriver National Institute of Child Health and Human Development. Am. J. Obstet. Gynecol. 2016, 215, S1–S46. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mir, I.N.; Leon, R.; Chalak, L.F. Placental Origins of Neonatal Diseases: Toward a Precision Medicine Approach. Pediatr. Res. 2021, 89, 377–383. [Google Scholar] [CrossRef]
- Brosens, I.; Pijnenborg, R.; Vercruysse, L.; Romero, R. The “Great Obstetrical Syndromes” Are Associated with Disorders of Deep Placentation. Am. J. Obstet. Gynecol. 2011, 204, 193–201. [Google Scholar] [CrossRef] [Green Version]
- Burton, G.J. Oxygen, the Janus Gas; Its Effects on Human Placental Development and Function. J. Anat. 2009, 215, 27–35. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Burton, G.J.; Yung, H.-W.; Cindrova-Davies, T.; Charnock-Jones, D.S. Placental Endoplasmic Reticulum Stress and Oxidative Stress in the Pathophysiology of Unexplained Intrauterine Growth Restriction and Early Onset Preeclampsia. Placenta 2009, 30, 43–48. [Google Scholar] [CrossRef] [Green Version]
- Hung, T.-H.; Burton, G.J. Hypoxia and Reoxygenation: A Possible Mechanism for Placental Oxidative Stress in Preeclampsia. Taiwan. J. Obstet. Gynecol. 2006, 45, 189–200. [Google Scholar] [CrossRef] [Green Version]
- Nardozza, L.M.M.; Caetano, A.C.R.; Zamarian, A.C.P.; Mazzola, J.B.; Silva, C.P.; Marçal, V.M.G.; Lobo, T.F.; Peixoto, A.B.; Araujo Júnior, E. Fetal Growth Restriction: Current Knowledge. Arch. Gynecol. Obs. 2017, 295, 1061–1077. [Google Scholar] [CrossRef] [PubMed]
- American College of Obstetricians and Gynecologists. ACOG Practice Bulletin No. 204: Fetal Growth Restriction. Obstet. Gynecol. 2019, 133, e97–e109. [Google Scholar] [CrossRef]
- Takagi, Y.; Nikaido, T.; Toki, T.; Kita, N.; Kanai, M.; Ashida, T.; Ohira, S.; Konishi, I. Levels of Oxidative Stress and Redox-Related Molecules in the Placenta in Preeclampsia and Fetal Growth Restriction. Virchows Arch. 2004, 444, 49–55. [Google Scholar] [CrossRef]
- Li, Y.R.; Trush, M. Defining ROS in Biology and Medicine. ROS 2016, 1, 9–21. [Google Scholar] [CrossRef]
- Herb, M.; Schramm, M. Functions of ROS in Macrophages and Antimicrobial Immunity. Antioxidants 2021, 10, 313. [Google Scholar] [CrossRef] [PubMed]
- Chatterjee, S. Oxidative Stress, Inflammation, and Disease. In Oxidative Stress and Biomaterials; Elsevier: Amsterdam, The Netherlands, 2016; pp. 35–58. ISBN 978-0-12-803269-5. [Google Scholar]
- Rudov, A.; Balduini, W.; Carloni, S.; Perrone, S.; Buonocore, G.; Albertini, M.C. Involvement of MiRNAs in Placental Alterations Mediated by Oxidative Stress. Oxidative Med. Cell. Longev. 2014, 2014, 103068. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Meng, W.; Hao, Y.; He, C.; Li, L.; Zhu, G. Exosome-Orchestrated Hypoxic Tumor Microenvironment. Mol. Cancer 2019, 18, 57. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Villarroya-Beltri, C.; Gutiérrez-Vázquez, C.; Sánchez-Cabo, F.; Pérez-Hernández, D.; Vázquez, J.; Martin-Cofreces, N.; Martinez-Herrera, D.J.; Pascual-Montano, A.; Mittelbrunn, M.; Sánchez-Madrid, F. Sumoylated HnRNPA2B1 Controls the Sorting of MiRNAs into Exosomes through Binding to Specific Motifs. Nat. Commun. 2013, 4, 2980. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ruiz, G.P.; Camara, H.; Fazolini, N.P.B.; Mori, M.A. Extracellular MiRNAs in Redox Signaling: Health, Disease and Potential Therapies. Free Radic. Biol. Med. 2021, 173, 170–187. [Google Scholar] [CrossRef] [PubMed]
- Tannetta, D.S.; Dragovic, R.A.; Gardiner, C.; Redman, C.W.; Sargent, I.L. Characterisation of Syncytiotrophoblast Vesicles in Normal Pregnancy and Pre-Eclampsia: Expression of Flt-1 and Endoglin. PLoS ONE 2013, 8, e56754. [Google Scholar] [CrossRef]
- Kura, B.; Szeiffova Bacova, B.; Kalocayova, B.; Sykora, M.; Slezak, J. Oxidative Stress-Responsive MicroRNAs in Heart Injury. Int. J. Mol. Sci. 2020, 21, 358. [Google Scholar] [CrossRef] [Green Version]
- Banerjee, J.; Khanna, S.; Bhattacharya, A. MicroRNA Regulation of Oxidative Stress. Oxidative Med. Cell. Longev. 2017, 2017, 2872156. [Google Scholar] [CrossRef] [Green Version]
- Carbonell, T.; Gomes, A.V. MicroRNAs in the Regulation of Cellular Redox Status and Its Implications in Myocardial Ischemia-Reperfusion Injury. Redox Biol. 2020, 36, 101607. [Google Scholar] [CrossRef]
- Leisegang, M.S.; Schröder, K.; Brandes, R.P. Redox Regulation and Noncoding RNAs. Antioxid. Redox Signal. 2018, 29, 793–812. [Google Scholar] [CrossRef]
- Essandoh, K.; Li, Y.; Huo, J.; Fan, G.-C. MiRNA-Mediated Macrophage Polarization and Its Potential Role in the Regulation of Inflammatory Response. Shock 2016, 46, 122–131. [Google Scholar] [CrossRef] [PubMed]
- Ozsurekci, Y.; Aykac, K. Oxidative Stress Related Diseases in Newborns. Oxidative Med. Cell. Longev. 2016, 2016, 2768365. [Google Scholar] [CrossRef] [Green Version]
- Cannavò, L.; Perrone, S.; Viola, V.; Marseglia, L.; Di Rosa, G.; Gitto, E. Oxidative Stress and Respiratory Diseases in Preterm Newborns. Int. J. Mol. Sci. 2021, 22, 12504. [Google Scholar] [CrossRef] [PubMed]
- Marseglia, L.; D’Angelo, G.; Granese, R.; Falsaperla, R.; Reiter, R.J.; Corsello, G.; Gitto, E. Role of Oxidative Stress in Neonatal Respiratory Distress Syndrome. Free Radic. Biol. Med. 2019, 142, 132–137. [Google Scholar] [CrossRef] [PubMed]
- Abdel Hamid, E.R.; Ali, W.H.; Azmy, A.; Ahmed, H.H.; Sherif, L.S.; Saleh, M.T. Oxidative Stress and Anti-Oxidant Markers in Premature Infants with Respiratory Distress Syndrome. Open Access Maced. J. Med. Sci. 2019, 7, 2858–2863. [Google Scholar] [CrossRef] [Green Version]
- De Almeida, V.O.; Pereira, R.A.; Amantéa, S.L.; Rhoden, C.R.; Colvero, M.O. Neonatal Diseases and Oxidative Stress in Premature Infants: An Integrative Review. J. Pediatr. 2022, 98, 455–462. [Google Scholar] [CrossRef]
- Poggi, C.; Dani, C. Sepsis and Oxidative Stress in the Newborn: From Pathogenesis to Novel Therapeutic Targets. Oxidative Med. Cell. Longev. 2018, 2018, 9390140. [Google Scholar] [CrossRef] [Green Version]
- Dani, C.; Cecchi, A.; Bertini, G. Role of Oxidative Stress as Physiopathologic Factor in the Preterm Infant. Minerva Pediatr. 2004, 56, 381–394. [Google Scholar]
- Bharadwaj, S.; Bhat, V.B.; Vickneswaran, V.; Adhisivam, B.; Zachariah, B.; Habeebullah, S. Oxidative Stress in Preeclamptic Mother—Newborn Dyads and Its Correlation with Early Neonatal Outcome—A Case Control Study. J. Matern. Fetal Neonatal. Med. 2018, 31, 1548–1553. [Google Scholar] [CrossRef]
- Barron, A.; McCarthy, C.M.; O’Keeffe, G.W. Preeclampsia and Neurodevelopmental Outcomes: Potential Pathogenic Roles for Inflammation and Oxidative Stress? Mol. Neurobiol. 2021, 58, 2734–2756. [Google Scholar] [CrossRef]
- Pavlek, L.R.; Vudatala, S.; Bartlett, C.W.; Buhimschi, I.A.; Buhimschi, C.S.; Rogers, L.K. MiR-29b Is Associated with Perinatal Inflammation in Extremely Preterm Infants. Pediatr. Res. 2021, 89, 889–893. [Google Scholar] [CrossRef] [PubMed]
- Gusar, V.; Ganichkina, M.; Chagovets, V.; Kan, N.; Sukhikh, G. MiRNAs Regulating Oxidative Stress: A Correlation with Doppler Sonography of Uteroplacental Complex and Clinical State Assessments of Newborns in Fetal Growth Restriction. J. Clin. Med. 2020, 9, 3227. [Google Scholar] [CrossRef] [PubMed]
- Gordijn, S.J.; Beune, I.M.; Thilaganathan, B.; Papageorghiou, A.; Baschat, A.A.; Baker, P.N.; Silver, R.M.; Wynia, K.; Ganzevoort, W. Consensus Definition of Fetal Growth Restriction: A Delphi Procedure: Consensus Definition of FGR. Ultrasound Obstet. Gynecol. 2016, 48, 333–339. [Google Scholar] [CrossRef] [PubMed]
- Silverman, W.A.; Andersen, D.H. A Controlled Clinical Trial of Effects of Water Mist on Obstructive Respiratory Signs, Death Rate and Necropsy Findings among Premature Infants. Pediatrics 1956, 17, 1–10. [Google Scholar]
- Downes, J.J.; Vidyasagar, D.; Morrow, G.M.; Boggs, T.R. Respiratory Distress Syndrome of Newborn Infants: I. New Clinical Scoring System (RDS Score) with Acid-Base and Blood- Gas Correlations. Clin. Pediatr. 1970, 9, 325–331. [Google Scholar] [CrossRef]
- Janota, J.; Simak, J.; Stranak, Z.; Matthews, T.; Clarke, T.; Corcoran, D. Critically Ill Newborns with Multiple Organ Dysfunction: Assessment by NEOMOD Score in a Tertiary NICU. Ir. J. Med. Sci. 2008, 177, 11–17. [Google Scholar] [CrossRef]
- Weiss, S.L.; Peters, M.J.; Alhazzani, W.; Agus, M.S.D.; Flori, H.R.; Inwald, D.P.; Nadel, S.; Schlapbach, L.J.; Tasker, R.C.; Argent, A.C.; et al. Surviving Sepsis Campaign International Guidelines for the Management of Septic Shock and Sepsis-Associated Organ Dysfunction in Children. Pediatr. Crit. Care Med. 2020, 21, e52–e106. [Google Scholar] [CrossRef]
- Schimke, I.; Kahl, P.-E.; Romaniuk, P.; Papies, B. Konzentration Thiobarbitursäure-reaktiver Substanzen (TBARS) im Serum nach Myokardinfarkt. Klin. Wochenschr. 1986, 64, 1237–1239. [Google Scholar] [CrossRef]
- Tietze, F. Enzymic Method for Quantitative Determination of Nanogram Amounts of Total and Oxidized Glutathione: Applications to Mammalian Blood and Other Tissues. Anal. Biochem. 1969, 27, 502–522. [Google Scholar] [CrossRef]
- Esterbauer, H.; Cheeseman, K.H. Determination of Aldehydic Lipid Peroxidation Products: Malonaldehyde and 4-Hydroxynonenal. In Methods in Enzymology; Elsevier: Amsterdam, The Netherlands, 1990; Volume 186, pp. 407–421. ISBN 978-0-12-182087-9. [Google Scholar]
- Burri, B.J.; Chan, S.G.; Berry, A.J.; Yarnell, S.K. Blood Levels of Superoxide Dismutase and Glutathione Peroxidase in Duchenne Muscular Dystrophy. Clin. Chim. Acta 1980, 105, 249–255. [Google Scholar] [CrossRef]
- Paglia, D.E.; Valentine, W.N. Studies on the Quantitative and Qualitative Characterization of Erythrocyte Glutathione Peroxidase. J. Lab. Clin. Med. 1967, 70, 158–169. [Google Scholar] [PubMed]
- Rørth, M.; Jensen, P.K. Determination of Catalase Activity by Means of the Clark Oxygen Electrode. Biochim. Biophys. Acta Enzymol. 1967, 139, 171–173. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of Relative Gene Expression Data Using Real-Time Quantitative PCR and the 2−ΔΔCT Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Burton, G.J.; Jauniaux, E. Oxidative Stress. Best Pract. Res. Clin. Obstet. Gynaecol. 2011, 25, 287–299. [Google Scholar] [CrossRef] [Green Version]
- Schoots, M.H.; Gordijn, S.J.; Scherjon, S.A.; van Goor, H.; Hillebrands, J.-L. Oxidative Stress in Placental Pathology. Placenta 2018, 69, 153–161. [Google Scholar] [CrossRef] [PubMed]
- Sun, C.; Groom, K.M.; Oyston, C.; Chamley, L.W.; Clark, A.R.; James, J.L. The Placenta in Fetal Growth Restriction: What Is Going Wrong? Placenta 2020, 96, 10–18. [Google Scholar] [CrossRef] [PubMed]
- Zelko, I.N.; Mariani, T.J.; Folz, R.J. Superoxide Dismutase Multigene Family: A Comparison of the CuZn-SOD (SOD1), Mn-SOD (SOD2), and EC-SOD (SOD3) Gene Structures, Evolution, and Expression. Free Radic. Biol. Med. 2002, 33, 337–349. [Google Scholar] [CrossRef]
- Chiarello, D.I.; Abad, C.; Rojas, D.; Toledo, F.; Vázquez, C.M.; Mate, A.; Sobrevia, L.; Marín, R. Oxidative Stress: Normal Pregnancy versus Preeclampsia. Biochim. Et Biophys. Acta Mol. Basis Dis. 2020, 1866, 165354. [Google Scholar] [CrossRef]
- Liu, L.; Zheng, B.; Jiang, Z.; Wu, S.; Jin, Q.; Lin, P.; Lin, Z.; Wang, L. Association of Elevated Cord Blood Oxidative Stress Biomarkers with Neonatal Outcomes in Mothers with Pre-Eclampsia: A Case-Control Study. Gynecol. Obstet. Investig. 2021, 86, 361–369. [Google Scholar] [CrossRef]
- Asiltas, B.; Surmen-Gur, E.; Uncu, G. Prediction of First-Trimester Preeclampsia: Relevance of the Oxidative Stress Marker MDA in a Combination Model with PP-13, PAPP-A and Beta-HCG. Pathophysiology 2018, 25, 131–135. [Google Scholar] [CrossRef]
- Biberoglu, E.; Biberoglu, K.; Kirbas, A.; Daglar, K.; Genc, M.; Avci, A.; Danisman, N. Circulating and Myometrial Markers of Oxidative Stress in Pregnant Women with Fetal Growth Restriction: Oxidative Stress in FGR. J. Obstet. Gynaecol. Res. 2016, 42, 29–35. [Google Scholar] [CrossRef] [PubMed]
- Buonocore, G.; Perrone, S.; Tataranno, M.L. Oxidative Stress in the Newborn. Oxidative Med. Cell. Longev. 2017, 2017, 1094247. [Google Scholar] [CrossRef] [PubMed]
- Adeoye, O.; Olawumi, J.; Opeyemi, A.; Christiania, O. Review on the Role of Glutathione on Oxidative Stress and Infertility. JBRA Assist. Reprod. 2017, 22, 61–66. [Google Scholar] [CrossRef] [PubMed]
- Ahmad, I.M.; Zimmerman, M.C.; Moore, T.A. Oxidative Stress in Early Pregnancy and the Risk of Preeclampsia. Pregnancy Hypertens. 2019, 18, 99–102. [Google Scholar] [CrossRef]
- Panfoli, I.; Candiano, G.; Malova, M.; De Angelis, L.; Cardiello, V.; Buonocore, G.; Ramenghi, L.A. Oxidative Stress as a Primary Risk Factor for Brain Damage in Preterm Newborns. Front. Pediatr. 2018, 6, 369. [Google Scholar] [CrossRef] [Green Version]
- Cascant-Vilaplana, M.M.; Sánchez-Illana, Á.; Piñeiro-Ramos, J.D.; Llorens-Salvador, R.; Quintás, G.; Oger, C.; Galano, J.-M.; Vigor, C.; Durand, T.; Kuligowski, J.; et al. Do Levels of Lipid Peroxidation Biomarkers Reflect the Degree of Brain Injury in Newborns? Antioxid. Redox Signal. 2021, 35, 1467–1475. [Google Scholar] [CrossRef]
- Jugović, D.; Tumbri, J.; Medić, M.; Jukić, M.K.; Kurjak, A.; Arbeille, P.; Salihagić-Kadić, A. New Doppler Index for Prediction of Perinatal Brain Damage in Growth-Restricted and Hypoxic Fetuses. Ultrasound Obstet. Gynecol. 2007, 30, 303–311. [Google Scholar] [CrossRef]
- Tanis, J.C.; Boelen, M.R.; Schmitz, D.M.; Casarella, L.; van der Laan, M.E.; Bos, A.F.; Bilardo, C.M. Correlation between Doppler Flow Patterns in Growth-Restricted Fetuses and Neonatal Circulation: Doppler and NIRS in FGR. Ultrasound Obstet. Gynecol. 2016, 48, 210–216. [Google Scholar] [CrossRef] [Green Version]
- Lee, S.; Lim, S.; Ham, O.; Lee, S.-Y.; Lee, C.Y.; Park, J.-H.; Lee, J.; Seo, H.-H.; Yun, I.; Han, S.M.; et al. ROS-Mediated Bidirectional Regulation of MiRNA Results in Distinct Pathologic Heart Conditions. Biochem. Biophys. Res. Commun. 2015, 465, 349–355. [Google Scholar] [CrossRef]
- Zhu, C.; Chen, T.; Liu, B. Inhibitory Effects of MiR-25 Targeting HMGB1 on Macrophage Secretion of Inflammatory Cytokines in Sepsis. Oncol. Lett. 2018, 16, 5027–5033. [Google Scholar] [CrossRef] [Green Version]
- Huang, F.; Zhang, J.; Yang, D.; Zhang, Y.; Huang, J.; Yuan, Y.; Li, X.; Lu, G. MicroRNA Expression Profile of Whole Blood Is Altered in Adenovirus-Infected Pneumonia Children. Mediat. Inflamm. 2018, 2018, 2320640. [Google Scholar] [CrossRef] [PubMed]
- Ying, H.; Kang, Y.; Zhang, H.; Zhao, D.; Xia, J.; Lu, Z.; Wang, H.; Xu, F.; Shi, L. MiR-127 Modulates Macrophage Polarization and Promotes Lung Inflammation and Injury by Activating the JNK Pathway. J. Immunol. 2015, 194, 1239–1251. [Google Scholar] [CrossRef] [PubMed]
- Zheng, G.; Xiang, W.; Pan, M.; Huang, Y.; Li, Z. Identification of the Association between Rs41274221 Polymorphism in the Seed Sequence of MicroRNA-25 and the Risk of Neonate Sepsis. J. Cell. Physiol. 2019, 234, 15147–15155. [Google Scholar] [CrossRef] [PubMed]
- Yao, L.; Liu, Z.; Zhu, J.; Li, B.; Chai, C.; Tian, Y. Clinical Evaluation of Circulating MicroRNA-25 Level Change in Sepsis and Its Potential Relationship with Oxidative Stress. Int. J. Clin. Exp. Pathol. 2015, 8, 7675–7684. [Google Scholar]
- Varga, Z.V.; Kupai, K.; Szűcs, G.; Gáspár, R.; Pálóczi, J.; Faragó, N.; Zvara, Á.; Puskás, L.G.; Rázga, Z.; Tiszlavicz, L.; et al. MicroRNA-25-Dependent up-Regulation of NADPH Oxidase 4 (NOX4) Mediates Hypercholesterolemia-Induced Oxidative/Nitrative Stress and Subsequent Dysfunction in the Heart. J. Mol. Cell. Cardiol. 2013, 62, 111–121. [Google Scholar] [CrossRef]
- Xie, T.; Liang, J.; Liu, N.; Wang, Q.; Li, Y.; Noble, P.W.; Jiang, D. MicroRNA-127 Inhibits Lung Inflammation by Targeting IgG Fcγ Receptor I. J. Immunol. 2012, 188, 2437–2444. [Google Scholar] [CrossRef] [Green Version]
- Özmeral Odabaşı, I. Neonatal Sepsis. Sisli Etfal 2020, 54, 142–158. [Google Scholar] [CrossRef]
- Dalili, H.; Sheikh, M.; Hardani, A.K.; Nili, F.; Shariat, M.; Nayeri, F. Comparison of the Combined versus Conventional Apgar Scores in Predicting Adverse Neonatal Outcomes. PLoS ONE 2016, 11, e0149464. [Google Scholar] [CrossRef]
- El Damaty, A.; Giannoni, L.; Unterberg, A.; Baechli, H. Thrombocytopenia: Is It a Prognostic Factor for Development of Post-Hemorrhagic Hydrocephalus in Neonates? Childs Nerv. Syst. 2021, 37, 519–527. [Google Scholar] [CrossRef]
- Rosales, C. Neutrophils at the Crossroads of Innate and Adaptive Immunity. J. Leukoc. Biol. 2020, 108, 377–396. [Google Scholar] [CrossRef]
- Pérez-Figueroa, E.; Álvarez-Carrasco, P.; Ortega, E.; Maldonado-Bernal, C. Neutrophils: Many Ways to Die. Front. Immunol. 2021, 12, 631821. [Google Scholar] [CrossRef] [PubMed]
- Nupponen, I.; Pesonen, E.; Andersson, S.; Mäkelä, A.; Turunen, R.; Kautiainen, H.; Repo, H. Neutrophil Activation in Preterm Infants Who Have Respiratory Distress Syndrome. Pediatrics 2002, 110, 36–41. [Google Scholar] [CrossRef] [PubMed]
- Guo, H.; Wang, M.; Wang, B.; Guo, L.; Cheng, Y.; Wang, Z.; Sun, Y.-Q.; Wang, Y.; Chang, Y.-J.; Huang, X.-J. PRDM1 Drives Human Primary T Cell Hyporesponsiveness by Altering the T Cell Transcriptome and Epigenome. Front. Immunol. 2022, 13, 879501. [Google Scholar] [CrossRef]
- Guo, Y.; Niu, S. MiR-25 Protects PC-12 Cells from H2O2 Mediated Oxidative Damage via WNT/β-Catenin Pathway. J. Spinal Cord Med. 2018, 41, 416–425. [Google Scholar] [CrossRef]
- Benz, F.; Roy, S.; Trautwein, C.; Roderburg, C.; Luedde, T. Circulating MicroRNAs as Biomarkers for Sepsis. Int. J. Mol. Sci. 2016, 17, 78. [Google Scholar] [CrossRef] [Green Version]
- Lin, H.; Jiang, L.; Ren, Y.; Sheng, F.; Wang, L.; Zhang, S. Expression Level, Correlation, and Diagnostic Value of Serum MiR-127 in Patients with Acute Respiratory Distress Syndrome. Evid. Based Complement. Altern. Med. 2021, 2021, 2257764. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| <32 GA | >32 GA | ||||||
|---|---|---|---|---|---|---|---|
| RDS (n = 8) | IP (n = 12) | NS (n = 18) | TT (n = 13) | RDS (n = 16) | IP (n = 12) | NS (n = 5) | |
| Gestational age at the time of delivery, weeks | 30.6 ± 1.12 * | 28.1 ± 1.9 | 28.06 ± 2.34 ‡ | 35.69 ± 2.56 ** | 33.73 ± 1.62 | 35.08 ± 3.03 | 37.2 ± 2.68 ‡ |
| BW, (grams) | 1531.7 ± 266.9 * | 1077.3 ± 253.4 | 942.3 ± 302.5 ‡ | 2435.8 ± 357.5 | 1848.0 ± 308.3 ‡ | 2218.8 ± 705.1 | 2586.4 ± 680.6 |
| Gender, n (%) | |||||||
| Male: | 3 (37.5) | 10 (83.3) | 11 (61.2) | 6 (46.1) | 8 (50) | 8 (66.6) | 4 (80) |
| Female: | 5 (62.5) | 2 (16.7) | 7 (38.8) | 7 (53.9) | 8 (50) | 4 (33.4) | 1 (20) |
| APGAR 1 min | 6.67 ± 0.5 * | 5.17 ± 0.75 | 4.72 ± 1.23 ‡ | 6.77 ± 1.48 | 6.93 ± 0.46 | 5.75 ± 2.63 | 7.4 ± 0.89 |
| APGAR 5 min | 7.44 ± 0.53 * | 6.83 ± 0.41 | 6.61 ± 0.85 ‡ | 7.77 ± 1.17 | 7.87 ± 0.35 | 6.92 ± 2.11 | 8.2 ± 0.84 |
| NEOMOD | 2.89 ± 0.6 * | 5.17 ± 1.47 | 5.17 ± 1.54 ‡ | 1.85 ± 0.55 & | 2.73 ± 0.7 ** | 3.58 ± 1.31 | 4 ± 1.41 ‡,Δ |
| Silverman | 2.56 ± 0.73 * | 3.5 ± 0.84 | 3.22 ± 0.55 ‡ | 1.62 ± 0.96 ** | 2.67 ± 0.9 | 1.92 ± 1.56 | 1.6 ± 1.52 |
| Downes | --- | --- | --- | 0.46 ± 0.97 | --- | 1.08 ± 1.62 | 1.2 ± 1.64 ‡ |
| IVH, n (%) Grade (1–2) Grade (3) | 1 (12.5) 1 --- | 12 (100) 10 2 | 10 (55.5) 6 4 | --- | --- | 2 (16.6) 2 --- | --- |
| WBC (6.0–17.5 × 109 c/L) (absolute count) | 8.75 ± 2.83 | 10.46 ± 4.45 | 7.76 ± 3.54 | 17.06 ± 5.46 ** | 12.38 ± 6.54 | 16.25 ± 8.75 | 10.7 ± 6.28 |
| NEUT (1.5–8.5 × 109 c/L) (absolute level) neutropenia (<1500 c/mcl), n (%) | 4055 ± 1377 --- | 2932 ± 1955 1 (8.3) | 2435 ± 2591 ‡ 3 (16.6) | 8727 ± 4468 ** --- | 5916 ± 5120 2 (12.5) | 6957 ± 6132 --- | 5378 ± 4272 1 (20) |
| NI (0.17–0.21) | 0.06 ± 0.07 | 0.06 ± 0.06 | 0.13 ± 0.12 | 0.05 ± 0.04 | 0.04 ± 0.05 | 0.06 ± 0.07 | 0.05 ± 0.03 |
| Platelet (180–400 × 109 c/L) thrombocytopenia (less 150 × 109 c/L), n (%) | 262.4 ± 43.1 * | 164.3 ± 61.2 | 179.7 ± 63.4 ‡ | 291.2 ± 60.9 &,Δ | 252.1 ± 61.0 | 223.3 ± 66.8 | 195.8 ± 32.7 ‡ |
| CRP (less 1.6 mg/l) | 1.57 ± 1.45 | 3.97 ± 7.25 | 1.9 ± 1.78 | 1.1 ± 0.75 | 0.87 ± 0.9 | 1.51 ± 2.19 | 5.57 ± 9.1 |
| Maternal diagnoses, n: | |||||||
| PE | 1 | 1 | 2 | --- | --- | 2 | --- |
| FGR | 1 | 4 | 4 | --- | --- | --- | --- |
| PE with FGR | --- | --- | 4 | --- | --- | --- | --- |
| GDM | --- | 1 | --- | --- | --- | --- | --- |
| Thrombophilia | --- | 1 | 5 | --- | --- | --- | --- |
| GH | --- | 3 | --- | --- | --- | --- | --- |
| AFH | --- | --- | 1 | --- | --- | 1 | --- |
| Parameter | r * | p ** | |
|---|---|---|---|
| Weight of newborn | GPx | 0.6 | 0.03 |
| SOD1 | −0.6 | 0.01 | |
| SOD2 | −0.6 | 0.02 | |
| Cat | −0.8 | 0.0005 | |
| Apgar 1 | MDA | −0.5 | 0.03 |
| GPx | 0.5 | 0.03 | |
| GSH/GSSG | 0.6 | 0.01 | |
| Cat | −0.6 | 0.03 | |
| Apgar 5 | GPx | 0.5 | 0.05 |
| GSH/GSSG | 0.7 | 0.007 | |
| SOD2 | −0.7 | 0.003 |
| Absence of Complication | Presence of Complication | p * | ||
|---|---|---|---|---|
| <34 GA | ||||
| ELBW | ||||
| MDA | 11.4 (10.3; 13.1) | 16.7 (16; 17.8) | 0.04 | |
| CAT | 83.8 (73.95; 88.2) | 36.1 (35; 40.4) | 0.009 | |
| tGSH | 343.2 (247.5; 376.4) | 4.6 (4.35; 4.75) | 0.01 | |
| GSH | 3.27 (3.18; 3.32) | 2.6 (2.25; 2.7) | 0.04 | |
| GSH/GSSG | 2.89 (2.8; 3.31) | 1.13 (0.92; 1.64) | 0.02 | |
| SOD1 | 34,576.9 (32,347.1; 35,896.6) | 44,368.92 (43,782.74; 46,867.36) | 0.01 | |
| SOD2 | 42,686.2 (41,245.2; 44,501.7) | 69,632.2 (65,415.3; 71,929.8) | 0.003 | |
| Cat | 52,101.5 (50,577.6; 53,371.4) | 78,296.4 (73,720.08; 80,756.8) | 0.004 | |
| Hyperechogenicity periventricular zone (by neurosonography) | ||||
| 4-HNE | 7.15 (6.08; 8.12) | 11 (10.85; 11.15) | 0.03 | |
| Cat | 53,371.4 (51,339.5; 58,266.9) | 80,756.8 (79,526.6; 81,987.1) | 0.02 | |
| >34 GA | ||||
| Intrauterine pneumonia | ||||
| 4-HNE | 6.7 (6; 7.9) | 5.1 (4.15; 5.5) | 0.04 | |
| Hyperbilirubinemia | ||||
| 4-HNE | 6.55 (5.93; 7.78) | 4.15 (3.68; 4.62) | 0.02 | |
| IVH | ||||
| SOD1 | 29,063.8 (9328.7; 32,210.6) | 7648.2 (7373.4; 7978.8) | 0.04 | |
| SOD2 | 36,105.7 (9065.3; 39,472.5) | 9903.1 (9113.9; 9943.6) | 0.05 | |
| Cerebral cyst (by neurosonography) | ||||
| CAT | 79.2 (69.2; 86.3) | 93.1 (92.9; 95.1) | 0.04 | |
| SOD1 | 29,063.8 (9328.7; 32,210.6) | 7098.6 (6973.08; 7373.4) | 0.03 | |
| SOD2 | 36,105.7 (9065.3; 39,472.5) | 9903.1 (9113.9; 9993.8) | 0.05 | |
| Respiratory Distress Syndrome | Intrauterine Pneumonia | Neonatal Sepsis | ||||||
|---|---|---|---|---|---|---|---|---|
| r * | p ** | r * | p ** | r * | p ** | |||
| APGAR 5—BW | 0.7 | 0.04 | APGAR 1—NI | −0.6 | 0.01 | miR-127-3p—APGAR 5 | −0.9 | 0.01 |
| APGAR 5—NI | −0.6 | 0.006 | APGAR 5—NI | −0.6 | 0.04 | NEOMOD—BW | −0.9 | 0.04 |
| Silverman—NEU | 0.5 | 0.04 | APGAR 1—Silverman | −0.8 | 0.04 | |||
| Silverman—LEU | 0.5 | 0.04 | APGAR 5—Silverman | −0.9 | 0.02 | |||
| Platelet—NEOMOD | −0.8 | 0.04 | ||||||
| <32 GA | ||||||||
| Thrombocytopenia—IVH | 0.6 | 0.008 | ||||||
| IVH—NEOMOD | 0.6 | 0.007 | ||||||
| miR-25-3p + miR-127-3p | AUC | Sensitivity | Specificity | Cutoff |
|---|---|---|---|---|
| RDS vs. IP < 32 GA | 0.75 | 0.83 | 0.75 | 0.74 |
| RDS vs. IP > 32 GA | 0.86 | 0.83 | 0.88 | 0.46 |
| NS vs. IP < 32 GA | 0.75 | 0.75 | 0.78 | 0.45 |
| NS vs. IP > 32 GA | 0.72 | 0.75 | 0.8 | 0.67 |
| IP (IVH) | 0.84 | 0.79 | 0.9 | 0.67 |
| miR-127-3p | ||||
| NS (IVH) | 0.68 | 0.64 | 0.83 | 0.49 |
| Gene Abbreviation (Ensembl ID) | Gene Name | Description | miRNA * (Reporter Assay, Western Blot, qPCR) | Pathway |
|---|---|---|---|---|
| BCL2L11 (ENSG00000153094) | Bcl-2-like protein 11 (BIM) | Belongs to the BCL-2 protein family. The protein encoded by this gene interacts with other members of the BCL-2 protein family and acts as an apoptosis activator. Its expression can be induced by the nerve growth factor (NGF) as well as the transcription factor FKHR-L1, which suggests the role of this gene in the apoptosis of neurons and lymphocytes. | miR-25-3p miR-9-5p miR-106b-5p miR-17-5p miR-32-5p miR-10b-5p miR-181a-5p miR-494-3p miR-375 miR-92a-3p miR-24-3p | FOXO-mediated transcription, Apoptosis and autophagy, MAPK signaling: oxidative stress, Cytokine signaling in immune system, PI3K-Akt signaling pathway. |
| CCL26 (ENSG00000006606) | C-C motif chemokine ligand 26 | Belongs to the family of cytokine genes involved in immunoregulatory and inflammatory processes. | miR-25-3p | MIF regulation of innate immune cells, CCR5 pathway in macrophages, Immune response CCR3 Pathway in eosinophils. |
| NFE2L2 (NRF2) (ENSG00000116044) | Nuclear Factor, Erythroid 2 Like 2 | A transcription factor that plays a key role in the response to oxidative stress. By binding to antioxidant response elements (ARE) present in the promoter region of many cytoprotective genes, it promotes their expression, thereby neutralizing reactive electrophiles. Critical regulator of innate immune response and survival in sepsis by maintaining redox homeostasis and curbing dysregulation of pro-inflammatory signaling pathways. Suppresses the inflammatory response of macrophages by blocking the transcription of pro-inflammatory cytokines and the induction of IL-6. | miR-340-5p miR-17-5p miR-93-5p miR-20a-5p miR-106b-5p | KEAP1-NFE2L2 pathway, Regulation of HMOX1 expression and activity, MAPK signaling: oxidative stress, Class I MHC mediated antigen processing and presentation, Oxidative stress response. |
| PRDM1 (ENSG00000057657) | B lymphocyte-induced maturation protein-1 | A transcription factor that mediates the innate and adaptive immune response of tissue-resident T-lymphocytes (resident memory T-cells, resident natural killers (TrNK) and natural killers (NKT) in non-lymphoid organs (skin, intestines), as well as the liver and kidneys, which provides immediate immunological protection against reactivating infections or viral reinfection. Controls the maturation of B-lymphocytes in Ig-secreting cells. Acts as a repressor of interferon-beta expression. | miR-127-3p miR-9-5p miR-30a-5p miR-125b-5p miR-222-3p let-7a-5p let-7b-5p let-7f-5p | NF-kappaB signaling, Leukocyte-intrinsic Hippo pathway functions, Vitamin D receptor pathway. |
| BCL6 (ENSG00000113916) | B-cell lymphoma 6 | Acts as a transcriptional repressor and modulates the transcription of STAT-dependent IL-4 responses in B cells. Necessary for the establishment and maintenance of immunological memory of T and B cells. Suppresses the proliferation of macrophages. It also controls neurogenesis by changing the composition of NOTCH-dependent transcription complexes on selective NOTCH targets, which leads to neuronal differentiation. | miR-127-3p miR-9-5p | Cytokine signaling in Immune system, FOXO-mediated transcription. NF-kappaB signaling, Notch signaling pathways, IL-4 and IL-13 signaling, BCR signaling pathway, IL4-mediated signaling events, Vitamin D receptor pathway. |
| HIF1a (ENSG00000100644) | Hypoxia Inducible Factor 1 Subunit Alpha | A transcription factor that functions as the main regulator of the adaptive cellular response to hypoxia. Activates transcription of more than 40 genes under hypoxic conditions involved in energy metabolism, angiogenesis, apoptosis, protein products which increase oxygen delivery or facilitate metabolic adaptation to hypoxia. Induces the expression of ACE2 and cytokines such as IL1B, TNF, IL6 and interferons in monocytes. | miR-20a-5p miR-18a-5p miR-17-5p let-7b-5p miR-93-5p miR-106b-5p miR-494-3p | Cellular response to hypoxia, Hypoxic and oxygen homeostasis regulation of HIF-1-alpha, Cytokine signaling in Immune system, Class I MHC mediated antigen processing and presentation, IL-4 and IL-13 signaling, Angiogenesis, Notch and PI3K-Akt signaling pathways, Vitamin D receptor pathway. |
| NFKB1 (ENSG00000109320) | Nuclear Factor Kappa B Subunit 1 | A transcriptional regulator that is activated by various intracellular and extracellular stimuli such as cytokines, free radicals, ultraviolet radiation, bacterial or viral products. Inadequate activation of NFKB is associated with a number of inflammatory diseases, while persistent inhibition of NFKB leads to abnormal development of immune cells or retardation cell growth. NFKB is a critical regulator of the immediate early response to viral infection. | miR-9-5p miR-92a-3p miR-195-5p let-7a-5p | Cytokine signaling in Immune system, Immune response IL-1, IL-6, IL-9, IL-17 and IL-23 signaling pathway, Immune response TLR signaling pathways, NFAT in immune response, CCR5 pathway in macrophages, Notch, MAPK and PI3K-Akt signaling pathways, Class I MHC mediated antigen processing and presentation, miRNAs involvement in the immune response in sepsis, Neuroinflammation and glutamatergic signaling, Oxidative damage and stress response. |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gusar, V.A.; Timofeeva, A.V.; Chagovets, V.V.; Vysokikh, M.Y.; Kan, N.E.; Manukhova, L.A.; Marey, M.V.; Sukhikh, G.T. Interrelation between miRNAs Expression Associated with Redox State Fluctuations, Immune and Inflammatory Response Activation, and Neonatal Outcomes in Complicated Pregnancy, Accompanied by Placental Insufficiency. Antioxidants 2023, 12, 6. https://doi.org/10.3390/antiox12010006
Gusar VA, Timofeeva AV, Chagovets VV, Vysokikh MY, Kan NE, Manukhova LA, Marey MV, Sukhikh GT. Interrelation between miRNAs Expression Associated with Redox State Fluctuations, Immune and Inflammatory Response Activation, and Neonatal Outcomes in Complicated Pregnancy, Accompanied by Placental Insufficiency. Antioxidants. 2023; 12(1):6. https://doi.org/10.3390/antiox12010006
Chicago/Turabian StyleGusar, Vladislava A., Angelika V. Timofeeva, Vitaliy V. Chagovets, Mikhail Yu. Vysokikh, Nataliya E. Kan, Ludmila A. Manukhova, Maria V. Marey, and Gennadiy T. Sukhikh. 2023. "Interrelation between miRNAs Expression Associated with Redox State Fluctuations, Immune and Inflammatory Response Activation, and Neonatal Outcomes in Complicated Pregnancy, Accompanied by Placental Insufficiency" Antioxidants 12, no. 1: 6. https://doi.org/10.3390/antiox12010006
APA StyleGusar, V. A., Timofeeva, A. V., Chagovets, V. V., Vysokikh, M. Y., Kan, N. E., Manukhova, L. A., Marey, M. V., & Sukhikh, G. T. (2023). Interrelation between miRNAs Expression Associated with Redox State Fluctuations, Immune and Inflammatory Response Activation, and Neonatal Outcomes in Complicated Pregnancy, Accompanied by Placental Insufficiency. Antioxidants, 12(1), 6. https://doi.org/10.3390/antiox12010006