
Anti-Wrinkling Effect of 3,4,5-tri-O-caffeoylquinic Acid from the Roots of Nymphoides peltata through MAPK/AP-1, NF-κB, and Nrf2 Signaling in UVB-Irradiated HaCaT Cells

 ,
,  ,
, 
Abstract
:1. Introduction
2. Materials and Methods
2.1. General
2.2. Plant Material
2.3. Isolation of Compounds from N. peltata Extract
3-O-β-D-Glucopyranosyl-Oleanolic Acid 28-O-β-D-Glucuronopyranoside (2)
2.4. Cell Culture and UVB Irradiation
2.5. Cell Viability
2.6. ROS Measurement (DCFDA Assay)
2.7. Luciferase Reporter Gene Assay
2.8. Real-Time Quantitative PCR (qPCR)
2.9. Western Blot Analysis
2.10. Determination of MMP-1 Secretions by ELISA
2.11. Statistical Analysis
3. Results
3.1. Isolation and Structural Identification of Compounds from N. peltata
3.2. Effects of Compounds Isolated from N. peltata on Nrf2–ARE Luciferase Activity in HaCaT Cells
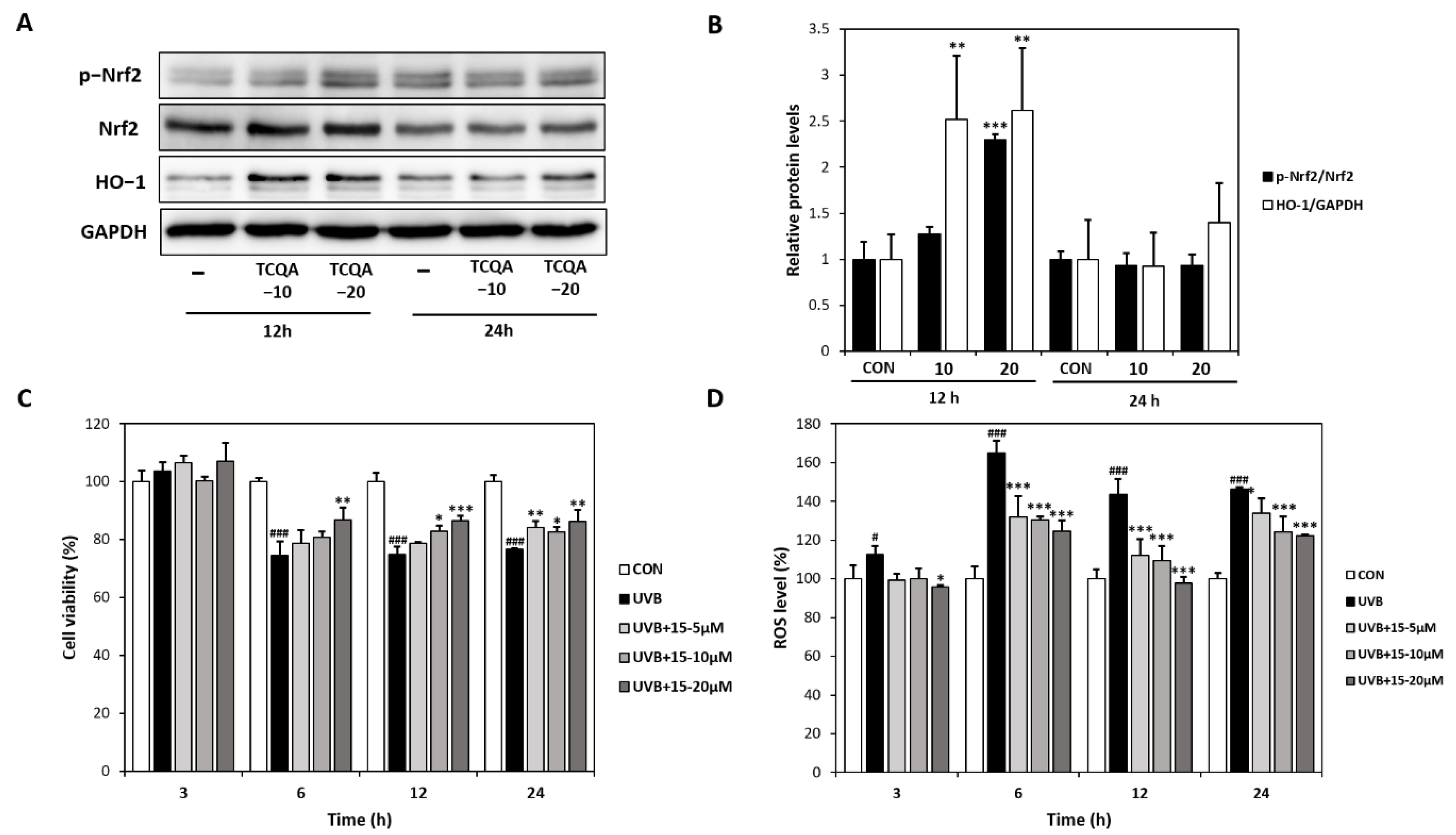
3.3. Effects of TCQA (15) on Antioxidant Activity in HaCaT Cells
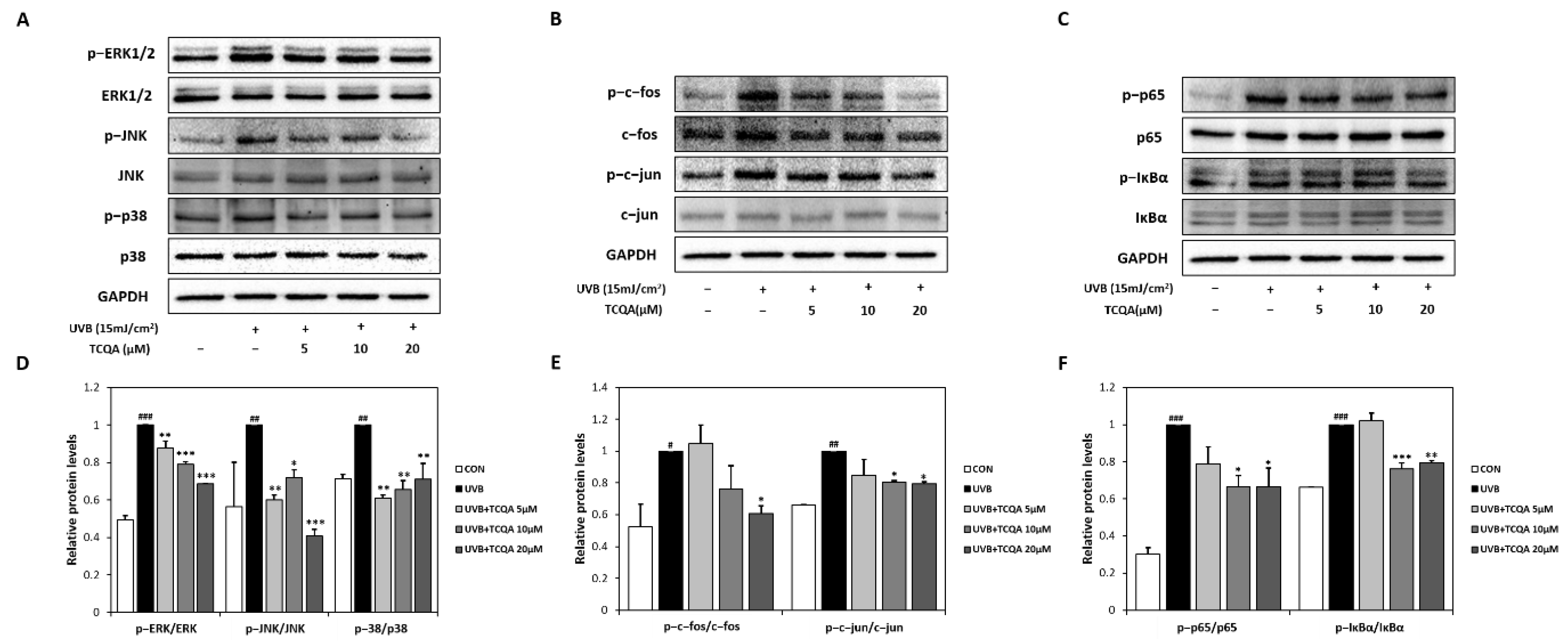
3.4. Effects of TCQA on MAPK-, AP-1-, and NF-κB-Related Transcription Factors in HaCaT Cells Exposed to UVB
3.5. Effects of TCQA on AP-1 Activity in PMA-Induced HaCaT Cells and NF-κb Activity in TNF-α/IFN-γ-Induced HaCaT Cells
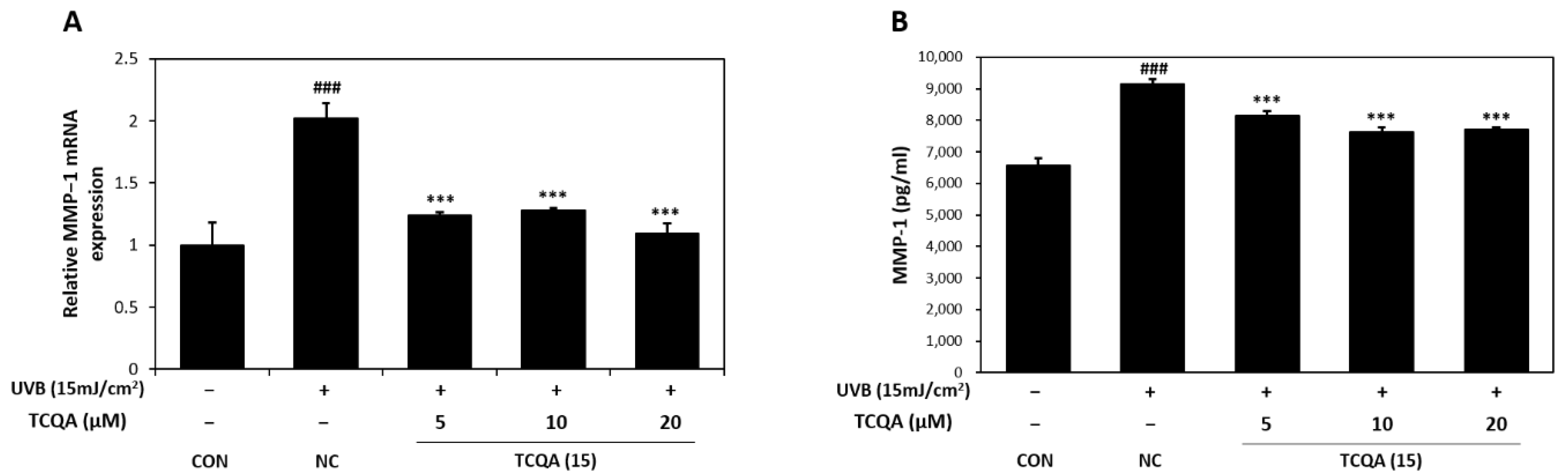
3.6. Effects of TCQA on MMP-1 Expression in HaCaT Cells Exposed to UVB
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Mohd Zaid, N.A.; Sekar, M.; Bonam, S.R.; Gan, S.H.; Lum, P.T.; Begum, M.Y.; Mat Rani, N.N.I.; Vaijanathappa, J.; Wu, Y.S.; Subramaniyan, V.; et al. Promising Natural Products in New Drug Design, Development, and Therapy for Skin Disorders: An Overview of Scientific Evidence and Understanding Their Mechanism of Action. Drug Des. Dev. Ther. 2022, 16, 23–66. [Google Scholar] [CrossRef] [PubMed]
- Franco, A.C.; Aveleira, C.; Cavadas, C. Skin Senescence: Mechanisms and Impact on Whole-Body Aging. Trends Mol. Med. 2022, 28, 97–109. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Y.; He, L.; Zhang, N.; Ma, L.; Yao, L. Photoprotective Effect of Artemisia sieversiana Ehrhart Essential Oil Against UVB-induced Photoaging in Mice. Photochem. Photobiol. 2022, 98, 958–968. [Google Scholar] [CrossRef]
- Tanveer, M.A.; Rashid, H.; Tasduq, S.A. Molecular Basis of Skin Photoaging and Therapeutic Interventions by Plant-Derived Natural Product Ingredients: A Comprehensive Review. Heliyon 2023, 9, e13580. [Google Scholar] [CrossRef] [PubMed]
- Bayo Jimenez, M.T.; Frenis, K.; Hahad, O.; Steven, S.; Cohen, G.; Cuadrado, A.; Münzel, T.; Daiber, A. Protective Actions of Nuclear Factor Erythroid 2-Related Factor 2 (NRF2) and Downstream Pathways against Environmental Stressors. Free Radic. Biol. Med. 2022, 187, 72–91. [Google Scholar] [CrossRef]
- Sharma, V.; Kaur, A.; Singh, T.G. Counteracting Role of Nuclear Factor Erythroid 2-Related Factor 2 Pathway in Alzheimer’s Disease. Biomed. Pharmacother. 2020, 129, 110373. [Google Scholar] [CrossRef]
- Kasai, S.; Shimizu, S.; Tatara, Y.; Mimura, J.; Itoh, K. Regulation of Nrf2 by Mitochondrial Reactive Oxygen Species in Physiology and Pathology. Biomolecules 2020, 10, 320. [Google Scholar] [CrossRef]
- Zhang, Q.; Pi, J.; Woods, C.G.; Andersen, M.E. A Systems Biology Perspective on Nrf2-Mediated Antioxidant Response. Toxicol. Appl. Pharmacol. 2010, 244, 84–97. [Google Scholar] [CrossRef]
- Liu, T.; Xia, Q.; Lv, Y.; Wang, Z.; Zhu, S.; Qin, W.; Yang, Y.; Liu, T.; Wang, X.; Zhao, Z.; et al. ErZhiFormula Prevents UV-Induced Skin Photoaging by Nrf2/HO-1/NQO1 Signaling: An in Vitro and in Vivo Studies. J. Ethnopharmacol. 2023, 309, 115935. [Google Scholar] [CrossRef]
- Chartoumpekis, D.; Ziros, P.G.; Psyrogiannis, A.; Kyriazopoulou, V.; Papavassiliou, A.G.; Habeos, I.G. Simvastatin Lowers Reactive Oxygen Species Level by Nrf2 Activation via PI3K/Akt Pathway. Biochem. Biophys. Res. Commun. 2010, 396, 463–466. [Google Scholar] [CrossRef]
- Papaccio, F.; D’Arino, A.; Caputo, S.; Bellei, B. Focus on the Contribution of Oxidative Stress in Skin Aging. Antioxidants 2022, 11, 1121. [Google Scholar] [CrossRef]
- Lee, C.S.; Bae, I.-H.; Han, J.; Choi, G.; Hwang, K.-H.; Kim, D.-H.; Yeom, M.-H.; Park, Y.-H.; Park, M. Compound K Inhibits MMP-1 Expression through Suppression of c-Src-Dependent ERK Activation in TNF-α-Stimulated Dermal Fibroblast. Exp. Dermatol. 2014, 23, 819–824. [Google Scholar] [CrossRef]
- Kirindage, K.G.I.S.; Jayasinghe, A.M.K.; Han, E.-J.; Jee, Y.; Kim, H.-J.; Do, S.G.; Fernando, I.P.S.; Ahn, G. Fucosterol Isolated from Dietary Brown Alga Sargassum horneri Protects TNF-α/IFN-γ-Stimulated Human Dermal Fibroblasts Via Regulating Nrf2/HO-1 and NF-κB/MAPK Pathways. Antioxidants 2022, 11, 1429. [Google Scholar] [CrossRef]
- Rahman, I. Oxidative Stress, Transcription Factors and Chromatin Remodelling in Lung Inflammation. Biochem. Pharmacol. 2002, 64, 935–942. [Google Scholar] [CrossRef]
- Jabłońska-Trypuć, A.; Matejczyk, M.; Rosochacki, S. Matrix Metalloproteinases (MMPs), the Main Extracellular Matrix (ECM) Enzymes in Collagen Degradation, as a Target for Anticancer Drugs. J. Enzym. Inhib. Med. Chem. 2016, 31, 177–183. [Google Scholar] [CrossRef] [PubMed]
- Pillai, S.; Oresajo, C.; Hayward, J. Ultraviolet Radiation and Skin Aging: Roles of Reactive Oxygen Species, Inflammation and Protease Activation, and Strategies for Prevention of Inflammation-Induced Matrix Degradation—A Review. Int. J. Cosmet. Sci. 2005, 27, 17–34. [Google Scholar] [CrossRef]
- Brenneisen, P.; Sies, H.; Scharffetter-Kochanek, K. Ultraviolet-B Irradiation and Matrix Metalloproteinases. Ann. N. Y. Acad. Sci. 2002, 973, 31–43. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.H.; Lee, M.J.; Lee, S.R.; Kim, K.H.; Cho, K.H.; Eun, H.C.; Chung, J.H. Augmentation of UV-Induced Skin Wrinkling by Infrared Irradiation in Hairless Mice. Mech. Ageing Dev. 2005, 126, 1170–1177. [Google Scholar] [CrossRef] [PubMed]
- Oh, J.H.; Joo, Y.H.; Karadeniz, F.; Ko, J.; Kong, C.-S. Syringaresinol Inhibits UVA-Induced MMP-1 Expression by Suppression of MAPK/AP-1 Signaling in HaCaT Keratinocytes and Human Dermal Fibroblasts. Int. J. Mol. Sci. 2020, 21, 3981. [Google Scholar] [CrossRef]
- Kim, M.-S.; Kim, Y.K.; Cho, K.H.; Chung, J.H. Regulation of Type I Procollagen and MMP-1 Expression after Single or Repeated Exposure to Infrared Radiation in Human Skin. Mech. Ageing Dev. 2006, 127, 875–882. [Google Scholar] [CrossRef]
- Wang, Y.; Wang, L.; Wen, X.; Hao, D.; Zhang, N.; He, G.; Jiang, X. NF-κB Signaling in Skin Aging. Mech. Ageing Dev. 2019, 184, 111160. [Google Scholar] [CrossRef] [PubMed]
- Sunil, M.A.; Sunitha, V.S.; Santhakumaran, P.; Mohan, M.C.; Jose, M.S.; Radhakrishnan, E.K.; Mathew, J. Protective Effect of (+)–Catechin against Lipopolysaccharide-Induced Inflammatory Response in RAW 264.7 Cells through Downregulation of NF-κB and P38 MAPK. Inflammopharmacology 2021, 29, 1139–1155. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Oh, J.-Y.; Lee, W.; Jeon, Y.-J. Fucoidan Isolated from Hizikia fusiforme Suppresses Ultraviolet B-Induced Photodamage by down-Regulating the Expressions of Matrix Metalloproteinases and pro-Inflammatory Cytokines via Inhibiting NF-κB, AP-1, and MAPK Signaling Pathways. Int. J. Biol. Macromol. 2021, 166, 751–759. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.; Li, L.; Huang, G.; Zhou, T.; Zhang, X.; Leng, X.; Chen, Z.; Lin, J. Embelia laeta Aqueous Extract Suppresses Acute Inflammation via Decreasing COX-2/iNOS Expression and Inhibiting NF-κB Pathway. J. Ethnopharmacol. 2021, 281, 114575. [Google Scholar] [CrossRef]
- Cavinato, M.; Waltenberger, B.; Baraldo, G.; Grade, C.V.C.; Stuppner, H.; Jansen-Dürr, P. Plant Extracts and Natural Compounds Used against UVB-Induced Photoaging. Biogerontology 2017, 18, 499–516. [Google Scholar] [CrossRef]
- Mukherjee, P.K.; Maity, N.; Nema, N.K.; Sarkar, B.K. Bioactive Compounds from Natural Resources against Skin Aging. Phytomedicine 2011, 19, 64–73. [Google Scholar] [CrossRef]
- Tippery, N.P.; Sokolik, J.T. Phylogenetics and Seed Morphology of African Nymphoides (Menyanthaceae). Nord. J. Bot. 2020, 38, njb02545. [Google Scholar] [CrossRef]
- Lee, B.Y.; Nam, G.H.; Lee, J.Y.; Park, C.H.; Lim, C.E.; Kim, M.H.; Lee, S.J.; Roh, T.K.; Lim, J.A.; Han, J.E. National List of Species of Korea (Vascular Plants); National Institute of Biological Resources: Incheon, Republic of Korea, 2011; p. 633.
- Nishihiro, J.; Uesugi, R.; Takagawa, S.; Washitani, I. Toward the Restoration of a Sustainable Population of a Threatened Aquatic Plant, Nymphoides peltata: Integrated Genetic/Demographic Studies and Practices. Biol. Conserv. 2009, 142, 1906–1912. [Google Scholar] [CrossRef]
- Johansson, S.; Göransson, U.; Luijendijk, T.; Backlund, A.; Claeson, P.; Bohlin, L. A Neutrophil Multitarget Functional Bioassay to Detect Anti-Inflammatory Natural Products. J. Nat. Prod. 2002, 65, 32–41. [Google Scholar] [CrossRef]
- Kim, T.-Y.; Park, N.-J.; Jegal, H.; Paik, J.-H.; Choi, S.; Kim, S.-N.; Yang, M.H. Nymphoides peltata Root Extracts Improve Atopic Dermatitis by Regulating Skin Inflammatory and Anti-Oxidative Enzymes in 2,4-Dinitrochlorobenzene (DNCB)-Induced SKH-1 Hairless Mice. Antioxidants 2023, 12, 873. [Google Scholar] [CrossRef]
- Lee, D.; Kim, J.-Y.; Kwon, H.C.; Kwon, J.; Jang, D.S.; Kang, K.S. Dual Beneficial Effects of α-Spinasterol Isolated from Aster Pseudoglehnii on Glucose Uptake in Skeletal Muscle Cells and Glucose-Stimulated Insulin Secretion in Pancreatic β-Cells. Plants 2022, 11, 658. [Google Scholar] [CrossRef] [PubMed]
- Kang, J.E.; Jeon, B.J.; Park, M.Y.; Kim, B.S. Inhibitory Activity of Sedum Middendorffianum-Derived 4-Hydroxybenzoic Acid and Vanillic Acid on the Type III Secretion System of Pseudomonas Syringae Pv. Tomato DC3000. Plant Pathol. J. 2020, 36, 608. [Google Scholar] [CrossRef]
- Colombo, R.; Batista, A.N.D.L.; Bomfim, G.C.; Burgos, R.C.; Cavalheiro, A.J.; Bolzani, V.D.S.; Silva, D.H.S.; Reimberg, M.C.H. Validated High-Performance Liquid Chromatographic Method for the Standardisation of Ptychopetalum olacoides Benth., Olacaceae, Commercial Extracts. Rev. Bras. Farmacogn. 2010, 20, 781–788. [Google Scholar] [CrossRef]
- Karthikeyan, R.; Devadasu, C.; Srinivasa Babu, P. Isolation, Characterization, and RP-HPLC Estimation of p-Coumaric Acid from Methanolic Extract of Durva Grass (Cynodon dactylon Linn.) (Pers.). Int. J. Anal. Chem. 2015, 2015, 201386. [Google Scholar] [CrossRef]
- Jeong, C.-H.; Jeong, H.R.; Choi, G.N.; Kim, D.-O.; Lee, U.K.; Heo, H.J. Neuroprotective and Anti-Oxidant Effects of Caffeic Acid Isolated from Erigeron annuus Leaf. Chin. Med. 2011, 6, 25. [Google Scholar] [CrossRef] [PubMed]
- Jain, P.G.; Surana, S.J. Isolation, Characterization and Hypolipidemic Activity of Ferulic Acid in High-Fat-Diet-Induced Hyperlipidemia in Laboratory Rats. EXCLI J. 2016, 15, 599. [Google Scholar] [CrossRef]
- Wong, S.K.; Lim, Y.Y.; Ling, S.K.; Chan, E.W.C. Caffeoylquinic Acids in Leaves of Selected Apocynaceae Species: Their Isolation and Content. Pharmacogn. Res. 2014, 6, 67. [Google Scholar] [CrossRef]
- Shi, S.; Huang, K.; Zhang, Y.; Zhao, Y.; Du, Q. Purification and Identification of Antiviral Components from Laggera pterodonta by High-Speed Counter-Current Chromatography. J. Chromatogr. B 2007, 859, 119–124. [Google Scholar] [CrossRef]
- Islam, M.S.; Yoshimoto, M.; Yahara, S.; Okuno, S.; Ishiguro, K.; Yamakawa, O. Identification and Characterization of Foliar Polyphenolic Composition in Sweetpotato (Ipomoea batatas L.) Genotypes. J. Agric. Food Chem. 2002, 50, 3718–3722. [Google Scholar] [CrossRef]
- Magalhaes, A.F.; Goulart de Azevedo Tozzi, A.M.; Santos, C.C.; Serrano, D.R.; Zanotti-Magalhaes, E.M.; Magalhaes, E.G.; Magalhaes, L.A. Saponins from Swartzia langsdorffii: Biological Activities. Mem. Inst. Oswaldo Cruz 2003, 98, 713–718. [Google Scholar] [CrossRef] [PubMed]
- Mizui, F.; Kasai, R.; Ohtani, K.; Tanaka, O. Saponins from Bran of Quinoa, Chenopodium quinoa WILLD. II. Chem. Pharm. Bull. 1990, 38, 375–377. [Google Scholar] [CrossRef]
- Neopane, D.; Ansari, V.A.; Singh, A. Ferulic Acid: Signaling Pathways in Aging. Drug Res. 2023, 73, 318–324. [Google Scholar] [CrossRef]
- Feng, J.; Zhang, P.; Chen, X.; He, G. PI3K and ERK/Nrf2 Pathways Are Involved in Oleanolic Acid-Induced Heme Oxygenase-1 Expression in Rat Vascular Smooth Muscle Cells. J. Cell. Biochem. 2011, 112, 1524–1531. [Google Scholar] [CrossRef]
- Ha, J.W.; Boo, Y.C. Siegesbeckiae Herba Extract and Chlorogenic Acid Ameliorate the Death of HaCaT Keratinocytes Exposed to Airborne Particulate Matter by Mitigating Oxidative Stress. Antioxidants 2021, 10, 1762. [Google Scholar] [CrossRef]
- Hwang, B.-M.; Noh, E.-M.; Kim, J.-S.; Kim, J.-M.; You, Y.-O.; Hwang, J.-K.; Kwon, K.-B.; Lee, Y.-R. Curcumin Inhibits UVB-Induced Matrix Metalloproteinase-1/3 Expression by Suppressing the MAPK-P38/JNK Pathways in Human Dermal Fibroblasts. Exp. Dermatol. 2013, 22, 371–374. [Google Scholar] [CrossRef]
- Hwang, E.; Lin, P.; Ngo, H.T.T.; Gao, W.; Wang, Y.-S.; Yu, H.-S.; Yi, T.-H. Icariin and Icaritin Recover UVB-Induced Photoaging by Stimulating Nrf2/ARE and Reducing AP-1 and NF-κB Signaling Pathways: A Comparative Study on UVB-Irradiated Human Keratinocytes. Photochem. Photobiol. Sci. 2018, 17, 1396–1408. [Google Scholar] [CrossRef] [PubMed]
- Khan, Z.; Chowdhury, N.; Sharmin, S.; Sohrab, M.H. Medicinal Values of Aquatic Plant Genus Nymphoides Grown in Asia: A Review. Asian Pac. J. Trop. Biomed. 2018, 8, 113. [Google Scholar] [CrossRef]
- Nakai, K.; Tsuruta, D. What Are Reactive Oxygen Species, Free Radicals, and Oxidative Stress in Skin Diseases? Int. J. Mol. Sci. 2021, 22, 10799. [Google Scholar] [CrossRef]
- Ikehata, H.; Yamamoto, M. Roles of the KEAP1-NRF2 System in Mammalian Skin Exposed to UV Radiation. Toxicol. Appl. Pharmacol. 2018, 360, 69–77. [Google Scholar] [CrossRef]
- Campbell, N.K.; Fitzgerald, H.K.; Dunne, A. Regulation of Inflammation by the Antioxidant Haem Oxygenase 1. Nat. Rev. Immunol. 2021, 21, 411–425. [Google Scholar] [CrossRef] [PubMed]
- Xie, Y.; Chen, Z.; Wu, Z. Four-Octyl Itaconate Attenuates UVB-Induced Melanocytes and Keratinocytes Apoptosis by Nrf2 Activation-Dependent ROS Inhibition. Oxidative Med. Cell. Longev. 2022, 2022, e9897442. [Google Scholar] [CrossRef]
- Wang, S.-H.; Chen, Y.-S.; Lai, K.-H.; Lu, C.-K.; Chang, H.-S.; Wu, H.-C.; Yen, F.-L.; Chen, L.-Y.; Lee, J.-C.; Yen, C.-H. Prinsepiae Nux Extract Activates NRF2 Activity and Protects UVB-Induced Damage in Keratinocyte. Antioxidants 2022, 11, 1755. [Google Scholar] [CrossRef]
- Guo, C.; Ning, X.; Zhang, J.; Zhang, C.; Wang, J.; Su, L.; Han, J.; Ma, N. Ultraviolet B Radiation Induces Oxidative Stress and Apoptosis in Human Lens Epithelium Cells by Activating NF-κB Signaling to down-Regulate Sodium Vitamin C Transporter 2 (SVCT2) Expression. Cell Cycle 2023, 22, 1450–1462. [Google Scholar] [CrossRef] [PubMed]
- Kriete, A.; Mayo, K.L. Atypical Pathways of NF-κB Activation and Aging. Exp. Gerontol. 2009, 44, 250–255. [Google Scholar] [CrossRef]
- Giridharan, S.; Srinivasan, M. Mechanisms of NF-κB p65 and Strategies for Therapeutic Manipulation. J. Inflamm. Res. 2018, 11, 407–419. [Google Scholar] [CrossRef] [PubMed]
- Liu, T.; Zhang, L.; Joo, D.; Sun, S.-C. NF-κB Signaling in Inflammation. Signal Transduct. Target. Ther. 2017, 2, 17023. [Google Scholar] [CrossRef]
- Young, C.N.; Koepke, J.I.; Terlecky, L.J.; Borkin, M.S.; Boyd, S.L.; Terlecky, S.R. Reactive Oxygen Species in Tumor Necrosis Factor-α-Activated Primary Human Keratinocytes: Implications for Psoriasis and Inflammatory Skin Disease. J. Investig. Dermatol. 2008, 128, 2606–2614. [Google Scholar] [CrossRef]
- Tanaka, K.; Asamitsu, K.; Uranishi, H.; Iddamalgoda, A.; Ito, K.; Kojima, H.; Okamoto, T. Protecting Skin Photoaging by NF-κB Inhibitor. Curr. Drug Metab. 2010, 11, 431–435. [Google Scholar] [CrossRef]
- García-García, V.A.; Alameda, J.P.; Page, A.; Casanova, M.L. Role of NF-κB in Ageing and Age-Related Diseases: Lessons from Genetically Modified Mouse Models. Cells 2021, 10, 1906. [Google Scholar] [CrossRef] [PubMed]
- Xue, N.; Liu, Y.; Jin, J.; Ji, M.; Chen, X. Chlorogenic Acid Prevents UVA-Induced Skin Photoaging through Regulating Collagen Metabolism and Apoptosis in Human Dermal Fibroblasts. Int. J. Mol. Sci. 2022, 23, 6941. [Google Scholar] [CrossRef]
- Li, R.; Tao, M.; Wu, T.; Zhou, Z.; Xu, T.; Pan, S.; Xu, X. A promising strategy for investigating the anti-aging effect of natural compounds: A case study of caffeoylquinic acids. Food Funct. 2021, 12, 8583. [Google Scholar] [CrossRef] [PubMed]
- Maurin, C.; Bailly, F.; Cotelle, P. Structure-Activity Relationships of HIV-1 Integrase Inhibitors-Enzyme-Ligand Interactions. Curr. Med. Chem. 2003, 10, 1795–1810. [Google Scholar] [CrossRef]
- Alcázar Magaña, A.; Kamimura, N.; Soumyanath, A.; Stevens, J.F.; Maier, C.S. Caffeoylquinic Acids: Chemistry, Biosynthesis, Occurrence, Analytical Challenges, and Bioactivity. Plant J. 2021, 107, 1299–1319. [Google Scholar] [CrossRef]
- Xu, J.-G.; Hu, Q.-P.; Liu, Y. Antioxidant and DNA-Protective Activities of Chlorogenic Acid Isomers. J. Agric. Food Chem. 2012, 60, 11625–11630. [Google Scholar] [CrossRef]
- Indy Tamayose, C.; dos Santos, E.A.; Roque, N.; Costa-Lotufo, L.V.; Pena Ferreira, M.J. Caffeoylquinic Acids: Separation Method, Antiradical Properties and Cytotoxicity. Chem. Biodivers. 2019, 16, e1900093. [Google Scholar] [CrossRef]
- Arulselvan, P.; Fard, M.T.; Tan, W.S.; Gothai, S.; Fakurazi, S.; Norhaizan, M.E.; Kumar, S.S. Role of Antioxidants and Natural Products in Inflammation. Oxidative Med. Cell. Longev. 2016, 2016, e5276130. [Google Scholar] [CrossRef]
- Ge, L.; Wan, H.; Tang, S.; Chen, H.; Li, J.; Zhang, K.; Zhou, B.; Fei, J.; Wu, S.; Zeng, X. Novel Caffeoylquinic Acid Derivatives from Lonicera japonica Thunb. Flower Buds Exert Pronounced Anti-HBV Activities. RSC Adv. 2018, 8, 35374–35385. [Google Scholar] [CrossRef]
- Liu, W.; Li, J.; Zhang, X.; Zu, Y.; Yang, Y.; Liu, W.; Xu, Z.; Gao, H.; Sun, X.; Jiang, X.; et al. Current Advances in Naturally Occurring Caffeoylquinic Acids: Structure, Bioactivity, and Synthesis. J. Agric. Food Chem. 2020, 68, 10489–10516. [Google Scholar] [CrossRef] [PubMed]
- Sasaki, K.; Han, J.; Shigemori, H.; Isoda, H. Caffeoylquinic Acid Induces ATP Production and Energy Metabolism in Human Neurotypic SH-SY5Y Cells. Nutr. Aging 2012, 1, 141–150. [Google Scholar] [CrossRef]
- Abdel Motaal, A.; Ezzat, S.M.; Tadros, M.G.; El-Askary, H.I. In Vivo Anti-Inflammatory Activity of Caffeoylquinic Acid Derivatives from Solidago virgaurea in Rats. Pharm. Biol. 2016, 54, 2864–2870. [Google Scholar] [CrossRef]
- Tang, B.; Huang, Y.; Yang, H.; Tang, P.; Li, H. Molecular Mechanism of the Binding of 3,4,5-Tri-O-caffeoylquinic Acid to Human Serum Albumin: Saturation Transfer Difference NMR, Multi-Spectroscopy, and Docking Studies. J. Photochem. Photobiol. B 2016, 165, 24–33. [Google Scholar] [CrossRef] [PubMed]
- Kim, E.K.; Choi, E.-J. Pathological Roles of MAPK Signaling Pathways in Human Diseases. Biochim. Biophys. Acta BBA Mol. Basis Dis. 2010, 1802, 396–405. [Google Scholar] [CrossRef]
- Wada, T.; Penninger, J.M. Mitogen-Activated Protein Kinases in Apoptosis Regulation. Oncogene 2004, 23, 2838–2849. [Google Scholar] [CrossRef] [PubMed]
- Long, Y.; Wang, W.; Zhang, Y.; Du, F.; Zhang, S.; Li, Z.; Deng, J.; Li, J. Photoprotective Effects of Dendrobium nobile Lindl. Polysaccharides against UVB-Induced Oxidative Stress and Apoptosis in HaCaT Cells. Int. J. Mol. Sci. 2023, 24, 6120. [Google Scholar] [CrossRef]
- Mantena, S.K.; Katiyar, S.K. Grape Seed Proanthocyanidins Inhibit UV-Radiation-Induced Oxidative Stress and Activation of MAPK and NF-κB Signaling in Human Epidermal Keratinocytes. Free Radic. Biol. Med. 2006, 40, 1603–1614. [Google Scholar] [CrossRef]
- Liou, J.-T.; Lin, C.-S.; Liao, Y.-C.; Ho, L.-J.; Yang, S.-P.; Lai, J.-H. JNK/AP-1 Activation Contributes to Tetrandrine Resistance in T-Cell Acute Lymphoblastic Leukaemia. Acta Pharmacol. Sin. 2017, 38, 1171–1183. [Google Scholar] [CrossRef] [PubMed]
- Zhu, H.; Hao, Z.; Xing, Z.; Tan, J.; Zhao, Y.; Li, M. Impinging Flow Induces Expression of Monocyte Chemoattractant Protein-1 in Endothelial Cells Through Activation of the c-Jun N-Terminal Kinase/c-Jun/p38/c-Fos Pathway. World Neurosurg. 2022, 164, e681–e693. [Google Scholar] [CrossRef]
- Muthusamy, V.; Piva, T.J. The UV Response of the Skin: A Review of the MAPK, NFκB and TNFα Signal Transduction Pathways. Arch. Dermatol. Res. 2010, 302, 5–17. [Google Scholar] [CrossRef]
- Janson, N.D.; Jehanathan, N.; Jung, S.; Priyathilaka, T.T.; Nam, B.-H.; Kim, M.-J.; Lee, J. Insight into the Molecular Function and Transcriptional Regulation of Activator Protein 1 (AP-1) Components c-Jun/c-Fos Ortholog in Red Lip Mullet (Liza haematocheila). Fish Shellfish Immunol. 2019, 93, 597–611. [Google Scholar] [CrossRef]
- Briganti, S.; Picardo, M. Antioxidant Activity, Lipid Peroxidation and Skin Diseases. What’s New. J. Eur. Acad. Dermatol. Venereol. 2003, 17, 663–669. [Google Scholar] [CrossRef]
- Xiao, Z.; Yang, S.; Chen, J.; Li, C.; Zhou, C.; Hong, P.; Sun, S.; Qian, Z.-J. Trehalose against UVB-Induced Skin Photoaging by Suppressing MMP Expression and Enhancing Procollagen I Synthesis in HaCaT Cells. J. Funct. Foods 2020, 74, 104198. [Google Scholar] [CrossRef]
- Kim, H.M.; Lee, D.E.; Park, S.D.; Kim, Y.-T.; Kim, Y.J.; Jeong, J.W.; Jang, S.S.; Ahn, Y.-T.; Sim, J.-H.; Huh, C.-S.; et al. Oral Administration of Lactobacillus plantarum HY7714 Protects Hairless Mouse Against Ultraviolet B-Induced Photoaging. J. Microbiol. Biotechnol. 2014, 24, 1583–1591. [Google Scholar] [CrossRef] [PubMed]
- Wlaschek, M.; Tantcheva-Poór, I.; Naderi, L.; Ma, W.; Schneider, L.A.; Razi-Wolf, Z.; Schüller, J.; Scharffetter-Kochanek, K. Solar UV Irradiation and Dermal Photoaging. J. Photochem. Photobiol. B 2001, 63, 41–51. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Compound 2 | |||||
|---|---|---|---|---|---|
| Position | δH (Multi, J in Hz) | δC | Position | δH (Multi, J in Hz) | δC |
| Oleanolic acid | Glucose | ||||
| 1 | α: 0.88 (m); β: 1.48 (m) | 38.6 CH2 | 1′ | 4.13 (d, 7.5) | 105.8 CH |
| 2 | α: 1.50 (m); β: 1.90 (m) | 25.9 CH2 | 2′ | 2.95 (m) | 74.3 CH |
| 3 | 3.01 (dd, 10.5, 3.5) | 88.4 CH | 3′ | 3.13 (m) | 77.1 CH |
| 4 | 39.2 C | 4′ | 3.13 (m) | 70.0 CH | |
| 5 | 0.71, overlap | 55.5 CH | 5′ | 3.13 (m) | 78.2 CH |
| 6 | α: 1.46 (m); β: 1.30 (m) | 18.2 CH2 | 6′ | 3.61 (dd, 11.0, 5.0); 3.43 (dd, 11.0, 5.5) | 61.1 CH2 |
| 7 | α: 1.36 (m); β: 1.21 (m) | 32.7 CH2 | 2′-OH | 4.87 (d, 4.0) | |
| 8 | 39.6 C | 3′-OH | 4.92 (br. s) | ||
| 9 | 1.48 (m) | 47.6 CH | 4′-OH | 5.04 (d, 4.0) | |
| 10 | 36.7 C | 6′-OH | 4.45 (t, 5.5) | ||
| 11 | 1.79 (m) | 23.4 CH2 | Glucuronic acid | ||
| 12 | 5.16 (br. s) | 122.2 CH2 | 1″ | 5.23 (d, 8.0) | 94.6 CH |
| 13 | 143.9 C | 2″ | 3.11 (m) | 72.8 CH | |
| 14 | 41.7 C | 3″ | 3.21 (m) | 77.1 CH | |
| 15 | α: 0.95 (m); β: 1.72 (m) | 27.6 CH2 | 4″ | 3.15 (m) | 72.6 CH |
| 16 | α: 1.95 (m); β: 1.59 (m) | 23.0 CH2 | 5″ | 3.28 (m) | 74.6 CH |
| 17 | 46.0 C | 6″ | 174.3 C | ||
| 18 | 2.73 (dd, 13.5, 3.0) | 41.2 CH | |||
| 19 | 46.4 CH2 | ||||
| 20 | α: 1.62 (m); β: 1.08 (m) | 30.8 C | |||
| 21 | α: 1.34 (m); β: 1.16 (m) | 33.6 CH2 | |||
| 22 | 1.51 (m) | 32.1 CH2 | |||
| 23-CH3 | 0.96 (s) | 28.1 CH3 | |||
| 24-CH3 | 0.74 (s) | 17.0 CH3 | |||
| 25-CH3 | 0.86 (s) | 15.7 CH3 | |||
| 26-CH3 | 0.67 (s) | 17.1 CH3 | |||
| 27-CH3 | 1.07 (s) | 26.0 CH3 | |||
| 28 | 175.7 C | ||||
| 29-CH3 | 0.87 (s) | 33.2 CH3 | |||
| 30-CH3 | 0.86 (s) | 23.8 CH3 | |||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kim, T.-Y.; Park, N.-J.; Jo, B.-G.; Lee, B.S.; Keem, M.-J.; Kwon, T.-H.; Kim, K.H.; Kim, S.-N.; Yang, M.H. Anti-Wrinkling Effect of 3,4,5-tri-O-caffeoylquinic Acid from the Roots of Nymphoides peltata through MAPK/AP-1, NF-κB, and Nrf2 Signaling in UVB-Irradiated HaCaT Cells. Antioxidants 2023, 12, 1899. https://doi.org/10.3390/antiox12101899
Kim T-Y, Park N-J, Jo B-G, Lee BS, Keem M-J, Kwon T-H, Kim KH, Kim S-N, Yang MH. Anti-Wrinkling Effect of 3,4,5-tri-O-caffeoylquinic Acid from the Roots of Nymphoides peltata through MAPK/AP-1, NF-κB, and Nrf2 Signaling in UVB-Irradiated HaCaT Cells. Antioxidants. 2023; 12(10):1899. https://doi.org/10.3390/antiox12101899
Chicago/Turabian StyleKim, Tae-Young, No-June Park, Beom-Geun Jo, Bum Soo Lee, Min-Ji Keem, Taek-Hwan Kwon, Ki Hyun Kim, Su-Nam Kim, and Min Hye Yang. 2023. "Anti-Wrinkling Effect of 3,4,5-tri-O-caffeoylquinic Acid from the Roots of Nymphoides peltata through MAPK/AP-1, NF-κB, and Nrf2 Signaling in UVB-Irradiated HaCaT Cells" Antioxidants 12, no. 10: 1899. https://doi.org/10.3390/antiox12101899
APA StyleKim, T. -Y., Park, N. -J., Jo, B. -G., Lee, B. S., Keem, M. -J., Kwon, T. -H., Kim, K. H., Kim, S. -N., & Yang, M. H. (2023). Anti-Wrinkling Effect of 3,4,5-tri-O-caffeoylquinic Acid from the Roots of Nymphoides peltata through MAPK/AP-1, NF-κB, and Nrf2 Signaling in UVB-Irradiated HaCaT Cells. Antioxidants, 12(10), 1899. https://doi.org/10.3390/antiox12101899