

Tumor Microbial Communities and Thyroid Cancer Development—The Protective Role of Antioxidant Nutrients: Application Strategies and Future Directions



Abstract
:1. Introduction
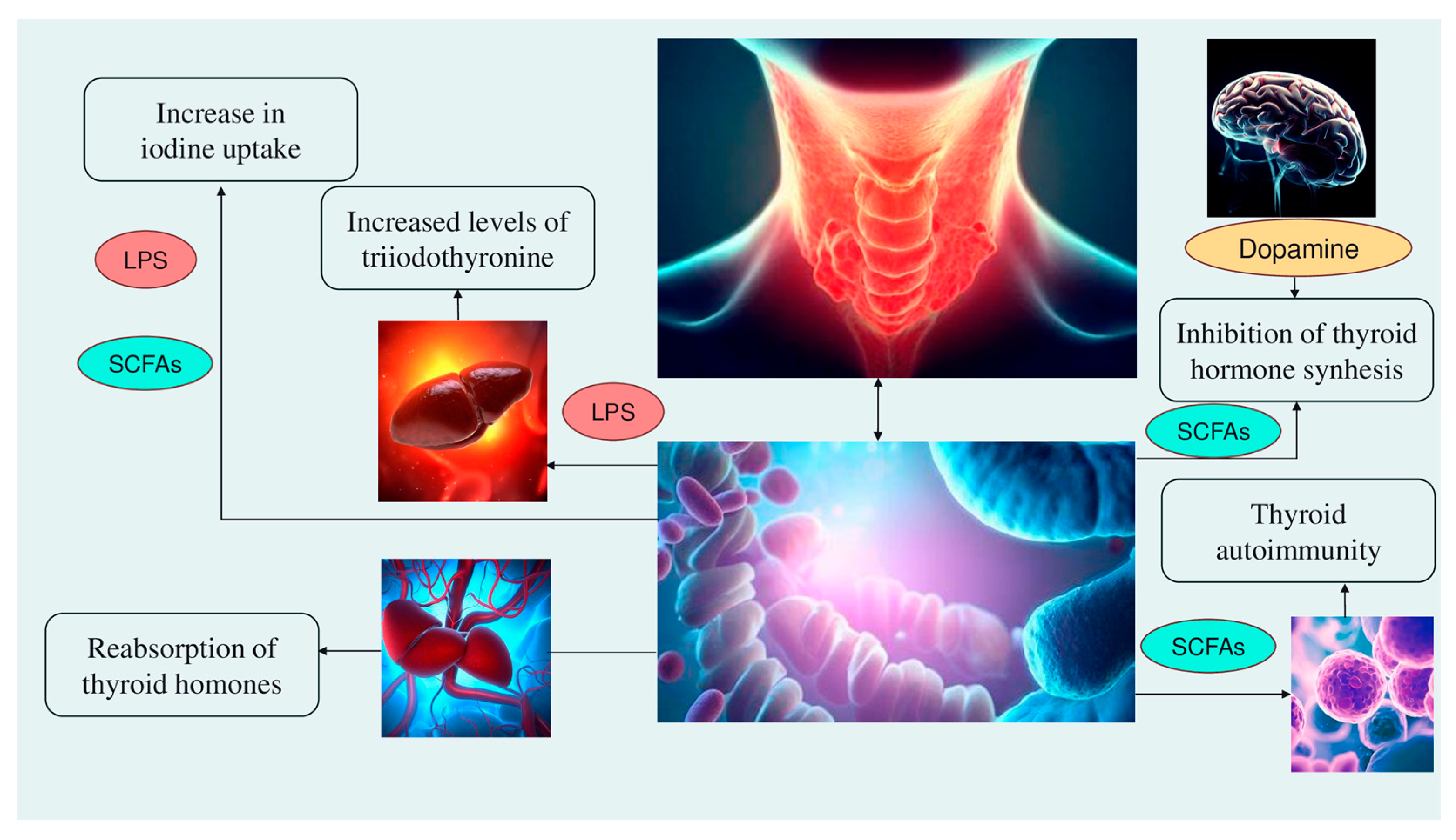
2. The Microbiota and Thyroid Axis
- Alteration of iodine uptake, the main rate-limiting step in thyroid hormonogenesis, affecting the activity of sodium iodide symporter (NIS) through two processes: (a) The binding of the Gram-negative bacterial endotoxin lipopolysaccharide (LPS), released by the gut microbiota, to the thyroid cell toll-like receptor 4 (TLR-4). TLR-4 in turn activates the nuclear factor kappa-light-chain-enhancer of activated B cells (NF-kB), which subsequently promotes NIS transcription through paired box 8 (PAX8) [38]. (b) Alternatively, enhancement of NIS expression may also occur through histone deacetylase (HDAC) inhibition by an important metabolite of the gut microbiota, butyrate (which belongs to the class of short-chain fatty acids—SCFAs; see later in Section 6) [39,40];
- Modulation of activities of iodothyronine deiodinases, enzymes responsible for the conversion of thyroxine (T4) to its active form T3 by type 1 and type 2 deiodinases (D1, expressed mostly in the liver, kidney, thyroid, and pituitary, and D2, expressed primarily in the thyroid, central nervous system, pituitary, developing cochlea, brown adipose tissue, and skeletal muscle [41]) or to reverse T3, its inactive form, by type 3 deiodinase—D3 [42]. This occurs through a complex thyroid–gut axis pathway involving LPS capable of inducing the decrease in D1 activity in the liver [43] and, at the same time, activating D2 in the mediobasal hypothalamus, ultimately promoting the conversion of T4 to T3 [44];
- Modulation of T3 and T4 bioavailability through the deconjugation of sulfoconjugated and glucuroconjugated iodothyronines by bacterial sulfate esterase or β-glucuronidase, respectively, thus inducing the reabsorption of thyroid hormones in the enterohepatic circulation. In humans, a recycling mechanism has been described for steroids hormones, biliary acids, and vitamins, while as for thyroid hormones, direct proof has been only established in animal models [31];
- Regulation of the SCFAs-mediated balance between T helper 17 (Th17) cells and regulatory T cells (Treg), two subtypes of CD4+ lymphocytes exerting opposite effects (release of pro-inflammatory cytokines, i.e., interleukin—IL-17 or anti-inflammatory IL-10, respectively), in autoimmune inflammatory diseases and immune tolerance [35,45]. All these immune cells play a role in the pathogenesis of autoimmune thyroid disease (AITD), like Hashimoto disease (HD) and Graves’ disease. For instance, Prevotella is correlated with reduced proinflammatory Th17 polarization and increased differentiation of anti-inflammatory Treg; therefore, it has been speculated that the Th17/Treg homeostasis regulation might be a potential pathogenic pathway for Prevotella in HT patients [32,46];
- Involvement of the microbiota–gut–brain signaling in dopamine release, synthesis, and bioavailability. Certain species up-/downregulate the system dopamine transporter/dopamine binding efficiency, while others are positively or negatively correlated with the activity of tyrosine hydroxylase, an enzyme involved in dopamine synthesis [47,48]. Furthermore, butyrate’s intrinsic HDAC inhibitor activity influences neurotransmitter levels [47]. Since dopamine inhibits synthesis and secretion of the thyroid-stimulating hormone (TSH), thyroid function may be affected [35] (Figure 1).
3. The Association between Microbiota and Thyroid Cancer
Microbial Communities in Thyroid Cancer Tissues
4. Oxidative Stress in Thyroid Cancer
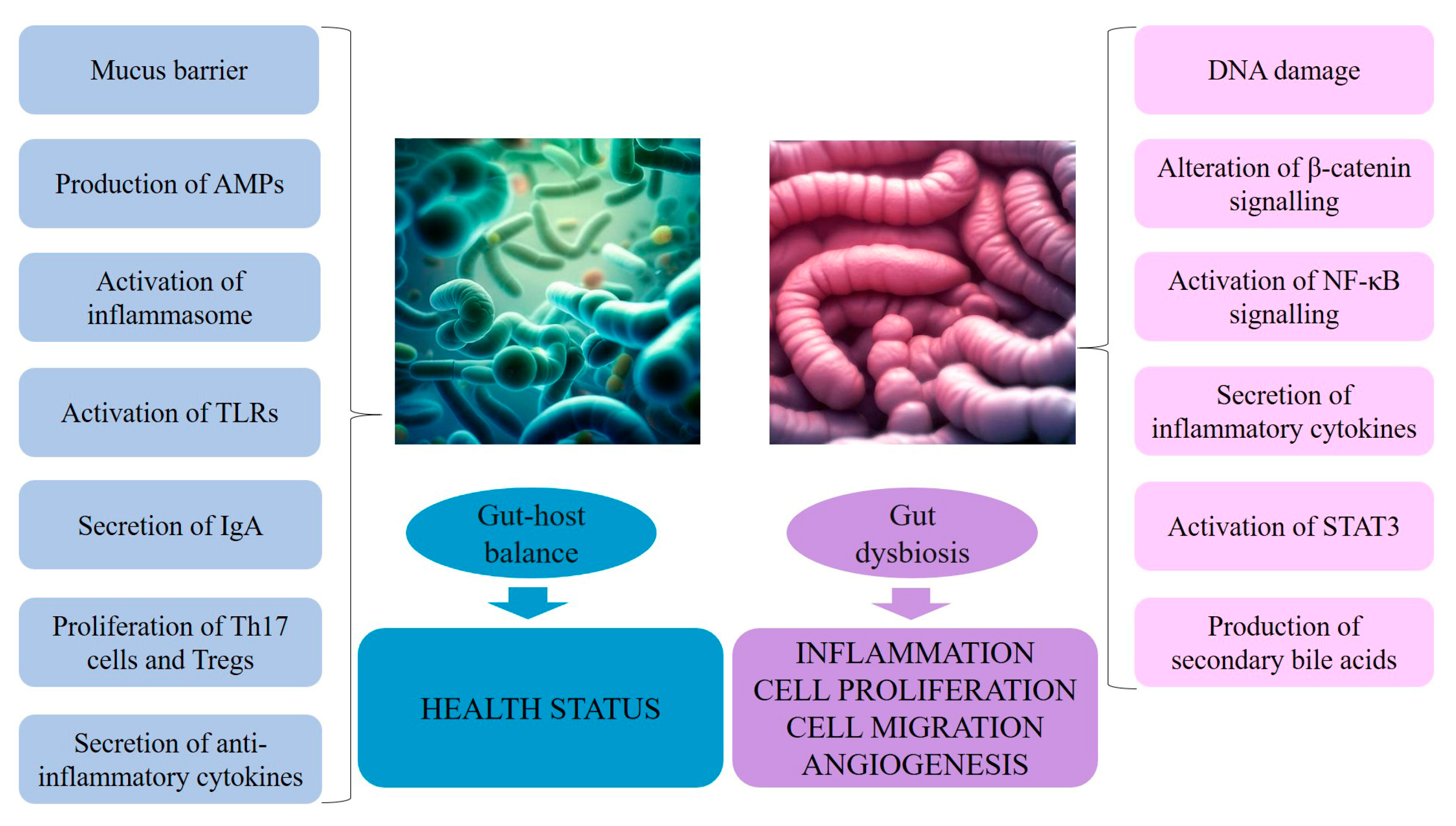
5. The Dual Relationship between the Microbiota and Cancer: The Biological Mechanisms
- DNA damage, both directly by inducing double-strand breaks and indirectly by eliciting high levels of ROS and RNS released from inflammatory cells, such as macrophages, as in colorectal cancer;
- β-Catenin signaling alterations, as observed in gastric cancer and colorectal adenomas and adenocarcinomas, leading to upregulation of genes involved in cellular proliferation, survival, and migration, as well as angiogenesis;
- Pro-inflammatory reactions induced by microorganism-associated molecular patterns that activate TLRs in several cell types, including macrophages, myofibroblasts, epithelial cells, and tumor cells, which, in loop, stimulate NF-κB, a master regulator of cancer-associated inflammation [136];
- Activation of the IL-23–IL-17 axis, which promotes tumor growth and progression [137], tumor necrosis factor–alpha (TNF-α)–TNF receptor signaling, and IL-11–IL-6 family member signaling, all activating the signal transducer and activator of transcription 3 (STAT3) (belonging to the family of STAT transcriptional factors that participate in the regulation of a variety of cellular process, including proliferation, differentiation, inflammation, and stemness) [138,139,140];
- Perturbations of the equilibrium between diet–gut microbiome–bile acid pool size/composition through the production of microbially generated secondary bile acids, like deoxycholic acid (DCA), responsible for activation of mitogenic and proinflammatory effects and subsequent promotion of liver cancer (see later in the text) [141,142] (Figure 2).
5.1. Gut Microorganisms: A Defense against Cancer
- Goblet cells, specialized epithelial cells, are essential to the formation of the mucus barriers [163];
- Paneth cells, secretory cells located in the crypts of Lieberkühn, produce antimicrobial peptides (AMPs) and proteins [164], whose activity is enhanced following signaling from local immune cells in response to the microbiota;
- Upon recognition of microbial peptidoglycan, nucleotide-binding oligomerization domain-containing protein 2 (NOD2) contributes to intestinal homeostasis by molecular signaling through the kinase receptor-interacting protein 2 and NF-κB, and inducing the production of AMPs and mucin [165];
- Microbial metabolites activate pathogen recognition receptors (PRRs), such as NOD-, leucine-rich repeat- and NOD-like receptor family pyrin domain-containing 6 (NLRP6) inflammasome (having a role in the maintenance of a stable microbial community in the intestine), which lead to the secretion of IL-18 and AMPs [165];
- Within adaptative mechanisms of microbial regulation, immunoglobulin A (IgA), secreted by plasma cells, terminally differentiated B cells residing in the intestinal lamina propria [168], acts primarily by inhibiting bacterial adherence to epithelial cells, also having direct effects on bacterial virulence [169];
- Pathogen-associated molecular patterns (e.g., LPS, flagellin) induce antigen-presenting cells, like dendritic cells (DCs); DCs then travel to mesentery, where they stimulate naïve T cells to differentiate into CD4+ Tregs and Th17 cells, which can migrate back to the gut mucosa or enter systemic circulation. While Tregs secrete the anti-inflammatory cytokine IL-10, Th17 cells, through the production of IL-17, can increase the Paneth cell production of antimicrobial peptides;
- SCFAs influence the immune response by a variety of signaling pathways, both in the innate immunity, e.g., activating NLRP3 that is responsible for IL-1β and IL-18 secretion [170]; inhibiting the production of cytokines, chemokines, and calprotectin produced by neutrophils [171]; inducing antimicrobial activity in the intestinal macrophage and increasing resistance to enteropathogens, and in the adaptative immunity, e.g., promoting the proliferation of Tregs [172]; and accelerating cellular metabolism and regulating gene expression to promote B cell differentiation into antibody-producing cells [173].
6. Diet: Enemy or Friend of the Gut Microbiota?
7. Antioxidant Nutrients at the Intersection of Gut Microbiota Health and Protection against Thyroid Cancer
7.1. Probiotics
7.2. Trace Elements
7.3. Vitamins
7.4. Polyphenols
8. Promoting Food Health: Should Citizens and Food Companies Meet Half Way?
9. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
| AITD | Autoimmune thyroid disease |
| AMPs | Antimicrobial peptides |
| ATC | Anaplastic thyroid cancer |
| CAT | Catalase |
| CRP | C-reactive protein |
| D1,,/3 | Deiodinase type 1,2,3 |
| DCs | Dendritic cells |
| DCA | Deoxycholic acid |
| DUOX | Dual oxidase |
| ERK | Extracellular signal-regulated kinase |
| FTC | Follicular thyroid cancer |
| GABA | Gamma-aminobutyric acid |
| FVPTC | Follicular variant of papillary thyroid cancer |
| GPx | Glutathione peroxidase |
| GPR | G protein-coupled receptors |
| GSH | Glutathione |
| H2O2 | Peroxide hydrogen |
| HD | Hashimoto’s disease |
| HDAC | Histone deacetylase |
| HIF-1α | Hypoxia-inducible factor-1 alpha |
| Hp | Helicobacter pylori |
| IL | Interleukin |
| LPS | Lipopolysaccharide |
| MAPK | Mitogen-activated protein kinase |
| NF-kB | Kappa-light-chain-enhancer of activated B cells |
| NIS | Sodium iodide symporter |
| NLRP3,6 | NOD-like receptor family pyrin domain-containing 3,6 |
| NO | Nitric oxide |
| NOCs | N-nitroso compounds |
| NOD | Nucleotide-binding oligomerization domain-containing protein |
| NOX | Nicotinamide adenine dinucleotide phosphate oxidase |
| PAX8 | Paired box 8 |
| PDTC | Poorly differentiated thyroid cancer |
| PRRs | Pathogen recognition receptors |
| PTC | Papillary thyroid cancer |
| RNS | Reactive nitrogen species |
| ROS | Reactive oxygen species |
| rRNA | Ribosomal RNA |
| SCFAs | Short-chain fatty acids |
| Se | Selenium |
| SOD | Superoxide dismutase |
| STAT3 | Signal transducer and activator of transcription 3 |
| T3 | Triiodothyronine |
| T4 | Thyroxine |
| TAMS | Tumor-associated macrophages |
| TC | Thyroid cancer |
| TCPTC | Tall cell papillary thyroid cancer |
| TGF-β1 | Transforming growth factor-beta 1 |
| Th | T helper cell |
| TrxR | Thioredoxin reductase |
| TLR | Toll-like receptor |
| TNF-α | Tumor necrosis factor-alpha |
| TPO | Thyroperoxidase |
| TR | Thyroid receptor |
| TSH | Thyroid-stimulating hormone |
| Treg | Regulatory T cell |
| Zn | Zinc |
References
- Liu, C.J.; Chen, S.Q.; Zhang, S.Y.; Wang, J.L.; Tang, X.D.; Yang, K.X.; Li, X.R. The comparison of microbial communities in thyroid tissues from thyroid carcinoma patients. J. Microbiol. 2021, 59, 988–1001. [Google Scholar] [CrossRef] [PubMed]
- Kitahara, C.M.; Schneider, A.B. Epidemiology of Thyroid Cancer. Cancer Epidemiol. Biomark. Prev. 2022, 31, 1284–1297. [Google Scholar] [CrossRef] [PubMed]
- Pizzato, M.; Li, M.; Vignat, J.; Laversanne, M.; Singh, D.; La Vecchia, C.; Vaccarella, S. The epidemiological landscape of thyroid cancer worldwide: GLOBOCAN estimates for incidence and mortality rates in 2020. Lancet Diabetes Endocrinol. 2022, 10, 264–272. [Google Scholar] [CrossRef] [PubMed]
- Lim, H.; Devesa, S.S.; Sosa, J.A.; Check, D.; Kitahara, C.M. Trends in Thyroid Cancer Incidence and Mortality in the United States, 1974–2013. JAMA 2017, 317, 1338–1348. [Google Scholar] [CrossRef] [PubMed]
- Olson, E.; Wintheiser, G.; Wolfe, K.M.; Droessler, J.; Silberstein, P.T. Epidemiology of Thyroid Cancer: A Review of the National Cancer Database, 2000–2013. Cureus 2019, 11, e4127. [Google Scholar] [CrossRef]
- Kitahara, C.M.; Sosa, J.A.; Shiels, M.S. Influence of Nomenclature Changes on Trends in Papillary Thyroid Cancer Incidence in the United States, 2000 to 2017. J. Clin. Endocrinol. Metab. 2020, 105, e4823–e4830. [Google Scholar] [CrossRef]
- Chung, R.; Guan, H.; Ponchiardi, C.; Cerda, S.; Marwaha, N.; Yilmaz, O.H.; Pinjic, E.; McAneny, D.; Lee, S.L.; Drake, F.T. Noninvasive Follicular Thyroid Neoplasm with Papillary-Like Nuclear Features: Epidemiology and Long-Term Outcomes in a Strictly Defined Cohort. Thyroid 2021, 31, 68–75. [Google Scholar] [CrossRef]
- Cabanillas, M.E.; McFadden, D.G.; Durante, C. Thyroid cancer. Lancet 2016, 388, 2783–2795. [Google Scholar] [CrossRef]
- Zhao, H.; Liu, C.H.; Cao, Y.; Zhang, L.Y.; Zhao, Y.; Liu, Y.W.; Liu, H.F.; Lin, Y.S.; Li, X.Y. Survival prognostic factors for differentiated thyroid cancer patients with pulmonary metastases: A systematic review and meta-analysis. Front. Oncol. 2022, 12, 990154. [Google Scholar] [CrossRef]
- Fiore, M.; Cristaldi, A.; Okatyeva, V.; Lo Bianco, S.; Oliveri Conti, G.; Zuccarello, P.; Copat, C.; Caltabiano, R.; Cannizzaro, M.; Ferrante, M. Dietary habits and thyroid cancer risk: A hospital-based case-control study in Sicily (South Italy). Food Chem. Toxicol. 2020, 146, 111778. [Google Scholar] [CrossRef]
- Barrea, L.; Gallo, M.; Ruggeri, R.M.; Giacinto, P.D.; Sesti, F.; Prinzi, N.; Adinolfi, V.; Barucca, V.; Renzelli, V.; Muscogiuri, G.; et al. Nutritional status and follicular-derived thyroid cancer: An update. Crit. Rev. Food Sci. Nutr. 2021, 61, 25–59. [Google Scholar] [CrossRef] [PubMed]
- Dai, D.; Yang, Y.; Yang, Y.; Dang, T.; Xiao, J.; Wang, W.; Teng, L.; Xu, J.; Ye, J.; Jiang, H. Alterations of thyroid microbiota across different thyroid microhabitats in patients with thyroid carcinoma. J. Transl. Med. 2021, 19, 488. [Google Scholar] [CrossRef] [PubMed]
- Kun, Y.; Wei, X.; Wang, H.; Nie, X.; Dai, Q. Exploring the oral-gut microbiota during thyroid cancer: Factors affecting the thyroid functions and cancer development. Food Sci. Nutr. 2023, 11, 5657–5674. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Zhang, F.; Li, Q.; Feng, C.; Teng, W. Iodine nutrition and papillary thyroid cancer. Front. Nutr. 2022, 9, 1022650. [Google Scholar] [CrossRef]
- Macvanin, M.T.; Gluvic, Z.; Zafirovic, S.; Gao, X.; Essack, M.; Isenovic, E.R. The protective role of nutritional antioxidants against oxidative stress in thyroid disorders. Front. Endocrinol. 2023, 13, 1092837. [Google Scholar] [CrossRef]
- Heydarzadeh, S.; Kia, S.K.; Zarkesh, M.; Pakizehkar, S.; Hosseinzadeh, S.; Hedayati, M. The Cross-Talk between Polyphenols and the Target Enzymes Related to Oxidative Stress-Induced Thyroid Cancer. Oxid. Med. Cell. Longev. 2022, 2022, 2724324. [Google Scholar] [CrossRef]
- Muzza, M.; Pogliaghi, G.; Colombo, C.; Carbone, E.; Cirello, V.; Palazzo, S.; Frattini, F.; Gentilini, D.; Gazzano, G.; Persani, L.; et al. Oxidative Stress Correlates with More Aggressive Features in Thyroid Cancer. Cancers 2022, 14, 5857. [Google Scholar] [CrossRef]
- Hou, K.; Wu, Z.X.; Chen, X.Y.; Wang, J.Q.; Zhang, D.; Xiao, C.; Zhu, D.; Koya, J.B.; Wei, L.; Li, J.; et al. Microbiota in health and diseases. Signal Transduct. Target. Ther. 2022, 7, 135. [Google Scholar] [CrossRef]
- Riaz Rajoka, M.S.; Thirumdas, R.; Mehwish, H.M.; Umair, M.; Khurshid, M.; Hayat, H.F.; Phimolsiripol, Y.; Pallarés, N.; Martí-Quijal, F.J.; Barba, F.J. Role of Food Antioxidants in Modulating Gut Microbial Communities: Novel Understandings in Intestinal Oxidative Stress Damage and Their Impact on Host Health. Antioxidants 2021, 10, 1563. [Google Scholar] [CrossRef]
- Feng, J.; Zhao, F.; Sun, J.; Lin, B.; Zhao, L.; Liu, Y.; Jin, Y.; Li, S.; Li, A.; Wei, Y. Alterations in the gut microbiota and metabolite profiles of thyroid carcinoma patients. Int. J. Cancer 2019, 144, 2728–2745. [Google Scholar] [CrossRef]
- Liu, S.; An, Y.; Cao, B.; Sun, R.; Ke, J.; Zhao, D. The Composition of Gut Microbiota in Patients Bearing Hashimoto’s Thyroiditis with Euthyroidism and Hypothyroidism. Int. J. Endocrinol. 2020, 2020, 5036959. [Google Scholar] [CrossRef] [PubMed]
- Su, X.; Zhao, Y.; Li, Y.; Ma, S.; Wang, Z. Gut dysbiosis is associated with primary hypothyroidism with interaction on gut-thyroid axis. Clin. Sci. 2020, 134, 1521–1535. [Google Scholar] [CrossRef] [PubMed]
- Jiang, W.; Yu, X.; Kosik, R.O.; Song, Y.; Qiao, T.; Tong, J.; Liu, S.; Fan, S.; Luo, Q.; Chai, L.; et al. Gut Microbiota May Play a Significant Role in the Pathogenesis of Graves’ Disease. Thyroid 2021, 31, 810–820. [Google Scholar] [CrossRef]
- Liu, J.; Qin, X.; Lin, B.; Cui, J.; Liao, J.; Zhang, F.; Lin, Q. Analysis of gut microbiota diversity in Hashimoto’s thyroiditis patients. BMC Microbiol. 2022, 22, 318. [Google Scholar] [CrossRef] [PubMed]
- Medina-Reyes, E.I.; Rodríguez-Ibarra, C.; Déciga-Alcaraz, A.; Díaz-Urbina, D.; Chirino, Y.I.; Pedraza-Chaverri, J. Food additives containing nanoparticles induce gastrotoxicity, hepatotoxicity and alterations in animal behavior: The unknown role of oxidative stress. Food Chem. Toxicol. 2020, 146, 111814. [Google Scholar] [CrossRef]
- Naliyadhara, N.; Kumar, A.; Gangwar, S.K.; Devanarayanan, T.N.; Hegde, M.; Alqahtani, M.S.; Abbas, M.; Sethi, G.; Kunnumakara, A. Interplay of dietary antioxidants and gut microbiome in human health: What has been learnt thus far? J. Funct. Foods. 2023, 100, 105365. [Google Scholar] [CrossRef]
- Chen, Y.; Wu, F.H.; Wu, P.Q.; Xing, H.Y.; Ma, T. The Role of The Tumor Microbiome in Tumor Development and Its Treatment. Front. Immunol. 2022, 13, 935846. [Google Scholar] [CrossRef]
- Ciernikova, S.; Sevcikova, A.; Stevurkova, V.; Mego, M. Tumor microbiome—An integral part of the tumor microenvironment. Front. Oncol. 2022, 12, 1063100. [Google Scholar] [CrossRef]
- Stone, T.W.; Darlington, L.G. Microbial carcinogenic toxins and dietary anti-cancer protectants. Cell. Mol. Life Sci. 2017, 74, 2627–2643. [Google Scholar] [CrossRef]
- Sepich-Poore, G.D.; Zitvogel, L.; Straussman, R.; Hasty, J.; Wargo, J.A.; Knight, R. The microbiome and human cancer. Science 2021, 371, eabc4552. [Google Scholar] [CrossRef]
- Virili, C.; Centanni, M. “With a little help from my friends”—The role of microbiota in thyroid hormone metabolism and enterohepatic recycling. Mol. Cell Endocrinol. 2017, 458, 39–43. [Google Scholar] [CrossRef]
- Jiang, W.; Lu, G.; Gao, D.; Lv, Z.; Li, D. The relationships between the gut microbiota and its metabolites with thyroid diseases. Front. Endocrinol. 2022, 13, 943408. [Google Scholar] [CrossRef] [PubMed]
- Yoo, J.Y.; Groer, M.; Dutra, S.V.O.; Sarkar, A.; McSkimming, D.I. Gut Microbiota and Immune System Interactions. Microorganisms 2020, 8, 1587. [Google Scholar] [CrossRef] [PubMed]
- Liu, Q.; Sun, W.; Zhang, H. Interaction of Gut Microbiota with Endocrine Homeostasis and Thyroid Cancer. Cancers 2022, 14, 2656. [Google Scholar] [CrossRef] [PubMed]
- Knezevic, J.; Starchl, C.; Tmava Berisha, A.; Amrein, K. Thyroid-Gut-Axis: How Does the Microbiota Influence Thyroid Function? Nutrients 2020, 12, 1769. [Google Scholar] [CrossRef]
- Sirakov, M.; Plateroti, M. The thyroid hormones and their nuclear receptors in the gut: From developmental biology to cancer. Biochim. Biophys. Acta. 2011, 1812, 938–946. [Google Scholar] [CrossRef]
- Fenneman, A.C.; Bruinstroop, E.; Nieuwdorp, M.; van der Spek, A.H.; Boelen, A.A. Comprehensive Review of Thyroid Hormone Metabolism in the Gut and Its Clinical Implications. Thyroid 2023, 33, 32–44. [Google Scholar] [CrossRef]
- Nicola, J.P.; Nazar, M.; Mascanfroni, I.D.; Pellizas, C.G.; Masini-Repiso, A.M. NF-kappaB p65 subunit mediates lipopolysaccharide-induced Na(+)/I(-) symporter gene expression by involving functional interaction with the paired domain transcription factor Pax8. Mol. Endocrinol. 2010, 24, 1846–1862. [Google Scholar] [CrossRef]
- Puppin, C.; D’Aurizio, F.; D’Elia, A.V.; Cesaratto, L.; Tell, G.; Russo, D.; Filetti, S.; Ferretti, E.; Tosi, E.; Mattei, T.; et al. Effects of histone acetylation on sodium iodide symporter promoter and expression of thyroid-specific transcription factors. Endocrinology 2005, 146, 3967–3974. [Google Scholar] [CrossRef]
- Zhou, L.; Zhang, M.; Wang, Y.; Dorfman, R.G.; Liu, H.; Yu, T.; Chen, X.; Tang, D.; Xu, L.; Yin, Y.; et al. Faecalibacterium prausnitzii Produces Butyrate to Maintain Th17/Treg Balance and to Ameliorate Colorectal Colitis by Inhibiting Histone Deacetylase 1. Inflamm. Bowel Dis. 2018, 24, 1926–1940. [Google Scholar] [CrossRef]
- Luongo, C.; Trivisano, L.; Alfano, F.; Salvatore, D. Type 3 deiodinase and consumptive hypothyroidism: A common mechanism for a rare disease. Front. Endocrinol. 2013, 4, 115. [Google Scholar] [CrossRef] [PubMed]
- Fröhlich, E.; Wahl, R. Microbiota and thyroid interaction in health and disease. Trends Endocrinol. Metab. 2019, 30, 479–490. [Google Scholar] [CrossRef] [PubMed]
- Boelen, A.; Kwakkel, J.; Alkemade, A.; Renckens, R.; Kaptein, E.; Kuiper, G.; Wiersinga, W.M.; Visser, T.J. Induction of type 3 deiodinase activity in inflammatory cells of mice with chronic local inflammation. Endocrinology 2005, 146, 5128–5134. [Google Scholar] [CrossRef]
- Fekete, C.; Gereben, B.; Doleschall, M.; Harney, J.W.; Dora, J.M.; Bianco, A.C.; Sarkar, S.; Liposits, Z.; Rand, W.; Emerson, C.; et al. Lipopolysaccharide induces type 2 iodothyronine deiodinase in the mediobasal hypothalamus: Implications for the nonthyroidal illness syndrome. Endocrinology 2004, 145, 1649–1655. [Google Scholar] [CrossRef] [PubMed]
- Asarat, M.; Apostolopoulos, V.; Vasiljevic, T.; Donkor, O. Short-Chain Fatty Acids Regulate Cytokines and Th17/Treg Cells in Human Peripheral Blood Mononuclear Cells in vitro. Immunol. Invest. 2016, 45, 205–222. [Google Scholar] [CrossRef]
- Li, J.; Sung, C.Y.; Lee, N.; Ni, Y.; Pihlajamaki, J.; Panagiotou, G.; El-Nezami, H. Probiotics modulated gut microbiota suppresses hepatocellular carcinoma growth in mice. Proc. Natl. Acad. Sci. USA 2016, 113, E1306–E1315. [Google Scholar] [CrossRef]
- Hamamah, S.; Aghazarian, A.; Nazaryan, A.; Hajnal, A.; Covasa, M. Role of Microbiota-Gut-Brain Axis in Regulating Dopaminergic Signaling. Biomedicines 2022, 10, 436. [Google Scholar] [CrossRef]
- Daubner, S.C.; Le, T.; Wang, S. Tyrosine hydroxylase and regulation of dopamine synthesis. Arch Biochem Biophys. 2011, 508, 1–12. [Google Scholar] [CrossRef]
- Azevedo, M.M.; Pina-Vaz, C.; Baltazar, F. Microbes and Cancer: Friends or Faux? Int. J. Mol. Sci. 2020, 21, 3115. [Google Scholar] [CrossRef]
- Veziant, J.; Villéger, R.; Barnich, N.; Bonnet, M. Gut Microbiota as Potential Biomarker and/or Therapeutic Target to Improve the Management of Cancer: Focus on Colibactin-Producing Escherichia coli in Colorectal Cancer. Cancers 2021, 13, 2215. [Google Scholar] [CrossRef]
- Lucas, C.; Barnich, N.; Nguyen, H.T.T. Microbiota, Inflammation and Colorectal Cancer. Int. J. Mol. Sci. 2017, 18, 1310. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Yang, Y.; Huycke, M.M. Microbiome-driven carcinogenesis in colorectal cancer: Models and mechanisms. Free Radic. Biol. Med. 2017, 105, 3–15. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Yu, K.C.; Hou, Y.Q.; Guo, M.; Yao, F.; Chen, Z.X. Gut microbiome in tumorigenesis and therapy of colorectal cancer. J. Cell. Physiol. 2023, 238, 94–108. [Google Scholar] [CrossRef] [PubMed]
- Yasunaga, J.I.; Matsuoka, M. Oncogenic spiral by infectious pathogens: Cooperation of multiple factors in cancer development. Cancer Sci. 2018, 109, 24–32. [Google Scholar] [CrossRef] [PubMed]
- IARC Working Group on the Evaluation of Carcinogenic Risks to Humans. Biological agents. IARC Monogr. Eval. Carcinog. Risks Hum. 2012, 100, 1–441. [Google Scholar]
- Locey, K.J.; Lennon, J.T. Scaling laws predict global microbial diversity. Proc. Natl. Acad. Sci. USA 2016, 113, 5970–5975. [Google Scholar] [CrossRef]
- Gopalakrishnan, V.; Helmink, B.A.; Spencer, C.N.; Reuben, A.; Wargo, J.A. The Influence of the Gut Microbiome on Cancer, Immunity, and Cancer Immunotherapy. Cancer Cell 2018, 33, 570–580. [Google Scholar] [CrossRef]
- Ni, J.; Huang, R.; Zhou, H.; Xu, X.; Li, Y.; Cao, P.; Zhong, K.; Ge, M.; Chen, X.; Hou, B.; et al. Analysis of the Relationship Between the Degree of Dysbiosis in Gut Microbiota and Prognosis at Different Stages of Primary Hepatocellular Carcinoma. Front. Microbiol. 2019, 10, 1458. [Google Scholar] [CrossRef]
- Zhang, J.; Zhang, F.; Zhao, C.; Xu, Q.; Liang, C.; Yang, Y.; Wang, H.; Shang, Y.; Wang, Y.; Mu, X.; et al. Dysbiosis of the gut microbiome is associated with thyroid cancer and thyroid nodules and correlated with clinical index of thyroid function. Endocrine 2019, 64, 564–574. [Google Scholar] [CrossRef]
- Gantuya, B.; El-Serag, H.B.; Matsumoto, T.; Ajami, N.J.; Oyuntsetseg, K.; Azzaya, D.; Uchida, T.; Yamaoka, Y. Gastric Microbiota in Helicobacter pylori-Negative and -Positive Gastritis Among High Incidence of Gastric Cancer Area. Cancers 2019, 11, 504. [Google Scholar] [CrossRef]
- Sohn, S.H.; Kim, N.; Jo, H.J.; Kim, J.; Park, J.H.; Nam, R.H.; Seok, Y.J.; Kim, Y.R.; Lee, D.H. Analysis of Gastric Body Microbiota by Pyrosequencing: Possible Role of Bacteria Other Than Helicobacter pylori in the Gastric Carcinogenesis. J. Cancer Prev. 2017, 22, 115–125. [Google Scholar] [CrossRef] [PubMed]
- Kumar, R.; Herold, J.L.; Schady, D.; Davis, J.; Kopetz, S.; Martinez-Moczygemba, M.; Murray, B.E.; Han, F.; Li, Y.; Callaway, E.; et al. Streptococcus gallolyticus subsp. gallolyticus promotes colorectal tumor development. PLoS Pathog. 2017, 13, e1006440. [Google Scholar] [CrossRef] [PubMed]
- Zhao, J.; Nian, L.; Kwok, L.Y.; Sun, T.; Zhao, J. Reduction in fecal microbiota diversity and short-chain fatty acid producers in Methicillin-resistant Staphylococcus aureus infected individuals as revealed by PacBio single molecule, real-time sequencing technology. Eur. J. Clin. Microbiol. Infect. Dis. 2017, 36, 1463–1472. [Google Scholar] [CrossRef] [PubMed]
- Silva, Y.P.; Bernardi, A.; Frozza, R.L. The Role of Short-Chain Fatty Acids From Gut Microbiota in Gut-Brain Communication. Front. Endocrinol. 2020, 11, 25. [Google Scholar] [CrossRef]
- Portincasa, P.; Bonfrate, L.; Vacca, M.; De Angelis, M.; Farella, I.; Lanza, E.; Khalil, M.; Wang, D.Q.; Sperandio, M.; Di Ciaula, A. Gut Microbiota and Short Chain Fatty Acids: Implications in Glucose Homeostasis. Int. J. Mol. Sci. 2022, 23, 1105. [Google Scholar] [CrossRef]
- Liu, X.F.; Shao, J.H.; Liao, Y.T.; Wang, L.N.; Jia, Y.; Dong, P.J.; Liu, Z.Z.; He, D.D.; Li, C.; Zhang, X. Regulation of short-chain fatty acids in the immune system. Front. Immunol. 2023, 14, 1186892. [Google Scholar] [CrossRef]
- Pessione, E. Lactic acid bacteria contribution to gut microbiota complexity: Lights and shadows. Front. Cell. Infect. Microbiol. 2012, 2, 86. [Google Scholar] [CrossRef]
- Zoidis, E.; Seremelis, I.; Kontopoulos, N.; Danezis, G.P. Selenium-Dependent Antioxidant Enzymes: Actions and Properties of Selenoproteins. Antioxidants 2018, 7, 66. [Google Scholar] [CrossRef]
- Gorini, F.; Sabatino, L.; Pingitore, A.; Vassalle, C. Selenium: An Element of Life Essential for Thyroid Function. Molecules 2021, 26, 7084. [Google Scholar] [CrossRef]
- Li, J.; Xie, H.; Li, A.; Cheng, J.; Yang, K.; Wang, J.; Wang, W.; Zhang, F.; Li, Z.; Dhillon, H.S.; et al. Distinct plasma lipids profiles of recurrent ovarian cancer by liquid chromatography-mass spectrometry. Oncotarget 2017, 8, 46834–46845. [Google Scholar] [CrossRef]
- Shen, S.; Yang, L.; Li, L.; Bai, Y.; Cai, C.; Liu, H. A plasma lipidomics strategy reveals perturbed lipid metabolic pathways and potential lipid biomarkers of human colorectal cancer. J. Chromatogr. B Analyt. Technol. Biomed. Life Sci. 2017, 1068–1069, 41–48. [Google Scholar] [CrossRef] [PubMed]
- Fu, Y.; Zou, T.; Shen, X.; Nelson, P.J.; Li, J.; Wu, C.; Yang, J.; Zheng, Y.; Bruns, C.; Zhao, Y.; et al. Lipid metabolism in cancer progression and therapeutic strategies. MedComm 2020, 2, 27–59. [Google Scholar] [CrossRef]
- Simsir, Y.I.; Cetinkalp, S.; Kabalak, T. Review of Factors Contributing to Nodular Goiter and Thyroid Carcinoma. Med. Princ. Pract. 2020, 29, 1–5. [Google Scholar]
- Azuma, H.; Inamoto, T.; Sakamoto, T.; Kiyama, S.; Ubai, T.; Shinohara, Y.; Maemura, K.; Tsuji, M.; Segawa, N.; Masuda, H.; et al. Gamma-aminobutyric acid as a promoting factor of cancer metastasis; induction of matrix metalloproteinase production is potentially its underlying mechanism. Cancer Res. 2003, 63, 8090–8096. [Google Scholar] [PubMed]
- Roberts, S.S.; Mendonça-Torres, M.C.; Jensen, K.; Francis, G.L.; Vasko, V. GABA receptor expression in benign and malignant thyroid tumors. Pathol. Oncol. Res. 2009, 15, 645–650. [Google Scholar] [CrossRef] [PubMed]
- Huang, D.; Wang, Y.; Thompson, J.W.; Yin, T.; Alexander, P.B.; Qin, D.; Mudgal, P.; Wu, H.; Liang, Y.; Tan, L.; et al. Cancer-cell-derived GABA promotes β-catenin-mediated tumour growth and immunosuppression. Nat. Cell Biol. 2022, 24, 230–241. [Google Scholar] [CrossRef]
- Lau, A.; Tymianski, M. Excitotoxicity. In Comprehensive Toxicology, 2nd ed.; Elsevier: McQueen, CA, USA, 2010; Volume 13, pp. 515–535. [Google Scholar]
- Qian, B.; Shen, S.; Zhang, J.; Jing, P. Effects of Vitamin B6 Deficiency on the Composition and Functional Potential of T Cell Populations. J. Immunol. Res. 2017, 2017, 2197975. [Google Scholar] [CrossRef]
- Yu, X.; Jiang, W.; Kosik, R.O.; Song, Y.; Luo, Q.; Qiao, T.; Tong, J.; Liu, S.; Deng, C.; Qin, S.; et al. Gut microbiota changes and its potential relations with thyroid carcinoma. J. Adv. Res. 2021, 35, 61–70. [Google Scholar] [CrossRef]
- Stojanov, S.; Berlec, A.; Štrukelj, B. The Influence of Probiotics on the Firmicutes/Bacteroidetes Ratio in the Treatment of Obesity and Inflammatory Bowel disease. Microorganisms 2020, 8, 1715. [Google Scholar] [CrossRef]
- Xu, Z.; Jiang, W.; Huang, W.; Lin, Y.; Chan, F.K.L.; Ng, S.C. Gut microbiota in patients with obesity and metabolic disorders—A systematic review. Genes Nutr. 2022, 17, 2. [Google Scholar] [CrossRef]
- Yuan, L.; Yang, P.; Wei, G.; Hu, X.; Chen, S.; Lu, J.; Yang, L.; He, X.; Bao, G. Tumor microbiome diversity influences papillary thyroid cancer invasion. Commun. Biol. 2022, 5, 864. [Google Scholar] [CrossRef] [PubMed]
- Xue, C.; Chu, Q.; Zheng, Q.; Yuan, X.; Su, Y.; Bao, Z.; Lu, J.; Li, L. Current understanding of the intratumoral microbiome in various tumors. Cell Rep. Med. 2023, 4, 100884. [Google Scholar] [CrossRef] [PubMed]
- Nejman, D.; Livyatan, I.; Fuks, G.; Gavert, N.; Zwang, Y.; Geller, L.T.; Rotter-Maskowitz, A.; Weiser, R.; Mallel, G.; Gigi, E.; et al. The human tumor microbiome is composed of tumor type-specific intracellular bacteria. Science 2020, 368, 973–980. [Google Scholar] [CrossRef] [PubMed]
- Baghban, R.; Roshangar, L.; Jahanban-Esfahlan, R.; Seidi, K.; Ebrahimi-Kalan, A.; Jaymand, M.; Kolahian, S.; Javaheri, T.; Zare, P. Tumor microenvironment complexity and therapeutic implications at a glance. Cell Commun. Signal. 2020, 18, 59. [Google Scholar] [CrossRef]
- Rossi, T.; Vergara, D.; Fanini, F.; Maffia, M.; Bravaccini, S.; Pirini, F. Microbiota-Derived Metabolites in Tumor Progression and Metastasis. Int. J. Mol. Sci. 2020, 21, 5786. [Google Scholar] [CrossRef]
- Scales, B.S.; Dickson, R.P.; LiPuma, J.J.; Huffnagle, G.B. Microbiology, genomics, and clinical significance of the Pseudomonas fluorescens species complex, an unappreciated colonizer of humans. Clin. Microbiol. Rev. 2014, 27, 927–948. [Google Scholar] [CrossRef]
- Reynolds, D.; Kollef, M. The Epidemiology and Pathogenesis and Treatment of Pseudomonas aeruginosa Infections: An Update. Drugs 2021, 81, 2117–2131. [Google Scholar] [CrossRef]
- Taglialegna, A. Pseudomonas against cancer. Nat. Rev. Microbiol. 2023, 21, 131. [Google Scholar] [CrossRef]
- Chaudhary, P.P.; Conway, P.L.; Schlundt, J. Methanogens in humans: Potentially beneficial or harmful for health. Appl. Microbiol. Biotechnol. 2018, 102, 3095–3104. [Google Scholar] [CrossRef]
- Liu, Y.; O’Brien, J.L.; Ajami, N.J.; Scheurer, M.E.; Amirian, E.S.; Armstrong, G.; Tsavachidis, S.; Thrift, A.P.; Jiao, L.; Wong, M.C.; et al. Lung tissue microbial profile in lung cancer is distinct from emphysema. Am. J. Cancer Res. 2018, 8, 1775–1787. [Google Scholar]
- Liu, X.; Shao, L.; Liu, X.; Ji, F.; Mei, Y.; Cheng, Y.; Liu, F.; Yan, C.; Li, L.; Ling, Z. Alterations of gastric mucosal microbiota across different stomach microhabitats in a cohort of 276 patients with gastric cancer. EBioMedicine 2019, 40, 336–348. [Google Scholar] [CrossRef] [PubMed]
- Higuchi, R.; Goto, T.; Hirotsu, Y.; Otake, S.; Oyama, T.; Amemiya, K.; Ohyama, H.; Mochizuki, H.; Omata, M. Sphingomonas and Phenylobacterium as Major Microbiota in Thymic Epithelial Tumors. J. Pers. Med. 2021, 11, 1092. [Google Scholar] [CrossRef]
- Richard, M.L.; Liguori, G.; Lamas, B.; Brandi, G.; da Costa, G.; Hoffmann, T.W.; Di Simone, P.M.; Calabrese, C.; Poggioli, G.; Langella, P.; et al. Mucosa-associated microbiota dysbiosis in colitis associated cancer. Gut Microbes 2018, 9, 131–142. [Google Scholar] [CrossRef] [PubMed]
- Tanaka, T.; Matsuno, Y.; Torisu, T.; Shibata, H.; Hirano, A.; Umeno, J.; Kawasaki, K.; Fujioka, S.; Fuyuno, Y.; Moriyama, T.; et al. Gastric microbiota in patients with Helicobacter pylori-negative gastric MALT lymphoma. Medicine 2021, 100, e27287. [Google Scholar] [CrossRef] [PubMed]
- Jeong, J.Y.; Kim, T.B.; Kim, J.; Choi, H.W.; Kim, E.J.; Yoo, H.J.; Lee, S.; Jun, H.R.; Yoo, W.; Kim, S.; et al. Diversity in the Extracellular Vesicle-Derived Microbiome of Tissues According to Tumor Progression in Pancreatic Cancer. Cancers 2020, 12, 2346. [Google Scholar] [CrossRef] [PubMed]
- Gnanasekar, A.; Castaneda, G.; Iyangar, A.; Magesh, S.; Perez, D.; Chakladar, J.; Li, W.T.; Bouvet, M.; Chang, E.Y.; Ongkeko, W.M. The intratumor microbiome predicts prognosis across gender and subtypes in papillary thyroid carcinoma. Comput. Struct. Biotechnol. J. 2021, 19, 1986–1997. [Google Scholar] [CrossRef]
- Ianniello, N.M.; Andrade, D.C.; Ivancic, S.; Eckardt, P.A.; Lemos Ramirez, J.C. Native valve infective endocarditis due to Micrococcus luteus in a non-Hodgkin’s lymphoma patient. IDCases 2019, 18, e00657. [Google Scholar] [CrossRef]
- Martín Guerra, J.M.; Martín Asenjo, M.; Rodríguez Martín, C. Bacteraemia by Micrococcus luteus in an inmunocompromised patient. Med. Clin. 2019, 152, 469–470. [Google Scholar] [CrossRef]
- Quach, J.U.; Diaz, M.J.; Huda, T.I.; Kinskey, J.C.; Zaman, S.; Desantis, J.E.; Cios, K.J.; Blanck, G. Bacterial Sequencing Reads in Blood Exome Files from Melanoma and Cervical Cancer Patients are Associated with Cancer Recurrence. Mol. Biotechnol. 2023, 65, 1476–1484. [Google Scholar] [CrossRef]
- Ding, J.; Wu, W.; Fang, J.; Zhao, J.; Jiang, L. Male sex is associated with aggressive behaviour and poor prognosis in Chinese papillary thyroid carcinoma. Sci. Rep. 2020, 10, 4141. [Google Scholar] [CrossRef]
- Kiseleva, E.P.; Mikhailopulo, K.I.; Sviridov, O.V.; Novik, G.I.; Knirel, Y.A.; Szwajcer Dey, E. The role of components of Bifidobacterium and Lactobacillus in pathogenesis and serologic diagnosis of autoimmune thyroid diseases. Benef. Microbes 2011, 2, 139–154. [Google Scholar] [CrossRef] [PubMed]
- Marchi, S.; Giorgi, C.; Suski, J.M.; Agnoletto, C.; Bononi, A.; Bonora, M.; De Marchi, E.; Missiroli, S.; Patergnani, S.; Poletti, F.; et al. Mitochondria-ros crosstalk in the control of cell death and aging. J. Signal Transduct. 2012, 2012, 329635. [Google Scholar] [CrossRef]
- Metere, A.; Frezzotti, F.; Graves, C.E.; Vergine, M.; De Luca, A.; Pietraforte, D.; Giacomelli, L. A possible role for selenoprotein glutathione peroxidase (GPx1) and thioredoxin reductases (TrxR1) in thyroid cancer: Our experience in thyroid surgery. Cancer Cell Int. 2018, 18, 7. [Google Scholar] [CrossRef]
- Ameziane-El-Hassani, R.; Buffet, C.; Leboulleux, S.; Dupuy, C. Oxidative stress in thyroid carcinomas: Biological and clinical significance. Endocr. Relat. Cancer 2019, 26, R131–R143. [Google Scholar] [CrossRef]
- Morshed, S.A.; Davies, T.F. Understanding Thyroid Cell Stress. J. Clin. Endocrinol. Metab. 2020, 105, e66–e69. [Google Scholar] [CrossRef]
- Karger, S.; Krause, K.; Engelhardt, C.; Weidinger, C.; Gimm, O.; Dralle, H.; Sheu-Grabellus, S.Y.; Schmid, K.W.; Fuhrer, D. Distinct pattern of oxidative DNA damage and DNA repair in follicular thyroid tumours. J. Mol. Endocrinol. 2012, 48, 193–202. [Google Scholar] [CrossRef]
- Qing, X.; Shi, D.; Lv, X.; Wang, B.; Chen, S.; Shao, Z. Prognostic significance of 8-hydroxy-2′-deoxyguanosine in solid tumors: A meta-analysis. BMC Cancer 2019, 19, 997. [Google Scholar] [CrossRef]
- Xing, M. Oxidative stress: A new risk factor for thyroid cancer. Endocr. Relat. Cancer 2012, 19, C7–C11. [Google Scholar] [CrossRef]
- Kościuszko, M.; Buczyńska, A.; Krętowski, A.J.; Popławska-Kita, A. Could Oxidative Stress Play a Role in the Development and Clinical Management of Differentiated Thyroid Cancer? Cancers 2023, 15, 3182. [Google Scholar] [CrossRef]
- Massart, C.; Hoste, C.; Virion, A.; Ruf, J.; Dumont, J.E.; Van Sande, J. Cell biology of H2O2 generation in the thyroid: Investigation of the control of dual oxidases (DUOX) activity in intact ex vivo thyroid tissue and cell lines. Mol. Cell Endocrinol. 2011, 343, 32–44. [Google Scholar] [CrossRef]
- Ameziane-El-Hassani, R.; Schlumberger, M.; Dupuy, C. NADPH oxidases: New actors in thyroid cancer? Nat. Rev. Endocrinol. 2016, 12, 485–894. [Google Scholar] [CrossRef]
- Szanto, I.; Pusztaszeri, M.; Mavromati, M. H2O2 Metabolism in Normal Thyroid Cells and in Thyroid Tumorigenesis: Focus on NADPH Oxidases. Antioxidants 2019, 8, 126. [Google Scholar] [CrossRef]
- Song, Y.; Driessens, N.; Costa, M.; De Deken, X.; Detours, V.; Corvilain, B.; Maenhaut, C.; Miot, F.; Van Sande, J.; Many, M.C.; et al. Roles of hydrogen peroxide in thyroid physiology and disease. J. Clin. Endocrinol. Metab. 2007, 92, 3764–3773. [Google Scholar] [CrossRef]
- Schweizer, U.; Chiu, J.; Köhrle, J. Peroxides and peroxide-degrading enzymes in the thyroid. Antioxid. Redox Signal. 2008, 10, 1577–1592. [Google Scholar] [CrossRef] [PubMed]
- Brandes, R.P.; Weissmann, N.; Schröder, K. Nox family NADPH oxidases: Molecular mechanisms of activation. Free Radic. Biol. Med. 2014, 76, 208–226. [Google Scholar] [CrossRef] [PubMed]
- Ameziane-El-Hassani, R.; Talbot, M.; de Souza Dos Santos, M.C.; Al Ghuzlan, A.; Hartl, D.; Bidart, J.M.; De Deken, X.; Miot, F.; Diallo, I.; de Vathaire, F.; et al. NADPH oxidase DUOX1 promotes long-term persistence of oxidative stress after an exposure to irradiation. Proc. Natl. Acad. Sci. USA 2015, 112, 5051–5056. [Google Scholar] [CrossRef] [PubMed]
- Ameziane-El-Hassani, R.; Boufraqech, M.; Lagente-Chevallier, O.; Weyemi, U.; Talbot, M.; Métivier, D.; Courtin, F.; Bidart, J.M.; El Mzibri, M.; Schlumberger, M.; et al. Role of H2O2 in RET/PTC1 chromosomal rearrangement produced by ionizing radiation in human thyroid cells. Cancer Res. 2010, 70, 4123–4132. [Google Scholar] [CrossRef] [PubMed]
- Detours, V.; Delys, L.; Libert, F.; Weiss Solís, D.; Bogdanova, T.; Dumont, J.E.; Franc, B.; Thomas, G.; Maenhaut, C. Genome-wide gene expression profiling suggests distinct radiation susceptibilities in sporadic and post-Chernobyl papillary thyroid cancers. Br. J. Cancer 2007, 97, 818–825. [Google Scholar] [CrossRef] [PubMed]
- Dom, G.; Tarabichi, M.; Unger, K.; Thomas, G.; Oczko-Wojciechowska, M.; Bogdanova, T.; Jarzab, B.; Dumont, J.E.; Detours, V.; Maenhaut, C. A gene expression signature distinguishes normal tissues of sporadic and radiation-induced papillary thyroid carcinomas. Br. J. Cancer. 2012, 107, 994–1000. [Google Scholar] [CrossRef] [PubMed]
- Muzza, M.; Colombo, C.; Cirello, V.; Perrino, M.; Vicentini, L.; Fugazzola, L. Oxidative stress and the subcellular localization of the telomerase reverse transcriptase (TERT) in papillary thyroid cancer. Mol. Cell. Endocrinol. 2016, 431, 54–61. [Google Scholar] [CrossRef]
- Weyemi, U.; Caillou, B.; Talbot, M.; Ameziane-El-Hassani, R.; Lacroix, L.; Lagent-Chevallier, O.; Al Ghuzlan, A.; Roos, D.; Bidart, J.M.; Virion, A.; et al. Intracellular expression of reactive oxygen species-generating NADPH oxidase NOX4 in normal and cancer thyroid tissues. Endocr. Relat. Cancer 2010, 17, 27–37. [Google Scholar] [CrossRef]
- Azouzi, N.; Cailloux, J.; Cazarin, J.M.; Knauf, J.A.; Cracchiolo, J.; Al Ghuzlan, A.; Hartl, D.; Polak, M.; Carré, A.; El Mzibri, M.; et al. NADPH Oxidase NOX4 Is a Critical Mediator of BRAFV600E-Induced Downregulation of the Sodium/Iodide Symporter in Papillary Thyroid Carcinomas. Antioxid. Redox Signal. 2017, 26, 864–877. [Google Scholar] [CrossRef] [PubMed]
- Xing, M.; Alzahrani, A.S.; Carson, K.A.; Viola, D.; Elisei, R.; Bendlova, B.; Yip, L.; Mian, C.; Vianello, F.; Tuttle, R.M.; et al. Association between BRAF V600E mutation and mortality in patients with papillary thyroid cancer. JAMA 2013, 309, 1493–1501. [Google Scholar] [CrossRef]
- Cazarin, J.; Dupuy, C.; Pires de Carvalho, D. Redox Homeostasis in Thyroid Cancer: Implications in Na+/I- Symporter (NIS) Regulation. Int. J. Mol. Sci. 2022, 23, 6129. [Google Scholar] [CrossRef] [PubMed]
- Choi, Y.W.; Kim, H.J.; Kim, Y.H.; Park, S.H.; Chwae, Y.J.; Lee, J.; Soh, E.Y.; Kim, J.H.; Park, T.J. B-RafV600E inhibits sodium iodide symporter expression via regulation of DNA methyltransferase 1. Exp. Mol. Med. 2014, 46, e120. [Google Scholar] [CrossRef]
- Tang, P.; Dang, H.; Huang, J.; Xu, T.; Yuan, P.; Hu, J.; Sheng, J.F. NADPH oxidase NOX4 is a glycolytic regulator through mROS-HIF1α axis in thyroid carcinomas. Sci. Rep. 2018, 8, 15897. [Google Scholar] [CrossRef] [PubMed]
- Liberti, M.V.; Locasale, J.W. The Warburg Effect: How Does it Benefit Cancer Cells? Trends Biochem. Sci. 2016, 41, 211–218, Erratum in: Trends Biochem. Sci. 2016, 41, 287. [Google Scholar] [CrossRef]
- Ryder, M.; Ghossein, R.A.; Ricarte-Filho, J.C.; Knauf, J.A.; Fagin, J.A. Increased density of tumor-associated macrophages is associated with decreased survival in advanced thyroid cancer. Endocr. Relat. Cancer 2008, 15, 1069–1074. [Google Scholar] [CrossRef]
- Zhang, J.; Li, H.; Wu, Q.; Chen, Y.; Deng, Y.; Yang, Z.; Zhang, L.; Liu, B. Tumoral NOX4 recruits M2 tumor-associated macrophages via ROS/PI3K signaling-dependent various cytokine production to promote NSCLC growth. Redox Biol. 2019, 22, 101116. [Google Scholar] [CrossRef]
- Cancer Genome Atlas Research Network. Integrated genomic characterization of papillary thyroid carcinoma. Cell 2014, 159, 676–690. [Google Scholar] [CrossRef]
- Schubert, L.; Mariko, M.L.; Clerc, J.; Huillard, O.; Groussin, L. MAPK Pathway Inhibitors in Thyroid Cancer: Preclinical and Clinical Data. Cancers 2023, 15, 710. [Google Scholar] [CrossRef] [PubMed]
- Schwabe, R.F.; Jobin, C. The microbiome and cancer. Nat. Rev. Cancer 2013, 13, 800–812. [Google Scholar] [CrossRef] [PubMed]
- Garrett, W.S. Cancer and the microbiota. Science 2015, 348, 80–86. [Google Scholar] [CrossRef]
- Docimo, G.; Cangiano, A.; Romano, R.M.; Pignatelli, M.F.; Offi, C.; Paglionico, V.A.; Galdiero, M.; Donnarumma, G.; Nigro, V.; Esposito, D.; et al. The Human Microbiota in Endocrinology: Implications for Pathophysiology, Treatment, and Prognosis in Thyroid Diseases. Front. Endocrinol. 2020, 11, 586529. [Google Scholar] [CrossRef]
- Xia, Y.; Shen, S.; Verma, I.M. NF-κB, an active player in human cancers. Cancer Immunol. Res. 2014, 2, 823–830. [Google Scholar] [CrossRef]
- Grivennikov, S.I.; Wang, K.; Mucida, D.; Stewart, C.A.; Schnabl, B.; Jauch, D.; Taniguchi, K.; Yu, G.Y.; Osterreicher, C.H.; Hung, K.E.; et al. Adenoma-linked barrier defects and microbial products drive IL-23/IL-17-mediated tumour growth. Nature 2012, 491, 254–258. [Google Scholar] [CrossRef] [PubMed]
- Grivennikov, S.I.; Karin, M. Inflammatory cytokines in cancer: Tumour necrosis factor and interleukin 6 take the stage. Ann. Rheum. Dis. 2011, 70, i104–i108. [Google Scholar] [CrossRef]
- Grivennikov, S.I. IL-11: A prominent pro-tumorigenic member of the IL-6 family. Cancer Cell 2013, 24, 145–147. [Google Scholar] [CrossRef]
- Yu, H.; Lee, H.; Herrmann, A.; Buettner, R.; Jove, R. Revisiting STAT3 signalling in cancer: New and unexpected biological functions. Nat. Rev. Cancer 2014, 14, 736–746. [Google Scholar] [CrossRef]
- Louis, P.; Hold, G.L.; Flint, H.J. The gut microbiota, bacterial metabolites and colorectal cancer. Nat. Rev. Microbiol. 2014, 12, 661–672. [Google Scholar] [CrossRef]
- Ridlon, J.M.; Kang, D.J.; Hylemon, P.B.; Bajaj, J.S. Bile acids and the gut microbiome. Curr. Opin. Gastroenterol. 2014, 30, 332–338. [Google Scholar] [CrossRef]
- Glick, D.; Barth, S.; Macleod, K.F. Autophagy: Cellular and molecular mechanisms. J. Pathol. 2010, 221, 3–12. [Google Scholar] [CrossRef]
- Wang, Y.; Du, J.; Wu, X.; Abdelrehem, A.; Ren, Y.; Liu, C.; Zhou, X.; Wang, S. Crosstalk between autophagy and microbiota in cancer progression. Mol. Cancer 2021, 20, 163. [Google Scholar] [CrossRef]
- Thorburn, A. Apoptosis and autophagy: Regulatory connections between two supposedly different processes. Apoptosis 2008, 13, 1–9. [Google Scholar] [CrossRef]
- Deretic, V. Autophagy in inflammation, infection, and immunometabolism. Immunity 2021, 54, 437–453. [Google Scholar] [CrossRef]
- Jiang, B.; Zhou, X.; Yang, T.; Wang, L.; Feng, L.; Wang, Z.; Xu, J.; Jing, W.; Wang, T.; Su, H.; et al. The role of autophagy in cardiovascular disease: Cross-interference of signaling pathways and underlying therapeutic targets. Front. Cardiovasc. Med. 2023, 10, 1088575. [Google Scholar] [CrossRef]
- Guo, F.; Liu, X.; Cai, H.; Le, W. Autophagy in neurodegenerative diseases: Pathogenesis and therapy. Brain Pathol. 2018, 28, 3–13. [Google Scholar] [CrossRef]
- Jin, M.; Zhang, Y. Autophagy and Autoimmune Diseases. Adv. Exp. Med. Biol. 2020, 1207, 405–408. [Google Scholar]
- Mulcahy Levy, J.M.; Thorburn, A. Autophagy in cancer: Moving from understanding mechanism to improving therapy responses in patients. Cell Death Differ. 2020, 27, 843–857. [Google Scholar] [CrossRef]
- Zhong, X.; Chen, O.; Zhou, T.; Lü, M.; Wan, J. Cytotoxin-Associated Gene A-Positive Helicobacter pylori Promotes Autophagy in Colon Cancer Cells by Inhibiting miR-125b-5p. Can. J. Infect. Dis. Med. Microbiol. 2021, 2021, 6622092. [Google Scholar] [CrossRef]
- Binder Gallimidi, A.; Fischman, S.; Revach, B.; Bulvik, R.; Maliutina, A.; Rubinstein, A.M.; Nussbaum, G.; Elkin, M. Periodontal pathogens Porphyromonas gingivalis and Fusobacterium nucleatum promote tumor progression in an oral-specific chemical carcinogenesis model. Oncotarget 2015, 6, 22613–22623. [Google Scholar] [CrossRef] [PubMed]
- Hu, F.; Song, D.; Yan, Y.; Huang, C.; Shen, C.; Lan, J.; Chen, Y.; Liu, A.; Wu, Q.; Sun, L.; et al. IL-6 regulates autophagy and chemotherapy resistance by promoting BECN1 phosphorylation. Nat. Commun. 2021, 12, 3651. [Google Scholar] [CrossRef] [PubMed]
- Cho, T.J.; Wee, S.W.; Woo, V.H.; Choi, J.I.; Kim, S.J.; Shin, H.I.; Lee, J.H.; Park, H.R. Porphyromonas gingivalis-induced autophagy suppresses cell proliferation through G1 arrest in oral cancer cells. Arch. Oral. Biol. 2014, 59, 370–378. [Google Scholar] [CrossRef] [PubMed]
- Liu, W.D.; Sun, W.; Hua, Y.Q.; Wang, S.G.; Cai, Z.D. Effect of rapamycin and chloroquine on osteosarcoma. Zhonghua Yi Xue Za Zhi 2017, 97, 1510–1514. (In Chinese) [Google Scholar]
- Plottel, C.S.; Blaser, M.J. Microbiome and malignancy. Cell Host Microbe 2011, 10, 324–335. [Google Scholar] [CrossRef]
- Maslowski, K.M.; Vieira, A.T.; Ng, A.; Kranich, J.; Sierro, F.; Yu, D.; Schilter, H.C.; Rolph, M.S.; Mackay, F.; Artis, D.; et al. Regulation of inflammatory responses by gut microbiota and chemoattractant receptor GPR43. Nature 2009, 461, 1282–1286. [Google Scholar] [CrossRef]
- Hu, S.; Dong, T.S.; Dalal, S.R.; Wu, F.; Bissonnette, M.; Kwon, J.H.; Chang, E.B. The microbe-derived short chain fatty acid butyrate targets miRNA-dependent p21 gene expression in human colon cancer. PLoS ONE 2011, 6, e16221. [Google Scholar] [CrossRef]
- Bindels, L.B.; Porporato, P.; Dewulf, E.M.; Verrax, J.; Neyrinck, A.M.; Martin, J.C.; Scott, K.P.; Buc Calderon, P.; Feron, O.; Muccioli, G.G.; et al. Gut microbiota-derived propionate reduces cancer cell proliferation in the liver. Br. J. Cancer 2012, 107, 1337–1344. [Google Scholar] [CrossRef]
- Luca, S.V.; Macovei, I.; Bujor, A.; Miron, A.; Skalicka-Woźniak, K.; Aprotosoaie, A.C.; Trifan, A. Bioactivity of dietary polyphenols: The role of metabolites. Crit. Rev. Food Sci. Nutr. 2020, 60, 626–659. [Google Scholar] [CrossRef]
- Belkaid, Y.; Hand, T.W. Role of the microbiota in immunity and inflammation. Cell 2014, 157, 121–141. [Google Scholar] [CrossRef]
- Helmink, B.A.; Khan, M.A.W.; Hermann, A.; Gopalakrishnan, V.; Wargo, J.A. The microbiome, cancer, and cancer therapy. Nat. Med. 2019, 25, 377–388. [Google Scholar] [CrossRef] [PubMed]
- Cortez, V.; Schultz-Cherry, S. The role of goblet cells in viral pathogenesis. FEBS J. 2021, 288, 7060–7072. [Google Scholar] [CrossRef] [PubMed]
- Wallaeys, C.; Garcia-Gonzalez, N.; Libert, C. Paneth cells as the cornerstones of intestinal and organismal health: A primer. EMBO Mol. Med. 2023, 15, e16427. [Google Scholar] [CrossRef]
- Levy, M.; Kolodziejczyk, A.A.; Thaiss, C.A.; Elinav, E. Dysbiosis and the immune system. Nat. Rev. Immunol. 2017, 17, 219–232. [Google Scholar] [CrossRef] [PubMed]
- Javaid, N.; Choi, S. Toll-like Receptors from the Perspective of Cancer Treatment. Cancers 2020, 12, 297. [Google Scholar] [CrossRef] [PubMed]
- Köhling, H.L.; Plummer, S.F.; Marchesi, J.R.; Davidge, K.S.; Ludgate, M. The microbiota and autoimmunity: Their role in thyroid autoimmune diseases. Clin. Immunol. 2017, 183, 63–74. [Google Scholar] [CrossRef]
- Ceglia, S.; Berthelette, A.; Howley, K.; Li, Y.; Mortzfeld, B.; Bhattarai, S.K.; Yiew, N.K.H.; Xu, Y.; Brink, R.; Cyster, J.G.; et al. An epithelial cell-derived metabolite tunes immunoglobulin A secretion by gut-resident plasma cells. Nat. Immunol. 2023, 24, 531–544. [Google Scholar] [CrossRef]
- Stacey, H.D.; Golubeva, D.; Posca, A.; Ang, J.C.; Novakowski, K.E.; Zahoor, M.A.; Kaushic, C.; Cairns, E.; Bowdish, D.M.E.; Mullarkey, C.E.; et al. IgA potentiates NETosis in response to viral infection. Proc. Natl. Acad. Sci. USA 2021, 118, e2101497118. [Google Scholar] [CrossRef]
- Macia, L.; Tan, J.; Vieira, A.T.; Leach, K.; Stanley, D.; Luong, S.; Maruya, M.; Ian McKenzie, C.; Hijikata, A.; Wong, C.; et al. Metabolite-sensing receptors GPR43 and GPR109A facilitate dietary fibre-induced gut homeostasis through regulation of the inflammasome. Nat. Commun. 2015, 6, 6734. [Google Scholar] [CrossRef]
- Li, G.; Lin, J.; Zhang, C.; Gao, H.; Lu, H.; Gao, X.; Zhu, R.; Li, Z.; Li, M.; Liu, Z. Microbiota metabolite butyrate constrains neutrophil functions and ameliorates mucosal inflammation in inflammatory bowel disease. Gut Microbes 2021, 13, 1968257. [Google Scholar] [CrossRef]
- Smith, P.M.; Howitt, M.R.; Panikov, N.; Michaud, M.; Gallini, C.A.; Bohlooly, Y.M.; Glickman, J.N.; Garrett, W.S. The microbial metabolites, short-chain fatty acids, regulate colonic Treg cell homeostasis. Science 2013, 341, 569–573. [Google Scholar] [CrossRef]
- Kim, M.; Qie, Y.; Park, J.; Kim, C.H. Gut Microbial Metabolites Fuel Host Antibody Responses. Cell Host Microbe 2016, 20, 202–214. [Google Scholar] [CrossRef]
- WHO. Healthy Diet. 2020. Available online: https://www.who.int/news-room/fact-sheets/detail/healthy-diet (accessed on 7 September 2023).
- Cena, H.; Calder, P.C. Defining a Healthy Diet: Evidence for The Role of Contemporary Dietary Patterns in Health and Disease. Nutrients 2020, 12, 334. [Google Scholar] [CrossRef] [PubMed]
- García-García, F.J.; Monistrol-Mula, A.; Cardellach, F.; Garrabou, G. Nutrition, Bioenergetics, and Metabolic Syndrome. Nutrients 2020, 12, 2785. [Google Scholar] [CrossRef] [PubMed]
- Matrone, A.; Ferrari, F.; Santini, F.; Elisei, R. Obesity as a risk factor for thyroid cancer. Curr. Opin. Endocrinol. Diabetes Obes. 2020, 27, 358–363. [Google Scholar] [CrossRef] [PubMed]
- Ma, X.N.; Ma, C.X.; Hou, L.J.; Fu, S.B. The association of obesity with thyroid carcinoma risk. Cancer Med. 2022, 11, 1136–1144. [Google Scholar] [CrossRef]
- Matrone, A.; Ceccarini, G.; Beghini, M.; Ferrari, F.; Gambale, C.; D’Aqui, M.; Piaggi, P.; Torregrossa, L.; Molinaro, E.; Basolo, F.; et al. Potential Impact of BMI on the Aggressiveness of Presentation and Clinical Outcome of Differentiated Thyroid Cancer. J. Clin. Endocrinol. Metab. 2020, 105, dgz312. [Google Scholar] [CrossRef]
- Elbasan, O.; Yavuz, D.G. Effects of concomitant obesity and diabetes on the aggressiveness and outcomes of differentiated thyroid cancer patients. Arch. Endocrinol. Metab. 2021, 65, 455–461. [Google Scholar] [CrossRef]
- Markaki, I.; Linos, D.; Linos, A. The influence of dietary patterns on the development of thyroid cancer. Eur. J. Cancer 2003, 39, 1912–1919. [Google Scholar] [CrossRef]
- Nguyen, L.T.D.; Gunathilake, M.; Lee, J.; Kim, J. Association between dietary habits and incident thyroid cancer: A prospective cohort study. Front. Nutr. 2023, 10, 1104925. [Google Scholar] [CrossRef]
- Sangsefidi, Z.S.; Ghafouri-Taleghani, F.; Zakavi, S.R.; Norouzy, A.; Kashanifar, R.; Pourbaferani, R.; Safarian, M.; Hosseinzadeh, M. Major dietary patterns and differentiated thyroid cancer. Clin. Nutr. ESPEN 2019, 33, 195–201. [Google Scholar] [CrossRef] [PubMed]
- Llaha, F.; Cayssials, V.; Farràs, M.; Agudo, A.; Sandström, M.; Eriksen, A.K.; Tjønneland, A.; Boutron-Ruault, M.C.; Laouali, N.; Truong, T.; et al. Adherence to mediterranean diet and the risk of differentiated thyroid cancer in a European cohort: The EPIC study. Front. Nutr. 2022, 9, 982369. [Google Scholar] [CrossRef] [PubMed]
- David, L.A.; Maurice, C.F.; Carmody, R.N.; Gootenberg, D.B.; Button, J.E.; Wolfe, B.E.; Ling, A.V.; Devlin, A.S.; Varma, Y.; Fischbach, M.A.; et al. Diet rapidly and reproducibly alters the human gut microbiome. Nature 2014, 505, 559–563. [Google Scholar] [CrossRef]
- De Filippo, C.; Cavalieri, D.; Di Paola, M.; Ramazzotti, M.; Poullet, J.B.; Massart, S.; Collini, S.; Pieraccini, G.; Lionetti, P. Impact of diet in shaping gut microbiota revealed by a comparative study in children from Europe and rural Africa. Proc. Natl. Acad. Sci. USA 2010, 107, 14691–14696. [Google Scholar] [CrossRef] [PubMed]
- Wu, G.D.; Chen, J.; Hoffmann, C.; Bittinger, K.; Chen, Y.Y.; Keilbaugh, S.A.; Bewtra, M.; Knights, D.; Walters, W.A.; Knight, R.; et al. Linking long-term dietary patterns with gut microbial enterotypes. Science 2011, 334, 105–108. [Google Scholar] [CrossRef]
- Hashemi, Z.; Fouhse, J.; Im, H.S.; Chan, C.B.; Willing, B.P. Dietary Pea Fiber Supplementation Improves Glycemia and Induces Changes in the Composition of Gut Microbiota, Serum Short Chain Fatty Acid Profile and Expression of Mucins in Glucose Intolerant Rats. Nutrients 2017, 9, 1236. [Google Scholar] [CrossRef]
- Tan, B.L.; Norhaizan, M.E.; Liew, W.P. Nutrients and Oxidative Stress: Friend or Foe? Oxid. Med. Cell. Longev. 2018, 2018, 9719584. [Google Scholar] [CrossRef]
- Muñoz, A.; Costa, M. Nutritionally mediated oxidative stress and inflammation. Oxid. Med. Cell. Longev. 2013, 2013, 610950. [Google Scholar] [CrossRef]
- Li, Y.; Hecht, S.S. Metabolic Activation and DNA Interactions of Carcinogenic N-Nitrosamines to Which Humans Are Commonly Exposed. Int. J. Mol. Sci. 2022, 23, 4559. [Google Scholar] [CrossRef]
- Zhu, Y.; Wang, P.P.; Zhao, J.; Green, R.; Sun, Z.; Roebothan, B.; Squires, J.; Buehler, S.; Dicks, E.; Zhao, J.; et al. Dietary N-nitroso compounds and risk of colorectal cancer: A case-control study in Newfoundland and Labrador and Ontario, Canada. Br. J. Nutr. 2014, 111, 1109–1117. [Google Scholar] [CrossRef]
- Seyyedsalehi, M.S.; Mohebbi, E.; Tourang, F.; Sasanfar, B.; Boffetta, P.; Zendehdel, K. Association of Dietary Nitrate, Nitrite, and N-Nitroso Compounds Intake and Gastrointestinal Cancers: A Systematic Review and Meta-Analysis. Toxics 2023, 11, 190. [Google Scholar] [CrossRef] [PubMed]
- Hughes, R.; Rowland, I.R. Metabolic activities of the gut microflora in relation to cancer. Microb. Ecol. Health Dis. 2000, 12, 179–185. [Google Scholar] [CrossRef]
- Ou, J.; DeLany, J.P.; Zhang, M.; Sharma, S.; O’Keefe, S.J. Association between low colonic short-chain fatty acids and high bile acids in high colon cancer risk populations. Nutr. Cancer 2012, 64, 34–40. [Google Scholar] [CrossRef] [PubMed]
- Kühn, T.; Stepien, M.; López-Nogueroles, M.; Damms-Machado, A.; Sookthai, D.; Johnson, T.; Roca, M.; Hüsing, A.; Maldonado, S.G.; Cross, A.J.; et al. Prediagnostic plasma bile acid levels and colon cancer risk: A prospective study. J. Natl. Cancer Inst. 2020, 112, 516–524. [Google Scholar] [CrossRef] [PubMed]
- Yang, R.; Qian, L. Research on Gut Microbiota-Derived Secondary Bile Acids in Cancer Progression. Integr. Cancer Ther. 2022, 21, 15347354221114100. [Google Scholar] [CrossRef] [PubMed]
- Flint, H.J.; Scott, K.P.; Duncan, S.H.; Louis, P.; Forano, E. Microbial degradation of complex carbohydrates in the gut. Gut Microbes 2012, 3, 289–306. [Google Scholar] [CrossRef]
- Morrison, D.J.; Preston, T. Formation of short chain fatty acids by the gut microbiota and their impact on human metabolism. Gut Microbes 2016, 7, 189–200. [Google Scholar] [CrossRef]
- Cong, J.; Zhou, P.; Zhang, R. Intestinal Microbiota-Derived Short Chain Fatty Acids in Host Health and Disease. Nutrients 2022, 14, 1977. [Google Scholar] [CrossRef]
- Louis, P.; Young, P.; Holtrop, G.; Flint, H.J. Diversity of human colonic butyrate-producing bacteria revealed by analysis of the butyryl-CoA:acetate CoA-transferase gene. Environ. Microbiol. 2010, 12, 304–314. [Google Scholar] [CrossRef]
- Lee, S.U.; In, H.J.; Kwon, M.S.; Park, B.O.; Jo, M.; Kim, M.O.; Cho, S.; Lee, S.; Lee, H.J.; Kwak, Y.S.; et al. β-Arrestin 2 mediates G protein-coupled receptor 43 signals to nuclear factor-κB. Biol. Pharm. Bull. 2013, 36, 1754–1759. [Google Scholar] [CrossRef]
- Smith, B.C.; Denu, J.M. Chemical mechanisms of histone lysine and arginine modifications. Biochim. Biophys. Acta 2009, 1789, 45–57. [Google Scholar] [CrossRef] [PubMed]
- Säemann, M.D.; Böhmig, G.A.; Osterreicher, C.H.; Burtscher, H.; Parolini, O.; Diakos, C.; Stöckl, J.; Hörl, W.H.; Zlabinger, G.J. Anti-inflammatory effects of sodium butyrate on human monocytes: Potent inhibition of IL-12 and up-regulation of IL-10 production. FASEB J. 2000, 14, 2380–2382. [Google Scholar] [CrossRef] [PubMed]
- Greene, W.C.; Chen, L.F. Regulation of NF-kappaB action by reversible acetylation. Novartis Found. Symp. 2004, 259, 208–225. [Google Scholar]
- Yonezawa, T.; Kobayashi, Y.; Obara, Y. Short-chain fatty acids induce acute phosphorylation of the p38 mitogen-activated protein kinase/heat shock protein 27 pathway via GPR43 in the MCF-7 human breast cancer cell line. Cell. Signal. 2007, 19, 185–193. [Google Scholar] [CrossRef] [PubMed]
- Seljeset, S.; Siehler, S. Receptor-specific regulation of ERK1/2 activation by members of the “free fatty acid receptor” family. J. Recept. Signal Transduct. Res. 2012, 32, 196–201. [Google Scholar] [CrossRef] [PubMed]
- Blaak, E.E.; Canfora, E.E.; Theis, S.; Frost, G.; Groen, A.K.; Mithieux, G.; Nauta, A.; Scott, K.; Stahl, B.; van Harsselaar, J.; et al. Short chain fatty acids in human gut and metabolic health. Benef. Microbes 2020, 11, 411–455. [Google Scholar] [CrossRef]
- Wang, Y.; Wu, Y.; Wang, Y.; Xu, H.; Mei, X.; Yu, D.; Wang, Y.; Li, W. Antioxidant Properties of Probiotic Bacteria. Nutrients 2017, 9, 521. [Google Scholar] [CrossRef]
- Mishra, V.; Shah, C.; Mokashe, N.; Chavan, R.; Yadav, H.; Prajapati, J. Probiotics as potential antioxidants: A systematic review. J. Agric. Food Chem. 2015, 63, 3615–3626. [Google Scholar] [CrossRef]
- Gliszczyńska-Świgło, A. Folates as antioxidants. Food Chem. 2007, 101, 1480–1483. [Google Scholar] [CrossRef]
- Leoni, S.G.; Sastre-Perona, A.; De la Vieja, A.; Santisteban, P. Selenium Increases Thyroid-Stimulating Hormone-Induced Sodium/Iodide Symporter Expression Through Thioredoxin/Apurinic/Apyrimidinic Endonuclease 1-Dependent Regulation of Paired Box 8 Binding Activity. Antioxid. Redox Signal. 2016, 24, 855–866. [Google Scholar] [CrossRef]
- Lin, B.; Zhao, F.; Liu, Y.; Wu, X.; Feng, J.; Jin, X.; Yan, W.; Guo, X.; Shi, S.; Li, Z.; et al. Randomized Clinical Trial: Probiotics Alleviated Oral-Gut Microbiota Dysbiosis and Thyroid Hormone Withdrawal-Related Complications in Thyroid Cancer Patients Before Radioiodine Therapy Following Thyroidectomy. Front. Endocrinol. 2022, 13, 834674. [Google Scholar] [CrossRef] [PubMed]
- Kieliszek, M.; Błażejak, S. Current Knowledge on the Importance of Selenium in Food for Living Organisms: A Review. Molecules 2016, 21, 609. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Roh, Y.J.; Han, S.J.; Park, I.; Lee, H.M.; Ok, Y.S.; Lee, B.C.; Lee, S.R. Role of Selenoproteins in Redox Regulation of Signaling and the Antioxidant System: A Review. Antioxidants 2020, 9, 383. [Google Scholar] [CrossRef] [PubMed]
- Huang, Z.; Rose, A.H.; Hoffmann, P.R. The role of selenium in inflammation and immunity: From molecular mechanisms to therapeutic opportunities. Antioxid. Redox Signal. 2012, 16, 705–743. [Google Scholar] [CrossRef] [PubMed]
- Moncayo, R.; Kroiss, A.; Oberwinkler, M.; Karakolcu, F.; Starzinger, M.; Kapelari, K.; Talasz, H.; Moncayo, H. The role of selenium, vitamin C, and zinc in benign thyroid diseases and of selenium in malignant thyroid diseases: Low selenium levels are found in subacute and silent thyroiditis and in papillary and follicular carcinoma. BMC Endocr. Disord. 2008, 8, 2. [Google Scholar] [CrossRef]
- O’Grady, T.J.; Kitahara, C.M.; DiRienzo, A.G.; Gates, M.A. The association between selenium and other micronutrients and thyroid cancer incidence in the NIH-AARP Diet and Health Study. PLoS ONE 2014, 9, e110886. [Google Scholar] [CrossRef]
- Xu, X.; Hendryx, M.; Liang, X.; Kahe, K.; Li, Y.; Luo, J. Dietary selenium intake and thyroid cancer risk in postmenopausal women. Nutrition 2022, 103–104, 111840. [Google Scholar] [CrossRef]
- Kasaikina, M.V.; Kravtsova, M.A.; Lee, B.C.; Seravalli, J.; Peterson, D.A.; Walter, J.; Legge, R.; Benson, A.K.; Hatfield, D.L.; Gladyshev, V.N. Dietary selenium affects host selenoproteome expression by influencing the gut microbiota. FASEB J. 2011, 25, 2492–2499. [Google Scholar] [CrossRef]
- Callejón-Leblic, B.; Selma-Royo, M.; Collado, M.C.; Abril, N.; García-Barrera, T. Impact of Antibiotic-Induced Depletion of Gut Microbiota and Selenium Supplementation on Plasma Selenoproteome and Metal Homeostasis in a Mice Model. J. Agric. Food Chem. 2021, 69, 7652–7662. [Google Scholar] [CrossRef]
- Severo, J.S.; Morais, J.B.S.; de Freitas, T.E.C.; Andrade, A.L.P.; Feitosa, M.M.; Fontenelle, L.C.; de Oliveira, A.R.S.; Cruz, K.J.C.; do Nascimento Marreiro, D. The Role of Zinc in Thyroid Hormones Metabolism. Int. J. Vitam. Nutr. Res. 2019, 89, 80–88. [Google Scholar] [CrossRef]
- Kucharzewski, M.; Braziewicz, J.; Majewska, U.; Gózdz, S. Copper, zinc, and selenium in whole blood and thyroid tissue of people with various thyroid diseases. Biol. Trace Elem. Res. 2003, 93, 9–18. [Google Scholar] [CrossRef] [PubMed]
- Kazi Tani, L.S.; Gourlan, A.T.; Dennouni-Medjati, N.; Telouk, P.; Dali-Sahi, M.; Harek, Y.; Sun, Q.; Hackler, J.; Belhadj, M.; Schomburg, L.; et al. Copper Isotopes and Copper to Zinc Ratio as Possible Biomarkers for Thyroid Cancer. Front. Med. 2021, 8, 698. [Google Scholar] [CrossRef] [PubMed]
- Pajarillo, E.A.B.; Lee, E.; Kang, D.K. Trace metals and animal health: Interplay of the gut microbiota with iron, manganese, zinc, and copper. Anim. Nutr. 2021, 7, 750–761. [Google Scholar] [CrossRef]
- Wimalawansa, S.J. Vitamin D Deficiency: Effects on Oxidative Stress, Epigenetics, Gene Regulation, and Aging. Biology 2019, 8, 30. [Google Scholar] [CrossRef]
- Roseland, J.M.; Phillips, K.M.; Patterson, K.Y.; Pehrsson, P.R.; Taylor, C.L. Vitamin D in foods: An evolution of knowledge. In Vitamin D, Volume 2: Health, Disease and TherapeuticsI, 4th ed.; Feldman, D., Pike, J.W., Bouillon, R., Giovannucci, E., Goltzman, D., Hewison, M., Eds.; Elsevier: Amsterdam, The Netherlands, 2018; pp. 41–78. [Google Scholar]
- Babić Leko, M.; Jureško, I.; Rozić, I.; Pleić, N.; Gunjača, I.; Zemunik, T. Vitamin D and the Thyroid: A Critical Review of the Current Evidence. Int. J. Mol. Sci. 2023, 24, 3586. [Google Scholar] [CrossRef] [PubMed]
- Moslemi, E.; Musazadeh, V.; Kavyani, Z.; Naghsh, N.; Shoura, S.M.S.; Dehghan, P. Efficacy of vitamin D supplementation as an adjunct therapy for improving inflammatory and oxidative stress biomarkers: An umbrella meta-analysis. Pharmacol. Res. 2022, 186, 106484. [Google Scholar] [CrossRef] [PubMed]
- Heine, G.; Niesner, U.; Chang, H.D.; Steinmeyer, A.; Zügel, U.; Zuberbier, T.; Radbruch, A.; Worm, M. 1,25-dihydroxyvitamin D(3) promotes IL-10 production in human B cells. Eur. J. Immunol. 2008, 38, 2210–2218. [Google Scholar] [CrossRef]
- Krajewska, M.; Witkowska-Sędek, E.; Rumińska, M.; Stelmaszczyk-Emmel, A.; Sobol, M.; Majcher, A.; Pyrżak, B. Vitamin D Effects on Selected Anti-Inflammatory and Pro-Inflammatory Markers of Obesity-Related Chronic Inflammation. Front. Endocrinol. 2022, 13, 920340. [Google Scholar] [CrossRef]
- Sepidarkish, M.; Farsi, F.; Akbari-Fakhrabadi, M.; Namazi, N.; Almasi-Hashiani, A.; Maleki Hagiagha, A.; Heshmati, J. The effect of vitamin D supplementation on oxidative stress parameters: A systematic review and meta-analysis of clinical trials. Pharmacol. Res. 2019, 139, 141–152. [Google Scholar] [CrossRef]
- Cordiano, R.; Di Gioacchino, M.; Mangifesta, R.; Panzera, C.; Gangemi, S.; Minciullo, P.L. Malondialdehyde as a Potential Oxidative Stress Marker for Allergy-Oriented Diseases: An Update. Molecules 2023, 28, 5979. [Google Scholar] [CrossRef]
- Tangestani, H.; Boroujeni, H.K.; Djafarian, K.; Emamat, H.; Shab-Bidar, S. Vitamin D and The Gut Microbiota: A Narrative Literature Review. Clin. Nutr. Res. 2021, 10, 181–191. [Google Scholar] [CrossRef] [PubMed]
- Zhao, J.; Wang, H.; Zhang, Z.; Zhou, X.; Yao, J.; Zhang, R.; Liao, L.; Dong, J. Vitamin D deficiency as a risk factor for thyroid cancer: A meta-analysis of case-control studies. Nutrition 2019, 57, 5–11. [Google Scholar] [CrossRef] [PubMed]
- Ahn, J.H.; Choi, H.; Kim, S.J.; Cho, S.W.; Lee, K.E.; Park, D.J.; Park, Y.J. The association between vitamin D supplementation and the long-term prognosis of differentiated thyroid cancer patients: A retrospective observational cohort study with propensity score matching. Front. Endocrinol. 2023, 14, 1163671. [Google Scholar] [CrossRef] [PubMed]
- Carr, A.C.; Maggini, S. Vitamin C and Immune Function. Nutrients 2017, 9, 1211. [Google Scholar] [CrossRef]
- Kaźmierczak-Barańska, J.; Boguszewska, K.; Adamus-Grabicka, A.; Karwowski, B.T. Two Faces of Vitamin C—Antioxidative and Pro-Oxidative Agent. Nutrients 2020, 12, 1501. [Google Scholar] [CrossRef]
- Pizzino, G.; Irrera, N.; Cucinotta, M.; Pallio, G.; Mannino, F.; Arcoraci, V.; Squadrito, F.; Altavilla, D.; Bitto, A. Oxidative Stress: Harms and Benefits for Human Health. Oxid. Med. Cell. Longev. 2017, 2017, 8416763. [Google Scholar] [CrossRef]
- Farasati Far, B.; Behnoush, A.H.; Ghondaghsaz, E.; Habibi, M.A.; Khalaji, A. The interplay between vitamin C and thyroid. Endocrinol. Diabetes Metab. 2023, 6, e432. [Google Scholar] [CrossRef]
- Tronci, L.; Serreli, G.; Piras, C.; Frau, D.V.; Dettori, T.; Deiana, M.; Murgia, F.; Santoru, M.L.; Spada, M.; Leoni, V.P.; et al. Vitamin C Cytotoxicity and Its Effects in Redox Homeostasis and Energetic Metabolism in Papillary Thyroid Carcinoma Cell Lines. Antioxidants 2021, 10, 809. [Google Scholar] [CrossRef]
- Su, X.; Shen, Z.; Yang, Q.; Sui, F.; Pu, J.; Ma, J.; Ma, S.; Yao, D.; Ji, M.; Hou, P. Vitamin C kills thyroid cancer cells through ROS-dependent inhibition of MAPK/ERK and PI3K/AKT pathways via distinct mechanisms. Theranostics 2019, 9, 4461–4473. [Google Scholar] [CrossRef]
- Su, X.; Li, P.; Han, B.; Jia, H.; Liang, Q.; Wang, H.; Gu, M.; Cai, J.; Li, S.; Zhou, Y.; et al. Vitamin C sensitizes BRAFV600E thyroid cancer to PLX4032 via inhibiting the feedback activation of MAPK/ERK signal by PLX4032. J. Exp. Clin. Cancer Res. 2021, 40, 34. [Google Scholar] [CrossRef]
- Wang, X.; Xu, S.; Zhang, L.; Cheng, X.; Yu, H.; Bao, J.; Lu, R. Vitamin C induces ferroptosis in anaplastic thyroid cancer cells by ferritinophagy activation. Biochem. Biophys. Res. Commun. 2021, 551, 46–53. [Google Scholar] [CrossRef] [PubMed]
- Li, X.Y.; Meng, L.; Shen, L.; Ji, H.F. Regulation of gut microbiota by vitamin C, vitamin E and β-carotene. Food Res. Int. 2023, 169, 112749. [Google Scholar] [CrossRef] [PubMed]
- Zamora-Ros, R.; Knaze, V.; Rothwell, J.A.; Hémon, B.; Moskal, A.; Overvad, K.; Tjønneland, A.; Kyrø, C.; Fagherazzi, G.; Boutron-Ruault, M.C.; et al. Dietary polyphenol intake in Europe: The European Prospective Investigation into Cancer and Nutrition (EPIC) study. Eur. J. Nutr. 2016, 55, 1359–1375. [Google Scholar] [CrossRef]
- de Mello Andrade, J.M.; Fasolo, D. Chapter 20—Polyphenol Antioxidants from Natural Sources and Contribution to Health Promotion. In Polyphenols in Human Health and Disease; Watson, R.R., Preedy, V.R., Zibadi, S., Eds.; Academic Press: Cambridge, MA, USA, 2014; pp. 253–265. [Google Scholar]
- Wan, M.L.Y.; Co, V.A.; El-Nezami, H. Dietary polyphenol impact on gut health and microbiota. Crit. Rev. Food Sci. Nutr. 2021, 61, 690–711. [Google Scholar] [CrossRef]
- Li, J.; Chen, C.; Yang, H.; Yang, X. Tea polyphenols regulate gut microbiota dysbiosis induced by antibiotic in mice. Food Res. Int. 2021, 141, 110153. [Google Scholar] [CrossRef]
- Wang, X.; Qi, Y.; Zheng, H. Dietary Polyphenol, Gut Microbiota, and Health Benefits. Antioxidants 2022, 11, 1212. [Google Scholar] [CrossRef]
- Kafantaris, I.; Kotsampasi, B.; Christodoulou, V.; Kokka, E.; Kouka, P.; Terzopoulou, Z.; Gerasopoulos, K.; Stagos, D.; Mitsagga, C.; Giavasis, I.; et al. Grape pomace improves antioxidant capacity and faecal microflora of lambs. J. Anim. Physiol. Anim. Nutr. 2017, 101, e108–e121. [Google Scholar] [CrossRef]
- Zhao, L.; Zhang, Q.; Ma, W.; Tian, F.; Shen, H.; Zhou, M. A combination of quercetin and resveratrol reduces obesity in high-fat diet-fed rats by modulation of gut microbiota. Food Funct. 2017, 8, 4644–4656. [Google Scholar] [CrossRef]
- Zamora-Ros, R.; Cayssials, V.; Franceschi, S.; Kyrø, C.; Weiderpass, E.; Hennings, J.; Sandström, M.; Tjønneland, A.; Olsen, A.; Overvad, K.; et al. Polyphenol intake and differentiated thyroid cancer risk in the European Prospective Investigation into Cancer and Nutrition (EPIC) cohort. Int. J. Cancer 2020, 146, 1841–1850. [Google Scholar] [CrossRef]
- Zamora-Ros, R.; Lujan-Barroso, L.; Achaintre, D.; Franceschi, S.; Kyrø, C.; Overvad, K.; Tjønneland, A.; Truong, T.; Lecuyer, L.; Boutron-Ruault, M.C.; et al. Blood polyphenol concentrations and differentiated thyroid carcinoma in women from the European Prospective Investigation into Cancer and Nutrition (EPIC) study. Am. J. Clin. Nutr. 2021, 113, 162–171. [Google Scholar] [CrossRef]
- Borzekowski, D.L.G.; Robinson, T.N. The 30-second effect: An experiment revealing the impact of television commercials on food preferences of preschoolers. J. Am. Diet. Assoc. 2001, 101, 42–46. [Google Scholar] [CrossRef] [PubMed]
- Robinson, T.N.; Borzekowski, D.L.; Matheson, D.M.; Kraemer, H.C. Effects of fast-food branding on young children’s taste preferences. Arch. Pediatr. Adolesc. Med. 2007, 161, 792–797. [Google Scholar] [CrossRef] [PubMed]
- Sarafoleanu, C.; Mella, C.; Georgescu, M.; Perederco, C. The importance of the olfactory sense in the human behavior and evolution. J. Med. Life 2009, 2, 196–198. [Google Scholar] [PubMed]
- Sharma, A.; Kumar, R.; Aier, I.; Semwal, R.; Tyagi, P.; Varadwaj, P. Sense of Smell: Structural, Functional, Mechanistic Advancements and Challenges in Human Olfactory Research. Curr. Neuropharmacol. 2019, 17, 891–911. [Google Scholar] [CrossRef]
- Arshamian, A.; Gerkin, R.C.; Kruspe, N.; Wnuk, E.; Floyd, S.; O’Meara, C.; Garrido Rodriguez, G.; Lundström, J.N.; Mainland, J.D.; Majid, A. The perception of odor pleasantness is shared across cultures. Curr. Biol. 2022, 32, 2061–2066.e3. [Google Scholar] [CrossRef]
- Modesti, M.; Tonacci, A.; Sansone, F.; Billeci, L.; Bellincontro, A.; Cacopardo, G.; Sanmartin, C.; Taglieri, I.; Venturi, F. E-Senses, Panel Tests and Wearable Sensors: A Teamwork for Food Quality Assessment and Prediction of Consumer’s Choices. Chemosensors 2022, 10, 244. [Google Scholar] [CrossRef]
- Tirandazi, P.; Bamakan, S.M.; Toghroljerdi, A. A review of studies on internet of everything as an enabler of neuromarketing methods and techniques. J. Supercomput. 2023, 7, 7835–7876. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Study Design | Sample | Country | Microbial Indicators | Other Clinical Data | Reference |
|---|---|---|---|---|---|
| Cross-sectional | Blood sample from 77 patients with PTC (37 patients in distant metastasis group and 40 in ablation group) after total thyroidectomy and radioiodine treatment | China | - | 31 differentially abundant metabolites between the ablation and distant metastasis groups: 12 metabolites with higher concentrations in the distant metastasis group, while 19 metabolites showing lower concentrations. 31 metabolites mostly involved in “alanine, aspartate and glutamate metabolism” and “inositol phosphate metabolism”. | [71] |
| Case-control | Peripheral blood and fecal samples from 30 preoperative patients diagnosed for TC and 35 healthy controls, 18 to 65 years of age | China | Chao index (richness) and Shannon index (richness and evenness) higher in the TC group. Β-diversity (Bray–Curtis method) significantly different between the two groups. Firmicutes and Proteobacteria increase and Bacteroidetes decrease in TC group. | TC-enriched genera, like Lactococcus, Ruminococcaceae_UCG_002, Intestinibacter, Clostridium_sensu_stricto_1, Christensenellaceae_ R-7_group, Lachnospiraceae_ND3007, and Turicibacter, positively correlated with Lpa titers. TC-enriched genera, including Ruminococcaceae_UCG-014 and Lactococcus, negatively correlated with the levels of ApoB. 72 significantly changed metabolites (mainly lipids) in TC group. Some TC-enriched genera, e.g., Klebsiella, Coprococcus_3, and [Eubacterium]_hallii_group, associated with lipid-related metabolites. The adipocytokine signaling, sphingolipid metabolism, cholesterol metabolism, and necroptosis pathways also significantly enriched in TC patients. | [20] |
| Case-control | Fecal specimens from 74 subjects: 20 patients suffering from differentiated TC, 18 patients with thyroid nodules, and 36 healthy controls | China | 35 unique species, like Thiobacillus, Rhodobacter, Rheinheimera, Mycobacterium, and Anaerotruncus, found in TC group. Shannon index not significant. ACE index (microbial abundance) higher in TC and thyroid nodule groups than in control group. UniFrac distance (β-diversity) significantly different between thyroid disease and healthy groups. The gut microbiome of TC group characterized by a dominance of Prevotella, Roseburia, Coprococcus, Anaerostipes, Ruminococcus, Neisseria, Streptococcus, and Porphyromonas. | In TC group, relationships between 22 taxa and 7 clinical indices: significantly positive associations observed between TSH and the genus Porphyromonas and between FT3 and the genus Streptococcus, and significantly inverse associations between TRAb and the families Clostridiaceae and Lachnospiraceae and between TPOAb and the genus Ruminococcus. | [59] |
| Cross-sectional | Stool samples collected prior to surgery from 90 patients with TC (88 cases with PTC and 2 cases with FTC; 56 cases of TCs with local lymph node metastasis—N1 group and 34 of TCs without local lymph node metastasis—N0 group) and 90 healthy controls | China | Reduced richness (Ace index) and diversity (Shannon index) in TC samples. Significant differences in β-diversity (UniFrac distance) between TC and healthy control groups. No significant difference in the Firmicutes/Bacteroidetes ratio between the two groups. Relatively higher abundance of Proteobacteria in TC patients. | No significant difference in Ace and Shannon indices between N0 and N1 subgroups. A four-genus microbial signature able to distinguish TC patients in N0 from those in N1 (Hungatella, Alistipes, Fusobacterium, and Phascolarctobacterium). Five categories related to the processing of genetic information (‘‘Aminoacyl-tRNA biosynthesis,” ‘‘Homologous recombination,” ‘‘Mismatch repair,” ‘‘DNA replication,” and ‘‘Nucleotide excision repair”) significantly increased in the healthy controls compared to TC patients. | [79] |
| Study Design | Sample | Country | Microbial Indicators | Other Clinical Data | Reference |
|---|---|---|---|---|---|
| Cross-sectional | 25 patients with TC (19 malignant and 6 benign tumors), aged 25–66 years. Stool samples collected before thyroidectomy. Tissue samples removed during surgery and divided into tumor, paratumor, and normal tissues. | China | α-diversity (Chao1, Shannon, and Simpson indices) different in all types of samples. All fecal diversities higher than those in tissues. Diversity in malignant patients lower than that in benign patients, and decreasing with distance from cancerous tissues. Proteobacteria the most abundant in all three types of tissues, while Firmicutes dominant in stool samples. | The microbiome from thyroid completely different from that of the gut, with 18 of the 30 pathways significantly different. Two of the most abundant operational taxonomic units, Pseudomonas mucidolens and Escherichia fergusonii, associated with several processes. Several microbes negatively related with PTH, while Neisseria perflava positively related with TSH and T3. | [1] |
| Retrospective | Tumor and peritumor tissues collected from 30 patients with PTC who underwent total thyroidectomy. | China | Chao1 index (richness) and Shannon index (richness and evenness) lower in tumor tissues than in peritumor tissues (the first index not significant). β-diversity (Bray–Curtis method) significantly different between the two groups. At the genus level, abundance of Sphingomonas and Aeromonas significantly increased in tumor tissues, whereas the abundance of Comamonas, Acinetobacter, Peptostreptococcus, and Proteus significantly increased in peritumor tissues. | Chao1 index significantly higher in N1 stage (presence of lymph node metastasis) than in N0 stage (absence of lymph node metastasis); Shannon index significantly lower in N1 stage than in N0 stage. Abundance of Sphingomonas significantly increased in N1 stage compared to N0 stage. No significant difference in Chao1 and Shannon indices between male and female patients. No difference in the diversity and composition between patients <55 and ≥55. | [12] |
| Retrospective | Raw whole-transcriptome RNA-sequencing, Level 3 normalized mRNA expression read counts, and DNA methylation 450 k sequencing data for untreated, nonirradiated tumor, and adjacent normal tissue were downloaded from the Genomic Data Commons legacy archive for 563 TC patients (354 CPTC, 101 FVPTC, 35 TCPTC, 135 male, 366 female tumor samples: 505 tumor and 58 normal tissue samples). | United States | 45 microbes in CPTC, 34 in FVPTC, and 33 in TCPTC differentially abundant between tumor and normal tissue. 33 microbes in male samples and 49 microbes in female samples to be differentially abundant between tumor and normal tissue. Micrococcus luteus, Frankia sp., Anabaena sp. K119, and uncultured Gammaproteobacteria bacterium all similarly overabundant in normal tissue in CPTC, FVPTC, and TCPTC. In males, Synechococcus sp. CC9311 overabundant in the normal samples, while in females, overabundant in tumor samples. | Frankia sp. and uncultured Gammaproteobacteria bacterium Gammaproteobacteria bacterium, which are abundant in all PTC normal tissue samples, correlated with lower MACIS score). Bradyrhizobium sp. BTAi1, which is uniquely abundant in TCPTC normal tissue, correlated with higher MACIS score. Frankia sp. and Anabaena sp. K119, both of which are overabundant in normal tissue samples of all PTC subtypes, negatively correlated with pathologic M stage, while Stenotrophomonas maltophilia, dysregulated in only CPTC and FVPTC, positively correlated with pathologic M stage. Microbe abundance in FVPTC correlated with the greatest number of chromosomal alterations and mutations. Microbe abundance in males more frequently and strongly correlated with a greater number of CNV. Microbes dysregulated in TCPTC and male patients correlated with higher expression of the BRAFV600E mutation. The majority of most enriched pathways uniquely dysregulated in the different subtypes related to cell growth, in males belonging to the tumor suppression-related group, and in females, belonging to the DNA checkpoint and damage-related group. Lower microbe abundance correlated with greater extent of methylation at known tumor suppressor genes. | [97] |
| Cross-sectional | Tumor samples from 80 patients with PTC. | China | α-diversity significantly lower in patients with T1/T2 PTC than in those with T3/T4 PTC (Shannon and Simpson indices). No significant differences in richness (Sobs index) between clinical stages. β-diversity (Bray–Curtis metric distances) significantly different with tumor progression, with differences in Psudomonas, Rhodococcus, and Sphingomas in abundance among various stages. Pseudomonas spp., presenting a higher abundance in tumors of patients with T1 and T2 PTC than in T3 or T4; Rhodococcus abundance significantly higher in T1 than in T3 PTC; and Sphingomonas more abundant in T1 and T2 than in T3. Eight-genera microbiome signature between T1_2 and T3_4 groups. α-diversity significantly lower in males compared to females, but no significant differences in β-diversity by sex. Rhodococcus, Ralstonia, Chryseobacterium, and Burkholderia-Caballeronia-Paraburkholderia more abundant in females than in males. | In T1_2 group, higher proportions of most metabolic pathways, while in T3_4 group, higher proportions of super pathways of purine nucleotide de novo biosynthesis and palmitate biosynthesis II. Positive associations between FT4 and Neisseria and norank_f__norank_o__Chloroplast, and FT3 and Treponema. Negative associations between FT4 and Klebsiella; T4 and Klebsiella and Escherichia-Shigella, T3 and Granulicatella; and TSH and norank_f__norank_o__Clostridia_UCG-014 and Prevotella. A negative correlation between anti-T SHR levels and Klebsiella and Burkholderia-Caballeronia-Paraburkholderia. Positive correlation of the anti-TG levels with Sphingomonas, Rhodococcus, Ralstonia, and Brevundimonas, but negative correlation with Anaerococcus and Akkermansia. Nine genera (UCG-002, Streptococcus, Parvimonas, Akkermansia, Bacteroides, Haemophilus, Selenomonas, Prevotella, and Bifidobacterium) negatively correlated with the anti-TPO levels. | [82] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gorini, F.; Tonacci, A. Tumor Microbial Communities and Thyroid Cancer Development—The Protective Role of Antioxidant Nutrients: Application Strategies and Future Directions. Antioxidants 2023, 12, 1898. https://doi.org/10.3390/antiox12101898
Gorini F, Tonacci A. Tumor Microbial Communities and Thyroid Cancer Development—The Protective Role of Antioxidant Nutrients: Application Strategies and Future Directions. Antioxidants. 2023; 12(10):1898. https://doi.org/10.3390/antiox12101898
Chicago/Turabian StyleGorini, Francesca, and Alessandro Tonacci. 2023. "Tumor Microbial Communities and Thyroid Cancer Development—The Protective Role of Antioxidant Nutrients: Application Strategies and Future Directions" Antioxidants 12, no. 10: 1898. https://doi.org/10.3390/antiox12101898
APA StyleGorini, F., & Tonacci, A. (2023). Tumor Microbial Communities and Thyroid Cancer Development—The Protective Role of Antioxidant Nutrients: Application Strategies and Future Directions. Antioxidants, 12(10), 1898. https://doi.org/10.3390/antiox12101898