

Antioxidant, Alpha-Glucosidase Inhibition Activities, In Silico Molecular Docking and Pharmacokinetics Study of Phenolic Compounds from Native Australian Fruits and Spices




Abstract
:
1. Introduction
2. Materials and Methods
2.1. Chemicals and Reagents
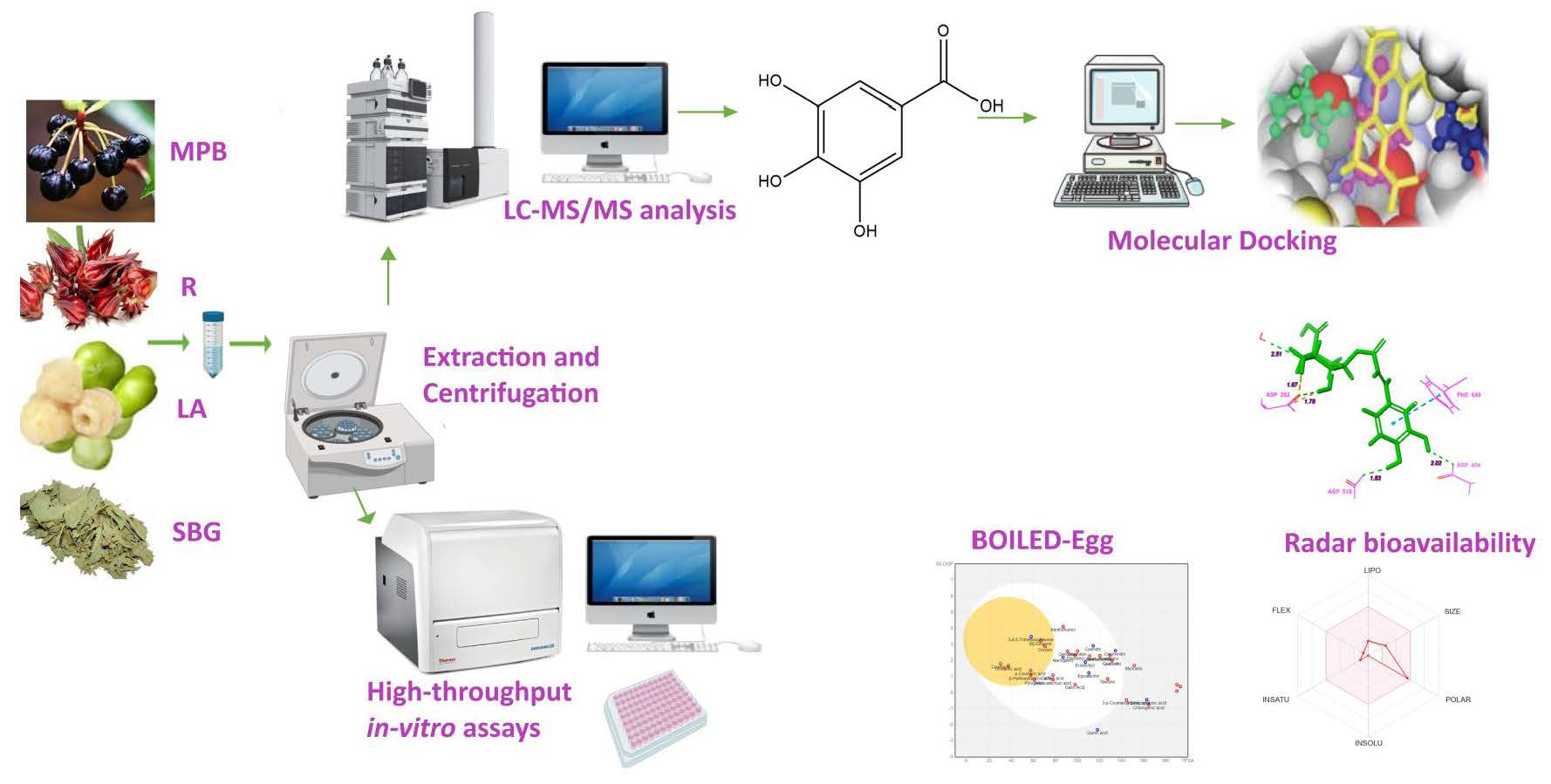
2.2. Preparation and Extraction of Phenolic Compounds
2.3. Measurement of Phenolic Contents and Biological Activities
2.4. LC-MS/MS Analysis
2.5. Molecular Docking and Pharmacokinetic Properties of Abundant Phenolic Compounds
2.6. Statistical Analysis
3. Results and Discussion
3.1. Measurement of Total Polyphenols (TPC, TFC, TMAC, TCT)
3.2. Biological Activities of Native Australian Fruits and Spices
3.3. Correlation Analysis
3.4. LC-MS Analysis
3.4.1. Phenolic Acids
Benzoic acid and Its Derivatives
Cinnamic Acids and Derivatives
3.4.2. Flavonoids
Anthocyanins
Flavanols
Flavanones
Flavones and Isoflavones
Flavonols, Dihydroflavonols, and Chalcones
3.4.3. Isoflavonoids
3.4.4. Tannins
3.4.5. Lignans and Stilbenes
3.4.6. Other Compounds
3.5. Quantification/Semi-Quantification of Targeted Phenolic Compounds
3.6. Molecular Docking
3.7. Pharmacokinetics Study of Selected Phenolic Compounds
3.7.1. Absorption and Distribution
3.7.2. Drug-Likeness
3.7.3. Metabolism, Excretion, and Toxicity
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ali, A.; Cottrell, J.J.; Dunshea, F.R. Identification and characterization of anthocyanins and non-anthocyanin phenolics from australian native fruits and their antioxidant, antidiabetic, and anti-alzheimer potential. Food Res. Int. 2022, 162, 111951. [Google Scholar] [CrossRef]
- Ali, A.; Zahid, H.F.; Cottrell, J.J.; Dunshea, F.R. A comparative study for nutritional and phytochemical profiling of coffea arabica (c. Arabica) from different origins and their antioxidant potential and molecular docking. Molecules 2022, 27, 5126. [Google Scholar]
- Tsao, R. Chemistry and biochemistry of dietary polyphenols. Nutrients 2010, 2, 1231–1246. [Google Scholar]
- Kiloni, S.M.; Akhtar, A.; Cáceres-Vélez, P.R.; Dunshea, F.; Jusuf, P. P06-05 zebrafish embryo acute toxicity and antioxidant characterization of native australian plants: Towards safe and effective glaucoma treatments. Toxicol. Lett. 2022, 368, S115. [Google Scholar]
- Cock, I.E. Medicinal and aromatic plants–australia. Ethnopharmacol. Encycl. Life Support Syst. EOLSS 2011, 1, 1–173. [Google Scholar]
- Richmond, R.; Bowyer, M.; Vuong, Q. Australian native fruits: Potential uses as functional food ingredients. J. Funct. Foods 2019, 62, 103547. [Google Scholar]
- Attique, S.A.; Hassan, M.; Usman, M.; Atif, R.M.; Mahboob, S.; Al-Ghanim, K.A.; Bilal, M.; Nawaz, M.Z. A molecular docking approach to evaluate the pharmacological properties of natural and synthetic treatment candidates for use against hypertension. Int. J. Environ. Res. Public Health 2019, 16, 923. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bahun, M.; Jukić, M.; Oblak, D.; Kranjc, L.; Bajc, G.; Butala, M.; Bozovičar, K.; Bratkovič, T.; Podlipnik, Č.; Poklar Ulrih, N. Inhibition of the sars-cov-2 3cl(pro) main protease by plant polyphenols. Food Chem. 2022, 373, 131594. [Google Scholar] [PubMed]
- Jukič, M.; Janežič, D.; Bren, U. Potential novel thioether-amide or guanidine-linker class of sars-cov-2 virus rna-dependent rna polymerase inhibitors identified by high-throughput virtual screening coupled to free-energy calculations. Int. J. Mol. Sci. 2021, 22, 11143. [Google Scholar] [CrossRef] [PubMed]
- Ali, A.; Wu, H.; Ponnampalam, E.N.; Cottrell, J.J.; Dunshea, F.R.; Suleria, H.A.R. Comprehensive profiling of most widely used spices for their phenolic compounds through lc-esi-qtof-ms2 and their antioxidant potential. Antioxidants 2021, 10, 721. [Google Scholar]
- Sharifi-Rad, J.; Song, S.; Ali, A.; Subbiah, V.; Taheri, Y.; Suleria, H.A.R. Lc-esi-qtof-ms/ms characterization of phenolic compounds from pyracantha coccinea m. Roem. And their antioxidant capacity. Cell. Mol. Biol. 2021, 67, 201–211. [Google Scholar] [CrossRef] [PubMed]
- Chou, O.; Ali, A.; Subbiah, V.; Barrow, C.J.; Dunshea, F.R.; Suleria, H.A.R. Lc-esi-qtof-ms/ms characterisation of phenolics in herbal tea infusion and their antioxidant potential. Fermentation 2021, 7, 73. [Google Scholar]
- Zahid, H.F.; Ali, A.; Ranadheera, C.S.; Fang, Z.; Dunshea, F.R.; Ajlouni, S. In vitro bioaccessibility of phenolic compounds and alpha-glucosidase inhibition activity in yoghurts enriched with mango peel powder. Food Biosci. 2022, 50, 102011. [Google Scholar]
- Bashmil, Y.M.; Ali, A.; BK, A.; Dunshea, F.R.; Suleria, H.A.R. Screening and characterization of phenolic compounds from australian grown bananas and their antioxidant capacity. Antioxidants 2021, 10, 1521. [Google Scholar]
- Ali, A.; Cottrell, J.J.; Dunshea, F.R. Lc-ms/ms characterization of phenolic metabolites and their antioxidant activities from australian native plants. Metabolites 2022, 12, 1016. [Google Scholar] [PubMed]
- Ali, A.; Kiloni, S.M.; Cáceres-Vélez, P.R.; Jusuf, P.R.; Cottrell, J.J.; Dunshea, F.R. Phytochemicals, antioxidant activities, and toxicological screening of native australian fruits using zebrafish embryonic model. Foods 2022, 11, 4038. [Google Scholar]
- Daina, A.; Michielin, O.; Zoete, V. Swissadme: A free web tool to evaluate pharmacokinetics, drug-likeness and medicinal chemistry friendliness of small molecules. Sci. Rep. 2017, 7, 42717. [Google Scholar]
- Ali, A.; Bashmil, Y.M.; Cottrell, J.J.; Suleria, H.A.R.; Dunshea, F.R. Lc-ms/ms-qtof screening and identification of phenolic compounds from australian grown herbs and their antioxidant potential. Antioxidants 2021, 10, 1770. [Google Scholar] [PubMed]
- Konczak, I.; Zabaras, D.; Dunstan, M.; Aguas, P. Antioxidant capacity and phenolic compounds in commercially grown native australian herbs and spices. Food Chem. 2010, 122, 260–266. [Google Scholar] [CrossRef]
- Cáceres-Vélez, P.R.; Ali, A.; Fournier-Level, A.; Dunshea, F.R.; Jusuf, P.R. Phytochemical and safety evaluations of finger lime, mountain pepper, and tamarind in zebrafish embryos. Antioxidants 2022, 11, 1280. [Google Scholar] [CrossRef] [PubMed]
- Vélez, P.R.C.; Ali, A.; Fournier-Level, A.; Dunshea, F.; Jusuf, P.R. P06-04 antioxidant activity and embryotoxicity of citrus australasica, tasmannia lanceolata and diploglottis australis extracts in zebrafish. Toxicol. Lett. 2022, 368, S114. [Google Scholar] [CrossRef]
- Hu, T.; Subbiah, V.; Wu, H.; Bk, A.; Rauf, A.; Alhumaydhi, F.A.; Suleria, H.A.R. Determination and characterization of phenolic compounds from australia-grown sweet cherries (prunus avium l.) and their potential antioxidant properties. ACS Omega 2021, 6, 34687–34699. [Google Scholar] [CrossRef]
- Lukmanto, S.; Roesdiyono, N.; Ju, Y.-H.; Indraswati, N.; Soetaredjo, F.E.; Ismadji, S. Supercritical co2 extraction of phenolic compounds in roselle (hibiscus sabdariffa l.). Chem. Eng. Commun. 2013, 200, 1187–1196. [Google Scholar] [CrossRef] [Green Version]
- Liu, H.; Qiu, N.; Ding, H.; Yao, R. Polyphenols contents and antioxidant capacity of 68 chinese herbals suitable for medical or food uses. Food Res. Int. 2008, 41, 363–370. [Google Scholar]
- Yang, W.-J.; Li, D.-P.; Li, J.-K.; Li, M.-H.; Chen, Y.-L.; Zhang, P.-Z. Synergistic antioxidant activities of eight traditional chinese herb pairs. Biol. Pharm. Bull. 2009, 32, 1021–1026. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yoo, K.M.; Lee, C.H.; Lee, H.; Moon, B.; Lee, C.Y. Relative antioxidant and cytoprotective activities of common herbs. Food Chem. 2008, 106, 929–936. [Google Scholar] [CrossRef]
- Tsai, M.-L.; Lin, C.-C.; Lin, W.-C.; Yang, C.-H. Antimicrobial, antioxidant, and anti-inflammatory activities of essential oils from five selected herbs. Biosci. Biotechnol. Biochem. 2011, 75, 1977–1983. [Google Scholar]
- Lin, C.-C.; Yang, C.-H.; Wu, P.-S.; Kwan, C.-C.; Chen, Y.-S. Antimicrobial, anti-tyrosinase and antioxidant activities of aqueous aromatic extracts from forty-eight selected herbs. J. Med. Plants Res. 2011, 5, 6203–6209. [Google Scholar]
- Garg, D.; Muley, A.; Khare, N.; Marar, T. Comparative analysis of phytochemical profile and antioxidant activity of some indian culinary herbs. Res. J. Pharm. Biol. Chem. Sci. 2012, 3, 845–854. [Google Scholar]
- Chen, I.-C.; Chang, H.-C.; Yang, H.-W.; Chen, G.-L. Evaluation of total antioxidant activity of several popular vegetables and chinese herbs: A fast approach with abts/h2o2/hrp system in microplates. J. Food Drug Anal. 2004, 12, 29–33. [Google Scholar] [CrossRef]
- Köksal, E.; Bursal, E.; Gülçin, İ.; Korkmaz, M.; Çağlayan, C.; Gören, A.C.; Alwasel, S.H. Antioxidant activity and polyphenol content of turkish thyme (thymus vulgaris) monitored by liquid chromatography and tandem mass spectrometry. Int. J. Food Prop. 2017, 20, 514–525. [Google Scholar] [CrossRef] [Green Version]
- Wojdyło, A.; Oszmiański, J.; Czemerys, R. Antioxidant activity and phenolic compounds in 32 selected herbs. Food Chem. 2007, 105, 940–949. [Google Scholar]
- Gülçin, İ. Antioxidant activity of caffeic acid (3, 4-dihydroxycinnamic acid). Toxicology 2006, 217, 213–220. [Google Scholar] [CrossRef] [PubMed]
- Gülçin, İ.; Huyut, Z.; Elmastaş, M.; Aboul-Enein, H.Y. Radical scavenging and antioxidant activity of tannic acid. Arab. J. Chem. 2010, 3, 43–53. [Google Scholar]
- Gülçın, İ.; Oktay, M.; Kıreçcı, E.; Küfrevıoǧlu, Ö.İ. Screening of antioxidant and antimicrobial activities of anise (pimpinella anisum l.) seed extracts. Food Chem. 2003, 83, 371–382. [Google Scholar]
- Syabana, M.A.; Yuliana, N.D.; Batubara, I.; Fardiaz, D. Antidiabetic activity screening and nmr profile of vegetable and spices commonly consumed in indonesia. Food Sci. Technol. 2020, 41, 254–264. [Google Scholar] [CrossRef]
- Taslimi, P.; Köksal, E.; Gören, A.C.; Bursal, E.; Aras, A.; Kılıç, Ö.; Alwasel, S.; Gülçin, İ. Anti-alzheimer, antidiabetic and antioxidant potential of satureja cuneifolia and analysis of its phenolic contents by lc-ms/ms. Arab. J. Chem. 2020, 13, 4528–4537. [Google Scholar]
- Cottrell, J.J.; Le, H.H.; Artaiz, O.; Iqbal, Y.; Suleria, H.A.; Ali, A.; Celi, P.; Dunshea, F.R. Recent advances in the use of phytochemicals to manage gastrointestinal oxidative stress in poultry and pigs. Anim. Prod. Sci. 2022, 62, 1140–1146. [Google Scholar]
- Granato, D.; Shahidi, F.; Wrolstad, R.; Kilmartin, P.; Melton, L.D.; Hidalgo, F.J.; Miyashita, K.; van Camp, J.; Alasalvar, C.; Ismail, A.B. Antioxidant activity, total phenolics and flavonoids contents: Should we ban in vitro screening methods? Food Chem. 2018, 264, 471–475. [Google Scholar] [CrossRef]
- Freeman, B.L.; Eggett, D.L.; Parker, T.L. Synergistic and antagonistic interactions of phenolic compounds found in navel oranges. J. Food Sci. 2010, 75, C570–C576. [Google Scholar] [CrossRef]
- Shan, B.; Cai, Y.Z.; Sun, M.; Corke, H. Antioxidant capacity of 26 spice extracts and characterization of their phenolic constituents. J. Agric. Food Chem. 2005, 53, 7749–7759. [Google Scholar]
- Mandal, S.M.; Chakraborty, D.; Dey, S. Phenolic acids act as signaling molecules in plant-microbe symbioses. Plant Signal. Behav. 2010, 5, 359–368. [Google Scholar]
- Goleniowski, M.; Bonfill, M.; Cusido, R.; Palazón, J. Phenolic acids. Nat. Prod. 2013, 2013, 1951–1953. [Google Scholar]
- Kakkar, S.; Bais, S. A review on protocatechuic acid and its pharmacological potential. ISRN Pharmacol. 2014, 2014, 952943. [Google Scholar] [PubMed] [Green Version]
- Hossain, M.B.; Rai, D.K.; Brunton, N.P.; Martin-Diana, A.B.; Barry-Ryan, C. Characterization of phenolic composition in lamiaceae spices by lc-esi-ms/ms. J. Agric. Food Chem. 2010, 58, 10576–10581. [Google Scholar] [PubMed]
- Dong, J.; Zhu, Y.; Gao, X.; Chang, Y.; Wang, M.; Zhang, P. Qualitative and quantitative analysis of the major constituents in chinese medicinal preparation dan-lou tablet by ultra high performance liquid chromatography/diode-array detector/quadrupole time-of-flight tandem mass spectrometry. J. Pharm. Biomed. Anal. 2013, 80, 50–62. [Google Scholar] [CrossRef]
- Kadam, D.; Palamthodi, S.; Lele, S.S. Lc-esi-q-tof-ms/ms profiling and antioxidant activity of phenolics from l. Sativum seedcake. J. Food Sci. Technol. 2018, 55, 1154–1163. [Google Scholar] [PubMed]
- Aherne, S.A.; O’Brien, N.M. Dietary flavonols: Chemistry, food content, and metabolism. Nutrition 2002, 18, 75–81. [Google Scholar]
- Oh, Y.S.; Lee, J.H.; Yoon, S.H.; Oh, C.H.; Choi, D.S.; Choe, E.; Jung, M.Y. Characterization and quantification of anthocyanins in grape juices obtained from the grapes cultivated in korea by hplc/dad, hplc/ms, and hplc/ms/ms. J. Food Sci. 2008, 73, C378–C389. [Google Scholar]
- Liu, Y.; Tikunov, Y.; Schouten, R.E.; Marcelis, L.F.M.; Visser, R.G.F.; Bovy, A. Anthocyanin biosynthesis and degradation mechanisms in solanaceous vegetables: A review. Front. Chem. 2018, 6, 52. [Google Scholar] [CrossRef]
- Murkovic, M. Phenolic compounds: Occurrence, classes, and analysis. In Encyclopedia of Food and Health; Caballero, B., Finglas, P.M., Toldrá, F., Eds.; Academic Press: Oxford, UK, 2016; pp. 346–351. [Google Scholar]
- Rockenbach, I.I.; Jungfer, E.; Ritter, C.; Santiago-Schübel, B.; Thiele, B.; Fett, R.; Galensa, R. Characterization of flavan-3-ols in seeds of grape pomace by ce, hplc-dad-msn and lc-esi-fticr-ms. Food Res. Int. 2012, 48, 848–855. [Google Scholar]
- Zahid, H.F.; Ali, A.; Ranadheera, C.S.; Fang, Z.; Ajlouni, S. Identification of phenolics profile in freeze-dried apple peel and their bioactivities during in vitro digestion and colonic fermentation. Int. J. Mol. Sci. 2023, 24, 1514. [Google Scholar]
- Thuan, N.H.; Pandey, R.P.; Thuy, T.T.; Park, J.W.; Sohng, J.K. Improvement of regio-specific production of myricetin-3-o-α-l-rhamnoside in engineered escherichia coli. Appl. Biochem. Biotechnol. 2013, 171, 1956–1967. [Google Scholar] [CrossRef]
- Habtemariam, S. A-glucosidase inhibitory activity of kaempferol-3-o-rutinoside. Nat. Prod. Commun. 2011, 6, 201–203. [Google Scholar] [CrossRef] [Green Version]
- Chang, Z.; Zhang, Q.; Liang, W.; Zhou, K.; Jian, P.; She, G.; Zhang, L. A comprehensive review of the structure elucidation of tannins from terminalia linn. Evid. Based Complement. Altern. Med. 2019, 2019, 8623909. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hou, K.; Wang, Z. Application of nanotechnology to enhance adsorption and bioavailability of procyanidins: A review. Food Rev. Int. 2021, 38, 738–752. [Google Scholar] [CrossRef]
- Kitts, D.D.; Yuan, Y.V.; Wijewickreme, A.N.; Thompson, L.U. Antioxidant activity of the flaxseed lignan secoisolariciresinol diglycoside and its mammalian lignan metabolites enterodiol and enterolactone. Mol. Cell. Biochem. 1999, 202, 91–100. [Google Scholar]
- Liu, Z.; Fei, Y.J.; Cao, X.H.; Xu, D.; Tang, W.J.; Yang, K.; Xu, W.X.; Tang, J.H. Lignans intake and enterolactone concentration and prognosis of breast cancer: A systematic review and meta-analysis. J. Cancer 2021, 12, 2787–2796. [Google Scholar] [PubMed]
- Konczak, I.; Zabaras, D.; Dunstan, M.; Aguas, P. Antioxidant capacity and hydrophilic phytochemicals in commercially grown native australian fruits. Food Chem. 2010, 123, 1048–1054. [Google Scholar]
- Pantiora, P.; Furlan, V.; Matiadis, D.; Mavroidi, B.; Perperopoulou, F.; Papageorgiou, A.C.; Sagnou, M.; Bren, U.; Pelecanou, M.; Labrou, N.E. Monocarbonyl curcumin analogues as potent inhibitors against human glutathione transferase p1-1. Antioxidants 2023, 12, 63. [Google Scholar] [CrossRef]
- Khalfaoui, A.; Noumi, E.; Belaabed, S.; Aouadi, K.; Lamjed, B.; Adnan, M.; Defant, A.; Kadri, A.; Snoussi, M.; Khan, M.A.; et al. Lc-esi/ms-phytochemical profiling with antioxidant, antibacterial, antifungal, antiviral and in silico pharmacological properties of algerian asphodelus tenuifolius (cav.) organic extracts. Antioxidants 2021, 10, 628. [Google Scholar] [CrossRef] [PubMed]
- Khan, J.; Deb, P.K.; Priya, S.; Medina, K.D.; Devi, R.; Walode, S.G.; Rudrapal, M. Dietary flavonoids: Cardioprotective potential with antioxidant effects and their pharmacokinetic, toxicological and therapeutic concerns. Molecules 2021, 26, 4021. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Variables | TPC mg GAE/g | TFC mg QE/g | TCT mg CE/g | TMAC mg C3GE/g |
|---|---|---|---|---|
| Rosella | 5.65 ± 0. 48 b | 1.33 ± 0.10 c | 1.26 ± 1.13 d | 0.08 ± 0.02 |
| Mountain pepper berries | 6.10 ± 0.34 cd | 1.73 ± 0.15 b | 2.37 ± 0.10 b | 0.17 ± 0.03 |
| Lemon aspen | 4.40 ± 0.38 c | 0.79 ± 0.04 d | 1.80 ± 0.35 c | 0.00 ± 0.00 |
| Strawberry gum | 36.57 ± 1.34 a | 15.69 ± 2.69 a | 8.05 ± 0.52 a | 0.00 ± 0.00 |
| Variables | TPC | TFC | TCT | TMAC | DPPH | ABTS | FRAP | PMA | FICA | •OH-RSA | RPA |
|---|---|---|---|---|---|---|---|---|---|---|---|
| TFC | 1.00 | ||||||||||
| TCT | 0.99 | 0.99 | |||||||||
| TMAC | −0.48 | −0.47 | −0.44 | ||||||||
| DPPH | 0.98 | 0.98 | 0.95 | −0.38 | |||||||
| ABTS | 0.62 | 0.63 | 0.66 | 0.38 | 0.66 | ||||||
| FRAP | 0.92 | 0.92 | 0.87 | −0.31 | 0.98 | 0.64 | |||||
| PMA | −0.21 | −0.20 | −0.13 | 0.90 | −0.18 | 0.62 | −0.19 | ||||
| FICA | 0.88 | 0.88 | 0.91 | −0.04 | 0.87 | 0.91 | 0.80 | 0.28 | |||
| •OH-RSA | 0.97 | 0.97 | 0.96 | −0.27 | 0.99 | 0.76 | 0.96 | −0.04 | 0.93 | ||
| RPA | 0.99 | 0.99 | 1.00 | −0.46 | 0.95 | 0.65 | 0.86 | −0.14 | 0.91 | 0.95 | |
| * α-glu | 0.79 | 0.80 | 0.84 | 0.11 | 0.79 | 0.95 | 0.71 | 0.43 | 0.99 | 0.87 | 0.83 |
| No. | Proposed Compounds | Molecular Formula | RT (min) | ESI +/− | Theoretical (m/z) | Observed (m/z) | Mass Error (ppm) | MS/MS | Samples |
|---|---|---|---|---|---|---|---|---|---|
| Phenolic acids | |||||||||
| Hydroxybenzoic acids and derivatives | |||||||||
| 1 | Gallic acid 4-O-glucoside | C13H16O10 | 5.212 | [M−H]− | 331.0671 | 331.0676 | 1.5 | 169, 125 | MPB |
| 2 | Gallic acid | C7H6O5 | 6.288 | [M−H]− | 169.0142 | 169.0140 | −1.2 | 125 | MPB, SBG, R |
| 3 | Protocatechuic acid 4-O-glucoside | C13H16O9 | 8.861 | [M−H]− | 315.0721 | 315.0699 | −7.0 | 153, 109 | R, MPB |
| 4 | Protocatechuic acid | C7H6O4 | 9.341 | [M−H]− | 153.0193 | 153.0179 | −9.1 | 109 | MPB, SBG, R |
| 5 | p-Hydroxybenzoic acid | C7H6O3 | 12.359 | [M−H]− | 137.0244 | 137.0244 | 0.0 | 93 | R, MPB |
| 6 | 3-O-Methylgallic acid | C8H8O5 | 14.708 | [M−H]− | 183.0299 | 183.0296 | −1.6 | 168, 124, 78 | R, SBG |
| 7 | Benzoic acid | C7H6O2 | 26.507 | [M−H]− | 121.0295 | 121.0300 | 4.1 | 77 | R, LA, SBG, MBP |
| 8 | 4-Hydroxybenzoic acid 4-O-glucoside | C13H16O8 | 35.326 | [M−H]− | 299.0772 | 299.0773 | 0.3 | 255, 137 | SBG, R |
| Cinnamic acids and derivatives | |||||||||
| 9 | 3-Feruloyquinic acid lactone | C17H18O8 | 3.926 | [M−H]− | 349.0929 | 349.0921 | −2.3 | 193, 191, 178 | SBG |
| 10 | p-Coumaroyl malic acid | C13H12O7 | 4.495 | [M−H]− | 279.0510 | 279.0503 | −2.5 | 163 | LA, SBG |
| 11 | Ferulic acid 4-O-glucuronide | C16H18O10 | 6.585 | [M−H]− | 369.0827 | 369.0828 | 0.3 | 193 | R, MPB, SBG, LA |
| 12 | 1-O-Caffeoyl-ꞵ-D-glucose | C15H18O9 | 13.291 | [M−H]− | 341.0873 | 341.0870 | −0.9 | 179, 135 | SBG |
| 13 | Feruloyl tartaric acid | C14H14O9 | 14.091 | [M−H]− | 325.0565 | 325.0562 | −0.9 | 193, 149, 105 | SBG |
| 14 | Rosmarinic acid | C18H16O8 | 14.395 | [M−H]− | 359.0772 | 359.0753 | −5.3 | 197, 179, 135 | MPB |
| 15 | trans p-Coumaric acid 4-glucoside | C15H18O8 | 15.213 | [M−H]− | 325.0929 | 325.0926 | −0.9 | 163, 119 | SBG, LA, MPB |
| 16 | 3-Caffeoylquinic acid | C16H18O9 | 15.691 | [M−H]− | 353.0878 | 353.0875 | −0.8 | 191, 179, 161, 135 | R, SBG, LA, MPB |
| 17 | Caffeic acid | C9H8O4 | 16.614 | [M−H]− | 179.0350 | 179.0343 | −3.9 | 135 | SBG, MPB, R, LA |
| 18 | Cinnamoyl glucose | C15H18O7 | 16.808 | [M−H]− | 309.0979 | 309.0979 | 0.0 | 147 | R |
| 19 | 1-O-Sinapoyl-ꞵ-D-glucose | C17H22O10 | 16.945 | [M−H]− | 385.1140 | 385.1139 | −0.3 | 223 | LA, MPB |
| 20 | Cinnamic acid | C9H8O2 | 17.423 | [M−H]− | 147.0451 | 147.0458 | 4.8 | 103 | SBG, MPB |
| 21 | 3-p-Coumaroylquinic acid | C16H18O8 | 18.518 | [M−H]− | 337.0929 | 337.0918 | −3.3 | 191, 119 | MPB, LA, R |
| 22 | 3-Feruloylquinic acid | C17H20O9 | 19.200 | [M−H]− | 367.1034 | 367.1029 | −1.4 | 191 | R, LA |
| 23 | 3-Sinapoylquinic acid | C18H22O10 | 21.997 | [M−H]− | 397.1140 | 397.1141 | 0.3 | 223, 191 | LA, R |
| 24 | p-Coumaroyl tartaric acid | C13H12O8 | 22.056 | [M−H]− | 295.0459 | 295.0464 | 1.7 | 115 | LA |
| 25 | p-Coumaric acid | C9H8O3 | 22.309 | [M−H]− | 163.0400 | 163.0400 | 0.0 | 119 | R, LA, MPB |
| 26 | 1,2-Disinapoylgentiobiose | C34H42O19 | 23.851 | [M−H]− | 753.2247 | 753.2247 | 0.0 | 223, 207 | SBG, LA |
| 27 | Sinapic acid | C11H12O5 | 24.062 | [M−H]− | 223.0612 | 223.0618 | 2.7 | 193, 179, 149, 134 | LA |
| 28 | Feruloyl glucose | C16H20O9 | 26.100 | [M−H]− | 355.1034 | 355.1038 | 1.1 | 223, 207 | LA, MPB |
| 29 | 1,2-Diferuloylgentiobiose | C32H38O17 | 26.359 | [M−H]− | 693.2036 | 693.2042 | 0.9 | 193, 134 | LA, MPB, SBG |
| 30 | 1,5-Dicaffeoylquinic acid | C25H24O12 | 26.770 | [M−H]− | 515.1195 | 515.1197 | 0.4 | 191, 179, 135 | R, SBG |
| 31 | p-Coumaroyl glycolic acid | C11H10O5 | 60.679 | [M+H]+ | 223.0601 | 223.0605 | 1.8 | 205, 147, 119 | R, MPB |
| Flavonoids | |||||||||
| Anthocyanins | |||||||||
| 32 | Delphinidin 3-O-sambubioside | C26H29O16 | 11.988 | [M]+ | 597.1456 | 597.1471 | 2.5 | 303 | R, MBP |
| 33 | Cyanidin 3-sambubioside | C26H29O15 | 13.177 | [M]+ | 581.1506 | 581.1526 | 3.4 | 287 | R |
| 34 | Cyanidin | C15H11O6 | 13.926 | [M]+ | 287.0556 | 287.0522 | −11.8 | 231, 139, 69 | MPB, R |
| 35 | Cyanidin 3-rutinoside | C27H31O15 | 13.621 | [M]+ | 595.1663 | 595.1660 | −0.5 | 287 | MPB |
| 36 | Cyanidin 3-O-glucoside | C21H21O11 | 14.461 | [M]+ | 449.1084 | 449.0994 | 20.4 | 287 | MPB, R |
| 37 | Peonidin 3-O-(6″-p-coumaroyl-glucoside) | C31H29O13 | 16.378 | [M]+ | 609.1608 | 609.1617 | 1.5 | 301 | MPB |
| 38 | Delphinidin 3-rutinoside | C27H31O16 | 20.460 | [M]+ | 611.1612 | 611.1623 | 1.8 | 449, 303 | R |
| 39 | Delphinidin 3-galatoside | C21H21O12 | 20.460 | [M]+ | 465.1033 | 465.1033 | 0.0 | 303 | R |
| 40 | Delphinidin 3-O-(6″-p-coumaroyl-glucoside) | C30H27O14 | 20.528 | [M]+ | 611.1401 | 611.1430 | 4.7 | 303 | MPB, R |
| 41 | Delphinidin | C15H11O7 | 20.528 | [M]+ | 303.0505 | 303.0495 | −3.3 | 303 | MPB, R |
| Flavanols | |||||||||
| 42 | Theaflavin 3-O-gallate | C36H28O16 | 4.172 | [M+H]+ | 717.1450 | 717.1418 | −4.5 | 699, 565, 139 | SBG |
| 43 | Prodelphinidin trimer GC-GC-C | C45H38O20 | 6.333 | [M−H]− | 897.1883 | 897.1906 | 2.6 | 879, 305, 289, 125 | SBG, R |
| 44 | (−)-Epigallocatechin | C15H14O7 | 12.207 | [M−H]− | 305.0667 | 305.0650 | −5.6 | 289, 245, 179 | R |
| 45 | 4′,4″-Dimethylepigallocatechin gallate | C24H22O11 | 13.524 | [M−H]− | 485.1089 | 485.1092 | 0.6 | 441, 319, 183, 139 | SBG |
| 46 | (−)-Epicatechin | C15H14O6 | 15.19 | [M−H]− | 289.0717 | 289.0711 | −2.1 | 245, 205 | SBG, R, MPB, |
| 47 | Cinnamtannin A2 | C60H50O24 | 17.559 | [M−H]− | 1153.2619 | 1153.2602 | −1.5 | 1135, 577, 289, 125 | SBG |
| 48 | Catechin 3′-glucoside | C21H24O11 | 20.08 | [M−H]− | 451.1246 | 451.1253 | 1.6 | 289, 245 | LA, MPB |
| Flavanones | |||||||||
| 49 | Naringin 6′-malonate | C30H34O17 | 3.858 | [M−H]− | 665.1723 | 665.1701 | −3.3 | 579 | SBG |
| 50 | 6″-Acetylliquiritin | C23H24O10 | 6.288 | [M−H]− | 459.1297 | 459.1313 | 3.5 | 441, 255 | SBG, R |
| 51 | Narirutin 4′-O-glucoside | C33H42O19 | 20.803 | [M−H]− | 741.2247 | 741.2269 | 3.0 | 579, 271 | LA, SBG |
| 52 | Hesperetin 5-glucoside | C22H24O11 | 23.183 | [M−H]− | 463.1246 | 463.1252 | 1.3 | 301 | LA |
| 53 | Hesperetin 3′-O-glucuronide | C22H22O12 | 23.249 | [M−H]− | 477.1038 | 477.1052 | 2.9 | 301 | SBG, LA |
| 54 | 6-Geranylnaringenin | C25H28O5 | 23.288 | [M−H]− | 407.1864 | 407.1864 | 0.0 | 287, 271 | SBG |
| 55 | Naringenin-7-O-glucoside | C21H22O10 | 25.833 | [M−H]− | 433.1135 | 433.1154 | 4.4 | 301, 271, 151, 119 | MPB, SBG |
| 56 | Eriodictyol-7-O-glucoside | C21H22O11 | 27.312 | [M−H]− | 449.1084 | 449.1046 | −8.5 | 287, 151 | SBG |
| 57 | Hesperetin | C16H14O6 | 27.569 | [M−H]− | 301.0717 | 301.0719 | 0.7 | 265, 221, 177, 137 | LA, SBG, MPB |
| 58 | Hesperetin 5′,7-O-diglucuronide | C28H30O18 | 37.143 | [M−H]− | 653.1359 | 653.1341 | −2.8 | 301 | LA |
| 59 | Eriodictyol | C15H12O6 | 37.692 | [M−H]− | 287.0556 | 287.0572 | 5.6 | 151, 135 | SBG |
| 60 | Naringenin | C15H12O5 | 43.436 | [M−H]− | 271.0607 | 271.0623 | 5.9 | 227, 151, 119, 107 | SBG |
| 61 | 8-Prenylnaringenin | C20H20O5 | 49.532 | [M−H]− | 339.1238 | 339.1230 | −2.4 | 221, 147 | MPB |
| 62 | 5,7-Dihydroxyflavanone | C15H12O6 | 52.05 | [M−H]− | 255.0658 | 255.0671 | 5.1 | 213, 151 | SBG |
| 63 | Hesperidin | C28H34O15 | 53.498 | [M+H]+ | 611.1971 | 611.1966 | −0.8 | 303 | SBG |
| 64 | 3′,4′,5′-Trimethoxyflavone | C18H16O5 | 56.326 | [M−H]− | 311.0920 | 311.0890 | −9.6 | 296, 267 | SBG |
| Flavones and isoflavones | |||||||||
| 65 | 3′-O-Methylmaysin | C28H30O14 | 3.942 | [M−H]− | 589.1563 | 589.1571 | 1.4 | 589 | R, LA |
| 66 | Tetramethylscutellarein | C19H18O6 | 5.213 | [M−H]− | 341.103 | 341.1030 | 0.0 | 341 | R, MPB |
| 67 | Velutin | C17H14O6 | 6.265 | [M−H]− | 313.0717 | 313.0713 | −1.3 | 313 | SBG |
| 68 | Diosmin | C28H32O15 | 16.891 | [M−H]− | 607.1668 | 607.1669 | 0.2 | 301 | MPB, LA |
| 69 | Azaleatin 3-arabinoside | C21H20O11 | 21.170 | [M−H]− | 447.0928 | 447.0903 | −5.6 | 299, 269 | MBP |
| 70 | Syringetin-3-O-glucoside | C23H24O13 | 24.105 | [M−H]− | 507.1144 | 507.1160 | 3.2 | 345 | LA |
| 71 | Luteolin | C15H10O6 | 28.520 | [M−H]− | 285.0404 | 285.0423 | 6.7 | 267, 175, 133, 107 | LA, MBP, R |
| 72 | Biochanin A 7-O-glucoside | C22H22O10 | 31.319 | [M−H]− | 445.1135 | 445.1156 | 4.7 | 283 | SBG |
| 73 | Apigenin 6-C-glucoside | C21H20O10 | 32.237 | [M−H]− | 431.0983 | 431.0967 | -3.7 | 271 | MPB |
| 74 | Apigenin | C15H10O5 | 38.457 | [M−H]− | 269.0450 | 269.0467 | 6.3 | 225, 149 | MPB |
| 75 | Chrysoeriol 7-O-glucoside | C22H22O11 | 40.140 | [M−H]− | 461.1089 | 461.1068 | −4.6 | 299 | MPB |
| 76 | Diosmetin | C16H12O6 | 40.251 | [M−H]− | 299.0561 | 299.0567 | 2.0 | 284, 265, 133 | LA, MPB |
| 77 | Wogonin | C16H12O5 | 51.656 | [M−H]− | 283.0607 | 283.0587 | −7.1 | 268 | MPB |
| 78 | Glycitein | C16H12O5 | 52.368 | [M−H]− | 283.0607 | 283.0617 | 3.5 | 268 | MPB |
| 79 | Chrysin | C15H10O4 | 52.451 | [M−H]− | 253.0501 | 253.0515 | 5.5 | 253 | SBG |
| Flavonols and dihydroflavonols | |||||||||
| 80 | 6-Hydroxykaempferol 3,6-diglucoside 7-glucuronide | C33H38O23 | 14.082 | [M−H]− | 801.1726 | 801.1826 | 12.5 | 447, 285 | MBP |
| 81 | Limocitrin | C17H14O8 | 17.162 | [M−H]− | 345.0616 | 345.0604 | −3.5 | 330, 315, 301, 181 | LA |
| 82 | Myricetin 3-O-glucoside | C21H20O13 | 19.041 | [M−H]− | 479.0831 | 479.0816 | −3.1 | 317 | R |
| 83 | Quercetin 3-(2-galloylglucoside) | C28H24O16 | 20.371 | [M−H]− | 615.0986 | 615.0936 | −8.1 | 301, 169 | SBG |
| 84 | * Rutin | C27H30O16 | 20.530 | [M−H]− | 609.1461 | 609.1443 | −3.0 | 301, 300, 271, 255 | MPB, R |
| 85 | Myricetin 3-O-rhamnoside (myricitrin) | C21H20O12 | 21.328 | [M−H]− | 463.0882 | 463.0849 | −7.1 | 317 | SBG, R |
| 86 | Kaempferol 3-rutinoside | C27H30O15 | 24.911 | [M−H]− | 593.1507 | 593.1511 | 0.7 | 285, 151 | MPB |
| 87 | Kaempferol 3-O-arabinoside | C20H18O10 | 25.902 | [M−H]− | 417.0822 | 417.0793 | −7.0 | 285 | MPB |
| 88 | Quercetin 3-rhamnoside (quercitrin) | C21H20O11 | 26.142 | [M−H]− | 447.0928 | 447.0941 | 2.9 | 301 | SBG |
| 89 | Dihydroquercetin 3-O-rhamnoside | C21H22O11 | 26.305 | [M−H]− | 449.1089 | 449.1095 | 1.3 | 303 | SBG, MPB |
| 90 | Quercetin 3-O-arabinoside | C20H18O11 | 25.269 | [M−H]− | 433.0776 | 433.0769 | −1.6 | 301, 271, 151 | MPB |
| 91 | Isorhamnetin 3-O-glucoside 7-O-rhamnoside | C28H32O16 | 26.574 | [M−H]− | 623.1617 | 623.1607 | −1.6 | 315 | R, LA |
| 92 | * Quercetin-3-O-glucoside | C21H20O12 | 27.258 | [M−H]− | 463.0882 | 463.0842 | −8.6 | 301, 271, 255, 151 | SBG, MBP, R |
| 93 | * Isorhamnetin | C16H12O7 | 29.384 | [M−H]− | 315.0510 | 315.0491 | −6.0 | 300, 271, 151 | LA, MPB, SBG |
| 94 | Kaempferol 3-O-glucoside | C21H20O11 | 30.214 | [M−H]− | 447.0928 | 447.0927 | −0.2 | 285, 255, 147 | MPB, R, LA |
| 95 | * Myricetin | C15H10O8 | 30.613 | [M−H]− | 317.0298 | 317.0314 | 3.3 | 179, 151 | SBG, MPB, R |
| 96 | * Taxifolin | C15H12O7 | 31.176 | [M−H]− | 303.0510 | 303.0505 | −1.7 | 217, 125 | LA, MPB |
| 97 | Isorhamnetin 3-O-glucuronide | C22H20O13 | 31.344 | [M−H]− | 491.0831 | 491.0819 | −2.4 | 315 | R |
| 98 | * Quercetin | C15H10O7 | 39.148 | [M−H]− | 301.0353 | 301.0352 | −0.3 | 271, 179, 151, 121 | SBG, LA, MPB, R |
| Chalcones | |||||||||
| 99 | Xanthohumol | C21H22O5 | 10.842 | [M−H]− | 353.1389 | 353.1399 | 2.8 | 295, 233 | SBG |
| 100 | Phloretin | C15H14O5 | 28.333 | [M−H]− | 273.0768 | 273.0780 | 4.3 | 167, 119 | SBG |
| 101 | Phloretin-2′-O-glucoside | C21H24O10 | 28.33 | [M−H]− | 435.1296 | 435.1303 | 1.5 | 273, 167 | SBG |
| Isoflavonoids | |||||||||
| 102 | Dihydroformononetin | C16H14O4 | 3.991 | [M−H]− | 269.0819 | 269.0816 | −1.1 | 253, 239, 223 | SBG, MPB |
| 103 | Equol 7-O-glucuronide | C21H22O9 | 6.310 | [M−H]− | 417.1191 | 417.1201 | 2.4 | 241 | SBG |
| 104 | 6″-O-Malonyldaidzin | C24H22O12 | 14.474 | [M−H]− | 501.1038 | 501.1027 | −2.2 | 415 | MPB, LA |
| 105 | 6″-O-Acetyldaidzin | C23H22O10 | 16.980 | [M−H]− | 457.1140 | 457.1138 | −0.4 | 415 | SBG, R |
| 106 | Daidzin 4′-O-glucuronide | C27H28O15 | 21.029 | [M−H]− | 591.1355 | 591.1353 | −0.3 | 415, 253 | LA, R, SBG |
| 107 | 3′,4′,7-Trihydroxyisoflavanone | C15H12O5 | 27.308 | [M−H]− | 271.0612 | 271.0611 | −0.4 | 239, 135, 121 | SBG |
| 108 | 3′-O-Methylviolanone | C18H18O6 | 27.762 | [M−H]− | 329.1030 | 329.1029 | −0.3 | 285, 163 | MPB, LA |
| 109 | Daidzein 7-O-glucuronide | C21H18O10 | 33.742 | [M−H]− | 429.0827 | 429.0807 | −4.7 | 253 | R |
| 110 | 3′-Hydroxymelanettin | C16H12O6 | 40.251 | [M−H]− | 299.0561 | 299.0567 | 2.0 | 284 | LA, MPB |
| 111 | 2′-Hydroxyformononetin | C16H12O5 | 52.124 | [M−H]− | 283.0612 | 283.0606 | −2.1 | 268 | SBG, MPB |
| Tannins | |||||||||
| 112 | Gallagic acid | C28H12O16 | 3.075 | [M−H]− | 603.0052 | 603.0041 | −1.8 | 587, 559, 549, 446 | R, MPB |
| 113 | 2-O-Galloylpunicalin | C41H26O26 | 6.333 | [M−H]− | 933.0639 | 933.0645 | 0.6 | 781, 169, 125 | SBG |
| 114 | Glucosyringic acid | C15H20O10 | 7.546 | [M−H]− | 359.0978 | 359.0914 | −17.8 | 315, 197, 153, 125 | MPB |
| 115 | Procyanidin trimer C1 | C45H38O18 | 16.230 | [M−H]− | 865.1985 | 865.2012 | 3.1 | 739, 713, 695 | SBG, R |
| 116 | Procyanidin B2 | C30H26O12 | 19.039 | [M−H]− | 577.1351 | 577.1323 | −4.9 | 451, 425, 289, 245 | MPB, LA, SBG, R |
| 117 | Punicafolin | C41H30O26 | 19.102 | [M−H]− | 937.0952 | 937.0966 | 1.5 | 169, 125 | SBG |
| 118 | Ellagic acid | C14H6O8 | 55.906 | [M−H]− | 300.9990 | 300.9988 | −0.7 | 284, 257 | LA, MBP, R |
| Stilbenes | |||||||||
| 119 | Piceatannol | C14H12O4 | 5.594 | [M−H]− | 243.0663 | 243.0653 | −4.1 | 225, 201 | SBG, MPB |
| 120 | Polydatin | C20H22O8 | 21.854 | [M−H]− | 389.1242 | 389.1245 | 0.8 | 227 | LA, MPB, SBG |
| 121 | Piceatannol 3-O-glucoside | C20H22O9 | 30.064 | [M−H]− | 405.1191 | 405.1188 | −0.7 | 243 | MPB |
| Lignans | |||||||||
| 122 | Sesamin | C20H18O6 | 4.879 | [M−H]− | 353.1030 | 353.1015 | −4.2 | 338, 163 | MPB |
| 123 | 2-Hydroxyenterolactone | C18H18O5 | 6.371 | [M−H]− | 313.1081 | 313.1091 | 3.2 | 255, 163 | LA, MPB |
| 124 | Silibinin | C25H22O10 | 16.794 | [M−H]− | 481.1140 | 481.1151 | 2.3 | 301, 179, 165, 151 | MPB |
| 125 | 7-Oxomatairesinol | C20H20O7 | 21.997 | [M−H]− | 371.1136 | 371.1138 | 0.5 | 358, 343, 328 | LA |
| 126 | Arctigenin | C21H24O6 | 23.288 | [M−H]− | 371.1500 | 371.1497 | −0.8 | 356, 312, 295 | SBG |
| 127 | Enterolactone | C18H18O4 | 47.739 | [M−H]− | 299.1288 | 299.1299 | 4.3 | 281, 187, 165 | MPB, SBG |
| 128 | 2-Hydroxyenterodiol | C18H22O5 | 53.013 | [M−H]− | 317.1394 | 317.1395 | 0.3 | 299, 287, 269, 257 | MPB |
| Other compounds | |||||||||
| 129 | Pyrogallol | C6H6O3 | 7.009 | [M−H]− | 125.0244 | 125.0242 | −1.6 | 107, 97, 79 | MPB |
| 130 | [6]-Gingerol | C17H26O4 | 12.249 | [M−H]− | 293.1758 | 293.1768 | 3.4 | 137 | SBG, MPB |
| 131 | Quinic acid | C7H12O6 | 4.189 | [M−H]− | 191.0561 | 191.0578 | 9.0 | 85 | MPB, SBG |
| 132 | 1,2,4,6-Tetragalloyl-ꞵ-D-glucopyranose | C34H28O22 | 19.144 | [M−H]− | 787.0999 | 787.0953 | −5.9 | 169, 125 | SBG |
| 133 | Umbelliferone | C9H6O3 | 19.396 | [M−H]− | 161.0244 | 161.0246 | 1.2 | 133 | MPB, R |
| 134 | 2-Hydroxybenzaldehyde | C7H6O2 | 20.620 | [M−H]− | 121.0269 | 121.0276 | 5.8 | 92, 77 | MPB |
| 135 | p-Coumaraldehyde | C9H8O2 | 29.139 | [M−H]− | 147.0451 | 147.0463 | 8.0 | 119 | MPB |
| 136 | Xanthotoxol | C11H6O4 | 47.927 | [M−H]− | 201.0193 | 201.0191 | −1.0 | 171 | MPB, LA, SBG |
| 137 | Carnosic acid | C20H28O4 | 55.899 | [M−H]− | 331.1915 | 331.1927 | 3.6 | 287 | SBG, R |
| 138 | Mellein | C10H10O3 | 62.141 | [M+H]+ | 179.0703 | 179.0694 | −5.0 | 135 | LA |
| Limonoids | |||||||||
| 139 | Limonin | C26H30O8 | 19.039 | [M−H]− | 469.1868 | 469.1859 | −1.9 | 229 | LA |
| 140 | Obacunoic acid | C26H32O8 | 25.201 | [M−H]− | 471.2024 | 471.2027 | 0.6 | 471 | LA, MPB, SBG |
| 141 | Nomilin | C28H34O9 | 51.280 | [M+H]+ | 515.2276 | 515.2280 | 0.8 | 515 | MPB, SBG |
| 142 | Obacunone | C26H30O7 | 19.253 | [M−H]− | 455.2065 | 455.2065 | 0.0 | 407, 163 | SBG, R, MPB, LA |
| 143 | Citrusin | C28H34O11 | 55.330 | [M+H]+ | 547.2174 | 547.2162 | −2.2 | 547 | LA |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ali, A.; Cottrell, J.J.; Dunshea, F.R. Antioxidant, Alpha-Glucosidase Inhibition Activities, In Silico Molecular Docking and Pharmacokinetics Study of Phenolic Compounds from Native Australian Fruits and Spices. Antioxidants 2023, 12, 254. https://doi.org/10.3390/antiox12020254
Ali A, Cottrell JJ, Dunshea FR. Antioxidant, Alpha-Glucosidase Inhibition Activities, In Silico Molecular Docking and Pharmacokinetics Study of Phenolic Compounds from Native Australian Fruits and Spices. Antioxidants. 2023; 12(2):254. https://doi.org/10.3390/antiox12020254
Chicago/Turabian StyleAli, Akhtar, Jeremy J. Cottrell, and Frank R. Dunshea. 2023. "Antioxidant, Alpha-Glucosidase Inhibition Activities, In Silico Molecular Docking and Pharmacokinetics Study of Phenolic Compounds from Native Australian Fruits and Spices" Antioxidants 12, no. 2: 254. https://doi.org/10.3390/antiox12020254
APA StyleAli, A., Cottrell, J. J., & Dunshea, F. R. (2023). Antioxidant, Alpha-Glucosidase Inhibition Activities, In Silico Molecular Docking and Pharmacokinetics Study of Phenolic Compounds from Native Australian Fruits and Spices. Antioxidants, 12(2), 254. https://doi.org/10.3390/antiox12020254