


Antioxidant Potential-Rich Betel Leaves (Piper betle L.) Exert Depigmenting Action by Triggering Autophagy and Downregulating MITF/Tyrosinase In Vitro and In Vivo



Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Materials and Extraction
2.2. Gas-Chromatography Mass Spectrometric Analysis
2.3. Antioxidant Activities
2.4. Cell Culture and Cell Viability Assay
2.5. Effect of PBLE on Mushroom, Mice, and Human Tyrosinase Activity
2.6. Effect of PBLE on Melanin Content
2.7. Zymography for Tyrosinase Activity Measurement
2.8. The Concentration of cAMP Determination
2.9. Animal Experiments
2.10. Antioxidant Enzyme Assays
2.11. Measurement of Lipid Peroxidation (LPO)
2.12. Preparation of Cell Lysates and Western Blotting
2.13. Statistical Analysis
3. Results
3.1. Gas Chromatography–Mass Spectrometry (GC–MS) Analysis of PBLE
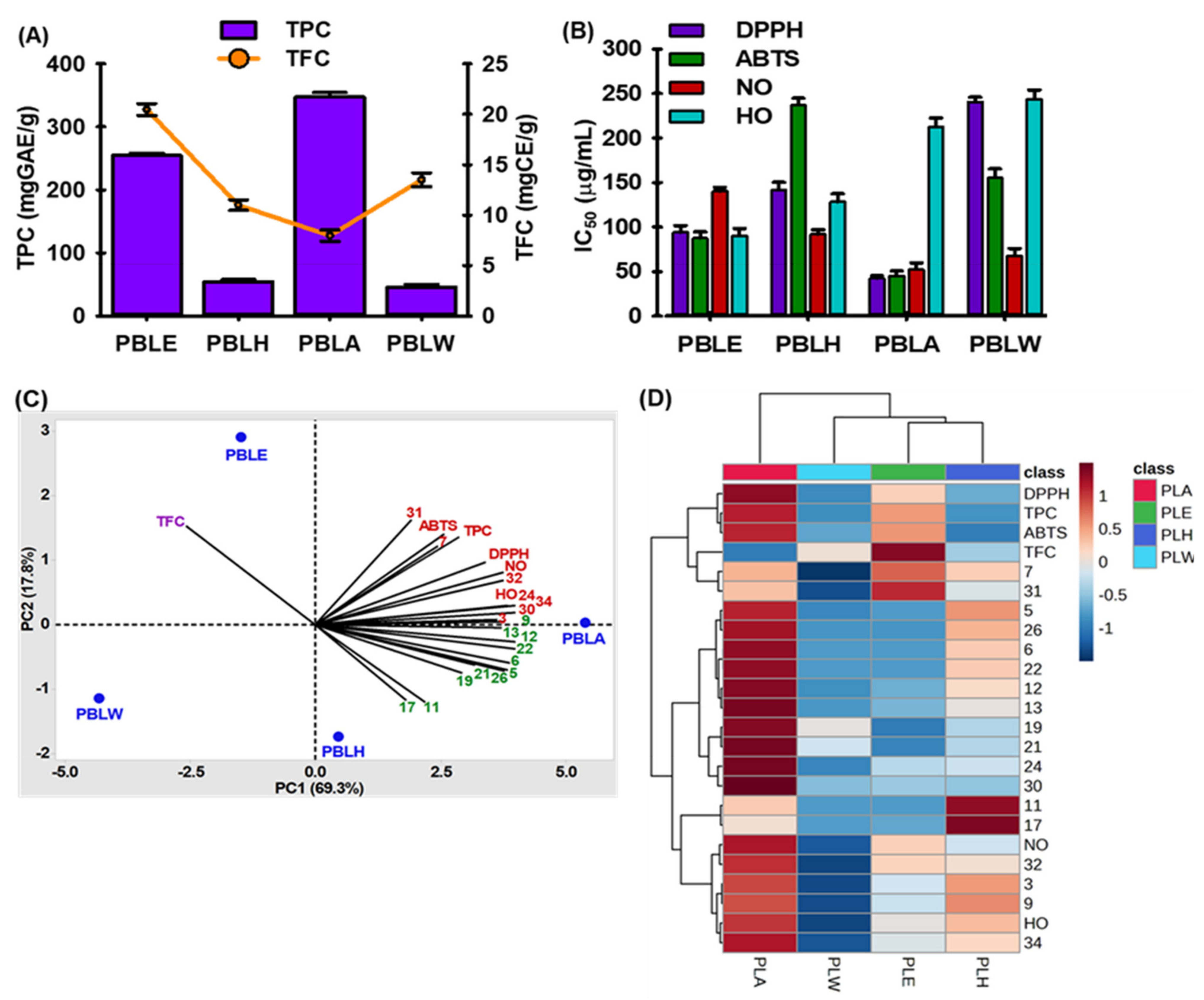
3.2. Total Phenol, Total Flavonoid, and Antioxidant Activities of PBL
3.3. Chemometric Analysis of PBL Extract/Fractions
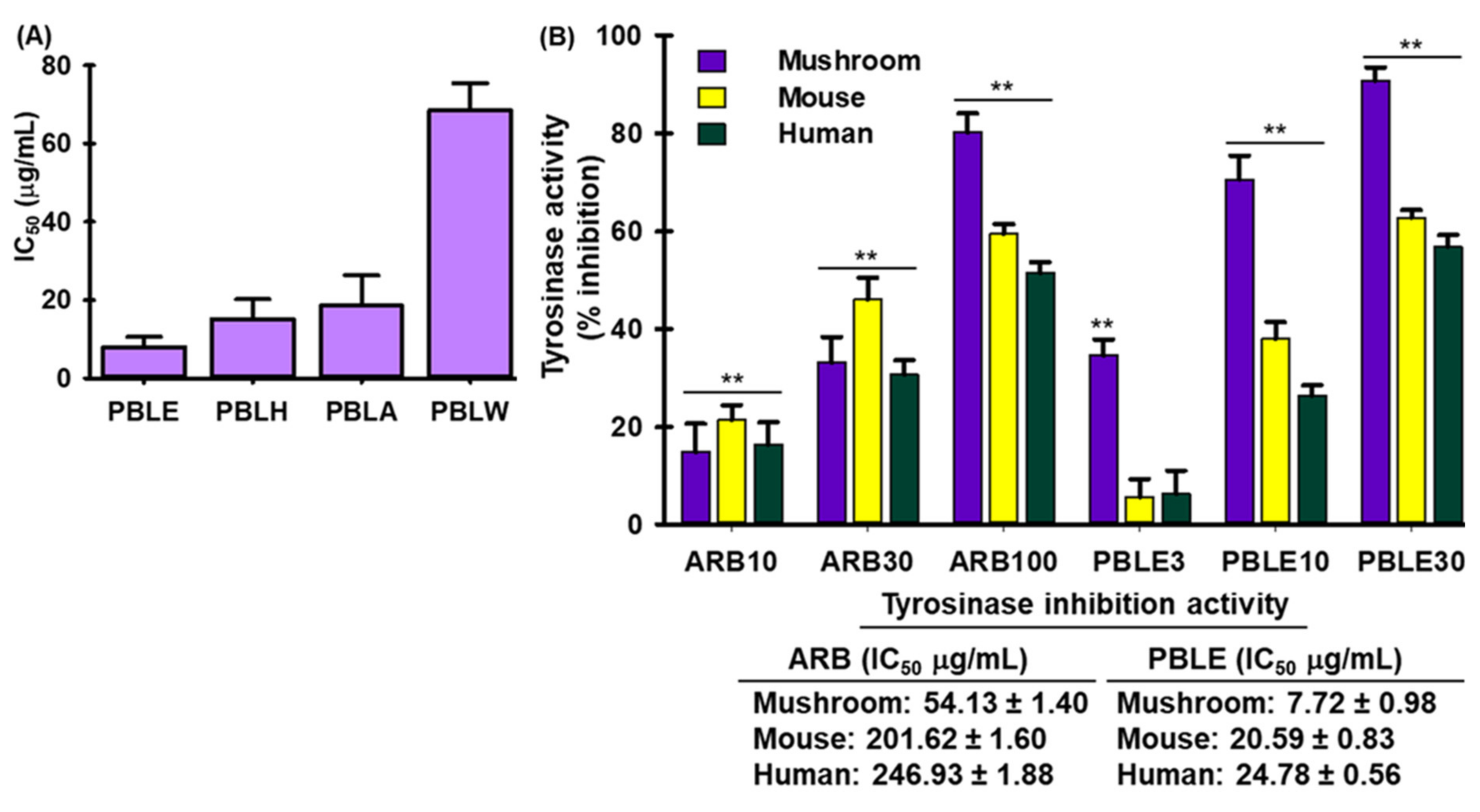
3.4. Effect of PBLE on Tyrosinase (Tyr) Activity
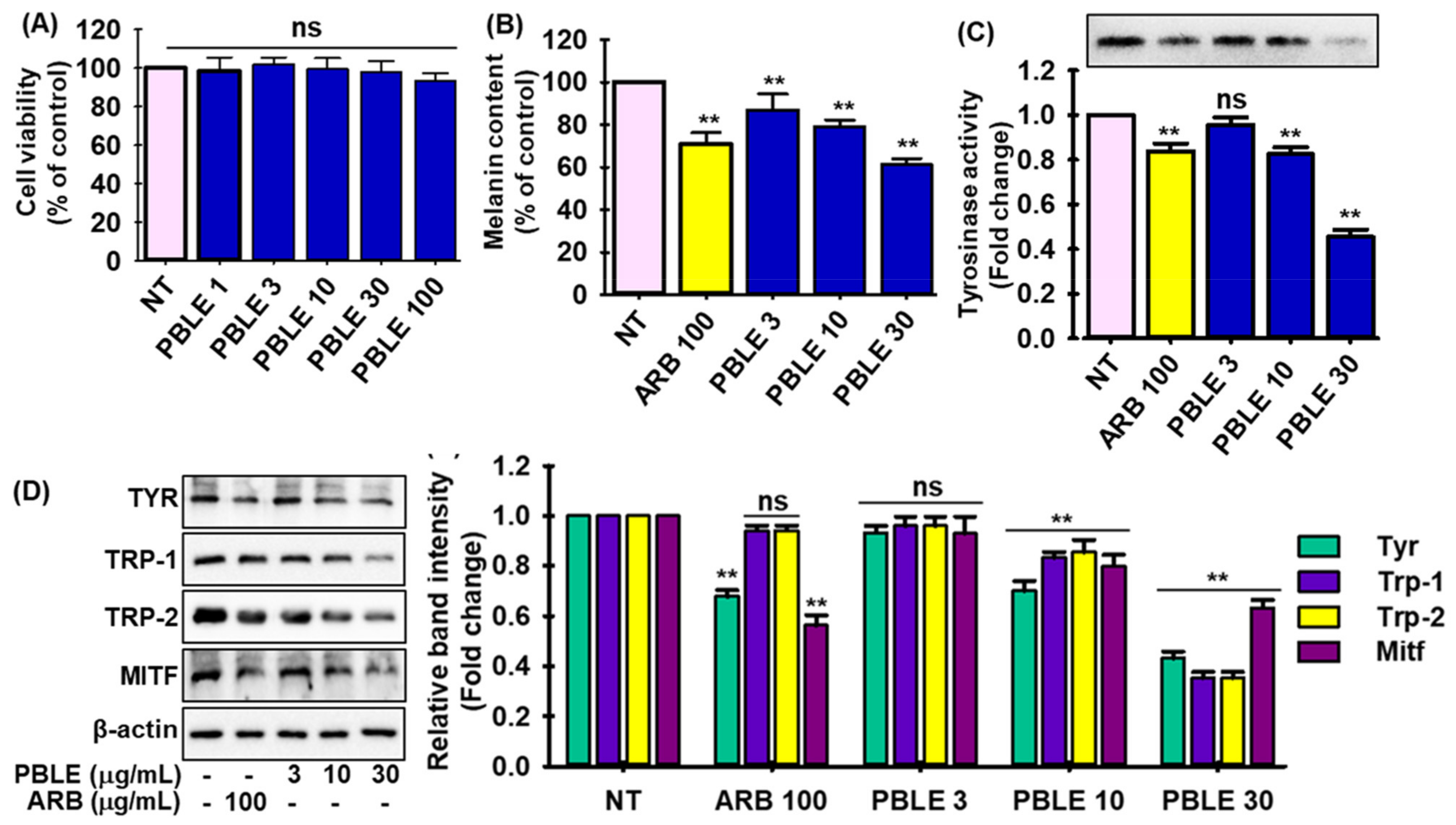
3.5. Depigmenting Effect of PBLE on Both Hyperpigmented Melanocyte Cells
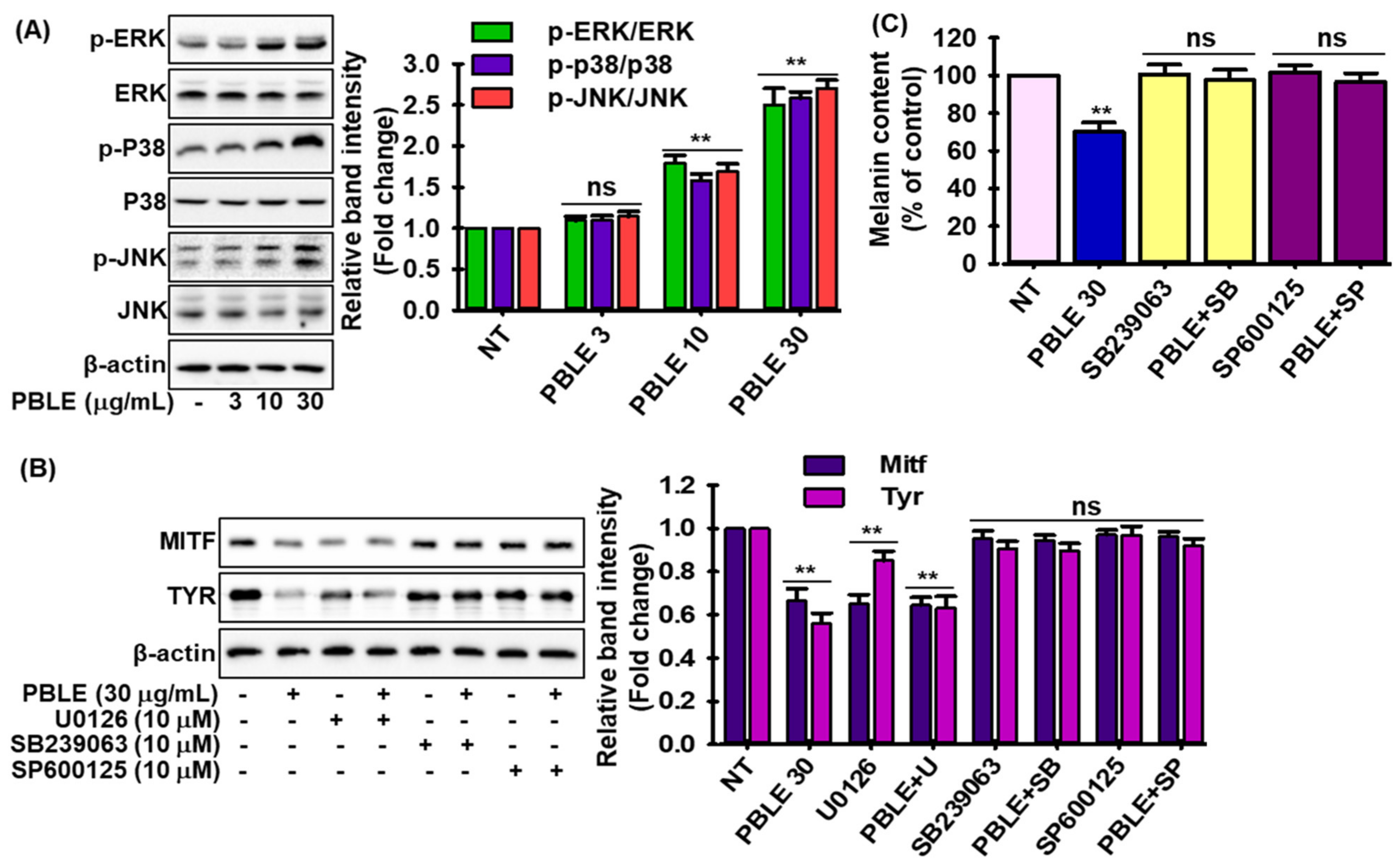
3.6. Effect of PBLE on the Melanogenesis-Related Signaling
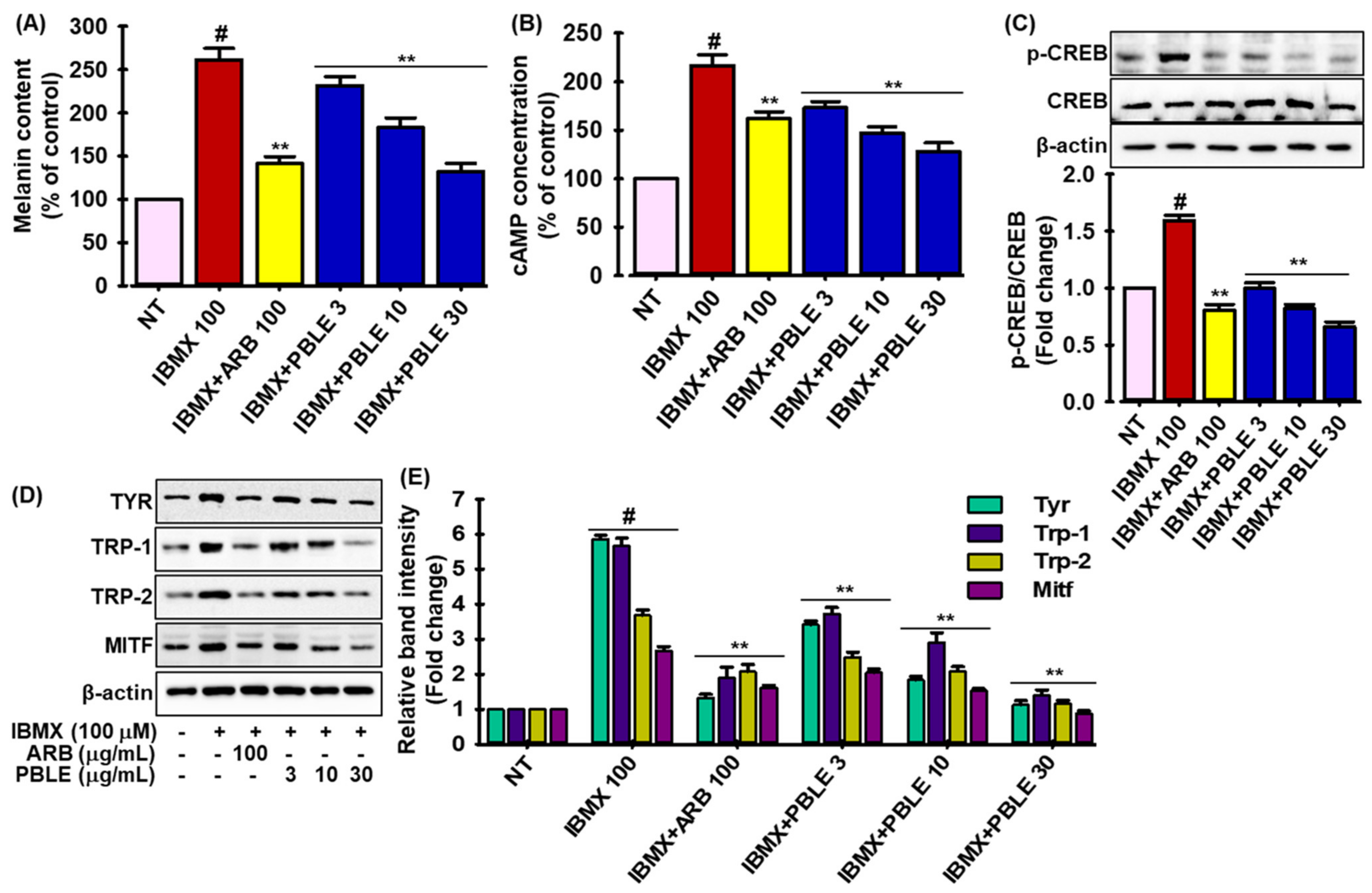
3.7. Effects of PBLE on IBMX (Isobutylmethylxanthine)-Induced Melanogenesis
3.8. Regulation of Autophagy by PBLE
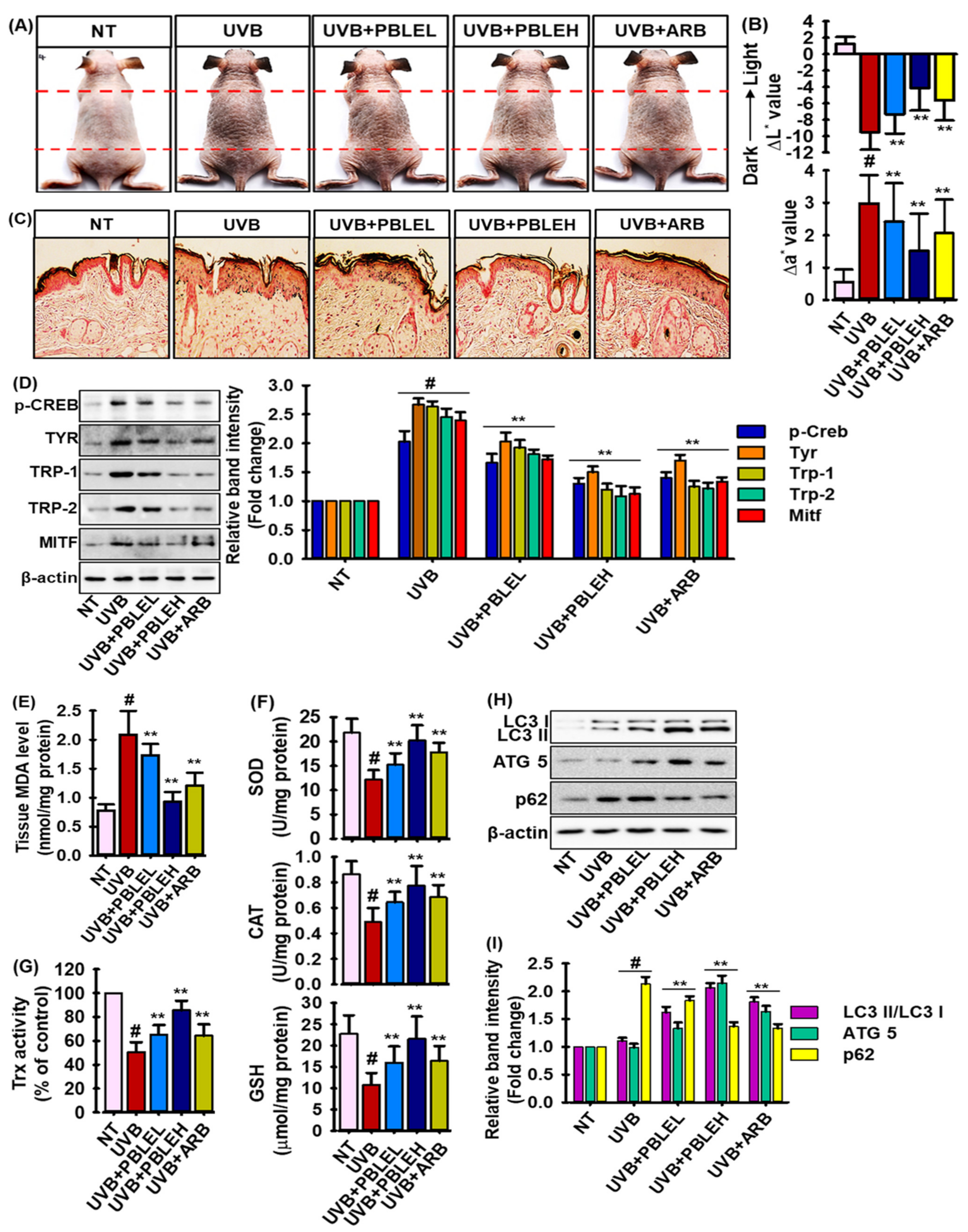
3.9. Depigmenting Effects of PBLE on UVB-Induced Pigmented HRM-2 Hairless Mice
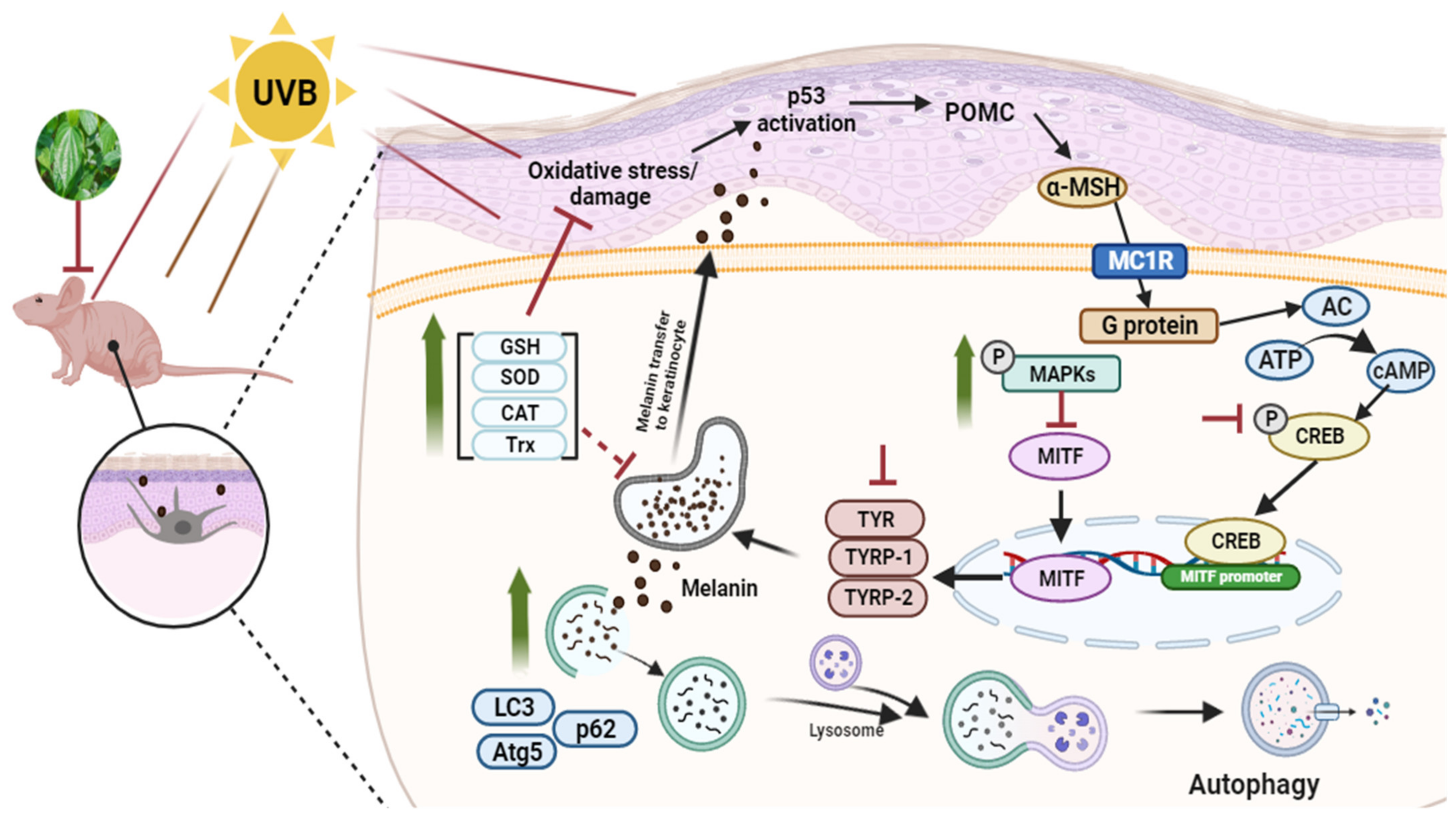
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- D’Mello, S.A.; Finlay, G.J.; Baguley, B.C.; Askarian-Amiri, M.E. Signaling pathways in melanogenesis. Int. J. Mol. Sci. 2016, 17, 1144. [Google Scholar] [CrossRef] [PubMed]
- Wang, N.; Hebert, D.N. Tyrosinase maturation through the mammalian secretory pathway: Bringing color to life. Pigment. Cell Res. 2006, 19, 3–18. [Google Scholar] [CrossRef] [PubMed]
- Ebanks, J.P.; Wickett, R.R.; Boissy, R.E. Mechanisms regulating skin pigmentation: The rise and fall of complexion coloration. Int. J. Mol. Sci. 2009, 10, 4066–4087. [Google Scholar] [CrossRef] [PubMed]
- Alam, M.B.; Ahmed, A.; Motin, M.A.; Kim, S.; Lee, S.H. Attenuation of melanogenesis by Nymphaea nouchali (Burm. f) flower extract through the regulation of cAMP/CREB/MAPKs/MITF and proteasomal degradation of tyrosinase. Sci. Rep. 2018, 8, 13928. [Google Scholar] [CrossRef]
- Miyamura, Y.; Coelho, S.G.; Wolber, R.; Miller, S.A.; Wakamatsu, K.; Zmudzka, B.Z.; Ito, S.; Smuda, C.; Passeron, T.; Choi, W. Regulation of human skin pigmentation and responses to ultraviolet radiation. Pigment. Cell Res. 2007, 20, 2–13. [Google Scholar] [CrossRef]
- Kim, Y.J.; Yokozawa, T. Modulation of oxidative stress and melanogenesis by proanthocyanidins. Biol. Pharm. Bull. 2009, 32, 1155–1159. [Google Scholar] [CrossRef]
- Jiménez-Cervantes, C.; Martínez-Esparza, M.A.; Pérez, C.; Daum, N.; Solano, F.; García-Borrón, J.C. Inhibition of melanogenesis in response to oxidative stress: Transient downregulation of melanocyte differentiation markers and possible involvement of microphthalmia transcription factor. J. Cell Sci. 2001, 114, 2335–2344. [Google Scholar] [CrossRef]
- Kamiński, K.; Kazimierczak, U.; Kolenda, T. Oxidative stress in melanogenesis and melanoma development. Contemp. Oncol. 2022, 26, 1–7. [Google Scholar] [CrossRef]
- Kondo, T.; Hearing, V.J. Update on the regulation of mammalian melanocyte function and skin pigmentation. Expert Rev. Dermatol. 2011, 6, 97–108. [Google Scholar] [CrossRef]
- Okamoto, K. Organellophagy: Eliminating cellular building blocks via selective autophagy. J. Cell Biol. 2014, 205, 435–445. [Google Scholar] [CrossRef] [Green Version]
- Stolz, A.; Ernst, A.; Dikic, I. Cargo recognition and trafficking in selective autophagy. Nat. Cell Biol. 2014, 16, 495–501. [Google Scholar] [CrossRef] [PubMed]
- Yang, Z.; Klionsky, D.J. Mammalian autophagy: Core molecular machinery and signaling regulation. Curr. Opin. Cell Biol. 2010, 22, 124–131. [Google Scholar] [CrossRef] [PubMed]
- Ganesan, A.K.; Ho, H.; Bodemann, B.; Petersen, S.; Aruri, J.; Koshy, S.; Richardson, Z.; Le, L.Q.; Krasieva, T.; Roth, M.G. Genome-wide siRNA-based functional genomics of pigmentation identifies novel genes and pathways that impact melanogenesis in human cells. PLoS Genet. 2008, 4, e1000298. [Google Scholar] [CrossRef] [PubMed]
- Ho, H.; Ganesan, A.K. The pleiotropic roles of autophagy regulators in melanogenesis. Pigment. Cell Melanoma Res. 2011, 24, 595–604. [Google Scholar] [CrossRef]
- Kumar, N.; Misra, P.; Dube, A.; Bhattacharya, S.; Dikshit, M.; Ranade, S. Piper betle Linn. a maligned Pan-Asiatic plant with an array of pharmacological activities and prospects for drug discovery. Curr. Sci. 2010, 99, 922–932. [Google Scholar]
- Pradhan, D.; Suri, K.; Pradhan, D.; Biswasroy, P. Golden heart of the nature: Piper betle L. J. Pharmacogn. Phytochem. 2013, 1, 147–167. [Google Scholar]
- Choudhary, D.; Kale, R.K. Antioxidant and non-toxic properties of Piper betle leaf extract: In vitro and in vivo studies. Phytotherapy Research: An International Journal Devoted to Pharmacological and Toxicological Evaluation of Natural Product Derivatives. Phytother. Res. 2002, 16, 461–466. [Google Scholar] [CrossRef]
- Nouri, L.; Nafchi, A.M.; Karim, A. Phytochemical, antioxidant, antibacterial, and α-amylase inhibitory properties of different extracts from betel leaves. Ind. Crops Prod. 2014, 62, 47–52. [Google Scholar] [CrossRef]
- Majumdar, B.; Chaudhuri, S.G.R.; Ray, A.; Bandyopadhyay, S.K. Effect of ethanol extract of Piper betle Linn leaf on healing of NSAID–induced experimental ulcer-A novel role of free radical scavenging action. Indian J. Exp. Biol. 2003, 41, 311–315. [Google Scholar]
- Parmar, V.S.; Jain, S.C.; Gupta, S.; Talwar, S.; Rajwanshi, V.K.; Kumar, R.; Azim, A.; Malhotra, S.; Kumar, N.; Jain, R. Polyphenols and alkaloids from Piper species. Phytochemistry 1998, 49, 1069–1078. [Google Scholar] [CrossRef]
- Saravanan, R.; Prakasam, A.; Ramesh, B.; Pugalendi, K. Influence of Piper betle on hepatic marker enzymes and tissue antioxidant status in ethanol-treated Wistar rats. J. Med. Food 2002, 5, 197–204. [Google Scholar] [CrossRef] [PubMed]
- Arambewela, L.; Arawwawala, M.; Rajapaksa, D. Piper betle: A potential natural antioxidant. Int. J. Food Sci. Technol. 2006, 41, 10–14. [Google Scholar] [CrossRef]
- Atiya, A.; Sinha, B.N.; Ranjan Lal, U. New chemical constituents from the Piper betle Linn.(Piperaceae). Nat. Prod. Res. 2018, 32, 1080–1087. [Google Scholar] [CrossRef] [PubMed]
- Madhumita, M.; Guha, P.; Nag, A. Bio-actives of betel leaf (Piper betle L.): A comprehensive review on extraction, isolation, characterization, and biological activity. Phytother. Res. 2020, 34, 2609–2627. [Google Scholar] [CrossRef] [PubMed]
- Alam, M.B.; Ju, M.K.; Lee, S.H. DNA Protecting Activities of Nymphaea nouchali (Burm. f) Flower Extract Attenuate t-BHP-Induced Oxidative Stress Cell Death through Nrf2-Mediated Induction of Heme Oxygenase-1 Expression by Activating MAP-Kinases. Int. J. Mol. Sci. 2017, 18, 2069. [Google Scholar] [CrossRef]
- Alam, M.B.; Bajpai, V.K.; Lee, J.; Zhao, P.; Byeon, J.H.; Ra, J.S.; Majumder, R.; Lee, J.S.; Yoon, J.I.; Rather, I.A.; et al. Inhibition of melanogenesis by jineol from Scolopendra subspinipes mutilans via MAP-Kinase mediated MITF downregulation and the proteasomal degradation of tyrosinase. Sci. Rep. 2017, 7, 45858. [Google Scholar] [CrossRef]
- Górny, M.; Bilska-Wilkosz, A.; Iciek, M.; Hereta, M.; Kamińska, K.; Kamińska, A.; Chwatko, G.; Rogóż, Z.; Lorenc-Koci, E. Alterations in the Antioxidant Enzyme Activities in the Neurodevelopmental Rat Model of Schizophrenia Induced by Glutathione Deficiency during Early Postnatal Life. Antioxidants 2020, 9, 538. [Google Scholar] [CrossRef]
- Tagrida, M.; Benjakul, S. Ethanolic extract of Betel (Piper betle L.) and Chaphlu (Piper sarmentosum Roxb.) dechlorophyllized using sedimentation process: Production, characteristics, and antioxidant activities. J. Food Biochem. 2020, 44, e13508. [Google Scholar] [CrossRef]
- Khorasani Esmaeili, A.; Mat Taha, R.; Mohajer, S.; Banisalam, B. Antioxidant activity and total phenolic and flavonoid content of various solvent extracts from in vivo and in vitro grown Trifolium pratense L. (Red Clover). BioMed Res. Int. 2015, 2015, 643285. [Google Scholar] [CrossRef]
- Yogeswari, S.; Bindu, K.H.; Kamalraj, S.; Ashokkumar, V.; Jayabaskaran, C. Antidiabetic, Antithrombin and Cytotoxic bioactive compounds in five cultivars of Piper betle L. Environ. Technol. Innov. 2020, 20, 101140. [Google Scholar] [CrossRef]
- Cabanes, J.; García-Cánovas, F.; Lozano, J.; Garcia-Carmona, F. A kinetic study of the melanization pathway between L-tyrosine and dopachrome. Biochim. Et Biophys. Acta (BBA)-Gen. Subj. 1987, 923, 187–195. [Google Scholar] [CrossRef]
- Jung, H.J.; Bang, E.; Kim, B.M.; Jeong, S.H.; Lee, G.H.; Chung, H.Y. Loganin Inhibits α-MSH and IBMX-induced Melanogenesis by Suppressing the Expression of Tyrosinase in B16F10 Melanoma Cells. J. Life Sci. 2019, 29, 1200–1207. [Google Scholar]
- Lee, K.W.; Kim, M.; Lee, S.H.; Kim, K.D. The Function of Autophagy as a Regulator of Melanin Homeostasis. Cells 2022, 11, 2085. [Google Scholar] [CrossRef]
- Lu, Y.; Tonissen, K.F.; Di Trapani, G. Modulating skin colour: Role of the thioredoxin and glutathione systems in regulating melanogenesis. Biosci. Rep. 2021, 41, BSR20210427. [Google Scholar] [CrossRef]
- García-Gavín, J.; González-Vilas, D.; Fernández-Redondo, V.; Toribio, J. Pigmented contact dermatitis due to kojic acid. A paradoxical side effect of a skin lightener. Contact Dermat. 2010, 62, 63–64. [Google Scholar] [CrossRef]
- Liu, W.-y.; Zou, C.-m.; Hu, J.-h.; Xu, Z.-j.; Si, L.-q.; Liu, J.-j.; Huang, J.-g. Kinetic Characterization of Tyrosinase-catalyzed Oxidation of Four Polyphenols. Curr. Med. Sci. 2020, 40, 239–248. [Google Scholar] [CrossRef]
- Cho, S.J.; Kwon, H.S. Tyrosinase Inhibitory Activities of Safrole from Myristica fragrans Houtt. J. Appl. Biol. Chem. 2015, 58, 295–301. [Google Scholar] [CrossRef]
- Lim, J.Y.; Ishiguro, K.; Kubo, I. Tyrosinase inhibitory p-coumaric acid from ginseng leaves. Phytother. Res. PTR 1999, 13, 371–375. [Google Scholar] [CrossRef]
- Ha, J.H.; Park, S.N. Mechanism underlying inhibitory effect of six dicaffeoylquinic acid isomers on melanogenesis and the computational molecular modeling studies. Bioorganic Med. Chem. 2018, 26, 4201–4208. [Google Scholar] [CrossRef]
- Sato, K.; Toriyama, M. Depigmenting effect of catechins. Molecules 2009, 14, 4425–4432. [Google Scholar] [CrossRef]
- Choi, M.-H.; Shin, H.-J. Anti-melanogenesis effect of quercetin. Cosmetics 2016, 3, 18. [Google Scholar] [CrossRef]
- Eom, Y.S.; Jeong, D.; Ryu, A.; Song, K.-H.; Im, D.S.; Lee, M.-Y. Daphne odora Exerts Depigmenting Effects via Inhibiting CREB/MITF and Activating AKT/ERK-Signaling Pathways. Curr. Issues Mol. Biol. 2022, 44, 3312–3323. [Google Scholar] [CrossRef]
- Orhan, I.E.; Deniz, F.S.S. Inhibition of Melanogenesis by Some Well-Known Polyphenolics: A Review. Curr. Pharm. Biotechnol. 2021, 22, 1412–1423. [Google Scholar] [CrossRef]
- Baek, S.h.; Lee, S.H. Sesamol decreases melanin biosynthesis in melanocyte cells and zebrafish: Possible involvement of MITF via the intracellular cAMP and p38/JNK signalling pathways. Exp. Dermatol. 2015, 24, 761–766. [Google Scholar] [CrossRef] [Green Version]
- Baek, S.H.; Nam, I.J.; Kwak, H.S.; Kim, K.C.; Lee, S.H. Cellular Anti-Melanogenic Effects of a Euryale ferox Seed Extract Ethyl Acetate Fraction via the Lysosomal Degradation Machinery. Int. J. Mol. Sci. 2015, 16, 9217–9235. [Google Scholar] [CrossRef]
- Zhu, W.; Zhao, Z.; Cheng, B. The role of autophagy in skin pigmentation. Eur. J. Dermatol. EJD 2020, 30, 655–662. [Google Scholar] [CrossRef]
- Katsuyama, Y.; Taira, N.; Yoshioka, M.; Okano, Y.; Masaki, H. Disruption of melanosome transport in melanocytes treated with theophylline causes their degradation by autophagy. Biochem. Biophys Res. Commun. 2017, 485, 126–130. [Google Scholar] [CrossRef]
- Lee, K.W.; Ryu, H.W.; Oh, S.S.; Park, S.; Madhi, H.; Yoo, J.; Park, K.H.; Kim, K.D. Depigmentation of α-melanocyte-stimulating hormone-treated melanoma cells by β-mangostin is mediated by selective autophagy. Exp. Derm. 2017, 26, 585–591. [Google Scholar] [CrossRef]
- Kim, E.S.; Chang, H.; Choi, H.; Shin, J.H.; Park, S.J.; Jo, Y.K.; Choi, E.S.; Baek, S.Y.; Kim, B.G.; Chang, J.W.; et al. Autophagy induced by resveratrol suppresses α-MSH-induced melanogenesis. Exp. Derm. 2014, 23, 204–206. [Google Scholar] [CrossRef]
- Kim, E.S.; Shin, J.H.; Seok, S.H.; Kim, J.B.; Chang, H.; Park, S.J.; Jo, Y.K.; Choi, E.S.; Park, J.S.; Yeom, M.H.; et al. Autophagy mediates anti-melanogenic activity of 3′-ODI in B16F1 melanoma cells. Biochem. Biophys. Res. Commun. 2013, 442, 165–170. [Google Scholar] [CrossRef]
- Kim, E.S.; Jo, Y.K.; Park, S.J.; Chang, H.; Shin, J.H.; Choi, E.S.; Kim, J.B.; Seok, S.H.; Kim, J.S.; Oh, J.S.; et al. ARP101 inhibits α-MSH-stimulated melanogenesis by regulation of autophagy in melanocytes. FEBS Lett. 2013, 587, 3955–3960. [Google Scholar] [CrossRef] [PubMed]
- Ma, C.; Zhang, D.; Ma, Q.; Liu, Y.; Yang, Y. Arbutin inhibits inflammation and apoptosis by enhancing autophagy via SIRT1. Adv. Clin. Exp. Med. 2021, 30, 535–544. [Google Scholar] [CrossRef] [PubMed]
- Zhang, B.; Zeng, M.; Li, B.; Wang, Y.; Kan, Y.; Wang, S.; Meng, Y.; Gao, J.; Feng, W.-s.; Zheng, X. Inhibition of oxidative stress and autophagy by arbutin in lipopolysaccharide-induced myocardial injury. Pharmacogn. Mag. 2019, 15, 507. [Google Scholar] [CrossRef]
- Lv, L.; Zhang, J.; Tian, F.; Li, X.; Li, D.; Yu, X. Arbutin protects HK-2 cells against high glucose-induced apoptosis and autophagy by up-regulating microRNA-27a. Artif. Cells Nanomed. Biotechnol. 2019, 47, 2940–2947. [Google Scholar] [CrossRef] [Green Version]
- Peng, H.-Y.; Lin, C.-C.; Wang, H.-Y.; Shih, Y.; Chou, S.-T. The melanogenesis alteration effects of Achillea millefolium L. essential oil and linalyl acetate: Involvement of oxidative stress and the JNK and ERK signaling pathways in melanoma cells. PLoS ONE 2014, 9, e95186. [Google Scholar] [CrossRef]
- Li, R.; Peng, A.; He, C.; Wang, X.; Shi, J.; Chen, L.; Wei, Y. Analysis of triptophenolide and its related compounds from Tripterygium wilfordii Hook.f by electrospray ionization tandem mass spectrometry. Int. J. Mass Spectrom. 2008, 278, 38–49. [Google Scholar] [CrossRef]
- Delijewski, M.; Wrześniok, D.; Otręba, M.; Beberok, A.; Buszman, E. Nicotine impact on melanogenesis and antioxidant defense system in HEMn-DP melanocytes. Mol. Cell. Biochem. 2014, 395, 109–116. [Google Scholar] [CrossRef]
- Tang, H.; Du, H.; Kuang, X.; Huang, H.; Zeng, J.; Long, C.; Zhu, B.; Fu, L.; Wang, H.; Zhang, Q.; et al. Arbutin Protects Retinal Pigment Epithelium Against Oxidative Stress by Modulating SIRT1/FOXO3a/PGC-1α/β Pathway. Front. Genet. 2022, 13, 922807. [Google Scholar] [CrossRef]
- Avci, P.; Sadasivam, M.; Gupta, A.; De Melo, W.C.M.A.; Huang, Y.-Y.; Yin, R.; Chandran, R.; Kumar, R.; Otufowora, A.; Nyame, T.; et al. Animal models of skin disease for drug discovery. Expert Opin. Drug Discov. 2013, 8, 331–355. [Google Scholar] [CrossRef]
- Ali, N.; Hosseini, M.; Vainio, S.; Taïeb, A.; Cario-André, M.; Rezvani, H.R. Skin equivalents: Skin from reconstructions as models to study skin development and diseases. Br. J. Dermatol. 2015, 173, 391–403. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| No. | RT | CAS | Name | MF | MW | Area (%) | |||
|---|---|---|---|---|---|---|---|---|---|
| PBLE | PBLH | PBLA | PBLW | ||||||
| 1 | 22.43 | 000470-82-6 | Eucalyptol | C10H18O | 154.25 | 0.01 | 1.21 | 0.81 | Nd |
| 2 | 25.27 | 000694-74-6 | 6-Amino-6-methylfulvene | C7H9N | 107.15 | 0.01 | 0.51 | 1.01 | Nd |
| 3 | 40.47 | 003856-25-5 | α-Copaene | C15H24 | 204.35 | 0.95 | 1.52 | 1.84 | 0.01 |
| 4 | 42.11 | 005208-59-3 | β-Bourbonene | C15H24 | 204.35 | 0.40 | 0.51 | 0.74 | 0.03 |
| 5 | 44.00 | 000078-70-6 | Linalool | C10H18O | 154.25 | 0.13 | 1.13 | 1.53 | 0.03 |
| 6 | 45.15 | 013474-59-4 | trans-α-Bergamotene | C15H24 | 204.35 | 0.09 | 1.09 | 2.09 | 0.09 |
| 7 | 46.81 | 000087-44-5 | trans-Caryophyllene | C15H24 | 204.35 | 1.80 | 1.35 | 1.48 | 0.03 |
| 8 | 49.65 | 025246-27-9 | Allo-aromadendrene | C15H24 | 204.35 | 0.04 | 2.14 | Nd | Nd |
| 9 | 50.99 | 006753-98-6 | α-Humulene | C15H24 | 204.35 | 0.83 | 1.40 | 1.59 | 0.07 |
| 10 | 51.87 | 028973-97-9 | cis-β-Farnesene | C15H24 | 204.35 | 0.08 | 2.08 | 1.48 | Nd |
| 11 | 52.59 | 178737-43-4 | cis-4,10-epoxy-Amorphane | C15H26O | 222.37 | 0.03 | 2.03 | 1.03 | 0.03 |
| 12 | 53.23 | 023986-74-5 | Germacrene D | C15H24 | 204.35 | 0.19 | 0.59 | 1.19 | 0.09 |
| 13 | 53.53 | 157477-72-0 | cis-Muurola-diene | C15H24 | 204.35 | 0.12 | 0.42 | 1.12 | 0.02 |
| 14 | 53.64 | 030021-74-0 | γ-Muurolene | C15H24 | 204.35 | 0.03 | 0.93 | 1.03 | Nd |
| 15 | 54.11 | 031983-22-9 | α-Muurolene | C15H24 | 204.35 | 0.48 | 0.02 | 1.05 | Nd |
| 16 | 54.24 | 000495-61-4 | β-Bisabolene | C15H24 | 204.35 | 0.26 | 1.26 | 0.86 | Nd |
| 17 | 57.27 | 029837-12-5 | Cubenene | C15H24 | 204.35 | 0.11 | 2.81 | 1.11 | 0.01 |
| 18 | 60.95 | 000127-41-3 | α-Ionone | C13H20O | 192.30 | 0.03 | 3.13 | 2.93 | Nd |
| 19 | 62.31 | 000100-51-6 | Benzyl alcohol | C7H8O | 108.14 | 0.02 | 1.32 | 4.62 | 1.92 |
| 20 | 63.47 | 001198-37-4 | 2,4-Dimethylquinoline | C11H11N | 157.21 | 0.03 | Nd | 1.03 | 0.03 |
| 21 | 64.13 | 000060-12-8 | 2-Phenylethanol | C8H10O | 122.07 | 0.02 | 0.82 | 3.12 | 1.02 |
| 22 | 64.30 | 021391-99-1 | α-Calacorene | C15H20 | 200.32 | 0.06 | 0.56 | 1.06 | 0.06 |
| 23 | 65.52 | 000079-77-6 | trans-β-Ionone | C13H20O | 192.30 | 0.02 | 3.02 | 2.32 | Nd |
| 24 | 69.12 | 000093-15-2 | Methyl eugenol | C11H14O2 | 178.23 | 0.73 | 0.84 | 2.54 | 0.08 |
| 25 | 69.38 | 002153-66-4 | Santrolina triene | C10H16 | 136.24 | 0.04 | 0.04 | Nd | Nd |
| 26 | 71.48 | 020129-39-9 | α-Corocalene | C15H20 | 200.32 | 0.06 | 1.26 | 2.06 | 0.06 |
| 27 | 72.15 | 000102-76-1 | Triacetin | C9H14O6 | 218.20 | 0.09 | 0.89 | 0.19 | 0.09 |
| 28 | 72.33 | 022567-17-5 | γ-Gurjunene | C15H24 | 204.35 | 0.02 | 0.12 | 0.08 | 0.02 |
| 29 | 74.56 | 077171-55-2 | (-)-Spathulenol | C15H24O | 220.35 | 0.51 | 0.83 | 0.82 | 0.02 |
| 30 | 76.39 | 000097-53-0 | Eugenol | C10H12O2 | 164.20 | 0.71 | 0.52 | 9.73 | 0.27 |
| 31 | 77.68 | 000501-19-9 | O-Eugenol | C10H12O2 | 164.20 | 51.47 | 26.01 | 35.25 | 0.30 |
| 32 | 80.62 | 001941-09-9 | O-Eugenol acetate | C12H14O3 | 206.24 | 17.37 | 16.06 | 26.72 | 0.20 |
| 33 | 82.56 | 092691-77-5 | Gossonorol | C15H22O | 218.33 | 0.06 | 0.76 | 0.06 | Nd |
| 34 | 83.94 | 000501-92-8 | Chavicol | C9H10O | 134.17 | 2.01 | 2.44 | 3.96 | 0.25 |
| 35 | 84.17 | 997146-73-2 | Dihydro actinidiolide | C11H16O2 | 180.24 | 0.04 | 0.02 | 0.04 | 0.04 |
| 36 | 90.04 | 000480-33-1 | (-)-Mellein | C10H10O3 | 178.18 | 0.05 | 0.15 | 0.05 | 0.05 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Alam, M.B.; Park, N.H.; Song, B.-R.; Lee, S.-H. Antioxidant Potential-Rich Betel Leaves (Piper betle L.) Exert Depigmenting Action by Triggering Autophagy and Downregulating MITF/Tyrosinase In Vitro and In Vivo. Antioxidants 2023, 12, 374. https://doi.org/10.3390/antiox12020374
Alam MB, Park NH, Song B-R, Lee S-H. Antioxidant Potential-Rich Betel Leaves (Piper betle L.) Exert Depigmenting Action by Triggering Autophagy and Downregulating MITF/Tyrosinase In Vitro and In Vivo. Antioxidants. 2023; 12(2):374. https://doi.org/10.3390/antiox12020374
Chicago/Turabian StyleAlam, Md Badrul, Na Hyun Park, Bo-Rim Song, and Sang-Han Lee. 2023. "Antioxidant Potential-Rich Betel Leaves (Piper betle L.) Exert Depigmenting Action by Triggering Autophagy and Downregulating MITF/Tyrosinase In Vitro and In Vivo" Antioxidants 12, no. 2: 374. https://doi.org/10.3390/antiox12020374
APA StyleAlam, M. B., Park, N. H., Song, B. -R., & Lee, S. -H. (2023). Antioxidant Potential-Rich Betel Leaves (Piper betle L.) Exert Depigmenting Action by Triggering Autophagy and Downregulating MITF/Tyrosinase In Vitro and In Vivo. Antioxidants, 12(2), 374. https://doi.org/10.3390/antiox12020374