
Targeting M2 Macrophages with a Novel NADPH Oxidase Inhibitor


 , ,
, ,  , and
, and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Reagents
2.2. Isolation of Blood Monocytes and Differentiation of Blood Monocytes by CSF1
2.3. Flow Cytometry Analysis of Human Macrophages
2.4. Cell Morphology
2.5. Phagocytosis
2.6. Cytokine Profile in Human Macrophages Supernatants
2.7. Confocal Imaging
2.7.1. Immunofluorescence of Living Cells
2.7.2. Experiment in Fixed Cells
2.8. Cell Viability Tests
2.9. Modeling
2.10. Animals and Experimental Protocols
2.11. Vascular Reactivity Studies on Pressure Myograph
3. Results
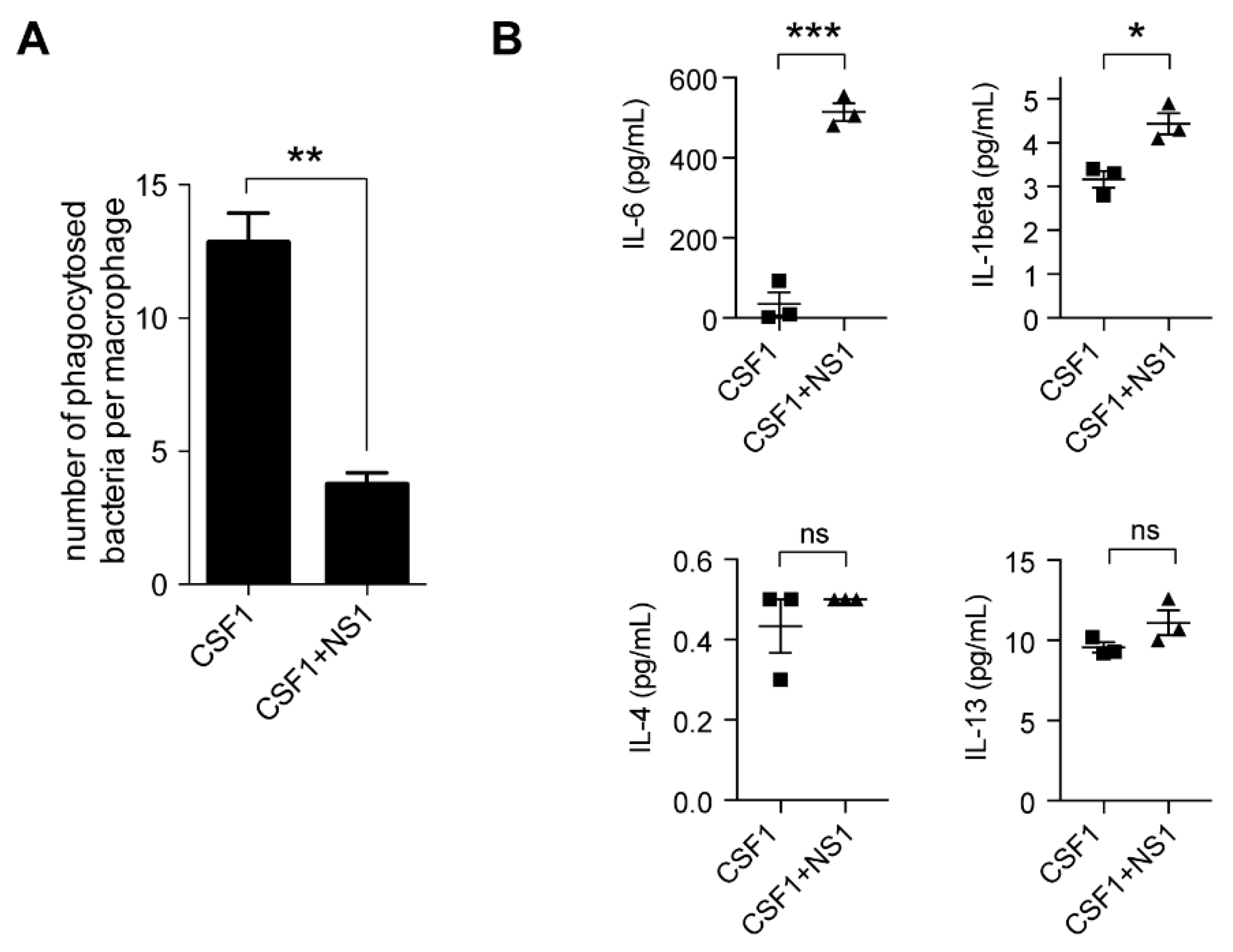
3.1. NS1 Inhibition of “M2” Macrophage Differentiation
3.2. Effect of NS1 on Vascular Tone and Metabolism of Smooth Muscle Cells
3.3. NADPH Sites of NOX2 and NOX4; Exploiting Their Variability to Design a NOX2-Specific Inhibitor
4. Discussion and Conclusions
4.1. NOX2 TAMs and NS1
4.2. NOX Inhibition in Cancer and NOX2 Inhibitors
4.3. NOX Inhibition and Hypoxia
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Qian, B.Z.; Pollard, J.W. Macrophage diversity enhances tumor progression and metastasis. Cell 2010, 141, 39–51. [Google Scholar] [CrossRef]
- Tariq, M.; Zhang, J.; Liang, G.; Ding, L.; He, Q.; Yang, B. Macrophage Polarization: Anti-Cancer Strategies to Target Tumor-Associated Macrophage in Breast Cancer. J. Cell. Biochem. 2017, 118, 2484–2501. [Google Scholar] [CrossRef] [PubMed]
- Ryder, M.; Ghossein, R.A.; Ricarte-Filho, J.C.; Knauf, J.A.; Fagin, J.A. Increased density of tumor-associated macrophages is associated with decreased survival in advanced thyroid cancer. Endocr. Relat. Cancer 2008, 15, 1069–1074. [Google Scholar] [CrossRef] [PubMed]
- Sionov, R.V.; Fridlender, Z.G.; Granot, Z. The Multifaceted Roles Neutrophils Play in the Tumor Microenvironment. Cancer Microenviron. 2015, 8, 125–158. [Google Scholar] [CrossRef] [PubMed]
- Griess, B.; Mir, S.; Datta, K.; Teoh-Fitzgerald, M. Scavenging reactive oxygen species selectively inhibits M2 macrophage polarization and their pro-tumorigenic function in part, via Stat3 suppression. Free Radic. Biol. Med. 2020, 147, 48–60. [Google Scholar] [CrossRef]
- Khou, S.; Popa, A.; Luci, C.; Bihl, F.; Meghraoui-Kheddar, A.; Bourdely, P.; Salavagione, E.; Cosson, E.; Rubod, A.; Cazareth, J.; et al. Tumor-Associated Neutrophils Dampen Adaptive Immunity and Promote Cutaneous Squamous Cell Carcinoma Development. Cancers 2020, 12, 1860. [Google Scholar] [CrossRef]
- Xu, Q.; Choksi, S.; Qu, J.; Jang, J.; Choe, M.; Banfi, B.; Engelhardt, J.F.; Liu, Z.G. NADPH Oxidases Are Essential for Macrophage Differentiation. J. Biol. Chem. 2016, 291, 20030–20041. [Google Scholar] [CrossRef]
- Curi, R.; de Siqueira Mendes, R.; de Campos Crispin, L.A.; Norata, G.D.; Sampaio, S.C.; Newsholme, P. A past and present overview of macrophage metabolism and functional outcomes. Clin. Sci. (Lond.) 2017, 131, 1329–1342. [Google Scholar] [CrossRef]
- Feron, O. Targeting the tumor vascular compartment to improve conventional cancer therapy. Trends Pharmacol. Sci. 2004, 25, 536–542. [Google Scholar] [CrossRef]
- Martinive, P.; De Wever, J.; Bouzin, C.; Baudelet, C.; Sonveaux, P.; Gregoire, V.; Gallez, B.; Feron, O. Reversal of temporal and spatial heterogeneities in tumor perfusion identifies the tumor vascular tone as a tunable variable to improve drug delivery. Mol. Cancer Ther. 2006, 5, 1620–1627. [Google Scholar] [CrossRef] [Green Version]
- Sonveaux, P.; Dessy, C.; Martinive, P.; Havaux, X.; Jordan, B.F.; Gallez, B.; Gregoire, V.; Balligand, J.L.; Feron, O. Endothelin-1 is a critical mediator of myogenic tone in tumor arterioles: Implications for cancer treatment. Cancer Res. 2004, 64, 3209–3214. [Google Scholar] [CrossRef]
- Manuel, R.; Lima, M.S.; Dilly, S.; Daunay, S.; Abbe, P.; Pramil, E.; Solier, S.; Guillaumond, F.; Tubiana, S.S.; Escargueil, A.; et al. Distinction between 2′- and 3′-Phosphate Isomers of a Fluorescent NADPH Analogue Led to Strong Inhibition of Cancer Cells Migration. Antioxidants 2021, 10, 723. [Google Scholar] [CrossRef] [PubMed]
- Rouaud, F.; Boucher, J.L.; Slama-Schwok, A.; Rocchi, S. Mechanism of melanoma cells selective apoptosis induced by a photoactive NADPH analogue. Oncotarget 2016, 7, 82804–82819. [Google Scholar] [CrossRef] [PubMed]
- Lejal, N.; Truchet, S.; Bechor, E.; Bouguyon, E.; Khedkar, V.; Bertho, N.; Vidic, J.; Adenot, P.; Solier, S.; Pick, E.; et al. Turning off NADPH oxidase-2 by impeding p67(phox) activation in infected mouse macrophages reduced viral entry and inflammation. Biochim. Biophys. Acta Gen. Subj. 2018, 1862, 1263–1275. [Google Scholar] [CrossRef] [PubMed]
- Rouaud, F.; Romero-Perez, M.; Wang, H.; Lobysheva, I.; Ramassamy, B.; Henry, E.; Tauc, P.; Giacchero, D.; Boucher, J.L.; Deprez, E.; et al. Regulation of NADPH-dependent Nitric Oxide and reactive oxygen species signalling in endothelial and melanoma cells by a photoactive NADPH analogue. Oncotarget 2014, 5, 10650–10664. [Google Scholar] [CrossRef]
- Liu, R.; Song, K.; Wu, J.-X.; Geng, X.-P.; Zheng, L.; Gao, X.; Peng, H.; Chen, L. Structure of human phagocyte NADPH oxidase in the resting state. eLife 2022, 11, e83743. [Google Scholar] [CrossRef]
- Jumper, J.; Evans, R.; Pritzel, A.; Green, T.; Figurnov, M.; Ronneberger, O.; Tunyasuvunakool, K.; Bates, R.; Zidek, A.; Potapenko, A.; et al. Highly accurate protein structure prediction with AlphaFold. Nature 2021, 596, 583–589. [Google Scholar] [CrossRef]
- Noreng, S.; Ota, N.; Sun, Y.; Ho, H.; Johnson, M.; Arthur, C.P.; Schneider, K.; Lehoux, I.; Davies, C.W.; Mortara, K.; et al. Structure of the core human NADPH oxidase NOX2. Nature Commun. 2022, 13, 6079. [Google Scholar] [CrossRef]
- Li, Y.; Wang, H.; Tarus, B.; Perez, M.R.; Morellato, L.; Henry, E.; Berka, V.; Tsai, A.L.; Ramassamy, B.; Dhimane, H.; et al. Rational design of a fluorescent NADPH derivative imaging constitutive nitric-oxide synthases upon two-photon excitation. Proc. Natl. Acad. Sci. USA 2012, 109, 12526–12531. [Google Scholar] [CrossRef]
- McWhorter, F.Y.; Wang, T.; Nguyen, P.; Chung, T.; Liu, W.F. Modulation of macrophage phenotype by cell shape. Proc. Natl. Acad. Sci. USA 2013, 110, 17253–17258. [Google Scholar] [CrossRef] [Green Version]
- Magnani, F.; Nenci, S.; Millana Fananas, E.; Ceccon, M.; Romero, E.; Fraaije, M.W.; Mattevi, A. Crystal structures and atomic model of NADPH oxidase. Proc. Natl. Acad. Sci. USA 2017, 114, 6764–6769. [Google Scholar] [CrossRef] [PubMed]
- Berendsen, H.J.; van der Spoel, D.; van Drunen, R. GROMACS: A message-passing parallel molecular dynamics implementation. Comput. Phys. Commun. 1995, 91, 43–56. [Google Scholar] [CrossRef]
- Dessy, C.; Saliez, J.; Ghisdal, P.; Daneau, G.; Lobysheva, I.; Frerart, F.; Belge, C.; Jnaoui, K.; Noirhomme, P.; Feron, O.; et al. Endothelial beta3-adrenoreceptors mediate nitric oxide-dependent vasorelaxation of coronary microvessels in response to the third-generation beta-blocker nebivolol. Circulation 2005, 112, 1198–1205. [Google Scholar] [CrossRef] [PubMed]
- Robles-Vera, I.; Visitacion, N.; Toral, M.; Sanchez, M.; Gomez-Guzman, M.; O’Valle, F.; Jimenez, R.; Duarte, J.; Romero, M. Toll-like receptor 7-driven lupus autoimmunity induces hypertension and vascular alterations in mice. J. Hypertens. 2020, 38, 1322–1335. [Google Scholar] [CrossRef]
- Solier, S.; Jacquel, A.; Martinou, J.C.; Pierron, G.; Rivière, J.; Dessen, P.; Dayris, T.; Hermine, O.; Dupuy, C.; Droin, N. A Caspase-7/NOX2 Axis Regulates the Migration of Monocytes in Response to Colony-Stimulating Factor-1. Available online: https://ssrn.com/abstract=3285805 (accessed on 16 November 2018).
- Beaumont, E.; Lambry, J.C.; Blanchard-Desce, M.; Martasek, P.; Panda, S.P.; van Faassen, E.E.; Brochon, J.C.; Deprez, E.; Slama-Schwok, A. NO formation by neuronal NO-synthase can be controlled by ultrafast electron injection from a nanotrigger. Chembiochem 2009, 10, 690–701. [Google Scholar] [CrossRef] [PubMed]
- Beaumont, E.; Lambry, J.C.; Gautier, C.; Robin, A.C.; Gmouh, S.; Berka, V.; Tsai, A.L.; Blanchard-Desce, M.; Slama-Schwok, A. Synchronous photoinitiation of endothelial NO synthase activity by a nanotrigger targeted at its NADPH site. J. Am. Chem. Soc. 2007, 129, 2178–2186. [Google Scholar] [CrossRef]
- Garcin, E.D.; Bruns, C.M.; Lloyd, S.J.; Hosfield, D.J.; Tiso, M.; Gachhui, R.; Stuehr, D.J.; Tainer, J.A.; Getzoff, E.D. Structural basis for isozyme-specific regulation of electron transfer in nitric-oxide synthase. J. Biol. Chem. 2004, 279, 37918–37927. [Google Scholar] [CrossRef]
- Lambry, J.C.; Beaumont, E.; Tarus, B.; Blanchard-Desce, M.; Slama-Schwok, A. Selective probing of a NADPH site controlled light-induced enzymatic catalysis. J. Mol. Recognit. 2010, 23, 379–388. [Google Scholar] [CrossRef]
- Sun, J.; Park, C.; Guenthner, N.; Gurley, S.; Zhang, L.; Lubben, B.; Adebayo, O.; Bash, H.; Chen, Y.; Maksimos, M.; et al. Tumor-associated macrophages in multiple myeloma: Advances in biology and therapy. J. Immunother. Cancer 2022, 10, e003975. [Google Scholar] [CrossRef]
- Mitchem, J.B.; Brennan, D.J.; Knolhoff, B.L.; Belt, B.A.; Zhu, Y.; Sanford, D.E.; Belaygorod, L.; Carpenter, D.; Collins, L.; Piwnica-Worms, D.; et al. Targeting tumor-infiltrating macrophages decreases tumor-initiating cells, relieves immunosuppression, and improves chemotherapeutic responses. Cancer Res. 2013, 73, 1128–1141. [Google Scholar] [CrossRef] [Green Version]
- Kim, E.-K.; Jang, M.; Song, M.-J.; Kim, D.; Kim, Y.; Jang, H.H. Redox-Mediated Mechanism of Chemoresistance in Cancer Cells. Antioxidants 2019, 8, 471. [Google Scholar] [CrossRef]
- de Paula Assis, L.H.; de Gabriel Dorighello, G.; De Oliveira, H.C. Pro-inflammatory polarization of macrophages is associated with reduced endoplasmic reticulum-mitochondria interaction. Biochem. Biophys. Res. Commun. 2022, 606, 661–667. [Google Scholar] [CrossRef]
- Laurindo, F.R.; Araujo, T.L.; Abrahão, T.B. Nox NADPH oxidases and the endoplasmic reticulum. Antioxid. Redox Signal. 2014, 20, 2755–2775. [Google Scholar] [CrossRef]
- Mukawera, E.; Chartier, S.; Williams, V.; Pagano, P.J.; Lapointe, R.; Grandvaux, N. Redox-modulating agents target NOX2-dependent IKKε oncogenic kinase expression and proliferation in human breast cancer cell lines. Redox Biol. 2015, 6, 9–18. [Google Scholar] [CrossRef] [PubMed]
- Pratt, S.J.P.; Lee, R.M.; Chang, K.T.; Hernández-Ochoa, E.O.; Annis, D.A.; Ory, E.C.; Thompson, K.N.; Bailey, P.C.; Mathias, T.J.; Ju, J.A.; et al. Mechanoactivation of NOX2-generated ROS elicits persistent TRPM8 Ca(2+) signals that are inhibited by oncogenic KRas. Proc. Natl. Acad. Sci. USA 2020, 117, 26008–26019. [Google Scholar] [CrossRef] [PubMed]
- Aydin, E.; Hallner, A.; Grauers Wiktorin, H.; Staffas, A.; Hellstrand, K.; Martner, A. NOX2 inhibition reduces oxidative stress and prolongs survival in murine KRAS-induced myeloproliferative disease. Oncogene 2019, 38, 1534–1543. [Google Scholar] [CrossRef] [PubMed]
- Konaté, M.M.; Antony, S.; Doroshow, J.H. Inhibiting the Activity of NADPH Oxidase in Cancer. Antioxid. Redox Signal. 2020, 33, 435–454. [Google Scholar] [CrossRef]
- Augsburger, F.; Filippova, A.; Rasti, D.; Seredenina, T.; Lam, M.; Maghzal, G.; Mahiout, Z.; Jansen-Dürr, P.; Knaus, U.G.; Doroshow, J.; et al. Pharmacological characterization of the seven human NOX isoforms and their inhibitors. Redox Biol. 2019, 26, 101272. [Google Scholar] [CrossRef]
- Lu, J.; Risbood, P.; Kane, C.T., Jr.; Hossain, M.T.; Anderson, L.; Hill, K.; Monks, A.; Wu, Y.; Antony, S.; Juhasz, A.; et al. Characterization of potent and selective iodonium-class inhibitors of NADPH oxidases. Biochem. Pharmacol. 2017, 143, 25–38. [Google Scholar] [CrossRef]
- Delprat, V.; Huart, C.; Feron, O.; Soncin, F.; Michiels, C. The impact of macrophages on endothelial cells is potentiated by cycling hypoxia: Enhanced tumor inflammation and metastasis. Front. Oncol. 2022, 12, 961753. [Google Scholar] [CrossRef]
- Michiels, C.; Tellier, C.; Feron, O. Cycling hypoxia: A key feature of the tumor microenvironment. Biochim. Biophys. Acta 2016, 1866, 76–86. [Google Scholar] [CrossRef] [PubMed]
- Shimokawa, H. Hydrogen peroxide as an endothelium-derived hyperpolarizing factor. Pflug. Arch. Eur. J. Physiol. 2010, 459, 915–922. [Google Scholar] [CrossRef] [PubMed]
- Bakalova, R.; Aoki, I.; Zhelev, Z.; Higashi, T. Cellular redox imbalance on the crossroad between mitochondrial dysfunction, senescence, and proliferation. Redox Biol. 2022, 53, 102337. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Compound | R1 | Chromophore | Protein | Interaction Energy Kcal/mol |
|---|---|---|---|---|
| NS1 | Ethyl |  | NOX2 | −74 |
| NS1 | Ethyl | NOX4 | −82 | |
| NS169 | Hook C5H9NH2 |  | NOX2 | −138 |
| NS169 | Hook | NOX4 | −75 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dilly, S.; Romero, M.; Solier, S.; Feron, O.; Dessy, C.; Slama Schwok, A. Targeting M2 Macrophages with a Novel NADPH Oxidase Inhibitor. Antioxidants 2023, 12, 440. https://doi.org/10.3390/antiox12020440
Dilly S, Romero M, Solier S, Feron O, Dessy C, Slama Schwok A. Targeting M2 Macrophages with a Novel NADPH Oxidase Inhibitor. Antioxidants. 2023; 12(2):440. https://doi.org/10.3390/antiox12020440
Chicago/Turabian StyleDilly, Sébastien, Miguel Romero, Stéphanie Solier, Olivier Feron, Chantal Dessy, and Anny Slama Schwok. 2023. "Targeting M2 Macrophages with a Novel NADPH Oxidase Inhibitor" Antioxidants 12, no. 2: 440. https://doi.org/10.3390/antiox12020440
APA StyleDilly, S., Romero, M., Solier, S., Feron, O., Dessy, C., & Slama Schwok, A. (2023). Targeting M2 Macrophages with a Novel NADPH Oxidase Inhibitor. Antioxidants, 12(2), 440. https://doi.org/10.3390/antiox12020440