
Increased SIRT1 Concentration Following Four Years of Selenium and Q10 Intervention Associated with Reduced Cardiovascular Mortality at 10-Year Follow-Up—Sub-Study of a Previous Prospective Double-Blind Placebo-Controlled Randomized Clinical Trial
 ,
,  ,
,  , and
, and
Abstract
:1. Introduction
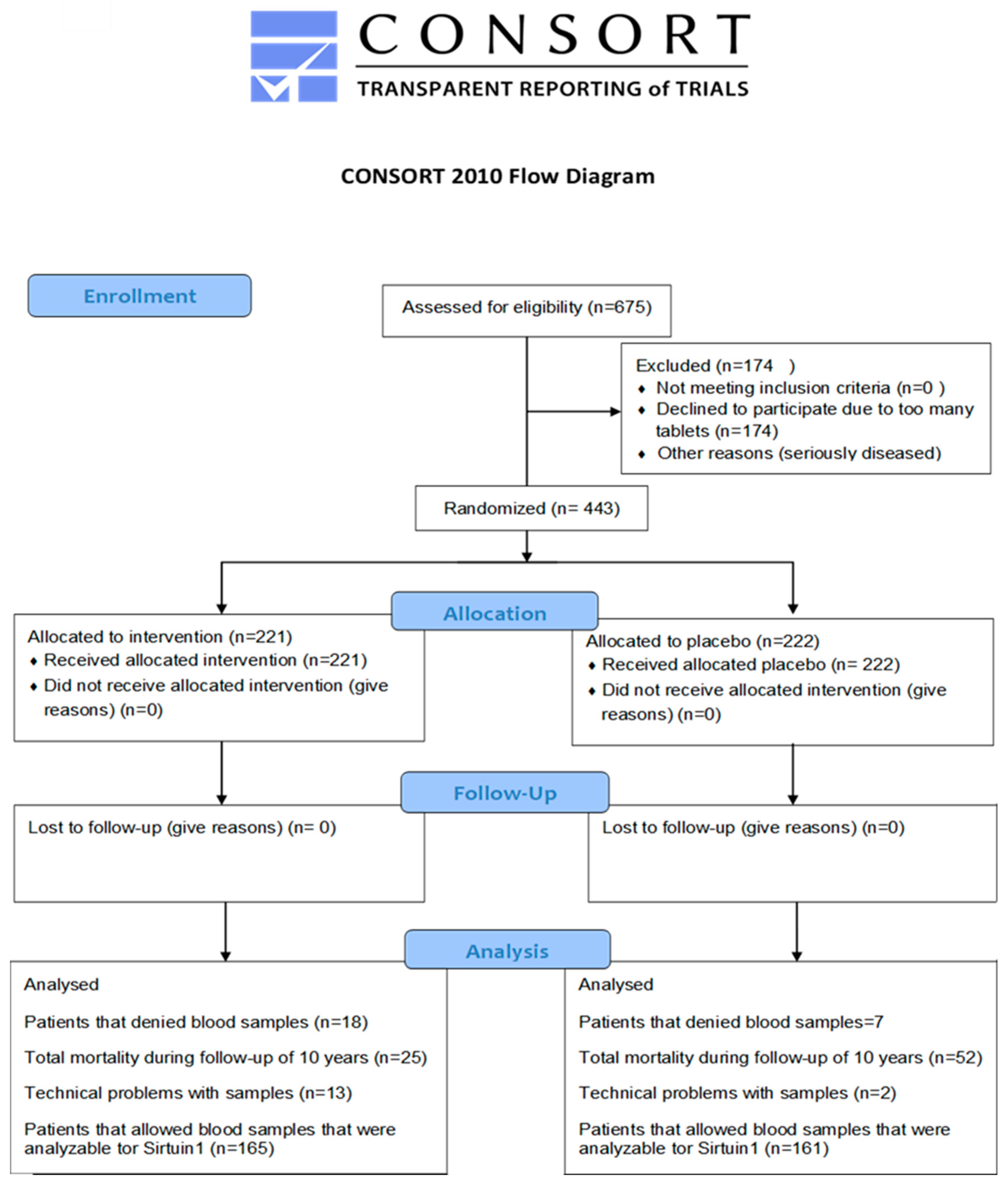
2. Materials and Methods
2.1. Study Population
2.2. Blood Sampling
2.3. SIRT1 Analysis
2.4. MicroRNA Analysis
2.5. Statistical Methods
3. Results
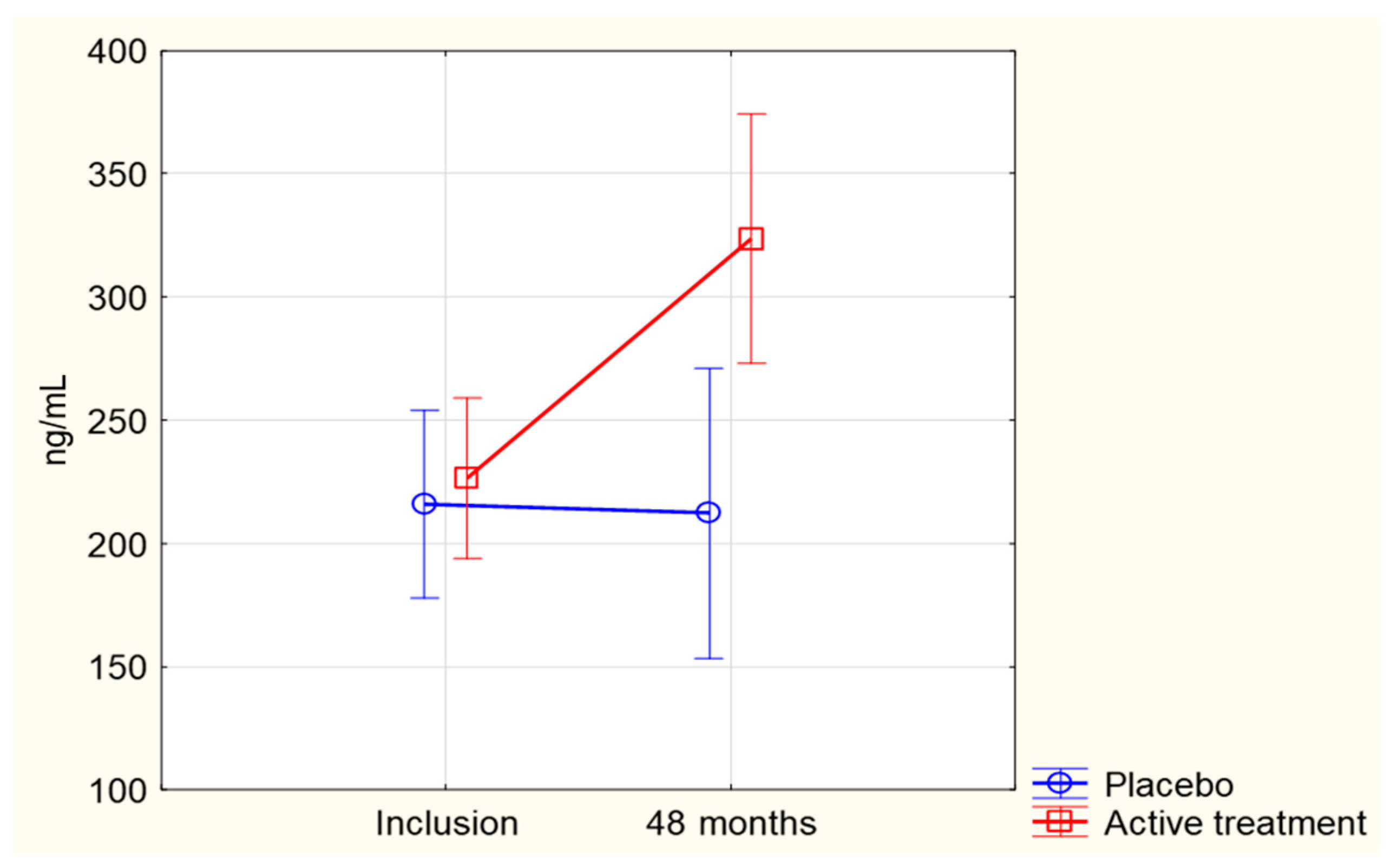
3.1. SIRT1 Concentration in Relation to the Se and Coenzyme Q10 Intervention
3.2. SIRT1 Changes as Related to CV Mortality
3.3. Association between Circulating SIRT1 and microRNAs at Baseline
4. Discussion
4.1. Effects of the Se/CoQ10 Intervention on SIRT1
4.2. Effects of SIRT1 in the CV System
4.3. Potential Mechanisms of Increased SIRT1 for Cardiac Protection
4.4. Regulation of SIRT1 by microRNAs
4.5. Limitations
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Rayman, M.P. The importance of selenium to human health. Lancet 2000, 356, 233–241. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alehagen, U.; Opstad, T.B.; Alexander, J.; Larsson, A.; Aaseth, J. Impact of Selenium on Biomarkers and Clinical Aspects Related to Ageing. A Review. Biomolecules 2021, 11, 1478. [Google Scholar] [CrossRef]
- Alehagen, U.; Alexander, J.; Aaseth, J. Supplementation with Selenium and Coenzyme Q10 Reduces Cardiovascular Mortality in Elderly with Low Selenium Status. A Secondary Analysis of a Randomised Clinical Trial. PLoS ONE 2016, 11, e0157541. [Google Scholar] [CrossRef] [Green Version]
- Alehagen, U.; Johansson, P.; Björnstedt, M.; Rosén, A.; Post, C.; Aaseth, J. Relatively high mortality risk in elderly Swedish subjects with low selenium status. Eur. J. Clin. Nutr. 2016, 70, 91–96. [Google Scholar] [CrossRef] [Green Version]
- Duntas, L.H. Selenium and inflammation: Underlying anti-inflammatory mechanisms. Horm. Metab. Res. 2009, 41, 443–447. [Google Scholar] [CrossRef] [PubMed]
- Flohé, L. The glutathione peroxidase reaction: Molecular basis of the antioxidant function of selenium in mammals. Curr. Top. Cell. Regul. 1985, 27, 473–478. [Google Scholar] [CrossRef] [PubMed]
- Aaseth, J.; Alexander, J.; Alehagen, U. Coenzyme Q(10) supplementation—In ageing and disease. Mech. Ageing Dev. 2021, 197, 111521. [Google Scholar] [CrossRef]
- Gutierrez-Mariscal, F.M.; Yubero-Serrano, E.M.; Villalba, J.M.; Lopez-Miranda, J. Coenzyme Q(10): From bench to clinic in aging diseases, a translational review. Crit. Rev. Food Sci. Nutr. 2019, 59, 2240–2257. [Google Scholar] [CrossRef]
- Kalén, A.; Appelkvist, E.L.; Dallner, G. Age-related changes in the lipid compositions of rat and human tissues. Lipids 1989, 24, 579–584. [Google Scholar] [CrossRef]
- Xia, L.; Nordman, T.; Olsson, J.M.; Damdimopoulos, A.; Björkhem-Bergman, L.; Nalvarte, I.; Eriksson, L.C.; Arnér, E.S.; Spyrou, G.; Björnstedt, M. The mammalian cytosolic selenoenzyme thioredoxin reductase reduces ubiquinone. A novel mechanism for defense against oxidative stress. J. Biol. Chem. 2003, 278, 2141–2146. [Google Scholar] [CrossRef] [Green Version]
- Alehagen, U.; Johansson, P.; Björnstedt, M.; Rosén, A.; Dahlström, U. Cardiovascular mortality and N-terminal-proBNP reduced after combined selenium and coenzyme Q10 supplementation: A 5-year prospective randomized double-blind placebo-controlled trial among elderly Swedish citizens. Int. J. Cardiol. 2013, 167, 1860–1866. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alehagen, U.; Aaseth, J.; Johansson, P. Reduced Cardiovascular Mortality 10 Years after Supplementation with Selenium and Coenzyme Q10 for Four Years: Follow-Up Results of a Prospective Randomized Double-Blind Placebo-Controlled Trial in Elderly Citizens. PLoS ONE 2015, 10, e0141641. [Google Scholar] [CrossRef]
- Alehagen, U.; Aaseth, J.; Alexander, J.; Johansson, P. Still reduced cardiovascular mortality 12 years after supplementation with selenium and coenzyme Q10 for four years: A validation of previous 10-year follow-up results of a prospective randomized double-blind placebo-controlled trial in elderly. PLoS ONE 2018, 13, e0193120. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alehagen, U.; Alexander, J.; Aaseth, J.; Larsson, A. Decrease in inflammatory biomarker concentration by intervention with selenium and coenzyme Q10: A subanalysis of osteopontin, osteoprotergerin, TNFr1, TNFr2 and TWEAK. J. Inflamm. 2019, 16, 5. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alehagen, U.; Lindahl, T.L.; Aaseth, J.; Svensson, E.; Johansson, P. Levels of sP-selectin and hs-CRP Decrease with Dietary Intervention with Selenium and Coenzyme Q10 Combined: A Secondary Analysis of a Randomized Clinical Trial. PLoS ONE 2015, 10, e0137680. [Google Scholar] [CrossRef]
- Alehagen, U.; Aaseth, J.; Johansson, P. Less increase of copeptin and MR-proADM due to intervention with selenium and coenzyme Q10 combined: Results from a 4-year prospective randomized double-blind placebo-controlled trial among elderly Swedish citizens. Biofactors 2015, 41, 443–452. [Google Scholar] [CrossRef]
- Alehagen, U.; Alexander, J.; Aaseth, J.; Larsson, A. Lindahl TL. Significant decrease of von Willebrand factor and plasminogen activator inhibitor-1 by providing supplementation with selenium and coenzyme Q10 to an elderly population with a low selenium status. Eur. J. Nutr. 2020, 59, 3581–3590. [Google Scholar] [CrossRef] [Green Version]
- Opstad, T.B.; Alexander, J.; Aaseth, J.O.; Larsson, A.; Seljeflot, I.; Alehagen, U. Selenium and Coenzyme Q(10) Intervention Prevents Telomere Attrition, with Association to Reduced Cardiovascular Mortality-Sub-Study of a Randomized Clinical Trial. Nutrients 2022, 14, 3346. [Google Scholar] [CrossRef]
- Giacconi, R.; Chiodi, L.; Boccoli, G.; Costarelli, L.; Piacenza, F.; Provinciali, M.; Malavolta, M. Reduced levels of plasma selenium are associated with increased inflammation and cardiovascular disease in an Italian elderly population. Exp. Gerontol. 2021, 145, 111219. [Google Scholar] [CrossRef]
- Haigis, M.C.; Sinclair, D.A. Mammalian sirtuins: Biological insights and disease relevance. Annu. Rev. Pathol. 2010, 5, 253–295. [Google Scholar] [CrossRef] [Green Version]
- Yamamoto, H.; Schoonjans, K.; Auwerx, J. Sirtuin functions in health and disease. Mol. Endocrinol. 2007, 21, 1745–1755. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kauppinen, A.; Suuronen, T.; Ojala, J.; Kaarniranta, K.; Salminen, A. Antagonistic crosstalk between NF-κB and SIRT1 in the regulation of inflammation and metabolic disorders. Cell. Signal. 2013, 25, 1939–1948. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.; Zhou, M.; Ge, Y.; Wang, X. SIRT1 and aging related signaling pathways. Mech. Ageing Dev. 2020, 187, 111215. [Google Scholar] [CrossRef] [PubMed]
- Ministrini, S.; Puspitasari, Y.M.; Beer, G.; Liberale, L.; Montecucco, F.; Camici, G.G. Sirtuin 1 in Endothelial Dysfunction and Cardiovascular Aging. Front. Physiol. 2021, 12, 733696. [Google Scholar] [CrossRef]
- Nogales, F.; Cebadero, O.; Romero-Herrera, I.; Rua, R.M.; Carreras, O.; Ojeda, M.L. Selenite supplementation modulates the hepatic metabolic sensors AMPK and SIRT1 in binge drinking exposed adolescent rats by avoiding oxidative stress. Food Funct. 2021, 12, 3022–3032. [Google Scholar] [CrossRef] [PubMed]
- DiNicolantonio, J.J.; McCarty, M.F.; O’Keefe, J.H. Coenzyme Q10 deficiency can be expected to compromise Sirt1 activity. Open Heart 2022, 9, e001927. [Google Scholar] [CrossRef]
- Samimi, F.; Baazm, M.; Eftekhar, E.; Rajabi, S.; Goodarzi, M.T.; Jalali Mashayekhi, F. Possible antioxidant mechanism of coenzyme Q10 in diabetes: Impact on Sirt1/Nrf2 signaling pathways. Res. Pharm. Sci. 2019, 14, 524–533. [Google Scholar] [CrossRef] [PubMed]
- Alehagen, U.; Johansson, P.; Aaseth, J.; Alexander, J.; Wågsäter, D. Significant changes in circulating microRNA by dietary supplementation of selenium and coenzyme Q10 in healthy elderly males. A subgroup analysis of a prospective randomized double-blind placebo-controlled trial among elderly Swedish citizens. PLoS ONE 2017, 12, e0174880. [Google Scholar] [CrossRef] [Green Version]
- Zia, A.; Sahebdel, F.; Farkhondeh, T.; Ashrafizadeh, M.; Zarrabi, A.; Hushmandi, K.; Samarghandian, S. A review study on the modulation of SIRT1 expression by miRNAs in aging and age-associated diseases. Int. J. Biol. Macromol. 2021, 188, 52–61. [Google Scholar] [CrossRef]
- Dhonukshe-Rutten, R.A.; Bouwman, J.; Brown, K.A.; Cavelaars, A.E.; Collings, R.; Grammatikaki, E.; de Groot, L.C.; Gurinovic, M.; Harvey, L.J.; Hermoso, M.; et al. EURRECA-Evidence-based methodology for deriving micronutrient recommendations. Crit. Rev. Food Sci. Nutr. 2013, 53, 999–1040. [Google Scholar] [CrossRef]
- Xia, Y.; Hill, K.E.; Li, P.; Xu, J.; Zhou, D.; Motley, A.K.; Wang, L.; Byrne, D.W.; Burk, R.F. Optimization of selenoprotein P and other plasma selenium biomarkers for the assessment of the selenium nutritional requirement: A placebo-controlled, double-blind study of selenomethionine supplementation in selenium-deficient Chinese subjects. Am. J. Clin. Nutr. 2010, 92, 525–531. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kitada, M.; Ogura, Y.; Koya, D. The protective role of Sirt1 in vascular tissue: Its relationship to vascular aging and atherosclerosis. Aging 2016, 8, 2290–2307. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Elibol, B.; Kilic, U. High Levels of SIRT1 Expression as a Protective Mechanism Against Disease-Related Conditions. Front. Endocrinol. 2018, 9, 614. [Google Scholar] [CrossRef] [PubMed]
- Mariani, S.; Fiore, D.; Basciani, S.; Persichetti, A.; Contini, S.; Lubrano, C.; Salvatori, L.; Lenzi, A.; Gnessi, L. Plasma levels of SIRT1 associate with non-alcoholic fatty liver disease in obese patients. Endocrine 2015, 49, 711–716. [Google Scholar] [CrossRef]
- Opstad, T.B.; Farup, P.G.; Rootwelt, H.; Aaseth, J.O. Changes in circulating sirtuin 1 after bariatric surgery. Nutr. Metab. Cardiovasc. Dis. 2022, 32, 2858–2864. [Google Scholar] [CrossRef]
- Suksomboon, N.; Poolsup, N.; Juanak, N. Effects of coenzyme Q10 supplementation on metabolic profile in diabetes: A systematic review and meta-analysis. J. Clin. Pharm. Ther. 2015, 40, 413–418. [Google Scholar] [CrossRef]
- Opstad, T.B.; Berg, T.J.; Holte, K.B.; Arnesen, H.; Solheim, S.; Seljeflot, I. Reduced leukocyte telomere lengths and sirtuin 1 gene expression in long-term survivors of type 1 diabetes: A Dialong substudy. J. Diabetes. Investig. 2021, 12, 1183–1192. [Google Scholar] [CrossRef]
- Ma, B.; Guo, B.; Chen, Z.; Li, Y. SIRT1 regulates hypoxia-induced oxidative stress in cardiomyocytes via PI3K/MTOR signaling. Cell Mol. Biol. 2022, 68, 48–53. [Google Scholar] [CrossRef]
- Packer, M. Cardioprotective Effects of Sirtuin-1 and Its Downstream Effectors: Potential Role in Mediating the Heart Failure Benefits of SGLT2 (Sodium-Glucose Cotransporter 2) Inhibitors. Circ. Heart Fail. 2020, 13, e007197. [Google Scholar] [CrossRef]
- Shengyu, C.; Yinhua, L.; Yuanhong, L.; Jinbo, Z.; Can, F.; Hao, X.; Changjiang, Z. Selenium alleviates heart remodeling through Sirt1/AKT/GSK-3β pathway. Int. Immunopharmacol. 2022, 111, 109158. [Google Scholar] [CrossRef]
- Mariani, S.; Costantini, D.; Lubrano, C.; Basciani, S.; Caldaroni, C.; Barbaro, G.; Poggiogalle, E.; Donini, L.M.; Lenzi, A.; Gnessi, L. Circulating SIRT1 inversely correlates with epicardial fat thickness in patients with obesity. Nutr. Metab. Cardiovasc. Dis. 2016, 26, 1033–1038. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Esmayel, I.M.; Hussein, S.; Gohar, E.A.; Ebian, H.F.; Mousa, M.M. Plasma levels of sirtuin-1 in patients with cerebrovascular stroke. Neurol. Sci. 2021, 42, 3843–3850. [Google Scholar] [CrossRef] [PubMed]
- Viana-Mattioli, S.; Nunes, P.; Cavalli, R.; Sandrim, V. Analysis of SIRT1 Expression in Plasma and in an In Vitro Model of Preeclampsia. Oxid. Med. Cell. Longev. 2020, 2020, 4561083. [Google Scholar] [CrossRef] [PubMed]
- Imai, S.; Guarente, L. NAD+ and sirtuins in aging and disease. Trends Cell Biol. 2014, 24, 464–471. [Google Scholar] [CrossRef]
- Mapelli, V.; Hillestrøm, P.R.; Kápolna, E.; Larsen, E.H.; Olsson, L. Metabolic and bioprocess engineering for production of selenized yeast with increased content of seleno-methylselenocysteine. Metab. Eng. 2011, 13, 282–293. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fang, M.; Guo, W.R.; Park, Y.; Kang, H.G.; Zarbl, H. Enhancement of NAD+-dependent SIRT1 deacetylase activity by methylselenocysteine resets the circadian clock in carcinogen-treated mammary epithelial cells. Oncotarget 2015, 6, 42879–42891. [Google Scholar] [CrossRef] [Green Version]
- Donato, A.J.; Magerko, K.A.; Lawson, B.R.; Durrant, J.R.; Lesniewski, L.A.; Seals, D.R. SIRT-1 and vascular endothelial dysfunction with ageing in mice and humans. J. Physiol. 2011, 589, 4545–4554. [Google Scholar] [CrossRef]
- Palacios, J.A.; Herranzm, D.; De Bonis, M.L.; Velasco, S.; Serrano, M.; Blasco, M.A. SIRT1 contributes to telomere maintenance and augments global homologous recombination. J. Cell Biol. 2010, 191, 1299–1313. [Google Scholar] [CrossRef] [Green Version]
- Higuchi, M.; Dusting, G.J.; Peshavariya, H.; Jiang, F.; Hsiao, S.T.; Chan, E.C.; Liu, G.S. Differentiation of human adipose-derived stem cells into fat involves reactive oxygen species and Forkhead box O1 mediated upregulation of antioxidant enzymes. Stem Cells Dev. 2013, 22, 878–888. [Google Scholar] [CrossRef] [Green Version]
- Pardo, P.S.; Boriek, A.M. SIRT1 Regulation in Ageing and Obesity. Mech. Ageing Dev. 2020, 188, 111249. [Google Scholar] [CrossRef]
- Gerasymchuk, M.; Cherkasova, V.; Kovalchuk, O.; Kovalchuk, I. The Role of microRNAs in Organismal and Skin Aging. Int. J. Mol. Sci. 2020, 21, 5281. [Google Scholar] [CrossRef] [PubMed]
- Yang, X.; Yang, Y.; Gan, R.; Zhao, L.; Li, W.; Zhou, H.; Wang, X.; Lu, J.; Meng, Q.H. Down-regulation of mir-221 and mir-222 restrain prostate cancer cell proliferation and migration that is partly mediated by activation of SIRT1. PLoS ONE 2014, 9, e98833. [Google Scholar] [CrossRef] [PubMed]
- Yang, Z.; Lin, S.D.; Zhan, F.; Liu, Y.; Zhan, Y.W. LncRNA GAS5 alleviates rheumatoid arthritis through regulating miR-222-3p/Sirt1 signalling axis. Autoimmunity 2021, 54, 13–22. [Google Scholar] [CrossRef] [PubMed]
- Long, J.K.; Dai, W.; Zheng, Y.W.; Zhao, S.P. miR-122 promotes hepatic lipogenesis via inhibiting the LKB1/AMPK pathway by targeting Sirt1 in non-alcoholic fatty liver disease. Mol. Med. 2019, 25, 26. [Google Scholar] [CrossRef] [PubMed] [Green Version]




{kind=link}
{kind=link}
{kind=link}
| Active Treatment Group n = 165 | Placebo Group n = 161 | p-Value | |
|---|---|---|---|
| Age years | 76.1 (3.2) | 76.4 (3.0) | 0.43 |
| Sex, Males/Females n | 85/80 | 82/79 | |
| History | |||
| Smoking, n (%) | 8 (4.8) | 8 (5.0) | 0.96 |
| Diabetes, n (%) | 34 (20.6) | 29 (18.0) | 0.55 |
| Hypertension, n (%) | 118 (71.5) | 116 (72.0) | 0.91 |
| IHD, n (%) | 25 (15.2) | 29 (18.0) | 0.49 |
| Atrial fibrillation, n (%) | 14 (8.5) | 16 (9.9) | 0.65 |
| NYHA class I, n (%) | 99 (60.0) | 87 (54.0) | 0.28 |
| NYHA class II, n (%) | 44 (26.7) | 44 (27.3) | 0.89 |
| NYHA class III, n (%) | 21 (12.7) | 28 (17.4) | 0.24 |
| NYHA class IV, n (%) | 0 | 0 | |
| Unclassified, n (%) | 1 (0.6) | 2 (1.2) | |
| Medications | |||
| ACEI/ARB, n (%) | 28 (17.0) | 34 (21.1) | 0.34 |
| Beta blockers, n (%) | 55 (33.3) | 46 (28.6) | 0.35 |
| Diuretics, n (%) | 46 (27.9) | 60 (37.3) | 0.07 |
| Statins, n (%) | 32 (19.4) | 37 (23.0) | 0.73 |
| Examinations | |||
| EF < 40%, n (%) | 7 (4.2) | 6 (3.7) | 0.81 |
| s-selenium s-coenzyme Q10 | 67.4 (14.2) 0.84 (0.31) | 67.2 (13.2) 0.88 (0.34) | 0.80 0.68 |
| CV-death, n (%) | 25 (15) | 52 (32) | <0.001 |
| Effects | Sum of Squares | Degrees of Freedom | Mean Squares | F | p |
|---|---|---|---|---|---|
| Intercept | 389,335 | 1 | 389,335 | 14.37 | 0.0002 |
| Age >75 years | 4755 | 1 | 4755 | 0.18 | 0.68 |
| CRP fold change | 77,029 | 1 | 77,029 | 2.84 | 0.09 |
| SIRT1 baseline | 13,786 | 1 | 13,786 | 0.51 | 0.48 |
| Smoker | 2494 | 1 | 2494 | 0.09 | 0.76 |
| Male sex | 18,523 | 1 | 18,523 | 0.68 | 0.41 |
| Hypertension | 86,222 | 1 | 86,222 | 3.18 | 0.08 |
| Diabetes | 5233 | 1 | 5233 | 0.19 | 0.66 |
| IHD | 35,406 | 1 | 35,406 | 1.31 | 0.26 |
| Active treatment | 210,429 | 1 | 210,429 | 7.77 | 0.006 |
| Error | 3,115,998 | 115 | 27,095 |
| MicroRNAs | n | r | p |
|---|---|---|---|
| miR-130a-3p | 32 | r = −0.466 | 0.007 |
| miR-19b-3p | 27 | r = −0.447 | 0.019 |
| miR-16-2-3p | 25 | r = −0.459 | 0.020 |
| miR-222-3p | 27 | r = −0.438 | 0.022 |
| miR-454-3p | 22 | r = −0.446 | 0.040 |
| miR-423-3p | 27 | r = 0.535 | 0.004 |
| miR-30b-5p | 27 | r = 0.523 | 0.005 |
| miR-181-5p | 27 | r = 0.488 | 0.010 |
| miR-191-5p | 27 | r = 0.393 | 0.042 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Opstad, T.B.; Alexander, J.; Aaseth, J.; Larsson, A.; Seljeflot, I.; Alehagen, U. Increased SIRT1 Concentration Following Four Years of Selenium and Q10 Intervention Associated with Reduced Cardiovascular Mortality at 10-Year Follow-Up—Sub-Study of a Previous Prospective Double-Blind Placebo-Controlled Randomized Clinical Trial. Antioxidants 2023, 12, 759. https://doi.org/10.3390/antiox12030759
Opstad TB, Alexander J, Aaseth J, Larsson A, Seljeflot I, Alehagen U. Increased SIRT1 Concentration Following Four Years of Selenium and Q10 Intervention Associated with Reduced Cardiovascular Mortality at 10-Year Follow-Up—Sub-Study of a Previous Prospective Double-Blind Placebo-Controlled Randomized Clinical Trial. Antioxidants. 2023; 12(3):759. https://doi.org/10.3390/antiox12030759
Chicago/Turabian StyleOpstad, Trine Baur, Jan Alexander, Jan Aaseth, Anders Larsson, Ingebjørg Seljeflot, and Urban Alehagen. 2023. "Increased SIRT1 Concentration Following Four Years of Selenium and Q10 Intervention Associated with Reduced Cardiovascular Mortality at 10-Year Follow-Up—Sub-Study of a Previous Prospective Double-Blind Placebo-Controlled Randomized Clinical Trial" Antioxidants 12, no. 3: 759. https://doi.org/10.3390/antiox12030759
APA StyleOpstad, T. B., Alexander, J., Aaseth, J., Larsson, A., Seljeflot, I., & Alehagen, U. (2023). Increased SIRT1 Concentration Following Four Years of Selenium and Q10 Intervention Associated with Reduced Cardiovascular Mortality at 10-Year Follow-Up—Sub-Study of a Previous Prospective Double-Blind Placebo-Controlled Randomized Clinical Trial. Antioxidants, 12(3), 759. https://doi.org/10.3390/antiox12030759