
Oxidative and Glycation Damage to Mitochondrial DNA and Plastid DNA during Plant Development


Abstract
:
1. Introduction
2. Cellular Metabolism and ROS
2.1. ROS during Abiotic and Biotic Stresses
2.2. ROS-Targeted Damage to Proteins and DNA
2.3. Assessing ROS and Oxidative Damage to orgDNA in Maize
3. Cellular Metabolism and Glycation
- Although the genotoxic effects of ROS have been intensively studied, most such reports do not mention glycation damage. Furthermore, the relative contributions to DNA damage from oxidation and glycation are not known.
- Defense against glycation damage to DNA involves the same enzymes that cleanse the nucleotide pool before polymerization (damage avoidance) and reverse the damage in DNA molecules (deglycase activity). That the structures of these enzymatic substrates are so different is remarkable.
- Perhaps most surprising is the tight coordination of defenses against DNA damage caused by oxidation and glycation.
Assessing Glycation Damage to Maize Proteins and orgDNA
4. Linkage of Damage Caused by Oxidation and Glycation
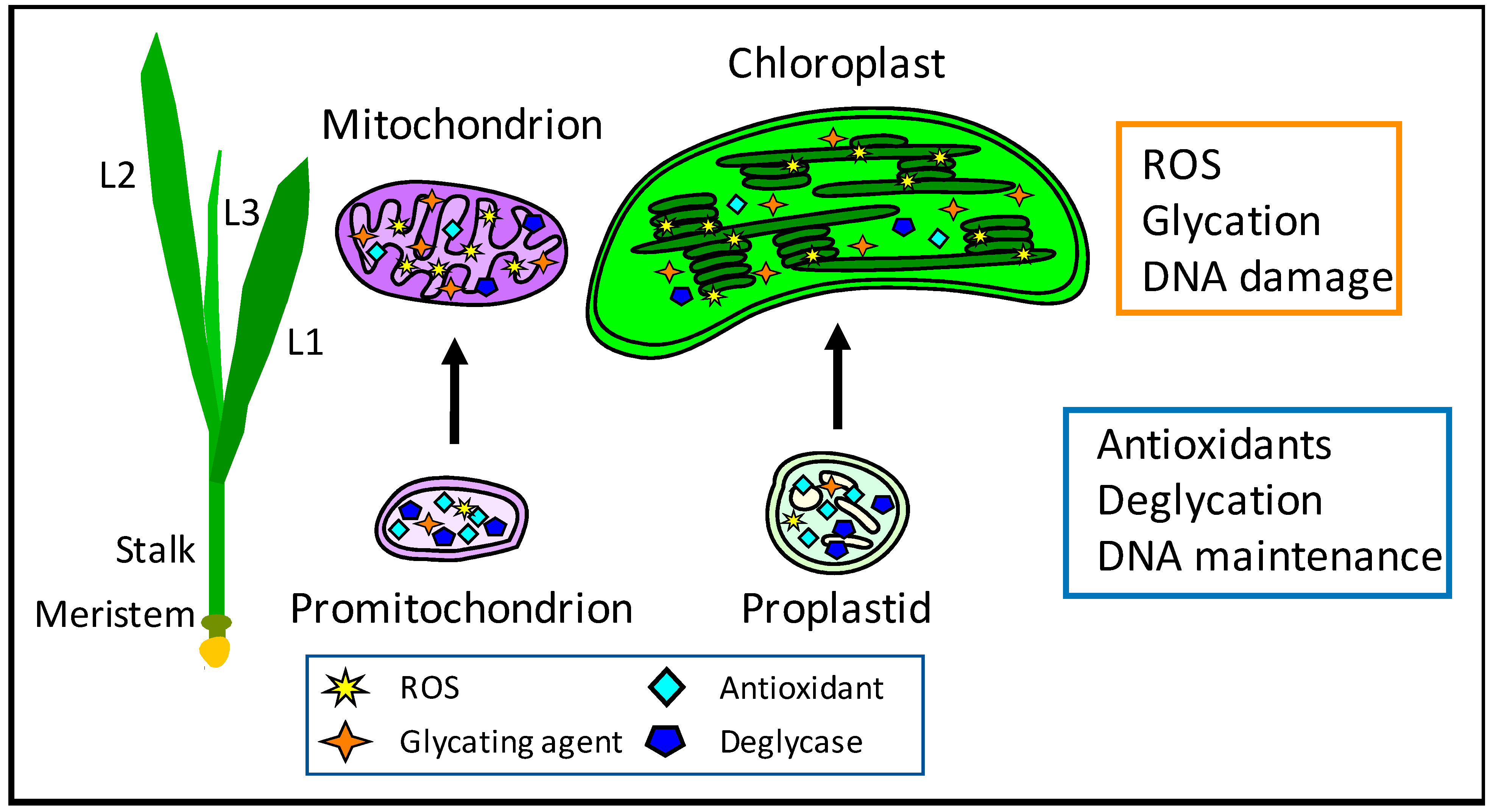
5. Integrity of orgDNA Molecules during Maize Development
6. Detection of Oxidative and Glycation Damages
6.1. Box 1. Measuring ROS, Oxidative Stress, and DNA Damage
6.2. Box 2. Measuring Glycating Agents and Damage to Proteins and DNA
7. Concluding Remarks
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Peralta-Castro, A.; García-Medel, P.L.; Baruch-Torres, N.; Trasviña-Arenas, C.H.; Juarez-Quintero, V.; Morales-Vazquez, C.M.; Brieba, L.G. Plant organellar DNA polymerases evolved multifunctionality through the acquisition of novel amino acid insertions. Genes 2020, 11, 1370. [Google Scholar] [CrossRef] [PubMed]
- Moretton, A.; Loizou, J.I. Interplay between cellular metabolism and the DNA damage response in cancer. Cancers 2020, 12, 2051. [Google Scholar] [CrossRef] [PubMed]
- Boesch, P.; Weber-Lotfi, F.; Ibrahim, N.; Tarasenko, V.; Cosset, A.; Paulus, F.; Lightowlers, R.N.; Dietrich, A. DNA repair in organelles: Pathways, organization, regulation, relevance in disease and aging. Biochim. Biophys. Acta 2011, 1813, 186–200. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Halliwell, B. Reactive species and antioxidants. Redox biology is a fundamental theme of aerobic life. Plant Physiol. 2006, 141, 312–322. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jubany-Marí, T.; Munné-Bosch, S.; López-Carbonell, M.; Alegre, L. Hydrogen peroxide is involved in the acclimation of the Mediterranean shrub, Cistus albidus L., to summer drought. J. Exp. Bot. 2009, 60, 107–120. [Google Scholar] [CrossRef] [Green Version]
- Roychoudhury, A.; Basu, S. Ascorbate-glutathione and plant tolerance to various abiotic stresses. In Oxidative Stress in Plants: Causes, Consequences and Tolerance; IK International Publishers: New Delhi, India, 2012; pp. 177–258. [Google Scholar]
- Janků, M.; Luhová, L.; Petřivalský, M. On the origin and fate of reactive oxygen species in plant cell compartments. Antioxidants 2019, 8, 105. [Google Scholar] [CrossRef] [Green Version]
- Tanaka, K.; Hanaoka, M. The early days of plastid retrograde signaling with respect to replication and transcription. Front. Plant Sci. 2012, 3, 301. [Google Scholar] [CrossRef] [Green Version]
- Foyer, C.H. Reactive oxygen species, oxidative signaling and the regulation of photosynthesis. Environ. Exp. Bot. 2018, 154, 134–142. [Google Scholar] [CrossRef]
- Kacprzak, S.M.; Mochizuki, N.; Naranjo, B.; Xu, D.; Leister, D.; Kleine, T.; Okamoto, H.; Terry, M.J. Plastid-to-nucleus retrograde signalling during chloroplast biogenesis does not require ABI4. Plant Physiol. 2019, 179, 18–23. [Google Scholar] [CrossRef] [Green Version]
- Apel, K.; Hirt, H. Reactive oxygen species: Metabolism, oxidative stress, and signal transduction. Annu. Rev. Plant Biol. 2004, 55, 373–399. [Google Scholar] [CrossRef] [Green Version]
- Foyer, C.H.; Noctor, G. Redox homeostasis and antioxidant signaling: A metabolic interface between stress perception and physiological responses. Plant Cell 2005, 17, 1866–1875. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Das, K.; Roychoudhury, A. Reactive oxygen species (ROS) and response of antioxidants as ROS-scavengers during environmental stress in plants. Front. Environ. Sci. 2014, 2, 53. [Google Scholar] [CrossRef] [Green Version]
- Roldan-Arjona, T.; Ariza, R.R. DNA demethylation. In DNA and RNA Modification Enzymes: Comparative Structure, Mechanism, Functions, Cellular Interactions and Evolution; Grosjean, H., Ed.; Landes Bioscience: Austin, TX, USA, 2009; pp. 149–161. [Google Scholar]
- Nisa, M.U.; Huang, Y.; Benhamed, M.; Raynaud, C. The plant DNA damage response: Signaling pathways leading to growth inhibition and putative role in response to stress conditions. Front. Plant Sci. 2019, 10, 653. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zandalinas, S.I.; Mittler, R. ROS-induced ROS release in plant and animal cells. Free Radic. Biol. Med. 2018, 122, 21–27. [Google Scholar] [CrossRef]
- Gill, S.S.; Tuteja, N. Reactive oxygen species and antioxidant machinery in abiotic stress tolerance in crop plants. Plant Physiol. Biochem. 2010, 48, 909–930. [Google Scholar] [CrossRef]
- Miller, G.; Suzuki, N.; Ciftci-Yilmaz, S.; Mittler, R. Reactive oxygen species homeostasis and signalling during drought and salinity stresses. Plant Cell Environ. 2010, 33, 453–467. [Google Scholar] [CrossRef] [PubMed]
- Gill, S.S.; Khan, N.A.; Anjum, N.A.; Tuteja, N. Amelioration of cadmium stress in crop plants by nutrients management: Morphological, physiological and biochemical aspects. In Plant Nutrition and Abiotic Stress Tolerance III; Anjum, N.A., Lopez-Lauri, F., Eds.; Global Science Books Ltd.: Ikenobe, Japan, 2011; pp. 1–23. [Google Scholar]
- Pitzschke, A.; Hirt, H. Mitogen-activated protein kinases and reactive oxygen species signaling in plants. Plant Physiol. 2006, 141, 351–356. [Google Scholar] [CrossRef] [Green Version]
- Galvez-Valdivieso, G.; Mullineaux, P.M. The role of reactive oxygen species in signalling from chloroplasts to the nucleus. Physiol. Plant 2010, 138, 430–439. [Google Scholar] [CrossRef]
- Mullineaux, P.M.; Exposito-Rodriguez, M.; Laissue, P.P.; Smirnoff, N.; Park, E. Spatial chloroplast-to-nucleus signalling involving plastid-nuclear complexes and stromules. Philos. Trans. R. Soc. Lond. B Biol. Sci. 2020, 375, 20190405. [Google Scholar] [CrossRef]
- Swanson, S.; Gilroy, S. ROS in plant development. Physiol. Plant 2010, 138, 384–392. [Google Scholar] [CrossRef]
- Imlay, J.A. The molecular mechanisms and physiological consequences of oxidative stress: Lessons from a model bacterium. Nat. Rev. Microbiol. 2013, 11, 443–454. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mohyeldin, A.; Garzón-Muvdi, T.; Quiñones-Hinojosa, A. Oxygen in stem cell biology: A critical component of the stem cell niche. Cell Stem Cell 2010, 7, 150–161. [Google Scholar] [CrossRef] [Green Version]
- Considine, M.J.; Diaz-Vivancos, P.; Kerchev, P.; Signorelli, S.; Agudelo-Romero, P.; Gibbs, D.J.; Foyer, C.H. Learning to breathe: Developmental phase transitions in oxygen status. Trends Plant Sci. 2017, 22, 140–153. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tripathi, D.; Nam, A.; Oldenburg, D.J.; Bendich, A.J. Reactive oxygen species, antioxidant agents, and DNA damage in developing maize mitochondria and plastids. Front. Plant Sci. 2020, 11, 596. [Google Scholar] [CrossRef] [PubMed]
- Huang, H.; Ullah, F.; Zhou, D.-X.; Yi, M.; Zhao, Y. Mechanisms of ROS regulation of plant development and stress responses. Front. Plant Sci. 2019, 10, 3389. [Google Scholar] [CrossRef]
- Weits, D.A.; Kunkowska, A.B.; Kamps, N.C.W.; Portz, K.M.S.; Packbier, N.K.; Nemec Venza, Z.; Gaillochet, C.; Lohmann, J.U.; Pedersen, O.; van Dongen, J.T.; et al. An apical hypoxic niche sets the pace of shoot meristem activity. Nature 2019, 569, 714–717. [Google Scholar] [CrossRef]
- Černý, M.; Habánová, H.; Berka, M.; Luklová, M.; Brzobohatý, B. Hydrogen Peroxide: Its role in plant biology and crosstalk with signalling networks. Int. J. Mol. Sci. 2018, 19, 2812. [Google Scholar] [CrossRef] [Green Version]
- Locato, V.; Cimini, S.; De Gara, L. ROS and redox balance as multifaceted players of cross-tolerance: Epigenetic and retrograde control of gene expression. J. Exp. Bot. 2018, 69, 3373–3391. [Google Scholar] [CrossRef] [Green Version]
- Smirnoff, N.; Arnaud, D. Hydrogen peroxide metabolism and functions in plants. New Phytol. 2019, 221, 1197–1214. [Google Scholar] [CrossRef] [Green Version]
- Soares, C.; Carvalho, M.E.A.; Azevedo, R.A.; Fidalgo, F. Plants facing oxidative challenges—A little help from the antioxidant networks. Environ. Exp. Bot. 2019, 161, 4–25. [Google Scholar] [CrossRef]
- Møller, I.M.; Jensen, P.E.; Hansson, A. Oxidative modifications to cellular components in plants. Annu. Rev. Plant Biol. 2007, 58, 459–481. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Evans, M.D.; Dizdaroglu, M.; Cooke, M.S. Oxidative DNA damage and disease: Induction, repair and significance. Mutat. Res. 2004, 567, 1–61. [Google Scholar] [CrossRef] [PubMed]
- De Medeiros, N.M.C.; De Medeiros, A.L.M.; Silva, H.C.; Scortecci, K.C. Recent advances in plant DNA repair. In Advances in Plant DNA Repair; IntechOpen: London, UK, 2015. [Google Scholar]
- Spampinato, C.P. Protecting DNA from errors and damage: An overview of DNA repair mechanisms in plants compared to mammals. Cell Mol. Life Sci. 2017, 74, 1693–1709. [Google Scholar] [CrossRef] [Green Version]
- Gutman, B.L.; Niyogi, K.K. Evidence for base excision repair of oxidative DNA damage in chloroplasts of Arabidopsis thaliana. J. Biol. Chem. 2009, 284, 17006–17012. [Google Scholar] [CrossRef] [Green Version]
- García-Medel, P.L.; Baruch-Torres, N.; Peralta-Castro, A.; Trasviña-Arenas, C.H.; Torres-Larios, A.; Brieba, L.G. Plant organellar DNA polymerases repair double-stranded breaks by microhomology-mediated end-joining. Nucleic Acids Res. 2019, 47, 3028–3044. [Google Scholar] [CrossRef]
- Markkanen, E. Not breathing is not an option: How to deal with oxidative DNA damage. DNA Repair 2017, 59, 82–105. [Google Scholar] [CrossRef] [PubMed]
- Bendich, A.J. Mitochondrial DNA, chloroplast DNA and the origins of development in eukaryotic organisms. Biol. Direct 2010, 5, 42. [Google Scholar] [CrossRef] [Green Version]
- Bendich, A.J. DNA abandonment and the mechanisms of uniparental inheritance of mitochondria and chloroplasts. Chromosome Res. 2013, 21, 287–296. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oldenburg, D.J.; Bendich, A.J. DNA maintenance in plastids and mitochondria of plants. Front. Plant Sci. 2015, 6, 883. [Google Scholar] [CrossRef] [Green Version]
- Oldenburg, D.J.; Bendich, A.J. Most chloroplast DNA of maize seedlings in linear molecules with defined ends and branched forms. J. Mol. Biol. 2004, 335, 953–970. [Google Scholar] [CrossRef]
- Oldenburg, D.J.; Kumar, R.A.; Bendich, A.J. The amount and integrity of mtDNA in maize decline with development. Planta 2013, 237, 603–617. [Google Scholar] [CrossRef]
- Kumar, R.A.; Oldenburg, D.J.; Bendich, A.J. Changes in DNA damage, molecular integrity, and copy number for plastid DNA and mitochondrial DNA during maize development. J. Exp. Bot. 2014, 65, 6425–6439. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zheng, Q.; Oldenburg, D.J.; Bendich, A.J. Independent effects of leaf growth and light on the development of the plastid and its DNA content in Zea species. J. Exp. Bot. 2011, 62, 2715–2730. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tripathi, D.; Oldenburg, D.J.; Bendich, A.J. Glycation damage to organelles and their DNA increases during maize seedling development. Sci. Rep. 2022, 12, 2688. [Google Scholar] [CrossRef] [PubMed]
- Skarstad, K.; Boye, E. Degradation of individual chromosomes in recA mutants of Escherichia coli. J. Bacteriol. 1993, 175, 5505–5509. [Google Scholar] [CrossRef] [Green Version]
- Martins, S.I.F.S.; Jongen, W.M.F.; van Boekel, M.A.J.S. A review of Maillard reaction in food and implications to kinetic modelling. Trends Food Sci. Technol. 2000, 11, 364–373. [Google Scholar] [CrossRef]
- Kutzli, I.; Weiss, J.; Gibis, M. Glycation of plant proteins via Maillard reaction: Reaction chemistry, technofunctional properties, and potential food application. Foods 2021, 10, 376. [Google Scholar] [CrossRef]
- Thornalley, P.J. Protein and nucleotide damage by glyoxal and methylglyoxal in physiological systems-role in ageing and disease. Drug Metab. Drug Interact. 2008, 23, 125–150. [Google Scholar] [CrossRef]
- Richarme, G.; Liu, C.; Mihoub, M.; Abdallah, J.; Leger, T.; Joly, N.; Liebart, J.C.; Jurkunas, U.V.; Nadal, M.; Bouloc, P.; et al. Guanine glycation repair by DJ-1/Park7 and its bacterial homologs. Science 2017, 357, 208–211. [Google Scholar] [CrossRef] [Green Version]
- Takeuchi, M.; Sakasai-Sakai, A.; Takata, T.; Takino, J.I.; Koriyama, Y.; Kikuchi, C.; Furukawa, A.; Nagamine, K.; Hori, T.; Matsunaga, T. Intracellular toxic AGEs (TAGE) triggers numerous types of cell damage. Biomolecules 2021, 11, 387. [Google Scholar] [CrossRef] [PubMed]
- Thornalley, P.J. The glyoxalase system: New developments towards functional characterization of a metabolic pathway fundamental to biological life. Biochem. J. 1990, 269, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Sankaranarayanan, S.; Jamshed, M.; Kumar, A.; Skori, L.; Scandola, S.; Wang, T.; Spiegel, D.; Samuel, M.A. Glyoxalase goes green: The expanding roles of glyoxalase in plants. Int. J. Mol. Sci 2017, 18, 898. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Richarme, G.; Mihoub, M.; Dairou, J.; Bui, L.C.; Leger, T.; Lamouri, A. Parkinsonism-associated protein DJ-1/Park7 is a major protein deglycase that repairs methylglyoxal-and glyoxal-glycated cysteine, arginine, and lysine residues. J. Biol. Chem. 2015, 290, 1885–1897. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jun, Y.W.; Kool, E.T. Small substrate or large? debate over the mechanism of glycation adduct repair by DJ-1. Cell Chem. Biol. 2020, 27, 1117–1123. [Google Scholar] [CrossRef] [PubMed]
- Prasad, M.; Kataria, P.; Ningaraju, S.; Buddidathi, R.; Bankapalli, K.; Swetha, C.; Susarla, G.; Venkatesan, R.; D’Silva, P.; Shivaprasad, P.V. Double DJ-1 domain containing Arabidopsis DJ-1D is a robust macromolecule deglycase. New Phytol. 2022, 236, 1061–1074. [Google Scholar] [CrossRef]
- Xu, X.M.; Lin, H.; Maple, J.; Björkblom, B.; Alves, G.; Larsen, J.P.; Møller, S.G. The Arabidopsis DJ-1a protein confers stress protection through cytosolic SOD activation. J. Cell Sci. 2010, 123, 1644–1651. [Google Scholar] [CrossRef] [Green Version]
- Lewandowska, A.; Vo, T.N.; Nguyen, T.H.; Wahni, K.; Vertommen, D.; Van Breusegem, F.; Young, D.; Messens, J. Bifunctional chloroplastic DJ-1B from Arabidopsis thaliana is an oxidation-robust holdase and a glyoxalase sensitive to H2O2. Antioxidants 2019, 8, 8. [Google Scholar] [CrossRef] [Green Version]
- Lin, J.; Nazarenus, T.J.; Frey, J.L.; Liang, X.; Wilson, M.A.; Stone, J.M. A plant DJ-1 homolog is essential for Arabidopsis thaliana chloroplast development. PLoS ONE 2011, 6, e23731. [Google Scholar] [CrossRef]
- Ghosh, A.; Kushwaha, H.R.; Hasan, M.R.; Pareek, A.; Sopory, S.K.; Singla-Pareek, S.L. Presence of unique glyoxalase III proteins in plants indicates the existence of shorter route for methylglyoxal detoxification. Sci. Rep. 2016, 6, 18358. [Google Scholar] [CrossRef] [Green Version]
- Nukaya, H.; Iwami, T.; Ishida, H.; Tsuji, K.; Suwa, Y.; Wakabayashi, K.; Nagao, M.; Sugimura, T.; Kosuge, T. N-2 acetylation of 2’-deoxyguanosine by coffee mutagens, methylglyoxal and hydrogen peroxide. Mutat. Res. 1990, 245, 251–257. [Google Scholar] [CrossRef]
- Abordo, E.A.; Minhas, H.S.; Thornalley, P.J. Accumulation of alpha-oxoaldehydes during oxidative stress: A role in cytotoxicity. Biochem. Pharmacol. 1999, 58, 641–648. [Google Scholar] [CrossRef] [PubMed]
- Fournet, M.; Bonté, F.; Desmoulière, A. Glycation damage: A possible hub for major pathophysiological disorders and aging. Aging Dis. 2018, 9, 880–900. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rosca, M.G.; Mustata, T.G.; Kinter, M.T.; Ozdemir, A.M.; Kern, T.S.; Szweda, L.I.; Brownlee, M.; Monnier, V.M.; Weiss, M.F. Glycation of mitochondrial proteins from diabetic rat kidney is associated with excess superoxide formation. Am. J. Physiol. Renal. Physiol. 2005, 289, F420–F430. [Google Scholar] [CrossRef] [Green Version]
- Morcos, M.; Morcos, M.; Du, X.; Sayed, A.; Hutter, H.; Pfisterer, F.; Kukudov, G.; Thornalley, P.; Baynes, J.; Thorpe, S.; et al. Life extension in Caenorhabditis elegans by overexpression of Glyoxalase I—A mechanistic integration of protein damage by glycation, oxidation and nitratio. Comp. Biochem. Physiol. Part A Mol. Integr. Physiol. 2007, 146 (Suppl. S4), S59. [Google Scholar] [CrossRef]
- De Bari, L.; Scirè, A.; Minnelli, C.; Cianfruglia, L.; Kalapos, M.P.; Armeni, T. Interplay among oxidative stress, methylglyoxal pathway and S-glutathionylation. Antioxidants 2021, 10, 19. [Google Scholar] [CrossRef]
- Breyer, V.; Weigel, I.; Huang, T.T.; Pischetsrieder, M. Endogenous mitochondrial oxidative stress in MnSOD-deficient mouse embryonic fibroblasts promotes mitochondrial DNA glycation. Free Radic. Biol. Med. 2012, 52, 1744–1749. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kang, J.H. Oxidative damage of DNA induced by methylglyoxal in vitro. Toxicol. Lett. 2003, 145, 181–187. [Google Scholar] [CrossRef]
- Akter, S.; Khan, M.S.; Smith, E.N.; Flashman, E. Measuring ROS and redox markers in plant cells. RSC Chem. Biol. 2021, 2, 1384–1401. [Google Scholar] [CrossRef]
- Marrocco, I.; Altieri, F.; Peluso, I. Measurement and clinical significance of biomarkers of oxidative stress in humans. Oxidative Med. Cell Longev. 2017, 2017, 6501046. [Google Scholar] [CrossRef]
- Katerji, M.; Filippova, M.; Duerksen-Hughes, P. Approaches and methods to measure oxidative stress in clinical samples: Research applications in the cancer field. Oxidative Med. Cell Longev. 2019, 2019, 1279250. [Google Scholar] [CrossRef] [Green Version]
- Amorati, R.; Valgimigli, L. Methods to measure the antioxidant activity of phytochemicals and plant extracts. J. Agric. Food Chem. 2018, 66, 3324–3329. [Google Scholar] [CrossRef]
- Hunter, S.E.; Jung, D.; Di Giulio, R.T.; Meyer, J.N. The QPCR assay for analysis of mitochondrial DNA damage, repair, and relative copy number. Methods 2010, 51, 444–451. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Parent, J.S.; Lepage, E.; Brisson, N. Divergent roles for the two PolI-like organelle DNA polymerases of Arabidopsis. Plant Physiol. 2011, 156, 254–262. [Google Scholar] [CrossRef] [Green Version]
- Wallace, S.S. DNA glycosylases search for and remove oxidized DNA bases. Environ. Mol. Mutagen 2013, 54, 691–704. [Google Scholar] [CrossRef] [Green Version]
- Bokhari, B.; Sharma, S. Stress marks on the genome: Use or Lose? Int. J. Mol. Sci. 2019, 20, 364. [Google Scholar] [CrossRef] [Green Version]
- Shen, J.; Deininger, P.; Hunt, J.D.; Zhao, H. 8-Hydroxy-2’-deoxyguanosine (8-OH-dG) as a potential survival biomarker in patients with non small-cell lung cancer. Cancer 2007, 109, 574–580. [Google Scholar] [CrossRef]
- Johnson, R.N.; Metcalf, P.A.; Baker, J.R. Fructosamine: A new approach to the estimation of serum glycosylprotein. an index of diabetic control. Clin. Chim. Acta 1983, 127, 87–95. [Google Scholar] [CrossRef] [PubMed]
- Mironova, R.; Niwa, T.; Handzhiyski, Y.; Sredovska, A.; Ivanov, I. Evidence for non-enzymatic glycosylation of Escherichia coli chromosomal DNA. Mol. Microbiol. 2005, 55, 1801–1811. [Google Scholar] [CrossRef] [PubMed]
- Oldenburg, D.J.; Rowan, B.A.; Kumar, R.A.; Bendich, A.J. On the fate of plastid DNA molecules during leaf development: Response to the Golczyk et al. commentary. Plant Cell 2014, 26, 855–861. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shaver, J.M.; Oldenburg, D.J.; Bendich, A.J. Changes in chloroplast DNA during development in tobacco, Medicago truncatula, pea, and maize. Planta 2006, 224, 72–82. [Google Scholar] [CrossRef]
- Oldenburg, D.J.; Bendich, A.J. Changes in the structure of DNA molecules and the amount of DNA per plastid during chloroplast development in maize. J. Mol. Biol. 2004, 344, 1311–1330. [Google Scholar] [CrossRef] [PubMed]
- Egelman, E.H. Mutate or die: Atomic structures explain bacterial SOS induction. Proc. Natl. Acad. Sci. USA 2023, 120, e2221605120. [Google Scholar] [CrossRef] [PubMed]
- Wright, A.F.; Murphy, M.P.; Turnbull, D.M. Do organellar genomes function as long-term redox damage sensors? Trends Genet. 2009, 25, 253–261. [Google Scholar] [CrossRef] [PubMed]
- Chevigny, N.; Schatz-Daas, D.; Lotfi, F.; Gualberto, J.M. DNA repair and the stability of the plant mitochondrial genome. Int. J. Mol. Sci. 2020, 21, 328. [Google Scholar] [CrossRef] [Green Version]


{kind=link}
{kind=link}
| ROS Assays 1 | Tissue | Condition |
| General ROS | Leaf > stalk 2 | Light > dark 3 |
| Hydrogen peroxide | Leaf > stalk | Light > dark |
| Superoxide 4 | Leaf > stalk | Light > dark |
| Superoxide dismutase | Leaf > stalk | Light > dark |
| Catalase | Leaf < stalk | Light < dark |
| Peroxidase | Leaf < stalk | Light < dark |
| Glutathione | Leaf < stalk | Light < dark |
| Ascorbic acid | Leaf < stalk | Light < dark |
| 8-oxoG in DNA | Leaf > stalk | Light > dark |
| Glycation assays 5 | ||
| MGO | Leaf > stalk | Light > dark |
| AGEs in proteins | Leaf > stalk | Light > dark |
| Amadori-adducts in DNA | Leaf > stalk | Light > dark |
| AGEs in DNA | Leaf > stalk | Light > dark |
| DJ-1 | Leaf < stalk | Light < dark |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tripathi, D.; Oldenburg, D.J.; Bendich, A.J. Oxidative and Glycation Damage to Mitochondrial DNA and Plastid DNA during Plant Development. Antioxidants 2023, 12, 891. https://doi.org/10.3390/antiox12040891
Tripathi D, Oldenburg DJ, Bendich AJ. Oxidative and Glycation Damage to Mitochondrial DNA and Plastid DNA during Plant Development. Antioxidants. 2023; 12(4):891. https://doi.org/10.3390/antiox12040891
Chicago/Turabian StyleTripathi, Diwaker, Delene J. Oldenburg, and Arnold J. Bendich. 2023. "Oxidative and Glycation Damage to Mitochondrial DNA and Plastid DNA during Plant Development" Antioxidants 12, no. 4: 891. https://doi.org/10.3390/antiox12040891
APA StyleTripathi, D., Oldenburg, D. J., & Bendich, A. J. (2023). Oxidative and Glycation Damage to Mitochondrial DNA and Plastid DNA during Plant Development. Antioxidants, 12(4), 891. https://doi.org/10.3390/antiox12040891