


The Eco-Immunological Relevance of the Anti-Oxidant Response in Invasive Molluscs

{kind=link}
{kind=link}
Abstract
:1. Introduction
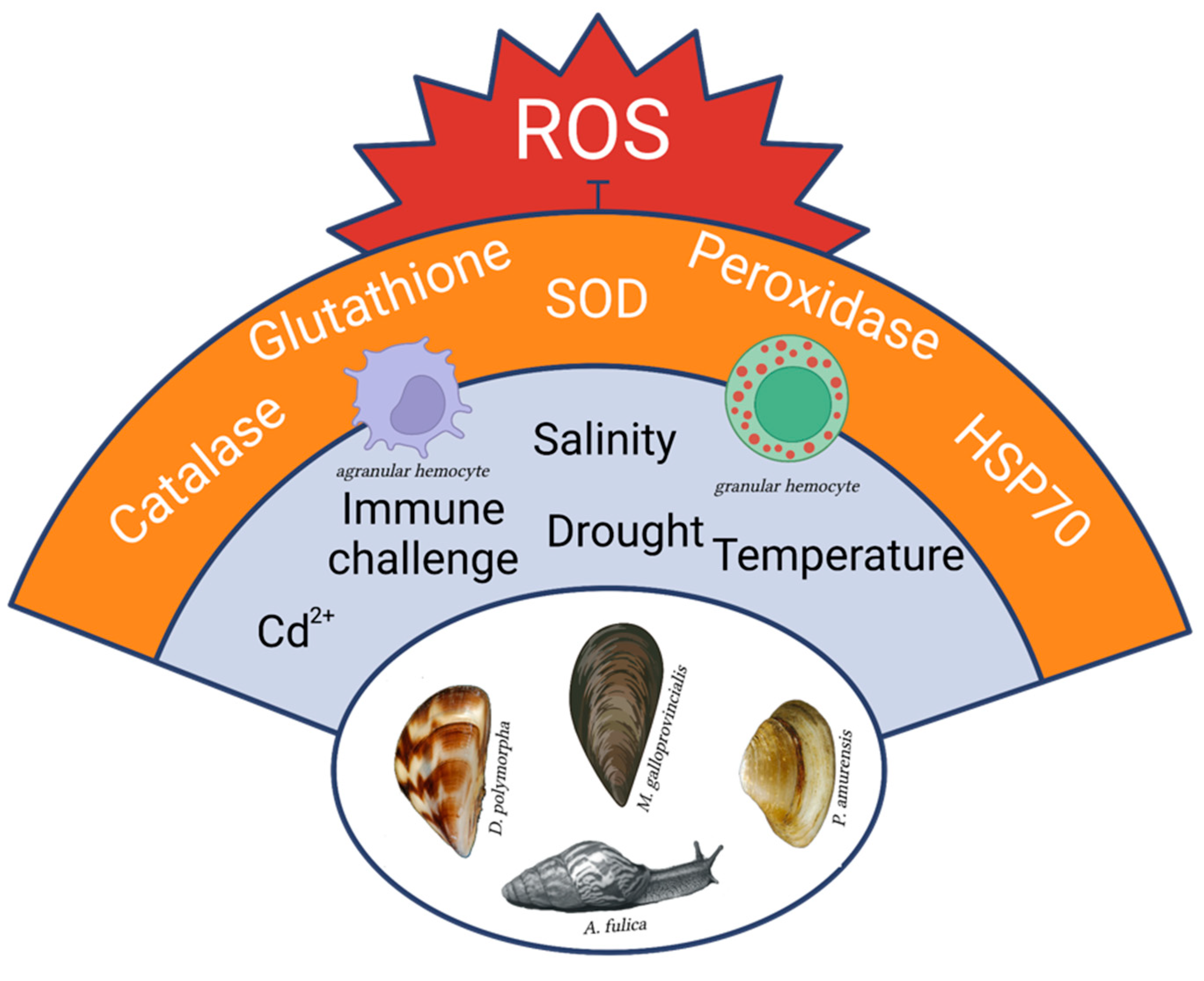
2. The Oxidative Stress Response against Immune or Environmental Challenges in Highly Invasive Mollusc Species
3. The Eco-Immunological Advantage of Managing Oxidative Stress Response
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Byers, J.E.; Blaze, J.A.; Dodd, A.C.; Hall, H.L.; Gribben, P.E. Exotic Asphyxiation: Interactions between Invasive Species and Hypoxia. Biol. Rev. 2023, 98, 150–167. [Google Scholar] [CrossRef] [PubMed]
- Stock, A.; Murray, C.C.; Gregr, E.J.; Steenbeek, J.; Woodburn, E.; Micheli, F.; Christensen, V.; Chan, K.M.A. Exploring Multiple Stressor Effects with Ecopath, Ecosim, and Ecospace: Research Designs, Modeling Techniques, and Future Directions. Sci. Total Environ. 2023, 869, 161719. [Google Scholar] [CrossRef] [PubMed]
- Capri, M.; Salvioli, S.; Monti, D.; Bucci, L.; Garagnani, P.; Ottaviani, E.; Franceschi, C. Eco-Immunology, Evolutive Aspects and Future Perspectives; Springer: Heidelberg, Germany, 2014; pp. 125–144. [Google Scholar] [CrossRef]
- Lowe, S.; Browne, M.; Boudjelas, S.; De Poorter, M. 100 of the World’s Worst Invasive Alien Species: A Selection from the Global Invasive Species Database; The Invasive Species Specialist Group (ISSG): Auckland, New Zealand; p. 12. First published as special lift-out in Aliens 12, December 2000. Updated and reprinted version: November 2004.
- Ng, T.H.; Kurtz, J. Dscam in Immunity: A Question of Diversity in Insects and Crustaceans. Dev. Comp. Immunol. 2020, 105, 103539. [Google Scholar] [CrossRef] [PubMed]
- Gerdol, M.; Moro, G.D.; Manfrin, C.; Venier, P.; Pallavicini, A. Big Defensins and Mytimacins, New AMP Families of the Mediterranean Mussel Mytilus galloprovincialis. Dev. Comp. Immunol. 2012, 36, 390–399. [Google Scholar] [CrossRef]
- Morris, G.; Gevezova, M.; Sarafian, V.; Maes, M. Redox Regulation of the Immune Response. Cell. Mol. Immunol. 2022, 19, 1079–1101. [Google Scholar] [CrossRef] [PubMed]
- Staerck, C.; Gastebois, A.; Vandeputte, P.; Calenda, A.; Larcher, G.; Gillmann, L.; Papon, N.; Bouchara, J.-P.; Fleury, M.J.J. Microbial Antioxidant Defense Enzymes. Microb. Pathog. 2017, 110, 56–65. [Google Scholar] [CrossRef]
- Sokolova, I. Bioenergetics in Environmental Adaptation and Stress Tolerance of Aquatic Ectotherms: Linking Physiology and Ecology in a Multi-Stressor Landscape. J. Exp. Biol. 2021, 224, jeb236802. [Google Scholar] [CrossRef]
- Betteridge, D.J. What Is Oxidative Stress? Metabolis 2000, 49, 3–8. [Google Scholar] [CrossRef]
- Pizzino, G.; Irrera, N.; Cucinotta, M.; Pallio, G.; Mannino, F.; Arcoraci, V.; Squadrito, F.; Altavilla, D.; Bitto, A. Oxidative Stress: Harms and Benefits for Human Health. Oxid. Med. Cell. Longev. 2017, 2017, 8416763. [Google Scholar] [CrossRef] [Green Version]
- Gostyukhina, O.L.; Yu, A.A.; Chelebieva, E.S.; Vodiasova, E.A.; Lantushenko, A.O.; Kladchenko, E.S. Adaptive Potential of the Mediterranean Mussel Mytilus galloprovincialis to Short-Term Environmental Hypoxia. Fish Shellfish Immun. 2022, 131, 654–661. [Google Scholar] [CrossRef]
- Sun, C.; Teng, J.; Wang, D.; Zhao, J.; Shan, E.; Wang, Q. The Adverse Impact of Microplastics and Their Attached Pathogen on Hemocyte Function and Antioxidative Response in the Mussel Mytilus galloprovincialis. Chemosphere 2023, 325, 138381. [Google Scholar] [CrossRef] [PubMed]
- Ramzan, R.; Vogt, S.; Kadenbach, B. Stress-Mediated Generation of Deleterious ROS in Healthy Individuals—Role of Cytochrome c Oxidase. J. Mol. Med. 2020, 98, 651–657. [Google Scholar] [CrossRef] [Green Version]
- Melis, J.P.M.; van Steeg, H.; Luijten, M. Oxidative DNA Damage and Nucleotide Excision Repair. Antioxid. Redox Signal. 2013, 18, 2409–2419. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tauffenberger, A.; Magistretti, P.J. Reactive Oxygen Species: Beyond Their Reactive Behavior. Neurochem. Res. 2021, 46, 77–87. [Google Scholar] [CrossRef] [PubMed]
- Eaton, L.; Pamenter, M.E. What to Do with Low O2: Redox Adaptations in Vertebrates Native to Hypoxic Environments. Comp. Biochem. Physiol. Part A Mol. Integr. Physiol. 2022, 271, 111259. [Google Scholar] [CrossRef]
- Dickson, K.B.; Zhou, J. Role of Reactive Oxygen Species and Iron in Host Defense against Infection. Front. Biosci. 2020, 25, 1600–1616. [Google Scholar] [CrossRef]
- Ferreira-Cravo, M.; Welker, A.F.; Hermes-Lima, M. Aestivation, Molecular and Physiological Aspects. Prog. Mol. Subcell Biol. 2009, 49, 47–61. [Google Scholar] [CrossRef]
- Nowakowska, A.; Świderska-Kołacz, G.; Rogalska, J.; Caputa, M. Antioxidants and Oxidative Stress in Helix pomatia Snails during Estivation. Comp. Biochem. Physiol. Part C Toxicol. Pharmacol. 2009, 150, 481–486. [Google Scholar] [CrossRef]
- Barbieri, R.F.; Lester, P.J.; Miller, A.S.; Ryan, K.G. A Neurotoxic Pesticide Changes the Outcome of Aggressive Interactions between Native and Invasive Ants. Proc. R. Soc. B Biol. Sci. 2013, 280, 20132157. [Google Scholar] [CrossRef] [Green Version]
- Xu, X.; Wang, X.; Yang, Y.; Ares, I.; Martínez, M.; Lopez-Torres, B.; Martínez-Larrañaga, M.-R.; Wang, X.; Anadón, A.; Martinez, M.-A. Neonicotinoids: Mechanisms of Systemic Toxicity Based on Oxidative Stress-Mitochondrial Damage. Arch. Toxicol. 2022, 96, 1493–1520. [Google Scholar] [CrossRef]
- McLaughlin, G.M.; Dearden, P.K. Invasive Insects: Management Methods Explored. J. Insect. Sci. 2019, 19, 17. [Google Scholar] [CrossRef] [PubMed]
- Kumar, J.; Ramlal, A.; Mallick, D.; Mishra, V. An Overview of Some Biopesticides and Their Importance in Plant Protection for Commercial Acceptance. Plants 2021, 10, 1185. [Google Scholar] [CrossRef] [PubMed]
- Malagoli, D.; Mandrioli, M.; Tascedda, F.; Ottaviani, E. Circulating Phagocytes: The Ancient and Conserved Interface between Immune and Neuroendocrine Function. Biol. Rev. 2017, 92, 369–377. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Malagoli, D.; Casarini, L.; Sacchi, S.; Ottaviani, E. Stress and Immune Response in the Mussel Mytilus galloprovincialis. Fish Shellfish Immun. 2007, 23, 171–177. [Google Scholar] [CrossRef]
- Malagoli, D.; Gobba, F.; Ottaviani, E. Effects of 50-Hz Magnetic Fields on the Signalling Pathways of FMLP-Induced Shape Changes in Invertebrate Immunocytes: The Activation of an Alternative “Stress Pathway” . Biochim. Biophys. Acta BBA-Gen. Subj. 2003, 1620, 185–190. [Google Scholar] [CrossRef]
- Malagoli, D.; Ottaviani, E. Yessotoxin Affects FMLP-induced Cell Shape Changes in Mytilus galloprovincialis Immunocytes. Cell. Biol. Int. 2004, 28, 57–61. [Google Scholar] [CrossRef]
- Franco-Martínez, L.; Martínez-Subiela, S.; Escribano, D.; Schlosser, S.; Nöbauer, K.; Razzazi-Fazeli, E.; Romero, D.; Cerón, J.J.; Tvarijonaviciute, A. Alterations in Haemolymph Proteome of Mytilus galloprovincialis Mussel after an Induced Injury. Fish Shellfish Immun. 2018, 75, 41–47. [Google Scholar] [CrossRef]
- Canesi, L.; Ciacci, C.; Fabbri, R.; Marcomini, A.; Pojana, G.; Gallo, G. Bivalve Molluscs as a Unique Target Group for Nanoparticle Toxicity. Mar. Environ. Res. 2012, 76, 16–21. [Google Scholar] [CrossRef]
- Georgoulis, I.; Feidantsis, K.; Giantsis, I.A.; Kakale, A.; Bock, C.; Pörtner, H.O.; Sokolova, I.M.; Michaelidis, B. Heat Hardening Enhances Mitochondrial Potential for Respiration and Oxidative Defence Capacity in the Mantle of Thermally Stressed Mytilus galloprovincialis. Sci. Rep. 2021, 11, 17098. [Google Scholar] [CrossRef]
- Smith, V.J.; Accorsi, A.; Malagoli, D. The Evolution of the Immune System; Academic Press: London, UK, 2016; pp. 1–28. [Google Scholar] [CrossRef]
- Evariste, L.; Auffret, M.; Audonnet, S.; Geffard, A.; David, E.; Brousseau, P.; Fournier, M.; Betoulle, S. Functional Features of Hemocyte Subpopulations of the Invasive Mollusk Species Dreissena polymorpha. Fish Shellfish Immun. 2016, 56, 144–154. [Google Scholar] [CrossRef]
- Guernic, A.L.; Geffard, A.; Rioult, D.; Bigot-Clivot, A.; Leprêtre, M.; Ladeiro, M.P. Cellular and Molecular Complementary Immune Stress Markers for the Model Species Dreissena polymorpha. Fish Shellfish Immun. 2020, 107, 452–462. [Google Scholar] [CrossRef] [PubMed]
- Binelli, A.; Torre, C.D.; Magni, S.; Parolini, M. Does Zebra Mussel (Dreissena polymorpha) Represent the Freshwater Counterpart of Mytilus in Ecotoxicological Studies? A Critical Review. Environ. Pollut. 2015, 196, 386–403. [Google Scholar] [CrossRef] [PubMed]
- Evariste, L.; Rioult, D.; Brousseau, P.; Geffard, A.; David, E.; Auffret, M.; Fournier, M.; Betoulle, S. Differential Sensitivity to Cadmium of Immunomarkers Measured in Hemocyte Subpopulations of Zebra Mussel Dreissena polymorpha. Ecotoxicol. Environ. Saf. 2017, 137, 78–85. [Google Scholar] [CrossRef] [PubMed]
- Miller, N.A.; Chen, X.; Stillman, J.H. Metabolic Physiology of the Invasive Clam, Potamocorbula amurensis: The Interactive Role of Temperature, Salinity, and Food Availability. PLoS ONE 2014, 9, e91064. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Werner, I. The Influence of Salinity on the Heat-Shock Protein Response of Potamocorbula amurensis (Bivalvia). Mar. Environ. Res. 2004, 58, 803–807. [Google Scholar] [CrossRef]
- Werner, I.; Hinton, D.E. Spatial Profiles of Hsp70 Proteins in Asian Clam (Potamocorbula amurensis) in Northern San Francisco Bay May Be Linked to Natural Rather than Anthropogenic Stressors. Mar. Environ. Res. 2000, 50, 379–384. [Google Scholar] [CrossRef]
- Lima, M.G.; Augusto, R.D.C.; Pinheiro, J.; Thiengo, S.C. Physiology and Immunity of the Invasive Giant African Snail, Achatina (Lissachatina) fulica, Intermediate Host of Angiostrongylus cantonensis. Dev. Comp. Immunol. 2020, 105, 103579. [Google Scholar] [CrossRef]
- Adema, C.M.; Harris, R.A.; van Deutekom-Mulder, E.C. A Comparative Study of Hemocytes from Six Different Snails: Morphology and Functional Aspects. J. Invertebr. Pathol. 1992, 59, 24–32. [Google Scholar] [CrossRef]
- Coaglio, A.L.; Ferreira, M.A.N.D.; dos Santos Lima, W.; de Jesus Pereira, C.A. Identification of a Phenoloxidase- and Melanin-Dependent Defence Mechanism in Achatina fulica Infected with Angiostrongylus vasorum. Parasites Vectors 2018, 11, 113. [Google Scholar] [CrossRef] [Green Version]
- Brinkmann, V.; Reichard, U.; Goosmann, C.; Fauler, B.; Uhlemann, Y.; Weiss, D.S.; Weinrauch, Y.; Zychlinsky, A. Neutrophil Extracellular Traps Kill Bacteria. Science 2004, 303, 1532–1535. [Google Scholar] [CrossRef]
- Lange, M.K.; Penagos-Tabares, F.; Muñoz-Caro, T.; Gärtner, U.; Mejer, H.; Schaper, R.; Hermosilla, C.; Taubert, A. Gastropod-Derived Haemocyte Extracellular Traps Entrap Metastrongyloid Larval Stages of Angiostrongylus vasorum, Aelurostrongylus abstrusus and Troglostrongylus brevior. Parasites Vectors 2017, 10, 50. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tunholi-Alves, V.M.; Tunholi, V.M.; Garcia, J.; Mota, E.M.; Castro, R.N.; Pontes, E.G.; Pinheiro, J. Unveiling the Oxidative Metabolism of Achatina fulica (Mollusca: Gastropoda) Experimentally Infected to Angiostrongylus cantonensis (Nematoda: Metastrongylidae). Parasitol. Res. 2018, 117, 1773–1781. [Google Scholar] [CrossRef]
- Williams, A.J.; Rae, R. Susceptibility of the Giant African Snail (Achatina fulica) Exposed to the Gastropod Parasitic Nematode Phasmarhabditis hermaphrodita. J. Invertebr. Pathol. 2015, 127, 122–126. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ottaviani, E.; Malagoli, D.; Capri, M.; Franceschi, C. Ecoimmunology: Is There Any Room for the Neuroendocrine System? Bioessays 2008, 30, 868–874. [Google Scholar] [CrossRef] [PubMed]
- Campbell, B.G.; Little, M.D. The Finding of Angiostrongylus cantonensis in Rats in New Orleans. Am. J. Trop. Med. Hyg. 1988, 38, 568–573. [Google Scholar] [CrossRef]
- Shozawa, A.; Suto, C. Hemocytes of Pomacea canaliculata: I. Reversible Aggregation Induced by Ca2+. Dev. Comp. Immunol. 1990, 14, 175–184. [Google Scholar] [CrossRef] [PubMed]
- Accorsi, A.; Bucci, L.; de Eguileor, M.; Ottaviani, E.; Malagoli, D. Comparative Analysis of Circulating Hemocytes of the Freshwater Snail Pomacea canaliculata. Fish Shellfish Immun. 2013, 34, 1260–1268. [Google Scholar] [CrossRef]
- Cueto, J.A.; Rodriguez, C.; Vega, I.A.; Castro-Vazquez, A. Immune Defenses of the Invasive Apple Snail Pomacea canaliculata (Caenogastropoda, Ampullariidae): Phagocytic Hemocytes in the Circulation and the Kidney. PLoS ONE 2015, 10, e0123964. [Google Scholar] [CrossRef] [Green Version]
- Boraldi, F.; Lofaro, F.D.; Accorsi, A.; Ross, E.; Malagoli, D.; Boraldi, F.; Lofaro, F.D. Toward the Molecular Deciphering of Pomacea canaliculata Immunity: First Proteomic Analysis of Circulating Hemocytes. Proteomics 2019, 19, 1800314. [Google Scholar] [CrossRef]
- Rodriguez, C.; Prieto, G.I.; Vega, I.A.; Castro-Vazquez, A. Assessment of the Kidney and Lung as Immune Barriers and Hematopoietic Sites in the Invasive Apple Snail Pomacea canaliculata. PeerJ 2018, 6, e5789. [Google Scholar] [CrossRef] [Green Version]
- Bergamini, G.; Ahmad, M.; Cocchi, M.; Malagoli, D. A New Protocol of Computer-Assisted Image Analysis Highlights the Presence of Hemocytes in the Regenerating Cephalic Tentacles of Adult Pomacea canaliculata. Int. J. Mol. Sci. 2021, 22, 5023. [Google Scholar] [CrossRef] [PubMed]
- Yang, C.; Tian, Y.; Lv, T.; Chang, X.; Zhang, M.; Gong, G.; Zhao, L.; Yang, S.; Chen, H. Histopathological Effects of Pedunsaponin A on Pomacea canaliculata. Pestic Biochem. Phys. 2018, 148, 151–158. [Google Scholar] [CrossRef] [PubMed]
- Yang, C.; Lv, T.; Wang, B.; Qiu, X.; Luo, L.; Zhang, M.; Yue, G.; Qin, G.; Xie, D.; Chen, H. The Damaging Effects of Pedunsaponin A on Pomacea canaliculata Hemocytes. Toxins 2019, 11, 390. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, C.; Ran, X.; Zhou, Y.; Huang, Y.; Yue, G.; Zhang, M.; Gong, G.; Chang, X.; Qiu, X.; Chen, H. Study on the Relationship of Hsp70 with the Temperature Sensitivity of Pedunsaponin a Poisoning Pomacea canaliculata. Pestic Biochem. Phys. 2022, 188, 105243. [Google Scholar] [CrossRef] [PubMed]
- Montanari, A.; Bergamini, G.; Ferrari, A.; Ferri, A.; Nasi, M.; Simonini, R.; Malagoli, D. The Immune Response of the Invasive Golden Apple Snail to a Nematode-Based Molluscicide Involves Different Organs. Biology 2020, 9, 371. [Google Scholar] [CrossRef]
- Accorsi, A.; Benatti, S.; Ross, E.; Nasi, M.; Malagoli, D. A Prokineticin-like Protein Responds to Immune Challenges in the Gastropod Pest Pomacea canaliculata. Dev. Comp. Immunol. 2017, 72, 37–43. [Google Scholar] [CrossRef] [PubMed]
- Giraud-Billoud, M.; Koch, E.; Vega, I.A.; Gamarra-Luques, C.; Castro-Vazquez, A. Urate Cells and Tissues in the South American Apple Snail Pomacea canaliculata. J. Mollus Stud. 2008, 74, 259–266. [Google Scholar] [CrossRef] [Green Version]
- Boraldi, F.; Lofaro, F.D.; Bergamini, G.; Ferrari, A.; Malagoli, D. Pomacea canaliculata Ampullar Proteome: A Nematode-Based Bio-Pesticide Induces Changes in Metabolic and Stress-Related Pathways. Biology 2021, 10, 1049. [Google Scholar] [CrossRef]
- Lennox, R.; Choi, K.; Harrison, P.M.; Paterson, J.E.; Peat, T.B.; Ward, T.D.; Cooke, S.J. Improving Science-Based Invasive Species Management with Physiological Knowledge, Concepts, and Tools. Biol. Invasions 2015, 17, 2213–2227. [Google Scholar] [CrossRef]
- Mooney, H.A.; Cleland, E.E. The Evolutionary Impact of Invasive Species. Proc. Natl. Acad. Sci. USA 2001, 98, 5446–5451. [Google Scholar] [CrossRef] [Green Version]
- Armitage, S.A.O.; Peuß, R.; Kurtz, J. Dscam and Pancrustacean Immune Memory—A Review of the Evidence. Dev. Comp. Immunol. 2015, 48, 315–323. [Google Scholar] [CrossRef] [PubMed]
- Yakovenko, I.; Donnyo, A.; Ioscovich, O.; Rosental, B.; Oren, M. The Diverse Transformer (Trf) Protein Family in the Sea Urchin Paracentrotus lividus Acts through a Collaboration between Cellular and Humoral Immune Effector Arms. Int. J. Mol. Sci. 2021, 22, 6639. [Google Scholar] [CrossRef] [PubMed]
- Schultz, J.H.; Bu, L.; Adema, C.M. Comparative Immunological Study of the Snail Physella acuta (Hygrophila, Pulmonata) Reveals Shared and Unique Aspects of Gastropod Immunobiology. Mol. Immunol. 2018, 101, 108–119. [Google Scholar] [CrossRef] [PubMed]
- Liu, D.; Yi, Q.; Wu, Y.; Lu, G.; Gong, C.; Song, X.; Sun, J.; Qu, C.; Liu, C.; Wang, L.; et al. A Hypervariable Immunoglobulin Superfamily Member from Crassostrea gigas Functions as Pattern Recognition Receptor with Opsonic Activity. Dev. Comp. Immunol. 2018, 86, 96–108. [Google Scholar] [CrossRef]
- Cantet, F.; Toubiana, M.; Parisi, M.-G.; Sonthi, M.; Cammarata, M.; Roch, P. Individual Variability of Mytimycin Gene Expression in Mussel. Fish Shellfish Immun. 2012, 33, 641–644. [Google Scholar] [CrossRef]
- Desriac, F.; Chevalier, P.; Brillet, B.; Leguerinel, I.; Thuillier, B.; Paillard, C.; Fleury, Y. Exploring the Hologenome Concept in Marine Bivalvia: Haemolymph Microbiota as a Pertinent Source of Probiotics for Aquaculture. FEMS Microbiol. Lett. 2014, 350, 107–116. [Google Scholar] [CrossRef] [Green Version]
- Liu, H.; Zha, S.; Yang, Z.; Zhang, W.; Lin, Z.; Wang, S.; Bao, Y. Acute Sulfide Exposure Induces Hemocyte Toxicity and Microbiota Dysbiosis in Blood Clam Tegillarca granosa. Aquat. Toxicol. 2022, 249, 106224. [Google Scholar] [CrossRef]
- Destoumieux-Garzón, D.; Canesi, L.; Oyanedel, D.; Travers, M.; Charrière, G.M.; Pruzzo, C.; Vezzulli, L. Vibrio–Bivalve Interactions in Health and Disease. Environ. Microbiol. 2020, 22, 4323–4341. [Google Scholar] [CrossRef]
- Vasquez, M.C.; Martinez, D.A.; Tomanek, L. Multiple Stressor Responses Are Regulated by Sirtuins in Mytilus Congeners. Comp. Biochem. Physiol. Part A Mol. Integr. Physiol. 2020, 246, 110719. [Google Scholar] [CrossRef]
- Collins, C.L.; Burnett, N.P.; Ramsey, M.J.; Wagner, K.; Zippay, M.L. Physiological Responses to Heat Stress in an Invasive Mussel Mytilus galloprovincialis Depend on Tidal Habitat. Mar. Environ. Res. 2020, 154, 104849. [Google Scholar] [CrossRef]
- Wojtal-Frankiewicz, A.; Bernasińska, J.; Frankiewicz, P.; Gwoździński, K.; Jurczak, T. The Role of Environmental Factors in the Induction of Oxidative Stress in Zebra Mussel (Dreissena polymorpha). Aquat. Ecol. 2017, 51, 289–306. [Google Scholar] [CrossRef] [Green Version]
- Ventura, L.D.; Sarpe, D.; Kopp, K.; Jokela, J. Variability in Phenotypic Tolerance to Low Oxygen in Invasive Populations of Quagga and Zebra Mussels. Aquat. Invasions 2016, 11, 267–276. [Google Scholar] [CrossRef]
- Carlton, J.; Tompson, J.; Schemel, L.; Nichols, F. Remarkable Invasion of San Francisco Bay (California, USA), by the Asian Clam Potamocorbula amurensis. I. Introduction and Dispersal. Mar. Ecol. Prog. Ser. 1990, 66, 81–94. [Google Scholar] [CrossRef]
- Paganini, A.; Kimmerer, W.; Stillman, J. Metabolic Responses to Environmental Salinity in the Invasive Clam Corbula amurensis. Aquat. Biol. 2010, 11, 139–147. [Google Scholar] [CrossRef] [Green Version]
- Salway, K.D.; Tattersall, G.J.; Stuart, J.A. Rapid Upregulation of Heart Antioxidant Enzymes during Arousal from Estivation in the Giant African Snail (Achatina fulica). Comp. Biochem. Physiol. Part A Mol. Integr. Physiol. 2010, 157, 229–236. [Google Scholar] [CrossRef]
- Giraud-Billoud, M.; Abud, M.A.; Cueto, J.A.; Vega, I.A.; Castro-Vazquez, A. Uric Acid Deposits and Estivation in the Invasive Apple-Snail, Pomacea canaliculata. Comp. Biochem. Physiol. Part A Mol. Integr. Physiol. 2011, 158, 506–512. [Google Scholar] [CrossRef] [PubMed]
- Hiong, K.C.; Loong, A.M.; Chew, S.F.; Ip, Y.K. Increases in Urea Synthesis and the Ornithine–Urea Cycle Capacity in the Giant African Snail, Achatina fulica, during Fasting or Aestivation, or after the Injection with Ammonium Chloride. J. Exp. Zool. Part A Comp. Exp. Biol. 2005, 303, 1040–1053. [Google Scholar] [CrossRef]
- Giraud-Billoud, M.; Campoy-Diaz, A.D.; Dellagnola, F.A.; Rodriguez, C.; Vega, I.A. Antioxidant Responses Induced by Short-Term Activity–Estivation–Arousal Cycle in Pomacea canaliculata. Front. Physiol. 2022, 13, 805168. [Google Scholar] [CrossRef]
- Giraud-Billoud, M.; Vega, I.A.; Tosi, M.E.R.; Abud, M.A.; Calderón, M.L.; Castro-Vazquez, A. Antioxidant and Molecular Chaperone Defences during Estivation and Arousal in the South American Apple Snail Pomacea canaliculata. J. Exp. Biol. 2012, 216, 614–622. [Google Scholar] [CrossRef] [Green Version]
- Rodriguez, C.; Campoy-Diaz, A.D.; Giraud-Billoud, M. Short-Term Estivation and Hibernation Induce Changes in the Blood and Circulating Hemocytes of the Apple Snail Pomacea canaliculata. Metabolites 2023, 13, 289. [Google Scholar] [CrossRef]
- Lochmiller, R.L.; Deerenberg, C. Trade-offs in Evolutionary Immunology: Just What Is the Cost of Immunity? Oikos 2000, 88, 87–98. [Google Scholar] [CrossRef] [Green Version]
- Malagoli, D.; Ottaviani, E. Life Is a Huge Compromise: Is the Complexity of the Vertebrate Immune-Neuroendocrine System an Advantage or the Price to Pay? Comp. Biochem. Physiol. Part A Mol. Integr. Physiol. 2010, 155, 134–138. [Google Scholar] [CrossRef] [PubMed]
- Gordon, S.; Martinez-Pomares, L. Physiological Roles of Macrophages. Pflügers Arch.-Eur. J. Physiol. 2017, 469, 365–374. [Google Scholar] [CrossRef] [Green Version]
- Molina-Cruz, A.; DeJong, R.J.; Charles, B.; Gupta, L.; Kumar, S.; Jaramillo-Gutierrez, G.; Barillas-Mury, C. Reactive Oxygen Species Modulate Anopheles gambiae Immunity against Bacteria and Plasmodium. J. Biol. Chem. 2008, 283, 3217–3223. [Google Scholar] [CrossRef] [Green Version]
- EFSA Panel on Plant Health (PLH). Scientific Opinion on the Assessment of the Potential Establishment of the Apple Snail in the EU. EFSA J. 2013, 11, 3487. [Google Scholar] [CrossRef]
- Sarma, R.R.; Munsi, M.; Ananthram, A.N. Effect of Climate Change on Invasion Risk of Giant African Snail (Achatina fulica Férussac, 1821: Achatinidae) in India. PLoS ONE 2015, 10, e0143724. [Google Scholar] [CrossRef] [Green Version]
- Thyrring, J.; Macleod, C.D.; Marshall, K.E.; Kennedy, J.; Tremblay, R.; Harley, C.D. Ocean Acidification Increases Susceptibility to Sub-Zero Air Temperatures in Ecosystem Engineers and Limits Poleward Range Shifts. Elife 2023, 12, e81080. [Google Scholar] [CrossRef]


Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Malagoli, D.; Franchi, N.; Sacchi, S. The Eco-Immunological Relevance of the Anti-Oxidant Response in Invasive Molluscs. Antioxidants 2023, 12, 1266. https://doi.org/10.3390/antiox12061266
Malagoli D, Franchi N, Sacchi S. The Eco-Immunological Relevance of the Anti-Oxidant Response in Invasive Molluscs. Antioxidants. 2023; 12(6):1266. https://doi.org/10.3390/antiox12061266
Chicago/Turabian StyleMalagoli, Davide, Nicola Franchi, and Sandro Sacchi. 2023. "The Eco-Immunological Relevance of the Anti-Oxidant Response in Invasive Molluscs" Antioxidants 12, no. 6: 1266. https://doi.org/10.3390/antiox12061266
APA StyleMalagoli, D., Franchi, N., & Sacchi, S. (2023). The Eco-Immunological Relevance of the Anti-Oxidant Response in Invasive Molluscs. Antioxidants, 12(6), 1266. https://doi.org/10.3390/antiox12061266